
Topoisomerase I Plays a Critical Role in Suppressing Genome Instability at a Highly Transcribed G-Quadruplex-Forming Sequence
Genome instability is not evenly distributed, but rather is highly elevated at certain genomic loci containing DNA sequences that can fold into non-canonical secondary structures. The four-stranded G-quadruplex or G4 DNA is one such DNA structure capable of instigating transcription and/or replication obstruction and subsequent genome instability. In this study, we used a reporter system to quantitatively measure the level of genome instability occurring at a G4 DNA motif integrated into the yeast genome. We showed that the disruption of Topoisomerase I function significantly elevated various types of genome instability at the highly transcribed G4 motif generating loss of heterozygosity and copy number alterations (deletions and duplications), both of which are frequently observed in cancer genomes.
Published in the journal:
. PLoS Genet 10(12): e32767. doi:10.1371/journal.pgen.1004839
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1004839
Summary
Genome instability is not evenly distributed, but rather is highly elevated at certain genomic loci containing DNA sequences that can fold into non-canonical secondary structures. The four-stranded G-quadruplex or G4 DNA is one such DNA structure capable of instigating transcription and/or replication obstruction and subsequent genome instability. In this study, we used a reporter system to quantitatively measure the level of genome instability occurring at a G4 DNA motif integrated into the yeast genome. We showed that the disruption of Topoisomerase I function significantly elevated various types of genome instability at the highly transcribed G4 motif generating loss of heterozygosity and copy number alterations (deletions and duplications), both of which are frequently observed in cancer genomes.
Introduction
In addition to the canonical Watson-Crick helical duplex (B-DNA), genomic DNA, especially repetitive sequences, can assume other types of structures such hairpins, Z-DNA, triplex DNA (H-DNA) or tetrahelical DNA structures [1]–[6]. Impediments to normal DNA metabolic processes including transcription and replication imposed by such secondary DNA structures explain the correlation between repetitive sequence elements and elevated genome instability. Genomic instability at purine-rich GAA•TTC repeats and CAG•CTG repeats, which can fold into three-stranded H-DNA [7] and a slipped hairpin structure [8], forms the molecular basis of multiple neurodegenerative diseases, such as Freidreich's Ataxia and Huntington's disease, respectively.
G-quadruplex or G4 DNA is another non-B secondary DNA structure that potentially interferes with normal DNA transactions [5], [9]–[11]. G4 DNA contains a stacked array of multiple G-quartets, which are comprised of four guanines interacting in a planar configuration [5], [10]. G4 DNA can be readily formed in solution by oligonucleotides containing multiple runs of guanines and by actively transcribed plasmid DNA [12], [13]. G4 DNA-forming sequences or G4 motifs are present in the genomes of diverse organisms and conserved throughout evolution; they number >375,000 in the human genome and >1,400 in the Saccharomyces cerevisiae nuclear genome [14]–[17]. The distribution of G4 motifs is highly concentrated at telomeres, rDNA loci, immunoglobulin heavy-chain switch regions, and G-rich minisatellites and significantly correlates with nucleosome-free regions and transcription start sites (TSSs) [10], [11], [18]. In oncogenes, G4 motifs are mostly enriched in the regions flanking TSSs, which suggests that G4 DNA may be involved in transcriptional regulation [15], [17].
G4 DNA becomes a structural barrier to transcription and replication in vitro indicating that it might play a significant role in genome instability [19]–[21]. In the absence of Pif1, a potent G4 DNA unwinding helicase, replication forks slow down near G4 motifs present in the yeast genome strengthening the argument that an unresolved G4 structure can lead to increased genome instability [22]. G4 motifs are frequently found at unstable genomic loci including proto-oncogenes and sites of frequent translocation breakpoints [23] and at preferred mitotic and meiotic DNA break sites [17]. In some human cancers, G4 motifs have been identified at frequent breakpoints involved in chromosomal translocations including the major breakpoint region in the proto-oncogene BCL2 [24]. Chromosomal translocations involving G-rich immunoglobulin switch regions have long been observed in various cancer cell lines [25]. The identification in silico of potential G4 DNA-forming sequences at sites of genome instability, however, has not yet been fully verified by in vivo demonstration of biological relevance of G4 DNA structure.
Seminal advances in understanding the genome instability induced by the repetitive, DNA secondary structure-forming sequences have been made using yeast and bacterial model systems [26]–[28]. A large tract of GAA•TTC repeats in yeast, for example, acted as a hotspot of gross chromosomal rearrangements (GCRs) and interstitial deletions [26], [27]. In bacteria, CTG•CAG repeats from the Myotonic Dystrophy gene induced large deletions when the repeats were highly transcribed [29]. When a guanine-run containing a human subtelomeric minisatellite was integrated into the yeast genome, it significantly elevated GCRs [30] and resulted in frequent repeat expansion and contraction [31]. The guanine-rich yeast telomere repeats, when placed within an intron of an interstitially located gene, cause various types of chromosomal rearrangements including deletions and inversions [32]. Accumulating evidence pointing to G4 motifs as genome instability hotspots underscores the importance of defining endogenous and exogenous factors that influence the integrity of genomic loci containing these motifs.
Active transcription, when oriented in the direction to place the guanine-runs in the transiently single stranded non-transcribed strand (NTS), was shown to stimulate formation of G4 DNA structure both in vitro and in bacterial cells [13]. To determine the effect of G4 DNA on yeast genome stability, we previously constructed a genomic reporter assay where a potential G4 DNA-forming sequence was highly transcribed to promote secondary DNA structure formation. By normalizing the extent of genome instability occurring at this reporter construct to that occurring at the exact same sequence transcribed in inverse orientation (that is, with the G4 motifs on the transcribed strand), we were able to apply a stringent control for the correlation between elevated genome instability and G4 DNA. Using this approach, we found that gene conversion recombination was significantly elevated by highly transcribing guanine-run containing sequence in a strictly strand-specific manner [33].
In the current report, we demonstrate that active transcription transforms a guanine-run containing sequence into a strong hotspot for gross chromosomal rearrangements and loss-of-heterozygosity (LOH). Our data also show that the direction of replication can significantly alter the level of instability at a potential G4 DNA-forming sequence suggesting that genomic context in terms of both transcription and replication is important when considering G4 motifs as potential genome instability hotspots. Finally, we identify a critical role of Topoisomerase I (Top1) in suppressing various types of genome rearrangements associated with co-transcriptionally formed G4 DNA.
Results
A genomic assay to measure gross chromosomal rearrangements
We previously showed that gene conversion resulting from ectopic recombination is increased due to co-transcriptionally formed G4 DNA. In order to determine whether co-transcriptionally formed G4 DNA can also elevate gross chromosomal rearrangements (GCRs), we modified the GCR reporter system previously described by Chen and Kolodner [34], [35]. In this reporter system, the URA3 gene was integrated into the left arm of chromosome V (CHR5) replacing the HXT13 gene located ∼8.5 kb centromere-distal to the CAN1 gene (Fig. 1A). The loss of functional CAN1 or URA3 results in resistance to the drug canavanine (Can) or 5-Fluoroorotic acid (5-FOA), respectively. Because the first essential gene on the left arm of CHR5, PCM1, is located ∼60 kb from the telomere, the region containing CAN1 and URA3 genes can be lost without affecting viability of haploids. The hypothetical rate of double drug resistance (CanR/5-FOAR) occurring via independent mutations in CAN1 and URA3 is approximately 10−12, which is significantly lower than the observed rate of deletion of the left arm of CHR5 (10−11–10−10 in a wild-type background) [35]. Therefore, by selecting for colonies resistant to both Can and 5-FOA, GCR events resulting in simultaneous loss of CAN1 and URA3 genes are detected.
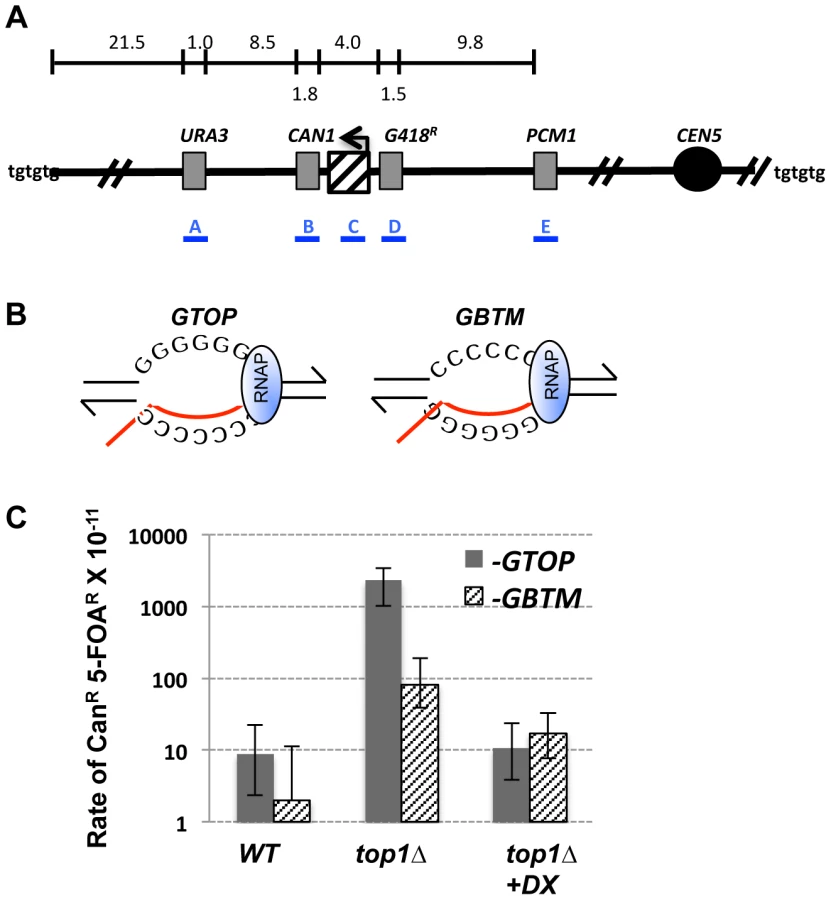
We modified the GCR assay by integrating the pTET-lys2-GTOP or -GBTM cassette immediately centromere-proximal to CAN1 (Fig. 1A). As previously described [36], the pTET-LYS2 cassette contains the LYS2 gene transcribed from the heterologous tetracycline-repressible promoter (pTET) linked to a marker gene conferring G418 resistance (G418R). Within the LYS2 ORF, at ∼390 bp from the start codon, a 760 bp fragment from the mouse immunoglobulin (Ig) switch Mu (Sμ) region was inserted to generate pTET-lys2-GTOP or -GBTM cassettes [33]. Switch regions, which are required for Ig heavy chain class-switch recombination (CSR), are comprised of multiple, degenerate guanine-rich repeats of several kb in length [37]. The Sμ sequence is a model G4 motif and was previously demonstrated to form G4 DNA structures both in vivo and in vitro when highly transcribed [13]. The sequence of the Sμ fragment incorporated into the LYS2 ORF (containing ∼17 of (GAGCT)nGGGGT repeats) is shown in Figure S1. This fragment was inserted either in the physiological (-GTOP) or in the inverted orientation (-GBTM), placing the guanine runs on the non-transcribed (NTS) or the transcribed strand (TS), respectively (Fig. 1B) [33].
GCR is elevated when Top1 is disrupted under high-transcription conditions
Topoisomerase 1 (Top1) is a highly conserved enzyme that relieves positive or negative torsional stress associated with transcription and replication [38], [39]. Top1 functions by covalently attaching to the 3′ end of nicked DNA, which is quickly re-ligated after swiveling of the DNA strands to remove supercoiling. Although not essential for viability in yeast, replication slows down and sometimes stalls in the absence of Top1, especially at highly transcribed regions [40]. We previously reported that disruption of Top1 leads to an increase in Sμ-induced gene conversion events [33].
In order to determine whether Top1 also plays a role in preventing GCRs initiating at G4 DNA, we deleted the TOP1 gene from strains containing pTET-lys2-GTOP and –GBTM constructs. In wild-type (WT) backgrounds under high-transcription conditions, the rates of CanR/5-FOAR events were 0.88×10−10 and 0.20×10−10 for pTET-lys2-GTOP and –GBTM constructs, respectively (Fig. 1C). These rates are not significantly different from each other and are comparable to GCR rates previously reported in the absence of an inserted pTET-lys2 cassette [34], [35]. Upon TOP1 deletion, the rate of GCR (CanR/5-FOAR) was significantly elevated for both the pTET-lys2-GTOP and -GBTM construct. Importantly, GCR occurred at a significantly higher (∼30-fold) rate for pTET-lys2–GTOP, where guanine-runs are present on the NTS, compared to the pTET-lys2–GBTM construct where guanine-runs are on the TS.
We tested whether active transcription of the guanine-run-containing sequence is required for the elevated GCRs by growing the top1Δ strains in medium containing doxycycline, an analog of tetracycline, which resulted in ∼60 - to 200-fold reductions in the transcription rates (Table S1). Repression of transcription from the pTET promoter by doxycycline resulted in a >200-fold decrease in the rate of CanR/5-FOAR for the strain associated with the pTET-lys2-GTOP construct (Fig. 1C). For the strain containing the pTET-lys2-GBTM construct, transcriptional repression also led to a significant decrease in the rate of CanR/5-FOAR.
In the absence of Top1, GCR events mainly occur proximal to G4 motifs and are frequently resolved by de novo telomere addition
In order to determine whether the GCRs in top1Δ backgrounds initiate at the G4 DNA containing reporter construct, we carried out PCR analysis to map the GCR initiating breakpoints. For the strain containing the pTET-lys2-GTOP construct in the top1Δ background, 27 of 30 CanR/5-FOAR isolates tested had lost the portion of LYS2 gene with the Sμ fragment insertion but still retained the G418R cassette located just centromere-proximal to this region (Fig. 1A and Table 1). Thus, 90% of the GCR events occurring in this strain initiated within the ∼4 kb region comprised of the pTET-lys2-GTOP cassette between G418R cassette and CAN1. This is proportionally greater than the GCR initiating in the same region for the pTET-lys2-GBTM construct in top1Δ strain (9 out 23; P<0.0005 by chi square analysis).

To further characterize the chromosome breakpoints in GCR events associated with the pTET-lys2-GTOP cassette, we carried out PCR with a degenerate primer annealing to generic yeast telomere sequence (CA16; 5′ CACCACACCCACACAC 3′) and a primer annealing to the 5′ untranslated region of the LYS2 gene. Out of 27 samples where the disruption of the pTET-lys2-GTOP cassette was confirmed by PCR mapping, telomere-anchored PCR products of 700 to 1900 bp were obtained for 16 samples. Subsequent sequencing of these fragments showed that, in 15 CanR/5-FOAR clones, de novo telomere additions occurred at various locations within the G4-forming Sμ fragment (Class I events in Fig. 2 and Fig. S1). Due to the high G/C content and repetitiveness of the Sμ sequence, sequencing analysis failed to identify the site of telomere addition in one of the PCR fragments. In concurrence with preferential telomere addition sites previously identified [41], the junctions of de novo telomere addition were located at GT dinucleotides and frequently at 5 to 6 nt clusters of GT-rich sequence. Additionally, mutations, deletions/insertions, and duplications within the Sμ fragment were detected in seven of the 15 CanR/5-FOAR clones with telomere additions.

The actively transcribed G4 motif induces complex genome rearrangements involving deletions and inverted duplications
We further characterized the genome rearrangements associated with G4 DNA using pulse field gel electrophoresis (PFGE) and microarray-based comparative genome hybridization (array-CGH). PFGE showed that, in the 16 samples where de novo telomere addition at Sμ was confirmed by sequencing (Class I), CHR5 was reduced in size by ∼35 kb and co-migrated with CHR8 (Fig. 2A and Fig. S2A). The loss of CHR5 sequences from the left telomere to the integration site of pTET-lys2 cassette (at 34,000 NT) was confirmed by array-CGH analysis (Fig. 2B and Fig. S2A).
In another 8 samples (Class II) where CHR5 appeared smaller by only ∼15 kb (Fig. 2A and Fig. S2B), array-CGH identified segmental deletions between the original location of HXT13 (at 23,000 NT) and the pTET-lys2 cassette. PCR and sequencing analysis showed that this recurrent deletion was mediated by a pair of 21 bp direct repeats introduced into the two respective sites of CHR5 as parts of plasmid constructs used for integration of the URA3Kl marker at HXT13 locus (pUG72; Euroscarf) and pTET promoter (pCM225; Euroscarf) (Fig. S3).
Finally, in class III events, deletion of CHR5 sequences from the left telomere to the pTET-lys2 integration site occurred in combination with duplications of the immediate proximal sequences on the left arm, and duplication of a terminal segment on the right arm (Fig. 2B and Fig. S2C–D). The two clones from class IIIa showed duplications from pTET-lys2 to the Watson-oriented YELWdelta1 dispersed long terminal repeat element (LTR), and the one example of class IIIb showed a duplication extending further to the full length Crick-oriented Ty1 element insertion at the URA3 locus (ura3-52 allele). All three class III clones had duplications of a segment of the right arm extending from position ∼446,000 NT (containing the full length Crick-oriented YERCTy1-1 element and the Watson-oriented YERWdelta20b LTR) all the way to the right telomere. These clones displayed longer versions of CHR5 of ∼700 kb (migrating just below CHR10) and ∼770 kb (co-migrating with CHR2 and CHR14) for class IIIa and class IIIb, respectively (Fig. 2A and Fig. S2C–D).
The class III chromosome sizes were consistent with the deletions and duplications detected by array-CGH and suggested a complex mechanism of formation. Similar array-CGH patterns were observed recently in the analyses of GCR events in yeast CHR5 as well as in humans [42], [43]. These studies described breakpoint structures consistent with an intra-strand fold-back mechanism in which a resected free 3′ end folds back on itself, re-anneals to a microhomology region and primes break-induced DNA replication (BIR). By plasmid-rescuing this regions subcloning and sequence analysis (See Materials and Methods), we confirmed that the duplicated regions proximal to the Sμ breakpoints in class IIIb clone A7 and class IIIa clone A8 were comprised of inverted duplications separated by single copy regions corresponding to the original ssDNA loops, as predicted by the fold-back mechanism (Figs. S4A and S4B). We were not successful in rescuing the duplicated regions from the Class IIIa clone A16. The recovered rearrangement structures observed in the A7 and A8 clones can be explained by two different models (Fig. S4C). In the first scenario, as the BIR event initiated by the intra-strand fold-back reached the Ty/LTR sequences on the left arm, it collapsed, re-annealed at the Ty/LTR sequences on the right arm (template switching), and continued on to reach the right telomere to produce a stable monocentric chromosome. The second possibility is that BIR continued all the way to the right telomere forming an unstable dicentric chromosome, which was then stabilized by a secondary homologous recombination event between Ty/LTR repeats leading to loss of one of the centromeres. Although our data does not allow us to distinguish between these two possibilities, we favor the template switching model (Model 1 in Fig. S4C) since BIR is generally thought to be impeded by centromeric structures, and BIR template switching has been shown to be frequent in yeast [44], [45].
A diploid system to examine the effect of G4 motifs on mitotic recombination
Spontaneous DNA breaks in diploid cells are frequently repaired by allelic mitotic recombination using as template either a sister chromatid or a homologous chromosome. We designed an assay that can measure G4 DNA-induced mitotic recombination between chromosome III (CHR3) homologs in diploids (Fig. 3A). First, we integrated the URA3 gene near the telomere of the left arm of CHR3 in a haploid strain derived from YPH45 (a S288c derivative). On the same arm of CHR3, about 44 kb centromere-proximal to the URA3 integration site, pTET-lys2-GTOP or -GBTM was integrated replacing HIS4. As described above for the CHR5 GCR assay, the pTET-lys2-GTOP and -GBTM cassettes contained the 760-bp fragment of Sμ sequence and were adjacent to an aminoglycoside phosphotransferase gene conferring resistance to the drug G418 (G418R). Because the direction of replication fork movement relative to the direction of transcription can affect recombination at highly transcribed regions [46], [47], each cassette was integrated in two orientations relative to the nearby replication origin ARS306. This yielded constructs in which the transcription and replication forks are co-directional (SAME) or in head-on orientation (OPPO) (Fig. 3B). The direction of replication fork movement through this region of CHR3 was previously confirmed by 2D-gel analysis [36].

Heterozygous diploids were generated by mating the YPH45-derived haploid strains described above to a haploid strain derived from YJM789, a clinically isolated strain with ∼0.5% sequence divergence relative to the S288c reference strain [48]. Because the YJM789-derived strain is Ura−, loss or mutation of the URA3 gene on the CHR3 from the YPH45 parent will result in resistance to 5-FOA in the heterozygous diploid cells. The types of genome rearrangements that can lead to loss of URA3 include (a) complete loss of YPH45 CHR3, (b) partial loss of the left arm of YPH45 CHR3, (c) Break Induced Replication (BIR) or reciprocal crossover (RCO) initiating between URA3 and CEN3 or (d) translocation/BIR events involving a heterologous chromosome. In RCO, the distal ends of the two CHR3s are exchanged without loss of genetic material. In BIR involving the homolog, the break in YPH45-CHR3 will be repaired through replication using YJM789-CHR3 from the break to telomere as template. In our assay, we cannot distinguish between these two mechanisms (Fig. S5). Because all of these events result in loss of heterozygosity (LOH) for the segment of CHR3 containing the URA3 marker, we hereafter refer to this assay as the LOH assay.
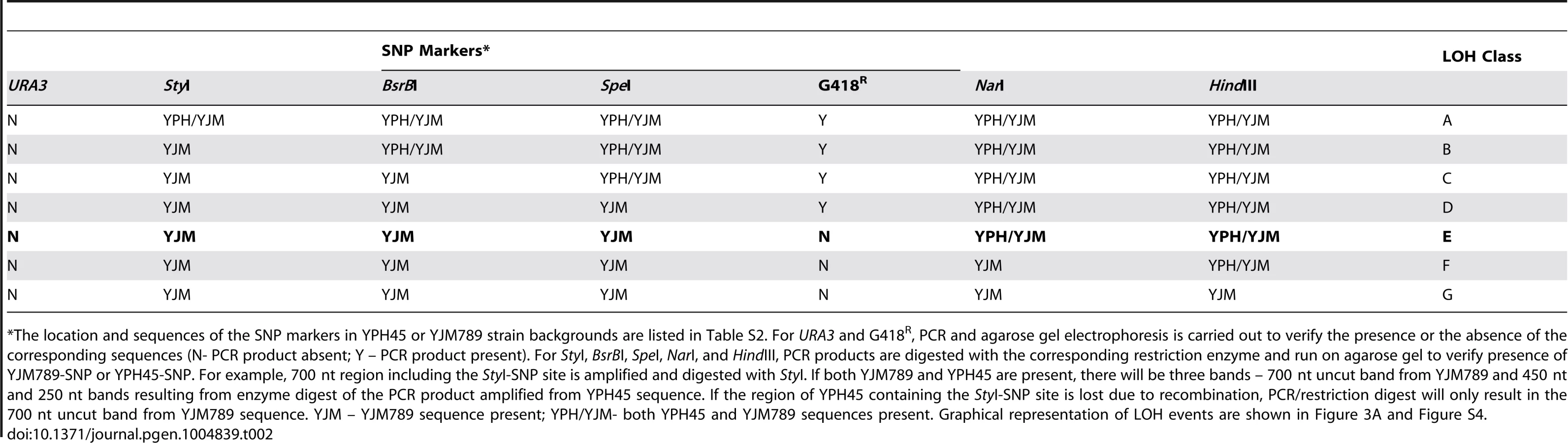
In order to map the position of LOH in 5-FOAR isolates, we devised a PCR-based restriction fragment length polymorphism (RFLP-SNP) assay described in Fig. S6. We defined the initiation of recombination point as between the last telomeric SNP site displaying LOH and the first centromeric SNP site displaying heterozygosity (Table 2). A 5-FOAR isolate was defined as resulting from recombination initiating at or near the G4 repeats (G418-pTET-lys2-GTOP or GBTM), when it was homozygous for the centromere-distal SpeI site and heterozygous for the centromere-proximal NarI site and the G418S cassette was not present (LOH class E in Table 2 and Figure 3A). When mitotic recombination is initiated by DNA breaks near the pTET-lys2-GTOP or –GBTM cassette, resection must extend into the region of the YPH45 CHR3 with homology to YJM789 CHR3 and, therefore, remove the pTET-lys2-GTOP or –GBTM cassette along with the G418R marker.
G4 insertion does not increase genome instability in WT backgrounds
To determine whether highly transcribed G4 motifs elevate LOH on CHR3, we measured the rate of 5-FOAR in WT strains containing pTET-LYS2 (no Sμ sequence) or pTET-lys2-GTOP cassette. In either the SAME or OPPO orientations, the overall rate of LOH (5-FOAR) associated with pTET-LYS2 or pTET-lys2-GTOP was not significantly different (Fig. 4A). Using the RFLP-SNP assay, we identified 3/46 or 9/46 LOH events initiated at the highly transcribed pTET-LYS2 cassette when in the SAME or the OPPO orientation, respectively (Table 3). These proportions were not statistically different from those for LOH initiating at pTET-lys2-GTOP in the SAME or OPPO orientation (by Fisher's exact test; P = 0.31 and 0.39, respectively).


Top1 disruption elevates mitotic recombination events initiating at a G4 motif-containing genomic locus
We observed a dramatic and specific increase in the rates of gene conversion [33] and GCR (see above) associated with highly transcribed Sμ sequences when Top1 was disrupted in a haploid YPH45 background. Importantly, both occurred at significantly higher rates when the guanine-run containing strand was on the NTS where its single stranded nature fosters G4 DNA formation. In order to determine whether LOH on CHR3 is similarly affected by the location of G4-forming sequence on the TS vs. NTS, we compared LOH rates associated with pTET-lys2-GTOP and -BTM constructs in top1Δ/top1Δ backgrounds. There was no significant difference between overall rates of LOH events associated with pTET-lys2-GTOP and -GBTM constructs when replication was in the SAME direction (Fig. 5A). However, when replication was in the OPPO orientation, the overall rate of LOH events was ∼3 fold higher for the pTET-lys2-GTOP than for the pTET-lys2-GBTM construct. The rates of LOH initiating at the pTET-lys2-GTOP/GBTM cassette in the SAME or OPPO orientation in top1Δ/top1Δ background were determined by analyzing 47–93 5-FOAR isolates by the RFLP-SNP assay (Table 4). When transcription from the pTET promoter was in the SAME orientation relative to replication originating at ARS306, the rate of LOH initiating at the G4-containing sequence was similar whether the guanine-runs were on the NTS (pTET-lys2-GTOP) or on the TS (pTET-lys2-GBTM) (Fig. 5B). However, the rate of LOH initiated near pTET-lys2-GTOP in the OPPO orientation was >20 fold higher than at pTET-lys2-GBTM in OPPO orientation and ∼4 fold higher than at pTET-lys2-GTOP in the SAME orientation.

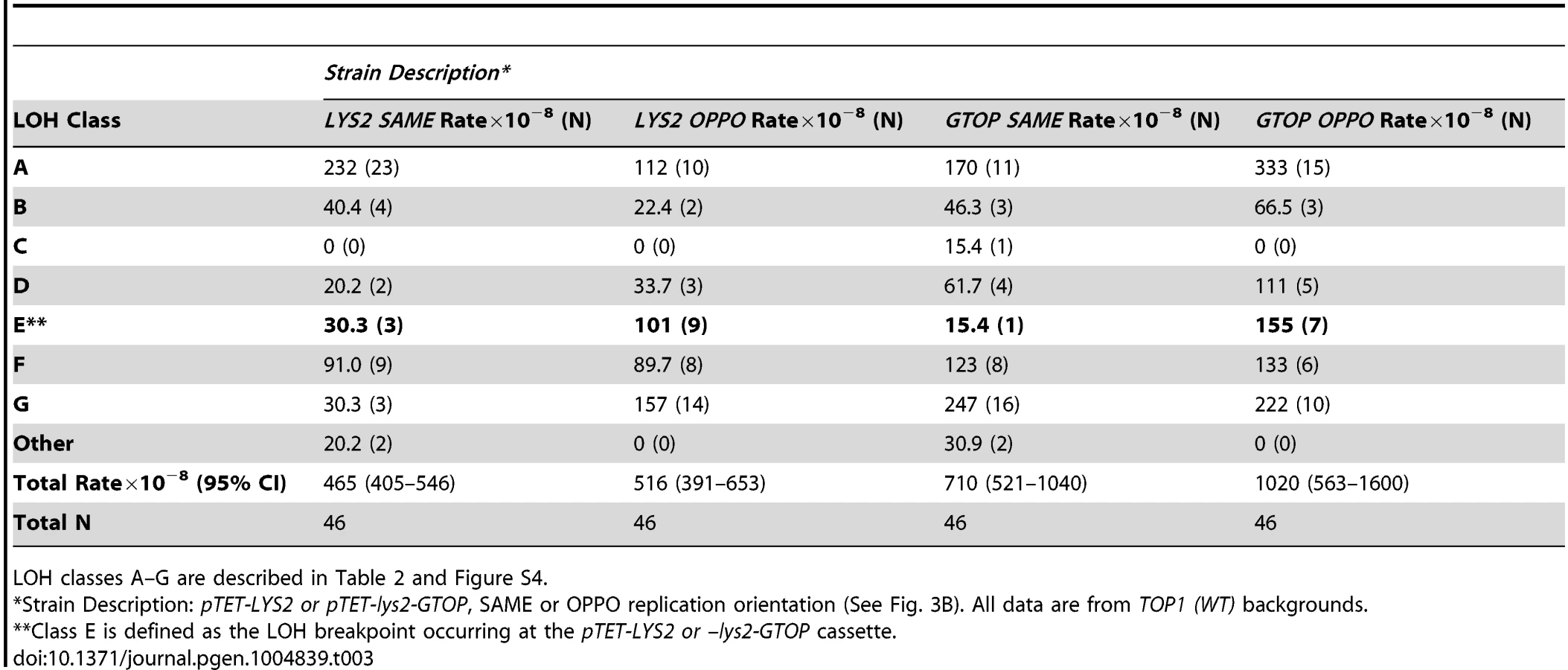
Upon Top1-disruption, when the replication and transcription is in “head-on” or OPPO orientation, the rate of loss of heterozygous SNP at NarI site (at 78380 NT) is also significantly elevated (Table 4). The NarI-SNP is not lost at a high rate in pTET-lys2-GBTM-containing strain, which suggests that recombination initiating at the pTET-lys2-GTOP cassette (at 68300 NT) are often associated with long conversion tracks resulting in the loss of NarI-SNP. This is consistent with the average length of conversion tracks associated with reciprocal crossover events, which is reported to be about 12 kb [49].
LOH events at G4 DNA mainly occur by crossover or break-induced replication in the absence of Top1
Using the RFLP-SNP assay described above, we characterized 93 5-FOAR isolates from the top1Δ/top1Δ strain containing the pTET-lys2-GTOP cassette in the OPPO orientation and identified 33 isolates with LOH events initiating near the G4 motif-containing reporter construct (Table 4 – LOH Class E). Such LOH events can occur by (1) mitotic recombination with the other CHR3 in the heterozygous diploid cell via reciprocal crossover (RCO) or break-induced replication (BIR) with the homolog, (2) the partial loss of the chromosome arm, or (3) translocation to another chromosome with a short stretch of homology (Figure 3A). In order to determine the types of rearrangements occurring at this locus, separation of the CHR3 homologs by PFGE was carried out for the 33 LOH Class E isolates (Figure S7). The CHR3 homologs of YPH45 and YJM789 differ in size by about 56 kb, likely reflecting different polymorphic subtelomeric gene content and retrotransposon insertions. In 32 out of 33 5-FOAR isolates analyzed, the YJM789-derived CHR3 was unchanged in size and the YPH45-derived CHR3 appeared slightly smaller than that of the parental haploid. A smaller CHR3 can be generated by RCO or BIR initiating on YPH45-derived CHR3 using YJM-789 derived CHR3 as the repair donor. The reduction in chromosome size was approximately 13 kb in 31/33 5-FOAR isolates (Fig. S7). In one of 33 isolates analyzed, YPH45-CHR3 was reduced in size by ∼24 kb. An ∼8.5 kb reduction was expected from the loss of hemizygous URA3 maker and the pTET-lys2-GTOP cassette with additional reduction in size resulting from the loss of other hemizygous sequences. Translocation to a heterologous chromosome was observed in 1/33 5-FOAR isolates. None of the analyzed isolates contained CHR3 shortened by ∼80 kb, which is predicted in case of the loss of left arm from the pTET-lys2-GTOP cassette to the telomere followed by a telomere addition at the break site (Figure 3A, bottom panel).
Replication-orientation significantly affects gene conversion rates associated with G4-forming sequence
In our previous report regarding the rate of gene conversion events induced by the highly transcribed Sμ sequence, the pTET-lys2-GTOP cassette was integrated in the orientation and location identical to the “OPPO” construct described above for the LOH assay (Fig. 6A). In this orientation, transcription from pTET promoter and replication originating at ARS306 are in the convergent or “head-on” orientation. In order to determine whether the rate of gene conversion is dependent on the relative orientation of transcription and replication, we deleted ARS306 by replacing the ARS consensus sequence (5′-WTTTAYRTTTW-3′) [50] with the gene encoding hygromycin B phosphotransferase (Hph). In the resulting ars306Δ strain, replication through the pTET-lys2-GTOP cassette originates from the ARS305 located about 27 kb away and is in co-directional orientation relative to transcription [51]. As previously reported, in the gene conversion assay, recombination initiating at the pTET-lys2 cassette can be completed using a truncated lys2 gene fragment integrated on CHR15 resulting in lysine prototrophy (Lys+) (Fig. 6A) [33]. In a top1Δ strain containing the pTET-lys2-GTOP cassette, reversing the replication orientation by deletion of ARS306 resulted in a three-fold decrease in the Lys+ rate indicating that replication-transcription conflict is a factor in elevated gene conversion initiating at co-transcriptionally formed G4 DNA (Fig. 6B). The deletion of ARS306 did not significantly affect the gene conversion rates at pTET-lys2-GBTM in top1Δ background or at pTET-lys2-GTOP or –GBTM cassette in WT backgrounds.

Discussion
G4 motifs have been implicated in various types of genome instability events. However, the large number of sequences predicted form G4 DNA structures have not all been validated as potential hotspots of genome rearrangements. We here focused on the level of transcription as a singularly important genomic context that distinguishes genetically unstable G4-forming sequence. At the Ig heavy chain locus, class-switch recombination (CSR) in activated B cells requires switch regions consisting of long repetitive sequences dense with guanine-runs [52]. In the pathogenic bacteria Neisseria gonorrhoeae, a G4 DNA-forming sequence was identified to be essential for the gene conversion occurring at the PilE locus, which facilitates evasion of host adaptive immune system by the production of variant pilin subunits [53]. Transcription of the guanine-run containing sequences is required to initiate recombination in both of these processes [54], [55]. One possible role played by transcription is to provide the strand separation necessary for guanine-runs to fold into G4 structures. In order to determine the effect of G4 DNA on yeast genome stability, we designed our genetic assays to assess the role of transcription in biological processes. For effective formation of G4 DNA during transcription, we placed a model G4 motif from the mouse Ig switch Mu region into the highly transcribed pTET-LYS2 cassette. The guanine-runs were placed on non-transcribed strand (-GTOP) to promote co-interaction in single-strand context; as a negative control, genomic instability associated with the same G4 sequence was measured when transcribed in inverse orientation (-GBTM) where guanines on the TS interact with the nascent RNA and are not available for G4 formation (Fig. 1B).
In the gene conversion assay we used previously, only those recombination events initiating specifically at the reporter construct could be phenotypically selected [33]. In the current study, we designed two other genomic assays that allowed us to survey instability initiated over large genomic areas that includes our G4 motif-containing reporter construct. Loss of part of a chromosome arm initiating over a ∼20 kb region of CHR5 in a haploid background (GCR assay) or recombination initiating over a ∼100 kb region of CHR3 in a diploid background (LOH assay) yielded selectable, CanR 5-FOAR or 5-FOAR colonies, respectively (Fig. 1A and 3A). By further analysis of the individual isolates, we were able to estimate the locations of GCR or mitotic recombination initiation. Importantly, this unbiased measurement of spontaneous genome rearrangements enabled us to define the conditions under which GCR and mitotic recombination resulting in LOH are specifically elevated at the highly transcribed G4 motif.
Eukaryotic Top1 has multiple functions during DNA transactions [56]. Both negative and positive supercoils accumulated during transcription are removed by Top1 activity. Together with the type II topoisomerase Top2, Top1 is recruited to the genomic regions undergoing replication [57] and has a role in relieving transcription-replication conflicts [40]. Top1 function can also adversely affect genome stability; its endo-ribonuclease activity generates unligatable single-strand breaks at ribonucleotides embedded in DNA, which leads to replication stress and accumulation of deletion mutations [58], [59]. We have shown here that, at the co-transcriptionally generated G4 DNA, Top1 activity is required to suppress various types of genome instability. Top1 disruption greatly elevated overall GCR rates when the pTET-lys2-GTOP was present on CHR5, with 90% of GCR breakpoints mapping to the G4 DNA-forming sequence (Fig. 1C and Table 1). This elevation was completely dependent on the level of transcription and the location of the guanine-runs on the NTS, reinforcing the conclusion that a potential G4 sequence motif is transformed by transcription into a genome instability hotspot. In the LOH assay, even though the overall mitotic recombination rate was not considerably elevated in the top1Δ/top1Δ background, a significantly higher proportion of the LOH tract breakpoints mapped to the pTET-lys2-GTOP but not to the pTET-lys2-GBTM cassette within the left arm of CHR3 (Fig. 5 and Table 4).
In absence of Top1, the rates of both gene conversion and LOH occurring at the G4-forming sequence were significantly higher when the transcription was in “head-on” or collisional orientation with replication fork movement than when it was in co-directional orientation (Figure 5B and 6B). This suggests that Top1-dependent suppression of G4-associated genome instability involves its activity of resolving transcription-replication conflict in addition to its activity of resolving transcription-associated torsional stress. This also suggests that, in addition to the level of transcription, the relative orientation of replication is an important genomic context that can render certain G4 motifs genetically unstable. Accumulation of stalled replication forks at gene-rich regions have been observed in Top1-deficient cells indicating that one of the ways Top1 prevents genome instability is to prevent replication fork collapse due to collision with transcription [40]. Alternatively, the significantly higher LOH and gene conversion rates observed when the transcription and replication are in “head-on” orientation can be due to the intrinsic asymmetry in the replication process. In “head-on” or co-directional orientation, G-runs are present in the leading strand or lagging strand, respectively. It is possible that, upon encountering G4 DNA, the lagging strand synthesis is less prone to replication arrest since re-priming downstream will allow continued replication fork movement. It was previously reported that replication orientation did not have an effect when the G4-forming human subtelomeric minisatelite CEB1 was placed into the yeast genome [30]. The rates of GCR at this G4 forming sequence were elevated to similar degrees whether the G-runs were on the leading strand or lagging strand. In this experiment, CEB1 was not transcribed, supporting the argument that the orientation bias we observed with the highly transcribed Sμ is due to conflict between replication and transcription.
In cultured mouse B cells, it was reported that class switch recombination (CSR) at Ig heavy chain locus was inhibited by camptothecin (CPT) treatment and significantly elevated by siRNA-mediated knock-down of Top1 [60]. It was suggested that G4 DNA formation is facilitated by reduced Top1 activity and that DNA ligation by Top1, which requires the proper alignment of 3′ and 5′ ends of the breaks, is inhibited by interaction with DNA secondary structures resulting in Top1 cleavage complex and unresolved DNA breaks that initiate CSR. However, we demonstrated here that genome instability associated a G4 motif is stimulated by the complete absence of the Top1 protein (top1Δ) (Fig. 1C, 5B and ref. [33]). DNA breaks initiating gene conversion, LOH or gross chromosomal rearrangements may originate from other sources such as G4-specific nucleases or from collapsed DNA replication forks.
Multiple helicases including human FANCJ, PIF1, BLM and yeast Sgs1 can unwind G4 structures in vitro [61], [62]. In BLM-deficient cells, G4 motifs are frequently found near the transcription start sites of those genes with perturbed expression profile suggesting a role of BLM helicase in G4-mediated gene regulation [63]. Using GCR assays similar to that described above, two independent investigations into the effect of mutation of the 5′-to-3′ DNA helicase Pif1 on the rate of GCR initiated by G4-forming sequences reported dramatically disparate results [30], [61]. Although greatly increased rates of FOAR/CanavanineR colonies were observed in both investigations, further analyses revealed that the 5-FOAR/CanavanineR colonies in yeast cells expressing mutant Pif1 arose mostly via rearrangements or partial loss of the chromosome in one case [30] but mainly via epigenetic silencing of the CAN1 and URA3 genes in another [61]. In case of the G4-associated GCR events occurring due to Top1-deficiency reported here, we demonstrated that the simultaneous resistance to 5-FOA and canavanine resulted from the loss of the region of the CHR5 containing CAN1 and URA3 genes (Table 1). In 60% of the GCR events involving the highly transcribed G4 motif, de novo telomere addition occurred within the guanine-run containing Sμ region (Fig. 2 and Fig. S1). Other types of events included 15 kb segmental deletions and complex genome rearrangements involving terminal deletions and segmental duplications (Fig. 2B. Fig. S2 and Fig. S3). When combined with co-transcriptionally formed G4 DNA, Top1 disruption significantly reshapes the genome not just through elevated non-crossover and allelic interhomolog recombination but also through gross deletions and duplications resulting in copy number variations. This result suggests that, whereas the function of Pif1 and BLM is possibly linked to the role of G4 DNA as an epigenetic and transcriptional regulator [17], [61], [63], Top1 functions directly to prevent chromosomal rearrangements and gross loss of genetic information associated with the G4 DNA, particularly at highly transcribed areas.
Activated transcription through G/C rich sequence can lead to formation of R-loops, which comprise of a long and stable hybrid between nascent RNA and template DNA strand [64]. R-loop accumulation and associated hyper-recombination can ensue when the mRNA packaging and export is disturbed in THO-TREX defective strains or when the degradation of RNA in RNA∶DNA hybrid is deficient due to absence of RNase H activity [65]. Accumulation of negative supercoils in Topoisomerase-deficient cells can also lead to R-loop accumulation [66]. Duquette et al reported that the combination of R-loop and G4 DNA, referred to as G-loop, is identifiable by electron microscopy when the Ig switch sequence is highly transcribed either in vitro or in bacteria [13]. At the pTET-lys2-GTOP cassette containing the Sμ sequence, therefore, the elevated genome instability could be the result of RNA∶DNA hybrid and/or G4 DNA. G-loop formation can be instigated by G4 DNA nucleation in the NTS, which lead to the stable annealing of the nascent RNA with the unpaired TS of DNA (Fig. 7). Alternatively, G-loop formation can initiate via the formation of RNA∶DNA hybrid, which leaves the NTS unpaired and free to fold into G4 structure. In this case, the higher stability of rG∶dC base pairing compared to rC∶dG could account for the greater instability we observed when the G-runs are on the NTS (-GTOP) [67]. During in vitro transcription, rG∶dC containing RNA∶DNA hybrid is critical for the formation of G-loop structure by Ig switch sequence [13], and required for the transcription blockage by a guanine-run [19]. We tested whether G4-induced hyper-recombination is dependent on RNA∶DNA hybrid formation by overexpressing RNase H1 in top1Δ background. RNase H1 is an enzyme that degrades RNA hybridized to DNA and was shown to counteract the hyper-recombination phenotype associated with R-loops in THO/TREX mutant background [65]. As shown in Fig. 7B, RNase H1 overexpression did not reduce the elevated recombination at the highly expressed pTET-lys2-GTOP cassette, which suggests that RNA∶DNA hybrid is not required for the elevated recombination occurring at the G4 motif in absence of Top1. We previously reported that, upon disruption of both RNase H1 and RNase H2 (rnh1Δ rnh2Δ), the rates of gene conversion for the pTET-lys2-GTOP and –GBTM constructs were elevated by 28 - and 8 - fold, respectively [33]. RNase H1 overexpression led to significant decreases in the rates of gene conversion in rnh1Δ rnh2Δ backgrounds indicating that RNA∶DNA hybrid is responsible for the elevated recombination at both the pTET-lys2-GTOP and –GBTM constructs in rnh1Δ rnh2Δ mutant strains (Fig. 7B). We postulate that RNA∶DNA hybrid and G4 DNA can each result in genome instability but that R-loop is not the primary cause of elevated recombination we observed for the pTET-lys2-GTOP upon disruption of Top1. A function of Top1 other than the prevention of R-loop formation is relevant in suppressing genome instability at G4 DNA, which will require further investigation to identify.

In summary, we report here the identification of Top1 as an important factor in suppressing genome rearrangements instigated by co-transcriptionally formed G4 DNA. In the absence of Top1, LOH-inducing mitotic recombination as well as GCR is highly elevated but only when the guanine-run containing sequence is located on the NTS and is highly transcribed. The exquisitely specific effect of Top1 at G4 DNA is underscored by recent reports that, without co-transcriptionally formed G4 DNA, Top1-disruption had no significant effect on GCR or gene conversion rate and even suppressed the GCR or gene conversion occurring in cells with defects in the ribonucleotide excision repair (RER) pathway [68], [69]. One possible explanation for its functional specificity is the high affinity binding of G4 DNA by Top1 demonstrated in in vitro experiments [70]–[72]. Top1 activity in suppressing G4-assoicated genome instability becomes even more important when transcription is in the collisional orientation with replication. This result suggests that, besides the transcription-conferred strand bias, the genomic location relative to a replication origin might determine which of the numerous G4 motifs so far identified in the eukaryotic genomes might be a hotspot of genome instability. The data presented in this report opens up the possibility that other factors suppressing G4-associated genome instability will be found among proteins with known physical and/or genetic interactions with Top1.
Materials and Methods
Yeast strains
Yeast strains used for the GCR assay and the gene conversion assay were derived from YPH45 (MATa, ura3-52 ade2-101 trp101; [73]). For construction of the GCR assay, procedures for deletion of endogenous LYS2 on chromosome II and insertion of tetR′-SSN6 repressor-expressing cassette at LEU2 locus (pCM244 from Euroscarf [74]) were as described previously for the gene conversion assay [33]. A PCR-amplified LYS2 gene fragment was then integrated upstream (centromere proximal) of the CAN1 ORF on the left arm of chromosome V. The LYS2 promoter was replaced with a PCR-generated cassette from pCM225 (Euroscarf) containing the pTET promoter with 7 repeats of tetO and the tetR-VP16 activator coding sequence. The replacement of LYS2 with either the lys2-GTOP or -GBTM allele was carried out using the two-step allele replacement method. Finally, the loxP-flanked URA3Kl cassette was amplified from pUG72 (Euroscarf; [75]) to replace the HXT13 gene through one-step allele replacement. The construction of pTET-LYS2 and pTET-lys2-GTOP or -GBTM (previously referred to as pTET-lys2-SμF and –SμR) cassettes and the genomic integration on chromosome III in the YPH45 strain background were previously described [33], [36]. For the loss of heterozygosity (LOH) assay, the YPH45-derived haploids were mated to an YJM789-derived haploid (MATα, ura3 lys2; [48]).
The plasmid pGAL1-RNH1 with the yeast RNH1 gene under the galactose-inducible GAL1 promoter was a gift from R. Crouch (NCI; Bethesda, MD).
Determination of Rates
For the GCR assay, 5 ml cultures in YEPD medium (1% yeast extract, 2% Bacto-peptone, 2% dextrose, and 250 µg/mL adenine hemisulfate, 2% agar for plates) were inoculated with single colonies and grown for 3 days at 30°C. Cells were then plated either on YEPD or synthetic complete dextrose medium lacking arginine (SCD-arg) and containing canavanine (60 mg/L) and 5-Fluoroorotic acid (5-FOA; 1 g/L). For the LOH assay, 1 ml YEPD cultures inoculated with single colonies were grown for 3 days at 30°C and plated on YEPD or SCD containing 5-FOA (1 g/L). For determination of gene conversion rates, growth and plating conditions were the same as previously described [33]. For RNH1 overexpression experiment, indicated yeast strains were transformed with pGAL1-RNH1 plasmid or pRS416. Individual Ura+ transformants were used to inoculate 1 ml cultures in SCD-Ura media supplemented with 1% raffinose and 2% galacotse. After 4 days growth at 30°C, appropriate dilutions of the cultures were plated on YEPD or SCD-Ura-Lys. For each strain, 12 to 36 cultures were used to determine rates and 95% confidence intervals using the Lea-Coulson method of median [76], [77]. Where indicated, rates were determined using the p0 method [78].
Molecular karyotype analysis of gross chromosomal rearrangements (GCRs)
Characterization of GCR events using pulse field gel electrophoresis (PFGE) and microarray-based comparative genome hybridization (array-CGH) were carried out as previously described [79]. The microarrays used were Agilent custom 8x15k design (AMID 028943), with 14,965 unique 60 nt oligonucleotide probes, and a median genomic spacing of 774 bp. Detailed microarray probe composition and hybridization data are available upon request.
Sequencing the inverted duplication breakpoints of Class III GCRs
The BglII-digested pAG25 plasmid [80] was integrated at a site proximal to the KanMX4 marker present in the class III clones. Genomic DNA from the clones containing the integrated plasmid was extracted and digested with EcoRV followed by re-ligation and rescue of re-circularized plasmids in E. coli. Restriction analyses of the rescued plasmids were consistent with inverted duplication structures. SacI restriction fragments containing the center of symmetry of the inverted duplicated regions were sub-cloned into a pUC18 plasmid vector and sequenced using primers positioned just outside the vector's multicloning site. The sequences of the inverted duplication breakpoints from clones A7 and A8 are shown in Fig. S4A and S4B. The secondary chromosomal rearrangements in class III had breakpoints at Ty/LTR sequences, and were consistent with the ectopic homologous recombination mechanism most often observed in yeast GCRs [81].
Supporting Information
Zdroje
1. MirkinSM (2008) Discovery of alternative DNA structures: a heroic decade (1979–1989). Front Biosci 13 : 1064–1071.
2. McMurrayCT (1999) DNA secondary structure: a common and causative factor for expansion in human disease. Proc Natl Acad Sci U S A 96 : 1823–1825.
3. WellsRD, DereR, HebertML, NapieralaM, SonLS (2005) Advances in mechanisms of genetic instability related to hereditary neurological diseases. Nucleic Acids Res 33 : 3785–3798.
4. McMurrayCT (2010) Mechanisms of trinucleotide repeat instability during human development. Nat Rev Genet 11 : 786–799.
5. BochmanML, PaeschkeK, ZakianVA (2012) DNA secondary structures: stability and function of G-quadruplex structures. Nat Rev Genet 13 : 770–780.
6. WangG, ZhaoJ, VasquezKM (2009) Methods to determine DNA structural alterations and genetic instability. Methods 48 : 54–62.
7. GacyAM, GoellnerGM, SpiroC, ChenX, GuptaG, et al. (1998) GAA instability in Friedreich's Ataxia shares a common, DNA-directed and intraallelic mechanism with other trinucleotide diseases. Mol cell 1 : 583–593.
8. GacyAM, GoellnerG, JuranicN, MacuraS, McMurrayCT (1995) Trinucleotide repeats that expand in human disease form hairpin structures in vitro. Cell 81 : 533–540.
9. DavisL, MaizelsN (2011) G4 DNA: at risk in the genome. EMBO J 30 : 3878–3879.
10. MaizelsN (2006) Dynamic roles for G4 DNA in the biology of eukaryotic cells. Nat Struct Mol Biol 13 : 1055–1059.
11. MaizelsN, GrayLT (2013) The G4 genome. PLoS Genet 9: e1003468.
12. SenD, GilbertW (1988) Formation of parallel four-stranded complexes by guanine-rich motifs in DNA and its implications for meiosis. Nature 334 : 364–366.
13. DuquetteML, HandaP, VincentJA, TaylorAF, MaizelsN (2004) Intracellular transcription of G-rich DNAs induces formation of G-loops, novel structures containing G4 DNA. Genes Dev 18 : 1618–1629.
14. HershmanSG, ChenQ, LeeJY, KozakML, YueP, et al. (2008) Genomic distribution and functional analyses of potential G-quadruplex-forming sequences in Saccharomyces cerevisiae. Nucleic Acids Res 36 : 144–156.
15. HuppertJL, BalasubramanianS (2005) Prevalence of quadruplexes in the human genome. Nucleic Acids Res 33 : 2908–2916.
16. ToddAK, JohnstonM, NeidleS (2005) Highly prevalent putative quadruplex sequence motifs in human DNA. Nucleic Acids Res 33 : 2901–2907.
17. CapraJA, PaeschkeK, SinghM, ZakianVA (2010) G-quadruplex DNA sequences are evolutionarily conserved and associated with distinct genomic features in Saccharomyces cerevisiae. PLoS Comput Biol 6: e1000861.
18. HuppertJL, BalasubramanianS (2007) G-quadruplexes in promoters throughout the human genome. Nucleic Acids Res 35 : 406–413.
19. BelotserkovskiiBP, LiuR, TornalettiS, KrasilnikovaMM, MirkinSM, et al. (2010) Mechanisms and implications of transcription blockage by guanine-rich DNA sequences. Proc Natl Acad Sci U S A 107 : 12816–12821.
20. KrasilnikovaMM, SamadashwilyGM, KrasilnikovAS, MirkinSM (1998) Transcription through a simple DNA repeat blocks replication elongation. EMBO J 17 : 5095–5102.
21. BelotserkovskiiBP, NeilAJ, SalehSS, ShinJH, MirkinSM, et al. (2013) Transcription blockage by homopurine DNA sequences: role of sequence composition and single-strand breaks. Nucleic Acids Res 41 : 1817–1828.
22. PaeschkeK, CapraJA, ZakianVA (2011) DNA replication through G-quadruplex motifs is promoted by the Saccharomyces cerevisiae Pif1 DNA helicase. Cell 145 : 678–691.
23. DuquetteML, HuberMD, MaizelsN (2007) G-rich proto-oncogenes are targeted for genomic instability in B-cell lymphomas. Cancer Res 67 : 2586–2594.
24. NambiarM, GoldsmithG, MoorthyBT, LieberMR, JoshiMV, et al. (2011) Formation of a G-quadruplex at the BCL2 major breakpoint region of the t(14;18) translocation in follicular lymphoma. Nucleic Acids Res 39 : 936–948.
25. WillisTG, DyerMJ (2000) The role of immunoglobulin translocations in the pathogenesis of B-cell malignancies. Blood 96 : 808–822.
26. TangW, DominskaM, GreenwellPW, HarvanekZ, LobachevKS, et al. (2011) Friedreich's ataxia (GAA)n*(TTC)n repeats strongly stimulate mitotic crossovers in Saccharomyces cerevisae. PLoS Genet 7: e1001270.
27. KimH-M, NarayananV, MieczkowskiP, PetesT, KrasilnikovaM, et al. (2008) Chromosome fragility at GAA tracts in yeast depends on repeat orientation and requires mismatch repair. The EMBO J 27 : 2896–3802.
28. ZhangY, ShishkinAA, NishidaY, Marcinkowski-DesmondD, SainiN, et al. (2012) Genome-wide screen identifies pathways that govern GAA/TTC repeat fragility and expansions in dividing and nondividing yeast cells. Mol Cell 48 : 254–265.
29. WojciechowskaM, BacollaA, LarsonJ, WellsR (2005) The myotonic dystrophy type 1 triplet repeat sequence induces gross deletions and inversions. J Biol Chem 280 : 941–993.
30. PiazzaA, SereroA, BouleJB, Legoix-NeP, LopesJ, et al. (2012) Stimulation of gross chromosomal rearrangements by the human CEB1 and CEB25 minisatellites in Saccharomyces cerevisiae depends on G-quadruplexes or Cdc13. PLoS Genet 8: e1003033.
31. RibeyreC, LopesJ, BouleJB, PiazzaA, GuedinA, et al. (2009) The yeast Pif1 helicase prevents genomic instability caused by G-quadruplex-forming CEB1 sequences in vivo. PLoS Genet 5: e1000475.
32. AksenovaAY, GreenwellPW, DominskaM, ShishkinAA, KimJC, et al. (2013) Genome rearrangements caused by interstitial telomeric sequences in yeast. Proc Natl Acad Sci U S A 110 : 19866–19871.
33. KimN, Jinks-RobertsonS (2011) Guanine repeat-containing sequences confer transcription-dependent instability in an orientation-specific manner in yeast. DNA Repair (Amst)
34. ChenC, KolodnerRD (1999) Gross chromosomal rearrangements in Saccharomyces cerevisiae replication and recombination defective mutants. Nat Genet 23 : 81–85.
35. MotegiA, MyungK (2007) Measuring the rate of gross chromosomal rearrangements in Saccharomyces cerevisiae: A practical approach to study genomic rearrangements observed in cancer. Methods 41 : 168–176.
36. KimN, AbdulovicAL, GealyR, LippertMJ, Jinks-RobertsonS (2007) Transcription-associated mutagenesis in yeast is directly proportional to the level of gene expression and influenced by the direction of DNA replication. DNA Repair (Amst) 6 : 1285–1296.
37. MuramatsuM, KinoshitaK, FagarasanS, YamadaS, ShinkaiY, et al. (2000) Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell 102 : 553–563.
38. WangJC (2002) Cellular roles of DNA topoisomerases: a molecular perspective. Nat Rev Mol Cell Biol 3 : 430–440.
39. WuHY, ShyySH, WangJC, LiuLF (1988) Transcription generates positively and negatively supercoiled domains in the template. Cell 53 : 433–440.
40. TuduriS, CrabbeL, ContiC, TourriereH, Holtgreve-GrezH, et al. (2009) Topoisomerase I suppresses genomic instability by preventing interference between replication and transcription. Nat Cell Biol 11 : 1315–1324.
41. PutnamCD, PennaneachV, KolodnerRD (2004) Chromosome healing through terminal deletions generated by de novo telomere additions in Saccharomyces cerevisiae. Proc Natl Acad Sci U S A 101 : 13262–13267.
42. PutnamCD, PallisK, HayesTK, KolodnerRD (2014) DNA repair pathway selection caused by defects in TEL1, SAE2, and de novo telomere addition generates specific chromosomal rearrangement signatures. PLoS Genet 10: e1004277.
43. HermetzKE, NewmanS, ConneelyKN, MartinCL, BallifBC, et al. (2014) Large inverted duplications in the human genome form via a fold-back mechanism. PLoS Genet 10: e1004139.
44. SmithCE, LlorenteB, SymingtonLS (2007) Template switching during break-induced replication. Nature 447 : 102–105.
45. TsaponinaO, HaberJE (2014) Frequent Interchromosomal Template Switches during Gene Conversion in S. cerevisiae. Mol Cell 55 : 615–625.
46. PradoF, AguileraA (2005) Impairment of replication fork progression mediates RNA polII transcription-associated recombination. EMBO J 24 : 1267–1276.
47. WellingerRE, PradoF, AguileraA (2006) Replication fork progression is impaired by transcription in hyperrecombinant yeast cells lacking a functional THO complex. Mol Cell Biol 26 : 3327–3334.
48. WeiW, McCuskerJH, HymanRW, JonesT, NingY, et al. (2007) Genome sequencing and comparative analysis of Saccharomyces cerevisiae strain YJM789. Proc Natl Acad Sci U S A 104 : 12825–12830.
49. LeeP, GreenwellP, DominskaM, GawelM, HamiltonM, et al. (2009) A fine-structure map of spontaneous mitotic crossovers in the yeast Saccharomyces cerevisiae. PLoS Genet 5.
50. TheisJF, NewlonCS (1997) The ARS309 chromosomal replicator of Saccharomyces cerevisiae depends on an exceptional ARS consensus sequence. Proc Natl Acad Sci U S A 94 : 10786–10791.
51. AbdulovicAL, MinesingerBK, Jinks-RobertsonS (2007) Identification of a strand-related bias in the PCNA-mediated bypass of spontaneous lesions by yeast Poleta. DNA Repair (Amst) 6 : 1307–1318.
52. DunnickW, HertzGZ, ScappinoL, GritzmacherC (1993) DNA sequences at immunoglobulin switch region recombination sites. Nucleic Acids Res 21 : 365–372.
53. CahoonLA, SeifertHS (2009) An alternative DNA structure is necessary for pilin antigenic variation in Neisseria gonorrhoeae. Science 325 : 764–767.
54. LeeCG, KinoshitaK, ArudchandranA, CerritelliSM, CrouchRJ, et al. (2001) Quantitative regulation of class switch recombination by switch region transcription. J Exp Med 194 : 365–374.
55. CahoonLA, SeifertHS (2013) Transcription of a cis-acting, noncoding, small RNA is required for pilin antigenic variation in Neisseria gonorrhoeae. PLoS Pathog 9: e1003074.
56. PommierY (2009) DNA topoisomerase I inhibitors: chemistry, biology, and interfacial inhibition. Chem Rev 109 : 2894–2902.
57. BermejoR, DoksaniY, CapraT, KatouYM, TanakaH, et al. (2007) Top1 - and Top2-mediated topological transitions at replication forks ensure fork progression and stability and prevent DNA damage checkpoint activation. Genes Dev 21 : 1921–1936.
58. KimN, HuangSY, WilliamsJS, LiYC, ClarkAB, et al. (2011) Mutagenic processing of ribonucleotides in DNA by yeast topoisomerase I. Science 332 : 1561–1564.
59. WilliamsJS, SmithDJ, MarjavaaraL, LujanSA, ChabesA, et al. (2013) Topoisomerase 1-mediated removal of ribonucleotides from nascent leading-strand DNA. Mol Cell 49 : 1010–1015.
60. KobayashiM, AidaM, NagaokaH, BegumNA, KitawakiY, et al. (2009) AID-induced decrease in topoisomerase 1 induces DNA structural alteration and DNA cleavage for class switch recombination. Proc Natl Acad Sci U S A 106 : 22375–22380.
61. PaeschkeK, BochmanML, GarciaPD, CejkaP, FriedmanKL, et al. (2013) Pif1 family helicases suppress genome instability at G-quadruplex motifs. Nature 497 : 458–462.
62. SunH, KarowJK, HicksonID, MaizelsN (1998) The Bloom's syndrome helicase unwinds G4 DNA. J Biol Chem 273 : 27587–27592.
63. NguyenGH, TangW, RoblesAI, BeyerRP, GrayLT, et al. (2014) Regulation of gene expression by the BLM helicase correlates with the presence of G-quadruplex DNA motifs. Proc Natl Acad Sci U S A 111 : 9905–9910.
64. HamperlS, CimprichKA (2014) The contribution of co-transcriptional RNA:DNA hybrid structures to DNA damage and genome instability. DNA Repair (Amst) 19 : 84–94.
65. HuertasP, AguileraA (2003) Cotranscriptionally formed DNA∶RNA hybrids mediate transcription elongation impairment and transcription-associated recombination. Mol Cell 12 : 711–721.
66. MasseE, DroletM (1999) Escherichia coli DNA topoisomerase I inhibits R-loop formation by relaxing transcription-induced negative supercoiling. J Biol Chem 274 : 16659–16664.
67. SugimotoN, NakanoS, KatohM, MatsumuraA, NakamutaH, et al. (1995) Thermodynamic parameters to predict stability of RNA/DNA hybrid duplexes. Biochemistry 34 : 11211–11216.
68. Allen-SolteroS, MartinezSL, PutnamCD, KolodnerRD (2014) A Saccharomyces cerevisiae RNase H2 Interaction Network Functions To Suppress Genome Instability. Mol Cell Biol 34 : 1521–1534.
69. PotenskiCJ, NiuH, SungP, KleinHL (2014) Avoidance of ribonucleotide-induced mutations by RNase H2 and Srs2-Exo1 mechanisms. Nature 511 : 251–254.
70. MarchandC, PourquierP, LacoGS, JingN, PommierY (2002) Interaction of human nuclear topoisomerase I with guanosine quartet-forming and guanosine-rich single-stranded DNA and RNA oligonucleotides. J Biol Chem 277 : 8906–8911.
71. ArimondoPB, RiouJF, MergnyJL, TaziJ, SunJS, et al. (2000) Interaction of human DNA topoisomerase I with G-quartet structures. Nucleic Acids Res 28 : 4832–4838.
72. ShuaiL, DengM, ZhangD, ZhouY, ZhouX (2010) Quadruplex-duplex motifs as new topoisomerase I inhibitors. Nucleosides Nucleotides Nucleic Acids 29 : 841–853.
73. SikorskiRS, HieterP (1989) A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 122 : 19–27.
74. BelliG, GariE, AldeaM, HerreroE (1998) Functional analysis of yeast essential genes using a promoter-substitution cassette and the tetracycline-regulatable dual expression system. Yeast 14 : 1127–1138.
75. GueldenerU, HeinischJ, KoehlerGJ, VossD, HegemannJH (2002) A second set of loxP marker cassettes for Cre-mediated multiple gene knockouts in budding yeast. Nucleic Acids Res 30: e23.
76. LeaDE, CoulsonCA (1949) The distribution of the numbers of mutants in bacterial populations. J Genet 49 : 264–285.
77. SpellRM, Jinks-RobertsonS (2004) Determination of mitotic recombination rates by fluctuation analysis in Saccharomyces cerevisiae. Methods Mol Biol 262 : 3–12.
78. FosterPL (2006) Methods for determining spontaneous mutation rates. Methods Enzymol 409 : 195–213.
79. ZhangH, ZeidlerAF, SongW, PucciaCM, MalcE, et al. (2013) Gene copy-number variation in haploid and diploid strains of the yeast Saccharomyces cerevisiae. Genetics 193 : 785–801.
80. GoldsteinAL, McCuskerJH (1999) Three new dominant drug resistance cassettes for gene disruption in Saccharomyces cerevisiae. Yeast 15 : 1541–1553.
81. ArguesoJL, WestmorelandJ, MieczkowskiPA, GawelM, PetesTD, et al. (2008) Double-strand breaks associated with repetitive DNA can reshape the genome. Proc Natl Acad Sci U S A 105 : 11845–11850.
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2014 Číslo 12
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Tetraspanin (TSP-17) Protects Dopaminergic Neurons against 6-OHDA-Induced Neurodegeneration in
- Maf1 Is a Novel Target of PTEN and PI3K Signaling That Negatively Regulates Oncogenesis and Lipid Metabolism
- The IKAROS Interaction with a Complex Including Chromatin Remodeling and Transcription Elongation Activities Is Required for Hematopoiesis
- Echoes of the Past: Hereditarianism and
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy