
A Big Catch for Germ Cell Tumour Research
article has not abstract
Published in the journal:
. PLoS Genet 9(4): e32767. doi:10.1371/journal.pgen.1003481
Category:
Perspective
doi:
https://doi.org/10.1371/journal.pgen.1003481
Summary
article has not abstract
Testicular germ cell tumour (TGCT) is the most common cancer in young men, and the incidence of TGCT is rising worldwide for unknown reasons [1], [2]. Treatments for TGCT are overall quite effective, but at the cost of significant toxicity [3], creating a powerful incentive for the development of more specific, molecularly guided therapies. TGCTs are generally thought to arise from a pluripotent fetal or embryonic germ cell [4]. Reflecting this pluripotency, these tumours can present in a wide range of histologic forms. Seminomas are TGCTs that retain features of pluripotent, primitive germ cells. In contrast, non-seminoma TGCTs exhibit differentiation into forms resembling somatic tissues (teratomas) or extraembryonic structures such as yolk sac (yolk sac tumour) or placenta (choriocarcinoma) (Figure 1). Family members of TGCT patients have a markedly increased risk of developing TGCT, strongly implicating an underlying genetic basis. Recent genome-wide association studies of TGCT have identified SNPs near ATF7IP, BAK1, DMRT1, KITLG, SPRY4, and TERT-CLPTM1L that increase TGCT risk [5]–[7]; however, the mechanisms associating most of these loci with tumourigenesis remain unclear.
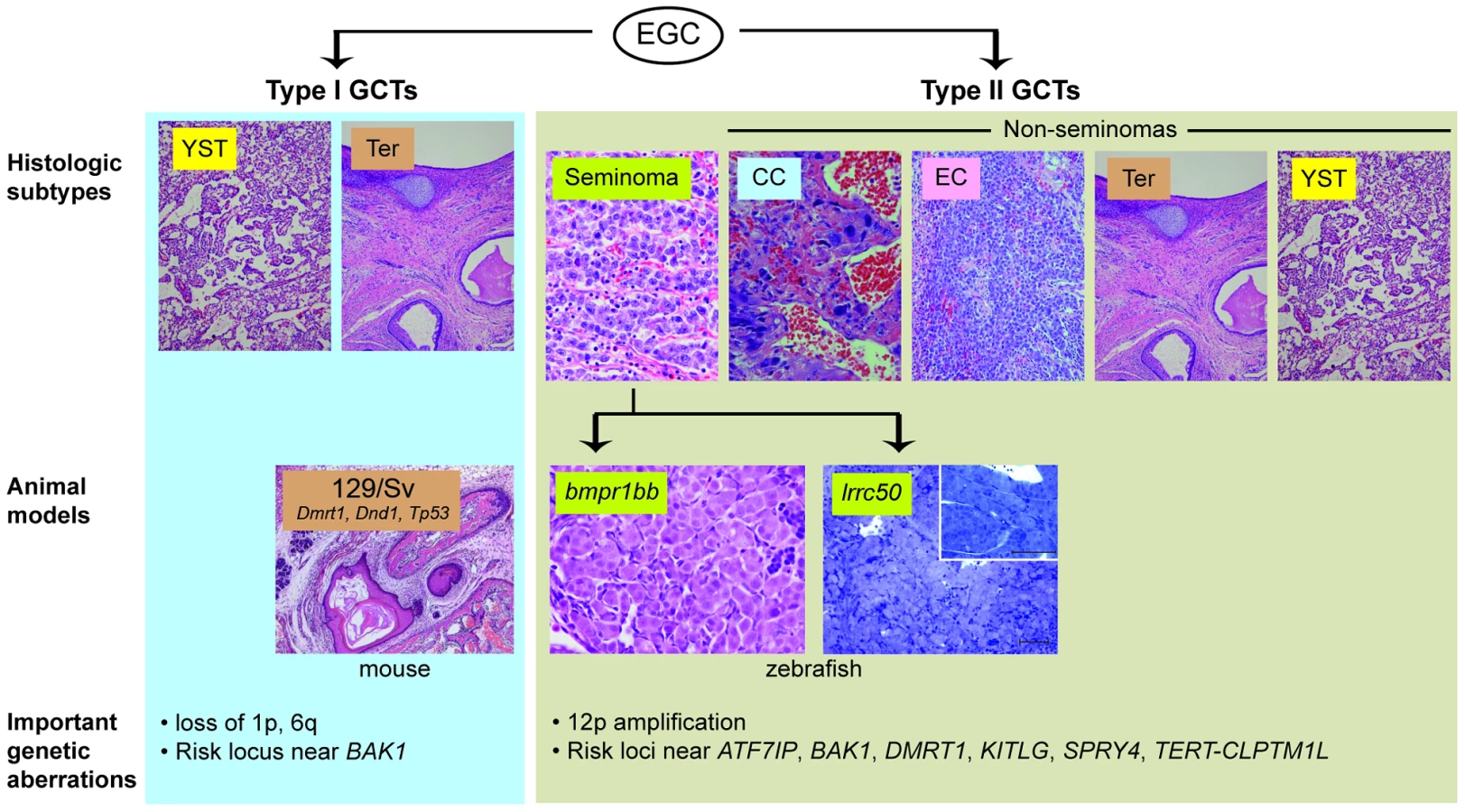
Researchers seeking to identify such mechanisms have been hindered by the limited TGCT animal models available thus far. Two heritable TGCT models have been previously described: one in mouse and one in zebrafish. The mouse model arose through the observation by Leroy Stevens in the late 1950s that testicular teratomas arise spontaneously at low frequency during embryonic development in mice of the 129/Sv strain [8]. This discovery, which ultimately led to the experimental derivation of embryonic stem cells [9], [10], has also proved to be a useful model of teratoma formation. A number of genes have been identified as modifier loci that increase teratoma incidence in the 129/Sv background, including Tp53, Dmrt1, and Dnd1, an RNA-binding protein that is central to germ cell maintenance [11]–[13]. Forward genetic screening led to the discovery of a second in vivo TGCT model. Zebrafish carrying nonsense mutations in alk6b/bmpr1bb, an ortholog of the human bone morphogenetic protein (BMP) receptor BMPR1B, develop TGCTs resembling human seminomas [14], [15]. This finding illuminated the importance of BMP signaling in germ cell development and implicated disruption of BMP signaling in human germ cell tumourigenesis [16]. These two models have provided insight into the roles of pluripotency and differentiation pathways in TGCT development; however, their direct correlation to human tumourigenesis has been limited, as genes such as DND1 and BMPR1B have not been found to be mutated in human TGCTs [14], [17].
In this issue of PLOS Genetics, Basten and coworkers describe a new zebrafish TGCT model with a direct connection to human TGCT mutations [18]. Zebrafish with homozygous mutations in the ciliary protein lrrc50 were previously described to have kidney cysts homologous to human polycystic kidney disease [19]. In this paper, the authors report that male lrrc50 heterozygotes develop testicular tumours late in life with near complete penetrance. Morphologically, these tumours, similar to those arising in bmpr1bb-deficient zebrafish, contain sheets of uniform, undifferentiated germ cells, resembling human seminoma. Loss of heterozygosity at the lrrc50 locus was found in some tumours, consistent with a role of lrrc50 as a tumour suppressor. The authors then conducted a mutational analysis of LRRC50 in a collection of human seminoma samples and identified different mutations in two pedigrees with family history of seminomas, as well as heterozygosity for a different germline LRRC50 mutation in five of 38 patients with sporadic seminomas. LRRC50 is thus the first gene specifically linked to seminoma predisposition in humans. The mutations were found to be functional nulls through their inability to complement lrrc50 knockdown in zebrafish embryos, an elegant example of the utility of the fish system for both gene discovery (forward genetics) and functional genomics (reverse genetics).
lrrc50 has heretofore been characterized solely as a ciliary motor protein, and its connection to GCT suppression is intriguing. Cilia have not previously been thought to be present in spermatogonia, but the authors show that normal spermatogonia do indeed have a cilium and that LRRC50 colocalized with the axoneme in spermatogonial stem cells. In addition, the authors provide evidence that its role may not be solely structural by showing that its expression is cell cycle–regulated and that it localizes with condensed chromosomes. The development of both renal cysts in homozygotes and seminomas in heterozygotes may implicate an underlying role for lrrc50 in early genitourinary development. Furthermore, the primary cilium has emerged as a signaling center for Hedgehog, Wnt, and other developmental pathways [20], and this, along with the TGCT phenotype of bmpr1bb mutants, raises the interesting possibility that interplay of these developmental signaling pathways is central to TGCT tumour suppression. Follow-up mechanistic studies will be critical to testing these hypotheses.
This paper provides another example of the power of zebrafish forward genetic screens for discovery of genes with novel roles in cancer and other diseases, and is a welcome addition to the list of animal models of TGCT. Some caveats apply when comparing analogous tumours arising in animals separated by a large evolutionary distance; for example, the fish seminomas are benign compared to human seminomas, and may well arise at a different stage of germ cell development. More significantly, neither the fish nor the mouse models currently reflect the biology of human non-seminomatous GCTs, such as embryonal carcinoma, choriocarcinoma, or yolk sac tumour. Clinically the non-seminomas are more likely to be metastatic and resistant to standard treatments, meaning that new models of these GCT subtypes are urgently needed to provide insight into tumour biology, as well as platforms for testing new therapeutic strategies for these cancers.
Zdroje
1. VisfeldtJ, JorgensenN, MullerJ, MollerH, SkakkebaekNE (1994) Testicular germ cell tumours of childhood in Denmark, 1943–1989: incidence and evaluation of histology using immunohistochemical techniques. J Pathol 174 : 39–47.
2. WuXC, ChenVW, SteeleB, RoffersS, KlotzJB, et al. (2003) Cancer incidence in adolescents and young adults in the United States, 1992–1997. J Adolesc Health 32 : 405–415.
3. Frazier AL, Amatruda JF (2009) Germ cell tumors. In: Fisher DE, Nathan D, Look AT, editors. Nathan and Oski's textbook of pediatric hematology-oncology. London: Elsevier.
4. OosterhuisJW, LooijengaLH (2005) Testicular germ-cell tumours in a broader perspective. Nat Rev Cancer 5 : 210–222.
5. RapleyEA, TurnbullC, Al OlamaAA, DermitzakisET, LingerR, et al. (2009) A genome-wide association study of testicular germ cell tumor. Nat Genet 41 : 807–810.
6. TurnbullC, RapleyEA, SealS, PernetD, RenwickA, et al. (2010) Variants near DMRT1, TERT and ATF7IP are associated with testicular germ cell cancer. Nat Genet 42 : 604–607.
7. PoynterJN, HootenAJ, FrazierAL, RossJA (2012) Associations between variants in KITLG, SPRY4, BAK1, and DMRT1 and pediatric germ cell tumors. Genes Chromosomes Cancer 51 : 266–271.
8. StevensLC, HummelKP (1957) A description of spontaneous congenital testicular teratomas in strain 129 mice. J Natl Cancer Inst 18 : 719–747.
9. MintzB, IllmenseeK (1975) Normal genetically mosaic mice produced from malignant teratocarcinoma cells. Proc Natl Acad Sci U S A 72 : 3585–3589.
10. StevensLC (1970) The development of transplantable teratocarcinomas from intratesticular grafts of pre - and postimplantation mouse embryos. Dev Biol 21 : 364–382.
11. KrentzAD, MurphyMW, KimS, CookMS, CapelB, et al. (2009) The DM domain protein DMRT1 is a dose-sensitive regulator of fetal germ cell proliferation and pluripotency. Proc Natl Acad Sci U S A 106 : 22323–22328.
12. LamMY, NadeauJH (2003) Genetic control of susceptibility to spontaneous testicular germ cell tumors in mice. APMIS 111 : 184–190; discussion 191.
13. YoungrenKK, CoveneyD, PengX, BhattacharyaC, SchmidtLS, et al. (2005) The Ter mutation in the dead end gene causes germ cell loss and testicular germ cell tumours. Nature 435 : 360–364.
14. NeumannJC, ChandlerGL, DamoulisVA, FustinoNJ, LillardK, et al. (2011) Mutation in the type IB bone morphogenetic protein receptor Alk6b impairs germ-cell differentiation and causes germ-cell tumors in zebrafish. Proc Natl Acad Sci U S A 108 : 13153–13158.
15. NeumannJC, DoveyJS, ChandlerGL, CarbajalL, AmatrudaJF (2009) Identification of a heritable model of testicular germ cell tumor in the zebrafish. Zebrafish 6 : 319–327.
16. FustinoN, RakhejaD, AteekCS, NeumannJC, AmatrudaJF (2011) Bone morphogenetic protein signalling activity distinguishes histological subsets of paediatric germ cell tumours. Int J Androl 34: e218–233.
17. LingerR, DudakiaD, HuddartR, TuckerK, FriedlanderM, et al. (2008) Analysis of the DND1 gene in men with sporadic and familial testicular germ cell tumors. Genes Chromosomes Cancer 47 : 247–252.
18. BastenSG, DavisEE, GillisAJM, van RooijenE, StoopH, et al. (2013) Mutations in LRRC50 predispose zebrafish and humans to seminomas. PLoS Genet 9: e1003384 doi:10.1371/journal. pgen.1003384.
19. van RooijenE, GilesRH, VoestEE, van RooijenC, Schulte-MerkerS, et al. (2008) LRRC50, a conserved ciliary protein implicated in polycystic kidney disease. J Am Soc Nephrol 19 : 1128–1138.
20. VelandIR, AwanA, PedersenLB, YoderBK, ChristensenST (2009) Primary cilia and signaling pathways in mammalian development, health and disease. Nephron Physiol 111: p39–53.
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2013 Číslo 4
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- The G4 Genome
- Neutral Genomic Microevolution of a Recently Emerged Pathogen, Serovar Agona
- The Histone Demethylase Jarid1b Ensures Faithful Mouse Development by Protecting Developmental Genes from Aberrant H3K4me3
- The Tissue-Specific RNA Binding Protein T-STAR Controls Regional Splicing Patterns of Pre-mRNAs in the Brain
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy