
Silencing of X-Linked MicroRNAs by Meiotic Sex Chromosome Inactivation
During male germ cell formation, the X and the Y chromosomes are inactivated. This process is conserved and it is essential for germ cell generation. It is believed that X/Y silencing affects all protein-coding genes, but the status of miRNAs and other non-coding genes needs further investigation. MicroRNAs from the X-chromosome (X-miRNAs) have been reported as potential silencing escapers, and they have been proposed to play a role in the inactivation mechanism itself. By looking at the individual cell level, we show unambiguously that X-miRNAs are subject to X/Y silencing, a finding that contradicts the current literature. Moreover, we generated mouse mutants in which we forced expression of X-miRNAs during X/Y silencing, and this lead to germ cell death. We propose that X/Y silencing can influence transcription of essential germ cell genes by regulating X-repressors.
Published in the journal:
. PLoS Genet 11(10): e32767. doi:10.1371/journal.pgen.1005461
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1005461
Summary
During male germ cell formation, the X and the Y chromosomes are inactivated. This process is conserved and it is essential for germ cell generation. It is believed that X/Y silencing affects all protein-coding genes, but the status of miRNAs and other non-coding genes needs further investigation. MicroRNAs from the X-chromosome (X-miRNAs) have been reported as potential silencing escapers, and they have been proposed to play a role in the inactivation mechanism itself. By looking at the individual cell level, we show unambiguously that X-miRNAs are subject to X/Y silencing, a finding that contradicts the current literature. Moreover, we generated mouse mutants in which we forced expression of X-miRNAs during X/Y silencing, and this lead to germ cell death. We propose that X/Y silencing can influence transcription of essential germ cell genes by regulating X-repressors.
Introduction
Meiotic sex chromosome inactivation (MSCI) describes the transcriptional silencing of the unsynapsed X and Y chromosomes at the onset of pachynema in mammalian male germ cells [1–5]. Inactivation of the sex chromosome results in the formation of a heterochromatic domain called the sex body [6]. MSCI is one example of a general mechanism, meiotic silencing, which inactivates any chromosome that is unsynapsed during male or female meiosis [7,8]. MSCI imposes a repressive chromatin signature on the X and Y chromosomes that is retained later, during spermiogenesis [9–12]. MSCI and its maintenance are regulated by a broad array of DNA double-strand break (DSB) repair proteins and chromatin modifications [2,5,13]. Male mice with chromosome abnormalities, e.g. XYY, or targeted mutations in meiotic synapsis or recombination genes, e.g. Spo11-/- and Brca1-/- frequently exhibit defective MSCI, and this results in misexpression of toxic sex-linked genes and midpachytene arrest [14–18].
Microarray [9,19], RNA-sequencing [20] and RNA FISH [16] studies have concluded that in mice MSCI is robust, and no example of an X-linked protein-coding gene that is actively transcribed during pachynema has yet been identified. This situation contrasts with that later in spermatogenesis, when expression of some X-linked genes from the repressed X chromosome is facilitated by various mechanisms including gene amplification [16] and establishment of active chromatin marks by the ubiquitin ligase RNF8 [21]. However, the activity of X-derived non-coding RNAs, and especially small RNAs, during pachynema is less well understood.
Interestingly, the X mouse chromosome is enriched in miRNA-encoding genes, and many of these are expressed in a testis-biased manner [22–24]. Song et al. conducted an extensive study of X - miRNA expression patterns, in which miRNA levels were evaluated by RT-qPCR in purified spermatogenic cell populations. 86% of X-linked miRNA transcripts were detected at high levels in pachytene spermatocytes, and it was been suggested that these genes escape MSCI [25]. High pachytene levels of X-miRNA transcripts have been confirmed by RT-qPCR [22,25], RNA-sequencing [23,26,27] and in situ hybridisation [25,27] approaches. Non-coding RNAs have a prominent role in gene silencing, e.g. in X chromosome inactivation [28], and repression of transposable elements and centromeric repeats [29], and it is therefore possible that X-linked miRNAs contribute to the process of MSCI itself. However, definitive proof that X-miRNA genes escape MSCI requires that nascent precursors of miRNAs, so-called pri-miRNAs, are generated during pachynema, and ideally, that these can be visualised as nascent transcripts originating from the otherwise inactive X chromosome, e.g. by techniques such as RNA FISH. We therefore sought to reappraise X-miRNA expression in the male germ line focusing on nascent transcripts.
Results
X-miRNAs are silenced during pachynema
In order to establish whether X-miRNAs are subject to MSCI, we examined their expression during mouse spermatogenesis using RNA FISH (Fig 1A–1G). We focused on spermatogonia, the early diploid germ cell progenitors in which the X chromosome is active, and pachytene spermatocytes, in which MSCI has taken place. There are currently 167 annotated miRNA genes on the X chromosome (source: miRBase version 21), the majority of which fall into clusters. We focused on six clusters, located at different sites on the X chromosome, and expressed in the testis (S1 Fig) [23]. Together these comprise 78 X-miRNAs, and 83% of them have been reported to escape MSCI [25]. We used a combination of antibody staining for the MSCI marker phosphorylated histone H2AFX (γH2AFX) [30], as well as DAPI nuclear staining, in order to accurately substage germ cells.

X-miRNA clusters 1 and 6 reside within introns of the genes Clcn5 and Ftx, respectively, and are transcribed from the same strand as the host genes (Fig 1A and 1F). At miRNA cluster 1 and cluster 6 loci, no putative promoter other than the host gene promoters can be detected upstream the miRNA genes, as assessed by H3K4me3 signal (S2 Fig), and expression of the miRNAs was shown to be dependent on the transcription of Clcn5 and Ftx parental RNAs [31,32]. Clcn5 and Ftx primary transcripts therefore represent the X-miRNA precursor transcripts. We used fluorescently-labelled, denatured fosmid DNA probes spanning the intronic Clcn5 and Ftx X-miRNA containing regions in order to detect X-miRNA precursor transcripts (X-pri-miRNA). Cluster 1 and 6 pri-miRNA FISH signals were observed in spermatogonia (53% and 75% expressing, n = 32 and 51 cells, respectively). However, no pri-miRNA expression could be detected in pachytene spermatocytes (0% expressing, n = 100 cells; Fig 1A, 1F and 1G).
Next, we used fosmid probes to examine expression of the remaining X-miRNA clusters 2, 3, 4 and 5. X-miRNAs located within these clusters do not lie within host genes. For all four clusters, we observed pri-miRNA FISH signals in spermatogonia (23%, 8%, 26% and 14% expressing, n = 43, 46, 42 and 41 cells, respectively; Fig 1B–1E and 1G). In contrast, RNA FISH signals were not observed in pachytene spermatocytes for any of the four clusters (0% expressing for each cluster, n = 100 cells each in each case; Fig 1B–1E and 1G).
The fosmids that we used for our RNA FISH experiments have an average size of 39kb. These probes will detect X-miRNA transcription, but could potentially also detect unannotated transcripts residing in the same locus. To exclude this possibility, we carried out two experiments. For cluster 5, we used recombineering to excise a 7kb segment containing the X-miRNA genes from the fosmid probe. When the resulting, modified fosmid was used for RNA FISH, no signals were observed in spermatogonia (0% displaying signals, n = 41 cells; S3 Fig). Secondly, we designed an RNA FISH protocol to assess transcription of specific X-pri-miRNAs. In this approach, we used ~40 nucleotide-long probes matching sequences present in the pri-miRNA, but not the pre-miRNA or the mature miRNA, at the base of the miRNA-containing stem-loop sequence (S4 Fig). We targeted the X-miRNA miR-465, present in six copies in cluster 4 (Fig 1D). Pri-miRNA signals were observed in spermatogonia but not in pachytene cells (0% expressing, n = 53 cells; S4 Fig). Thus, in conclusion, we observed transcription of all six X-miRNA clusters (total 78 X-miRNAs) in spermatogonia. However, we could not detect expression for any of these X-miRNAs during pachynema.
X-miRNAs are subject to MSCI
The absence of cluster 1 to 6 X-pri-miRNA FISH signals in pachytene spermatocytes suggests that these genes are subject to MSCI. To test this possibility, we repeated our RNA FISH analysis on a mouse model in which MSCI is defective. In Spo11 null male mice, a domain of γH2AFX is formed at pachynema, but this rarely encompasses the X and Y chromosomes, and it is therefore termed the “pseudo sex body” [15,33]. The failure to execute H2AFX phosphorylation on the XY bivalent causes misexpression of sex-linked genes during pachynema in this mutant [16]. We performed pri-miRNA FISH for four representative X-miRNA clusters: 1, 3, 4 and 6 (Fig 2). Pachytene spermatocytes were identified in Spo11 null males by the presence of the γH2AFX-labelled pseudo sex body.
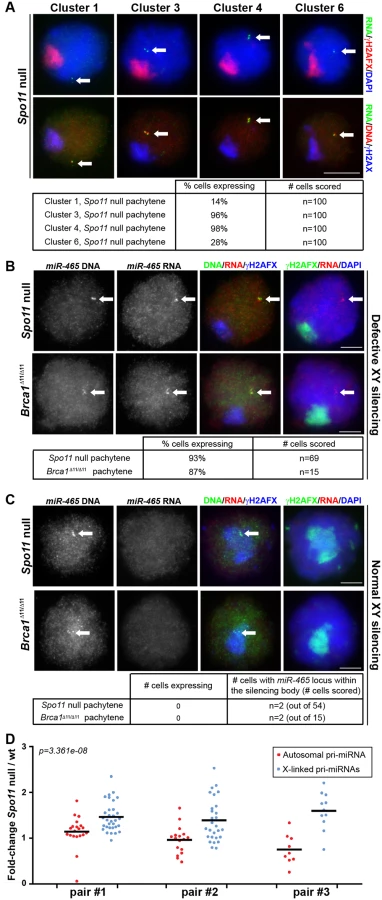
Notably, in Spo11 null pachytene spermatocytes we observed pri-miRNA FISH signals for all four gene clusters studied. Expression was observed in 14%, 96%, 98% and 28% of pachytene cells for clusters 1, 3, 4 and 6, respectively (n = 100 cells in each case; Fig 2A). We subsequently repeated the analysis of cluster 4 X-miRNAs using our oligonucleotide RNA FISH approach that specifically detects pri-miRNAs for the six copies of miR-465. We observed FISH pri-miRNA signals in 93% of Spo11 null pachytene cells (n = 69 cells; Fig 2B). In addition, we performed miR-465-specific pri-miRNA FISH in a second MSCI mutant, the Brca1 Δ11/Δ11 model [17,18]. We detected misexpression of this miRNA in 87% of pachytene spermatocytes. (n = 15 cells; Fig 2B). We conclude that defective MSCI leads to X-miRNA misexpression during pachynema.
Although MSCI is defective in most Spo11-/- and Brca1 Δ11/Δ11 pachytene cells, in both models domains of γH2AFX are occasionally seen covering sub-regions of the X chromosome [16,17]. We predicted that in these rare spermatocytes, X-miRNAs encompassed within γH2AFX regions should be normally silenced. This proved to be the case: in the few Spo11 null and Brca1 pachytene cells in which the miR-465 locus, identified using DNA FISH, lay within a γH2AFX domain (Spo11 null: n = 2 out of 54 cells; Brca1Δ11/Δ11: n = 2 out of 15 cells), no pri-miR-465 expression could be observed (Fig 2C). Thus, the expression status of these X-miRNAs is tightly linked to the presence of the meiotic silencing marker γH2AFX. We conclude that in wild type pachytene spermatocytes, the X-linked miRNAs studied herein are silenced during pachynema as a result of MSCI.
To corroborate our X-miRNA FISH data, we next compared expression levels of individual pri-miRNAs in wild type and Spo11 null sibling testes by RT-qPCR at 15.5days post-partum (dpp; Fig 2D). At this age, most spermatocytes are in pachynema, and genes subject to MSCI are expected to be overexpressed in Spo11 null relative to wild type males. We examined transcript levels for a number of X-linked and autosomal pri-miRNAs, and expressed these as a Spo11 null / wild type ratio. Experiments were performed in triplicate, each time using a different Spo11 null and wild type sibling. In each case, Spo11 null / wild type ratios for autosomal miRNAs averaged ca. 1, indicating no difference in pachytene expression levels between the two genotypes (Fig 2D). Conversely, the ratio for X-linked pri-miRNAs significantly exceeded one (p = 3.361e-08; Fig 2D), thereby confirming that X-linked miRNAs are upregulated in the absence of MSCI.
Finally, we used transgenesis to further investigate whether silencing of X-miRNAs in pachynema is due to MSCI. Previous experiments have demonstrated that X-genes present as transgenes on autosomes continue to be expressed during pachynema [34]. This is because unlike the X chromosome, autosomes are synapsed during pachynema and therefore escape the effects of meiotic silencing. To establish whether pachytene silencing of X-linked miRNA genes was due to their location on the X chromosome, we generated a single copy transgenic line in which X-linked miRNA gene clusters 3 and 4 were located together on an autosome by random BAC integration (X-miRBAC line 1; Fig 3A). We chose a BAC that includes a region of local H3K4me3 enrichment upstream of the miRNA gene cluster, indicative of a putative promoter (Fig 3A). Using pri-miRNA microarrays, we confirmed that cluster 3 / 4 X-miRNAs were overexpressed in X-miRBAC line 1 testes relative to non-transgenic siblings (Fig 3B). We then performed pri-miRNA FISH in pachytene spermatocytes from X-miRBAC line 1 transgenics using BAC probes covering clusters 3 and 4. We observed expression of cluster 3/4 miRNAs from both the X chromosome and the autosomal transgene prior to pachynema (Fig 3C). However, during pachynema, while the X-located 3/4 miRNAs were silenced, those located on the transgene continued to express (100%, n = 50). We observed the same results using our miR-465-specific pri-miRNA FISH protocol on X-miRBAC line 1 (S5 Fig) (100%, n = 9). Thus, silencing of X-integrated miRNAs during pachynema is due to MSCI.

Pachytene X-miRNA expression causes spermatogenic defects
Defects in MSCI cause pachytene arrest, due to misexpression of toxic sex-linked genes, e.g. the Y chromosome genes Zfy1 and Zfy2 [14]. Our analyses indicated that X-miRNAs are subject to MSCI. We therefore wondered whether misexpression of these genes during pachynema would give rise to spermatogenic defects. Interestingly, in our X-miRBAC line 1 males, which carry the autosomally-integrated single copy X-miRNA 3 and 4 cluster transgene, we observed reduced testis weights relative to non-transgenic brothers from as early as five weeks post-partum (Fig 4A). Importantly, histological and TUNEL analysis of X-miRBAC line 1 testis sections revealed spermatogenic defects, principally germ cell apoptosis at stage IV, corresponding to midpachynema, and stage XII, corresponding to the meiotic divisions (Fig 4B, S7 Fig).
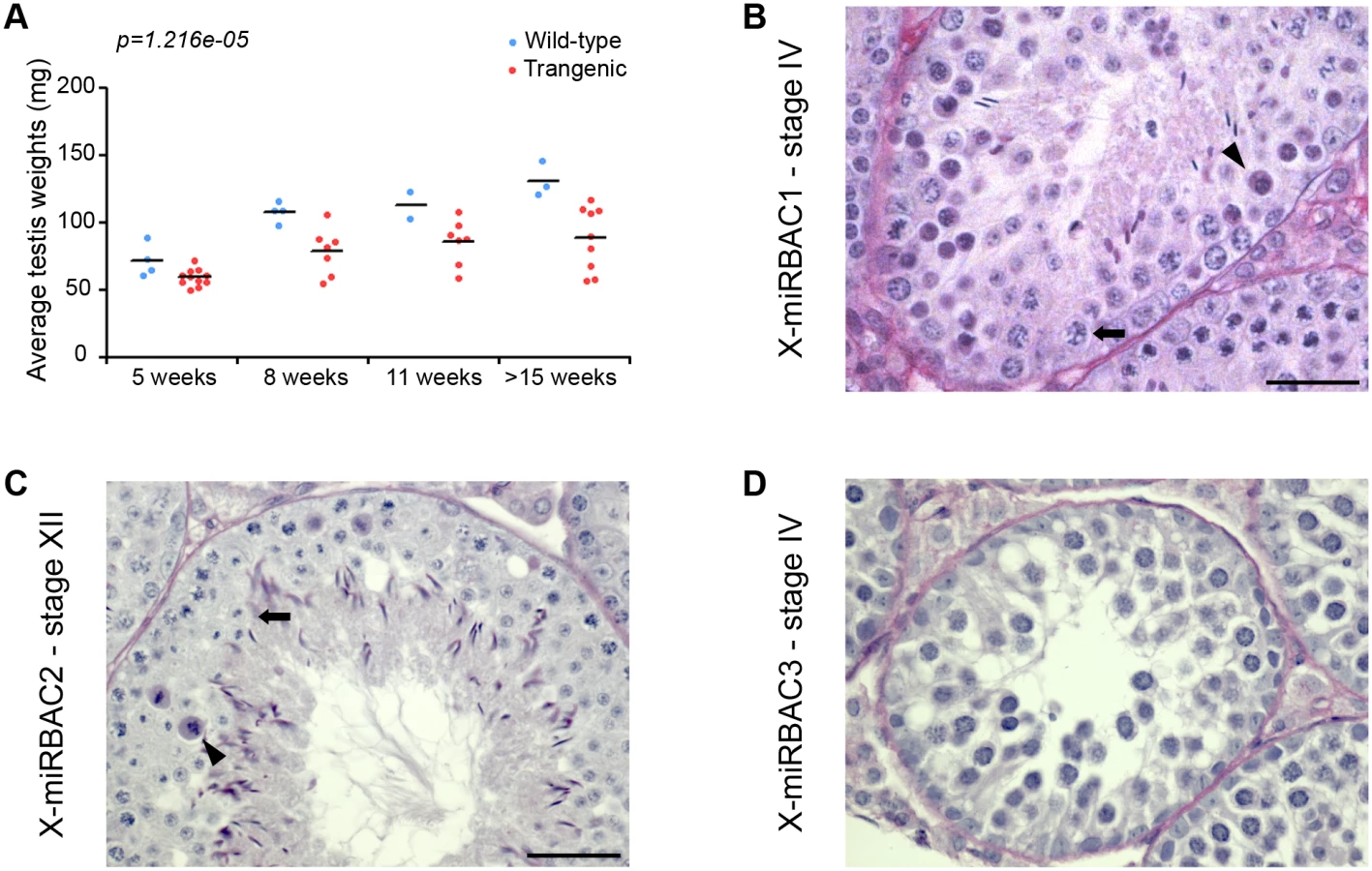
In order to exclude the possibility that the spermatogenic defects observed in X-mirBAC line 1 males resulted from a transgene integration effects, we subsequently generated two more cluster 3/4 X-miRNA autosomal transgenic lines, with three (X-miRBAC line 2) and eleven (X-miRBAC line 3) transgene copies (S6 Fig). X-miRBAC line 2 showed predominant apoptosis at stage XII (Fig 4C, S7 Fig), while X-miRBAC line 3 exhibited marked apoptosis at mid and late pachynema (Fig 4D, S7 Fig). We conclude that inappropriate expression of X-miRNAs from the X-linked clusters 3 and 4 at pachynema induces spermatogenic defects.
Discussion
MSCI is a robust silencing process, affecting most or all protein-coding genes on the mouse X chromosome. However, it is unclear whether silencing also affects X-linked miRNAs. Here, using RNA FISH and other transcriptional assays, we find that X-linked miRNA genes are expressed before prophase I but are silent during pachynema. We therefore conclude that X-miRNAs are subject to MSCI.
It is important to highlight that we did not study all miRNAs on the X chromosome. It is therefore formally possible that X-miRNAs omitted in our analyses behave differently with respect to MSCI. We find this unlikely, because we chose miRNAs from multiple, distinct clusters on the X chromosome, and we included many X-miRNAs that were previously reported to escape silencing [25]. A priori, one could also argue that our inability to detect X-miRNA FISH signals during pachynema is because our RNA FISH experiments lack the sensitivity required to detect gene expression during this stage of prophase I, rather than because these genes are subject to MSCI. We doubt that this is the case, because we were able to detect expression of these species during pachynema both in MSCI mutants, and in mice carrying autosomally-located X-miRNA transgenes. Finally, our conclusion that X-miRNAs are subject to MSCI was corroborated by RT-qPCR analysis in MSCI mutants versus controls. Taken together, our data support a model in which X-miRNAs behave like X-linked protein-coding genes with respect to X silencing.
How can our findings accommodate earlier work? Several independent reports have documented high levels of X-miRNA expression in pachytene spermatocytes [22,23,25,26], and there can be little doubt that this contrasts with the generally low level expression detected for protein-coding X-genes. However, in most existing studies X-miRNAs were assayed at the level of mature miRNAs. Notably, with the exception of some rare cases [35,36], miRNAs have an unusually long half-life, on average 5 days, which exceeds that of protein-coding RNAs by ten-fold [37]. The abundant expression of miRNAs during pachynema might therefore be due to their high transcript stability, rather than ongoing generation of nascent miRNA precursors.
Interestingly, our work shows not only that X-miRNAs are subject to MSCI, but also that failure to silence them can result in spermatogenic defects which are manifest in the case of the cluster 3/4 X-miRNAs as arrest predominantly at stages IV and XII. Thus, X-miRNAs join the Y-encoded Zfy1/2 genes as being male “pachytene-lethal” genes. Our findings show that MSCI must be extensive, silencing genes not only on the Y chromosome but also on the X chromosome. Given that miRNAs act as gene repressors, the phenotypes resulting from their ongoing expression in our miRNA transgenic males are presumably due to inappropriate target downregulation as a consequence of miRNA overexpression, or to an inability to appropriately upregulate target genes with meiotic and/or post-meiotic functions. In this model, MSCI could function as a chromosome-based mechanism for regulating expression of repressors. From a broader perspective, the large-scale silencing of genes across the X chromosome by MSCI is likely to influence myriad transcriptomic networks within germ cells. As a consequence, MSCI could regulate multiple facets of the mammalian germ cell development program.
Materials and Methods
Ethics statement
All animal procedures were in accordance with the United Kingdom Animal Scientific Procedures Act 1986 and were subject to local ethical review.
Mice
All mice were maintained on an MF1 background. The miRBAC transgenic lines were produced by microinjection of purified BAC BMQ-333E20 into fertilized eggs from CBA/Ca x C57Bl/10 F1s. Spo11 null and Brca1Δ11/Δ11 mice have been described previously [18,38].
Genome browser tracks
Browser tracks were generated in R using the Givz package. The UCSC mouse genome assembly mm10 was used as a basis for analysis. Annotation of transcripts was obtained from the UCSC knownGene database. Genomic coordinates of fosmid and BAC probes were obtained from the CHORI BACPAC resource center.
RNAseq analysis
A small RNA testis library was downloaded from GEO under accession identifier GSE40499 (Meunier et al.). Adapter sequences were removed from the reads (ATCTCGTATGCCGTCTTCTGCTTG), and 15 to 23nt-long reads were selected for analysis. Reads were aligned to the mouse mm10 genome using Bowtie (version 1.6.0) with the parameters -m 50—best—strata -v 2. MiRNA gene coordinates were obtained from miRBAse (version 21). MiRNA duplicates sharing a copy on the X chromosome and a copy on an autosome were removed from the analysis. MiRNA read counts were generated in R using the QuasR package as documented in the reference manual [39]. Read counts are expressed as read counts per million reads mapping to miRNA genes.
FISH and immunofluorescence
RNA and DNA FISH was carried out with digoxigenin - and biotin-labelled probes respectively, using fosmid and BAC genomic clones (cluster 1: WI1-603H11; cluster 2: WI1-1995I23; cluster 3: WI1-1646F11; cluster 4: WI1-2045C16; cluster 5: WI1-2828J23; cluster 6: WI1-2859G17; miRBAC: BMQ-333E20). The technique was described previously [40]. For miR-465-specific pri-miRNA FISH, a mix of nine amino-allyl-modified oligonucleotides labeled with fluorolink Cy3 were used as probes (S1 Table). We used the anti-γH2AFX antibody (Upstate, 16–193; dilution 1/100) for immunofluorescence post-RNA FISH.
RT-qPCR
Total RNA was extracted from frozen testis tissues with Trizol (Invitrogen), treated with DNAse I (Invitrogen), and reverse transcribed with random hexamers (Invitrogen) and Superscript II (Invitrogen) according to the manufacturer’s instructions. Quantitative PCR was performed with pre-designed Taqman pri-miRNA and U6 snRNA Taqman assays according to the manufacturer's instructions. Relative expression was calculated with the ΔCt method using U6 as a normaliser. Pri-miRNA expression ratio for individual pri-miRNAs are provided in S2 Table.
ChIPseq analysis
A library of H3K4me3 ChIP in adult germline precursor cells and corresponding input controls were downloaded from GEO under accession identifier GSE49624 [41]. Reads were aligned to the mouse mm10 genome using Bowtie (version 1.6.0) with the parameters Bowtie -m 1—best –strata.
Microarray
Affymetrix Mouse miRNA 2.0 microarrays were performed to measure miRNA expression in testis of three wild type and two homozygous transgenic siblings at 37dpp. Total RNA was extracted with Trizol (Invitrogen), treated with DNAse I (Invitrogen), and column-purified (Ambion). Microarray hybridizations were performed according to the manufacturer's instructions. Microarray signal intensity was extracted and normalized using Affymetrix' miRNA QC Tool using default parameters. Statistical analyses were performed using R and the limma package. Fold-changes and FDR-adjusted p-values were computed by fitting a linear model for each microRNA. Standard errors were smoothed using empirical Bayes (eBayes function of the limma package).
Transgene copy number estimation
The transgene copy number of the miRNA BAC transgenic line was estimated by qPCR on genomic DNA, with a technique adapted from the one described in. The data was normalised with Atr PCR for ΔCt calculations, and quantification of Jun copy number was used as a quality control. Primer sequences are provided in S3 Table.
Recombineering
Recombination-mediated genetic engineering of fosmid WI1-2828J23 was performed to delete a 7kb fragment encompassing miRNA genes miR-201 and miR-547 using standard procedures. The primers used for recombineering are
LN_X8-F (taactagtaagtctaatatattgttgtttaaaacctactgctttgtgctcggcctggtgatgatggcgggatcgttg)
LN_X8-R (ggcccaatgaattatattttctagtacctctcagtatacaaatcaccaactcagaagaactcgtcaagaaggcgata).
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. McKee BD, Handel MA (1993) Sex chromosomes, recombination, and chromatin conformation. Chromosoma 102 : 71–80. 8432196
2. Inagaki A, Schoenmakers S, Baarends WM (2010) DNA double strand break repair, chromosome synapsis and transcriptional silencing in meiosis. Epigenetics 5 : 255–266. 20364103
3. Turner JM (2007) Meiotic sex chromosome inactivation. Development 134 : 1823–1831. 17329371
4. Yan W, McCarrey JR (2009) Sex chromosome inactivation in the male. Epigenetics 4 : 452–456. 19838052
5. Ichijima Y, Sin HS, Namekawa SH (2012) Sex chromosome inactivation in germ cells: emerging roles of DNA damage response pathways. Cell Mol Life Sci.
6. Solari AJ (1964) The Morphology and Ultrastructure of the Sex Vesicle in the Mouse. Exp Cell Res 36 : 160–168. 14222737
7. Turner JM, Mahadevaiah SK, Fernandez-Capetillo O, Nussenzweig A, Xu X, et al. (2005) Silencing of unsynapsed meiotic chromosomes in the mouse. Nat Genet 37 : 41–47. 15580272
8. Baarends WM, Wassenaar E, van der Laan R, Hoogerbrugge J, Sleddens-Linkels E, et al. (2005) Silencing of unpaired chromatin and histone H2A ubiquitination in mammalian meiosis. Mol Cell Biol 25 : 1041–1053. 15657431
9. Namekawa SH, Park PJ, Zhang LF, Shima JE, McCarrey JR, et al. (2006) Postmeiotic sex chromatin in the male germline of mice. Curr Biol 16 : 660–667. 16581510
10. Sin HS, Ichijima Y, Koh E, Namiki M, Namekawa SH (2012) Human postmeiotic sex chromatin and its impact on sex chromosome evolution. Genome Res 22 : 827–836. doi: 10.1101/gr.135046.111 22375025
11. Turner JM, Mahadevaiah SK, Ellis PJ, Mitchell MJ, Burgoyne PS (2006) Pachytene asynapsis drives meiotic sex chromosome inactivation and leads to substantial postmeiotic repression in spermatids. Dev Cell 10 : 521–529. 16580996
12. Greaves IK, Rangasamy D, Devoy M, Marshall Graves JA, Tremethick DJ (2006) The X and Y chromosomes assemble into H2A.Z-containing [corrected] facultative heterochromatin [corrected] following meiosis. Mol Cell Biol 26 : 5394–5405. 16809775
13. Lu LY, Yu X (2015) Double-strand break repair on sex chromosomes: challenges during male meiotic prophase. Cell Cycle 14 : 516–525. doi: 10.1080/15384101.2014.998070 25565522
14. Royo H, Polikiewicz G, Mahadevaiah SK, Prosser H, Mitchell M, et al. (2010) Evidence that meiotic sex chromosome inactivation is essential for male fertility. Curr Biol 20 : 2117–2123. doi: 10.1016/j.cub.2010.11.010 21093264
15. Barchi M, Mahadevaiah S, Di Giacomo M, Baudat F, de Rooij DG, et al. (2005) Surveillance of different recombination defects in mouse spermatocytes yields distinct responses despite elimination at an identical developmental stage. Mol Cell Biol 25 : 7203–7215. 16055729
16. Mueller JL, Mahadevaiah SK, Park PJ, Warburton PE, Page DC, et al. (2008) The mouse X chromosome is enriched for multicopy testis genes showing postmeiotic expression. Nat Genet 40 : 794–799. doi: 10.1038/ng.126 18454149
17. Turner JM, Aprelikova O, Xu X, Wang R, Kim S, et al. (2004) BRCA1, histone H2AX phosphorylation, and male meiotic sex chromosome inactivation. Curr Biol 14 : 2135–2142. 15589157
18. Xu X, Aprelikova O, Moens P, Deng CX, Furth PA (2003) Impaired meiotic DNA-damage repair and lack of crossing-over during spermatogenesis in BRCA1 full-length isoform deficient mice. Development 130 : 2001–2012. 12642502
19. Khil PP, Smirnova NA, Romanienko PJ, Camerini-Otero RD (2004) The mouse X chromosome is enriched for sex-biased genes not subject to selection by meiotic sex chromosome inactivation. Nat Genet 36 : 642–646. 15156144
20. Margolin G, Khil PP, Kim J, Bellani MA, Camerini-Otero RD (2014) Integrated transcriptome analysis of mouse spermatogenesis. BMC Genomics 15 : 39. doi: 10.1186/1471-2164-15-39 24438502
21. Sin HS, Barski A, Zhang F, Kartashov AV, Nussenzweig A, et al. (2012) RNF8 regulates active epigenetic modifications and escape gene activation from inactive sex chromosomes in post-meiotic spermatids. Genes Dev 26 : 2737–2748. doi: 10.1101/gad.202713.112 23249736
22. Ro S, Park C, Sanders KM, McCarrey JR, Yan W (2007) Cloning and expression profiling of testis-expressed microRNAs. Dev Biol 311 : 592–602. 17936267
23. Meunier J, Lemoine F, Soumillon M, Liechti A, Weier M, et al. (2013) Birth and expression evolution of mammalian microRNA genes. Genome Res 23 : 34–45. doi: 10.1101/gr.140269.112 23034410
24. Guo X, Su B, Zhou Z, Sha J (2009) Rapid evolution of mammalian X-linked testis microRNAs. BMC Genomics 10 : 97. doi: 10.1186/1471-2164-10-97 19257908
25. Song R, Ro S, Michaels JD, Park C, McCarrey JR, et al. (2009) Many X-linked microRNAs escape meiotic sex chromosome inactivation. Nat Genet 41 : 488–493. doi: 10.1038/ng.338 19305411
26. Buchold GM, Coarfa C, Kim J, Milosavljevic A, Gunaratne PH, et al. (2010) Analysis of microRNA expression in the prepubertal testis. PLoS One 5: e15317. doi: 10.1371/journal.pone.0015317 21206922
27. Modzelewski AJ, Holmes RJ, Hilz S, Grimson A, Cohen PE (2012) AGO4 regulates entry into meiosis and influences silencing of sex chromosomes in the male mouse germline. Dev Cell 23 : 251–264. doi: 10.1016/j.devcel.2012.07.003 22863743
28. Augui S, Nora EP, Heard E (2011) Regulation of X-chromosome inactivation by the X-inactivation centre. Nat Rev Genet 12 : 429–442. doi: 10.1038/nrg2987 21587299
29. Sabin LR, Delas MJ, Hannon GJ (2013) Dogma derailed: the many influences of RNA on the genome. Mol Cell 49 : 783–794. doi: 10.1016/j.molcel.2013.02.010 23473599
30. Mahadevaiah SK, Turner JM, Baudat F, Rogakou EP, de Boer P, et al. (2001) Recombinational DNA double-strand breaks in mice precede synapsis. Nat Genet 27 : 271–276. 11242108
31. Ruiz-Lafuente N, Alcaraz-Garcia MJ, Sebastian-Ruiz S, Garcia-Serna AM, Gomez-Espuch J, et al. (2015) IL-4 Up-Regulates MiR-21 and the MiRNAs Hosted in the CLCN5 Gene in Chronic Lymphocytic Leukemia. PLoS One 10: e0124936. doi: 10.1371/journal.pone.0124936 25909590
32. Chureau C, Chantalat S, Romito A, Galvani A, Duret L, et al. (2011) Ftx is a non-coding RNA which affects Xist expression and chromatin structure within the X-inactivation center region. Hum Mol Genet 20 : 705–718. doi: 10.1093/hmg/ddq516 21118898
33. Bellani MA, Romanienko PJ, Cairatti DA, Camerini-Otero RD (2005) SPO11 is required for sex-body formation, and Spo11 heterozygosity rescues the prophase arrest of Atm-/ - spermatocytes. J Cell Sci 118 : 3233–3245. 15998665
34. Okamoto I, Arnaud D, Le Baccon P, Otte AP, Disteche CM, et al. (2005) Evidence for de novo imprinted X-chromosome inactivation independent of meiotic inactivation in mice. Nature 438 : 369–373. 16227973
35. Rissland OS, Hong SJ, Bartel DP (2011) MicroRNA destabilization enables dynamic regulation of the miR-16 family in response to cell-cycle changes. Mol Cell 43 : 993–1004. doi: 10.1016/j.molcel.2011.08.021 21925387
36. Krol J, Busskamp V, Markiewicz I, Stadler MB, Ribi S, et al. (2010) Characterizing light-regulated retinal microRNAs reveals rapid turnover as a common property of neuronal microRNAs. Cell 141 : 618–631. doi: 10.1016/j.cell.2010.03.039 20478254
37. Gantier MP, McCoy CE, Rusinova I, Saulep D, Wang D, et al. (2011) Analysis of microRNA turnover in mammalian cells following Dicer1 ablation. Nucleic Acids Res 39 : 5692–5703. doi: 10.1093/nar/gkr148 21447562
38. Baudat F, Manova K, Yuen JP, Jasin M, Keeney S (2000) Chromosome synapsis defects and sexually dimorphic meiotic progression in mice lacking Spo11. Mol Cell 6 : 989–998. 11106739
39. Gaidatzis D, Lerch A, Hahne F, Stadler MB (2015) QuasR: quantification and annotation of short reads in R. Bioinformatics 31 : 1130–1132. doi: 10.1093/bioinformatics/btu781 25417205
40. Mahadevaiah SK, Costa Y, Turner JM (2009) Using RNA FISH to study gene expression during mammalian meiosis. Methods Mol Biol 558 : 433–444. doi: 10.1007/978-1-60761-103-5_25 19685339
41. Hammoud SS, Low DH, Yi C, Carrell DT, Guccione E, et al. (2014) Chromatin and transcription transitions of mammalian adult germline stem cells and spermatogenesis. Cell Stem Cell 15 : 239–253. doi: 10.1016/j.stem.2014.04.006 24835570
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2015 Číslo 10
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Single Strand Annealing Plays a Major Role in RecA-Independent Recombination between Repeated Sequences in the Radioresistant Bacterium
- The Rise and Fall of an Evolutionary Innovation: Contrasting Strategies of Venom Evolution in Ancient and Young Animals
- Genome Wide Identification of SARS-CoV Susceptibility Loci Using the Collaborative Cross
- DCA1 Acts as a Transcriptional Co-activator of DST and Contributes to Drought and Salt Tolerance in Rice
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy