
A Genetic Cascade of Modulates Nucleolar Size and rRNA Pool in
Among the RNA/protein bodies within the nucleus, nucleoli are essential factories for ribosome production and assembly. The size and morphology of the nucleolus is thus a cytological manifestation of protein biosynthesis and is closely coordinated with cell biology and even malignancy. However, without membrane delimitation, the principles that define nucleoli size are poorly understood. Caenorhabditis elegans represents an ideal model to address this question owing to distinct tissue distribution of nucleolar sizes and a mutant, ncl-1, which exhibits larger-than-normal nucleoli. We report here a genetic cascade of microRNA let-7 and translation repressor NCL-1, which tightly controls abundance of FIB-1/fibrillarin. This network ultimately contributes to developmental control of nucleolar size and function.
Published in the journal:
. PLoS Genet 11(10): e32767. doi:10.1371/journal.pgen.1005580
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1005580
Summary
Among the RNA/protein bodies within the nucleus, nucleoli are essential factories for ribosome production and assembly. The size and morphology of the nucleolus is thus a cytological manifestation of protein biosynthesis and is closely coordinated with cell biology and even malignancy. However, without membrane delimitation, the principles that define nucleoli size are poorly understood. Caenorhabditis elegans represents an ideal model to address this question owing to distinct tissue distribution of nucleolar sizes and a mutant, ncl-1, which exhibits larger-than-normal nucleoli. We report here a genetic cascade of microRNA let-7 and translation repressor NCL-1, which tightly controls abundance of FIB-1/fibrillarin. This network ultimately contributes to developmental control of nucleolar size and function.
Introduction
Among the RNA/protein bodies within the nucleus, nucleoli bear the essential function of being the factories for ribosome subunit production and assembly, a stress sensor for cell cycle control, as well as a site for hepatitis D virus (HDV) replication and adenovirus-associated virus (AAV) assembly [1–3]. The size and morphology of the nucleolus is a cytological manifestation of ribosome biogenesis and therefore protein biosynthesis and is closely coordinated with cell growth and development [4]. Accordingly, these attributes sometimes are also physiological indicators of cell cycle, cancer growth and malignancy as well as stem cells differentiation and pluripotency [5, 6]. However, without membrane delimitation, the principles that define nucleoli size and shape are poorly understood. Furthermore, spatiotemporal regulation of nucleolar size and output, particularly in coordination with development and in non-dividing cells, are not fully characterized.
Caenorhabditis elegans represents an exploitable model for further interrogating nucleolus biology owing to distinct distribution of nucleolar sizes in different cell types. A C. elegans mutant, ncl-1, described as a recessive mutation with enlarged nucleoli in nearly all cells of the worm [7, 8], has this phenotype consistent with its role as a suppressor of rRNA biosynthesis. C. elegans ncl-1 phenotypes can be rescued by its Drosophila homolog, brat [9, 10]. Mutations in the fly brat gene have a similar phenotype to the defect of ncl-1 mutants in C. elegans, affecting nucleolar size. In addition, brat mutants induce brain tumor formation [11]. These homologous proteins belong to a TRIM/RBCC/NHL (NCL-1, HT2A, and LIN-41] family characterized by the presence of a RING domain, a B-box zinc finger, and a coil-coiled domain [12, 13]. Because its lack of an RNA binding motif, Brat protein was previously shown to associate with the 3’UTR of the hunchback transcript in partnership with two RNA-binding proteins Pumillio (PUF) and Nanos (NOS), and suppress expression of Hunchback protein at the translational level [14].
In this study, we dissected the molecular mechanism through which NCL-1 controls nucleolar size and function, and pinpointed fibrillarin—the rRNA 2’-O-methyltransferase and pre-rRNA processing factor [1, 15–17]–as a downstream effector. Further, this regulation is dynamically coordinated with development as part of a functional axis driven by let-7, a critical developmental regulator of heterochronic development in worms and flies [18–20] and of cancer formation and stem cell maintenance in the mammals [21].
Results
Suppression of nucleolar size and rRNA expression by NCL-1 is associated with nucleolar protein FIB-1
Although abundantly expressed in the gonads of C. elegans [9], the effect of ncl-1 on the nucleoli of germ cells was not characterized. In intact gonads of wild-type (N2) young adult worms, nucleolar structure is nearly absent in the -1 oocyte, which is immediately adjacent to the spermatheca (Fig 1A, upper panel). In contrast, the nucleolus was readily detectable in the -1 oocyte of ncl-1(e1942) mutant (Fig 1A, lower panel). While nucleoli were evident in the germ cells and the -3 and -2 oocytes of both worms, ncl-1 worms exhibited considerably larger average nucleoli size ranging from 119% to 176% of wild-type diameter (Fig 1A and 1B). Profiling of the ncl-1 mRNA expression by RT-qPCR revealed a progressive decline in mRNA abundance from the embryo to and throughout the four larva stages, followed by subsequent up-regulation in the adult (S1 Fig). This developmental stage-specific expression is consistent with previous in situ immuno-staining of NCL-1 that demonstrated its expression in the proximal gonad and early embryos and the subsequent gradual disappearance in the late stages of embryos [9]. Further, this expression is in line with the non-detectable to small sizes of nucleoli in the -1 oocyte and early embryos (Fig 1C, left panel), supporting the notion that NCL-1 is a negative regulator of nucleolar size.

We also examined nucleolar morphology in worms devoid of functional fib-1. Consistent with its significance, fib-1 mutation led to lethality [22]; we thus characterized fib-1 mutant larvae (L1 stage) and found that nucleoli therein displayed size reduction (S2 Fig). To next determine if FIB-1/fibrillarin is involved in the nucleolar appearance and size, we depleted fib-1 in ncl-1(e1942) worms by RNAi feeding and measured the nucleolar size. The increase in nucleolar size in the blastomeres of ncl-1(e1942) worms (Fig 1C, middle panel) was significantly reversed by fib-1 abrogation as shown by image analysis (Fig 1C, right panel, and 1D). This observation supports a notion that the amount of FIB-1 expression is directly associated with the control of nucleolar size by NCL-1. Moreover, Western blot and RT-qPCR analyses showed that worms expressing a greater amount of FIB-1 generally had a higher level of rRNA abundance (Fig 1D and 1F). Conversely, knockdown of FIB-1 led to an overall reduction in the rRNA levels, further indicating a positive role of FIB-1 in this functional regard.
NCL-1 is a suppressor of FIB-1 expression
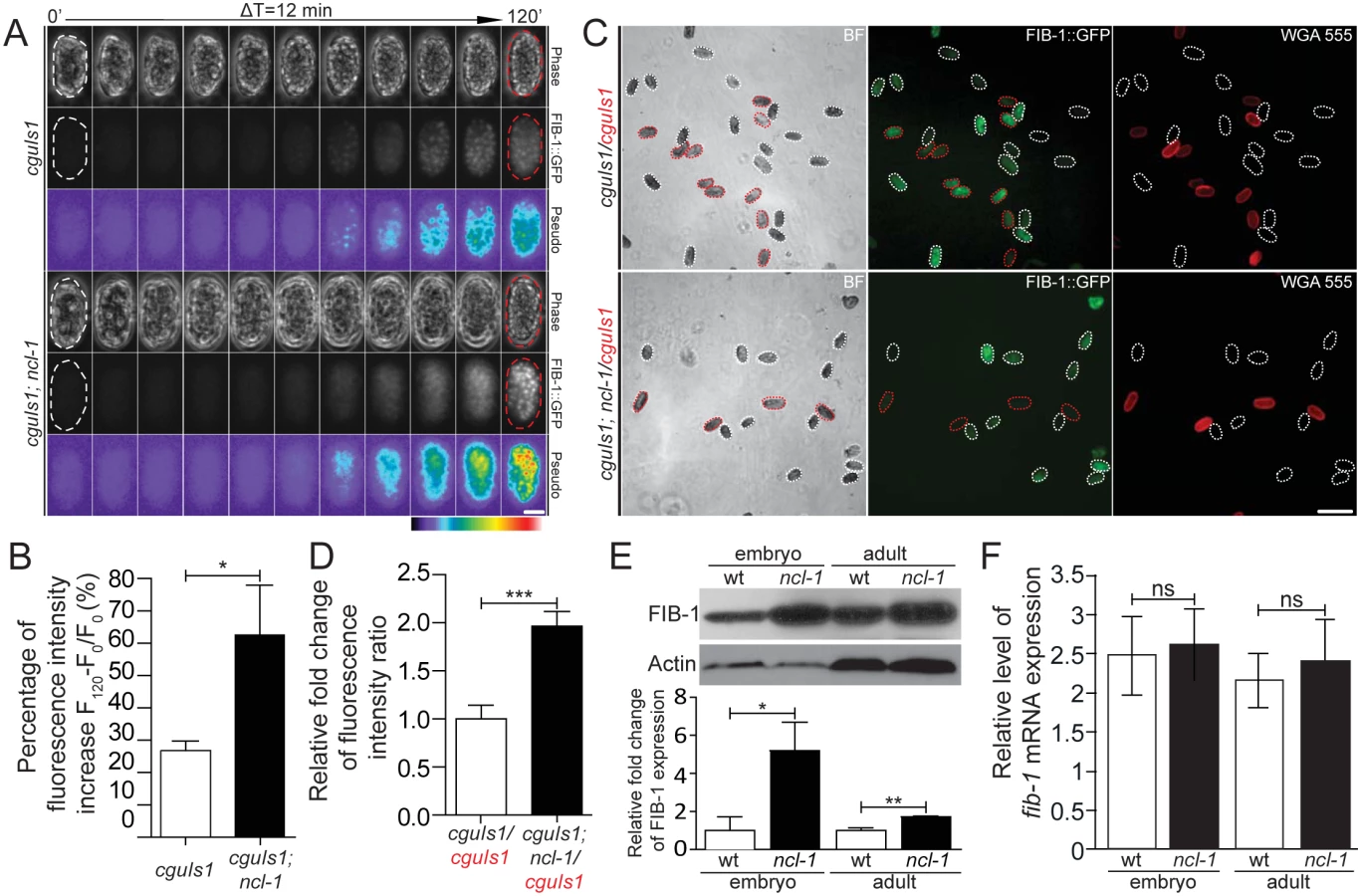
To examine whether NCL-1-mediated nucleolar size alternations is through the regulation of FIB-1 expression, we generated a pair of transgenic worms that express FIB-1::GFP chimeric protein in both the N2 and ncl-1 backgrounds [respectively designated as cguIs1 (strain SJL1) and ncl-1(e1942); cguIs1 (strain SJL14), see S1 Table]. Time-lapse fluorescence microscopy of embryos was performed to trace the level of GFP expression during early stages, and showed progressively higher GFP signals (Fig 2A and S1–S3 Movies). Dynamic up-regulation of GFP levels was more prominent in the ncl-1(e1942); cguIs1 embryos (62.8%) than in cguIs1 (26.9%) (Fig 2B). Random collections of embryos from both transgenic worms were further examined to quantify the GFP intensity of each embryo in the same field (Fig 2C) and subsequently revealed that the embryos in the absence of NCL-1 exhibited higher levels of FIB-1::GFP (about 2 fold) (Fig 2D). Further expression analyses consistently showed elevated levels of FIB-1 in ncl-1(e1942) embryos (5.2 fold) and adult worms (1.7 fold) (Fig 2E). Unexpectedly, RT-qPCR analysis revealed comparable levels of fib-1 mRNA in wild type and ncl-1(e1942) in embryo and adult stages (Fig 2F). Taken together, these findings indicate that ncl-1 is an upstream negative regulator of fib-1 expression at the post-transcriptional/translational stage.
NCL-1 cooperates with PUF and NANOS to modulate fib-1 mRNA translation
We next aimed to test whether NCL-1 acts as its fly homologue Brat, which suppresses its target gene at the translational level by binding to the 3' UTR of transcripts [14]. Towards this end, we created two more pairs of transgenic worms [cguIs2 and ncl-1(e1942); cguIs2 (strain SJL2/strain SJL15), and cguIs19 and ncl-1(e1942); cguIs19 (strain SJL34/strain SJL38), see S1 Table]; SJL2 and SJL15 harbored a plasmid similar to cguIs1 worms that contains the full-length fib-1 3' UTR, while in SJL34 and SJL38 the fib-1 3' UTR was replaced with unc-54 3' UTR sequence (Fig 3A). In agreement with the above observations, enlarged nucleoli and a significantly increased levels of FIB-1::GFP expression were both evident in the tail hypodermis of ncl-1(e1942); cguIs1 and ncl-1(e1942); cguIs2 worms (Fig 3B, top two panels at right, and S3 Fig). In contrast, for the transgene harboring the unc-54 3' UTR, ncl-1 inactivation did not lead to discernable difference in GFP intensity, despite the occurrence of enlarged nucleoli of cells in cguIs19; ncl-1 transgenic worms (Fig 3C). These observations and the quantitative data for the whole worms (Fig 3D and 3E) strongly support the notion that, rather than being the consequence of altered nucleolus, the suppression of FIB-1 may arise from direct targeting of its 3' UTR by NCL-1.

Since Brat mediates its repressive role through other RNA-binding factors, we further tested the roles of C. elegans pumillio and nanos in the translational suppression of fib-1. A potentially direct involvement of these RNA-binding proteins was first supported by the sequence analysis of the fib-1 3’UTR, which revealed a consensus PUF binding motif (Fig 4A). To demonstrate the link between this 3’UTR element and NCL-1-dependent control, we then generated worms with 3’UTR reporter carrying mutations in the PUF binding sequence (cguEx18; Figs 3A and 4A) [23, 24]. Fluorescence microscopy showed that, in comparison to the wild-type reporter (Fig 3B and 3D), this particular transgene exhibited considerably diminished responsiveness to the loss of ncl-1 (Fig 4B and 4C), giving rise to a lower level of fluorescence intensity. In further support to the roles of the PUF proteins, RNAi knockdown puf-5, puf-8 and puf-9 and nos-2 in cguIs1 worms resulted in the appearance of brighter GFP signals (Fig 4D and 4E). However, such effect of nos/puf knockdown (puf-8 and puf-9 in particular) on the GFP reporter was reduced in the cguEx18 worms, in which the PUF binding sequence was altered (Fig 4F). Consistently with the ncl-1 knockdown and mutant worms, immunoblotting showed a rise in FIB-1::GFP abundance in these knockdown worms (Fig 4G). Collectively, these data imply that ncl-1 may coordinate with puf-5, -8, -9 and nos-2 to act directly on the 3' UTR element of fib-1, likely through a similar regulatory mechanism exhibited by brat, pumillio and nanos in the fly [14]. This demonstration of a response element in the fib-1 3’UTR and its regulatory relevance would certainly strengthen a specific and direct control mechanism.

The let-7-ncl-1-fib-1 pathway controls the nucleolus size and rRNA pool
Since another TRIM/RBCC/NHL family protein, LIN-41, is regulated by let-7 [13, 25], we tested for the potential involvement of microRNAs in the regulation of NCL-1. Indeed, potential let-7 and mir-49 target sequences were identified in the 3' UTR of ncl-1 (S4 Fig). As one of the best-known and evolutionarily conserved microRNAs [18, 26, 27], let-7 was selected for further investigation of possible role in ncl-1 expression. Two transcriptional reporters–Pncl-1::gfp::3' UTRncl-1 and Pncl-1::gfp::3' UTRncl-1(m), respectively harboring the wild type and mutated let-7 presumptive sites (Fig 5A), were constructed to each generate multiple independent integration lines of transgenic worms (S1 Table). Immunoblotting results revealed that transgenic worms bearing Pncl-1::gfp::3' UTRncl-1 (strain SJL8) expressed less GFP than those of Pncl-1::gfp::3' UTRncl-1(m) (strain SJL12) (Fig 5B), despite comparable copy numbers and mRNA levels of the transgene between the two strains (Fig 5C). These results thus indicated a loss of responsiveness to let-7 suppression. In further support to the let-7-ncl-1 link, fluorescence microscopic analysis of the seam cells and vulva, which are known to express let-7 [28, 29], indeed showed pronounced GFP reporter expression in the context of defective let-7 targeting (Fig 5D and 5E; S4 and S5 Movies).

We next interrogated the significance of let-7 in the regulation of nucleolar size by assessing vulva cells in the temperature-sensitive, loss-of-function let-7(n2853) mutants. Mutants grown at non-permissive temperature (25°C) displayed a significant reduction in nucleolar size in these cells, by 25% as compared to those at permissive condition (15°C) (Fig 5F and 5G). However, such temperature-sensitive nucleolar size alteration was not observed in a double mutant let-7; ncl-1 (strain SJL39) (Fig 5F and 5G), implying that let-7 acts upstream of ncl-1 transcript to directly suppress NCL-1 translation and regulate nucleolar sizes of the vulva cells. We further verified the link of let-7 to NCL-1-mediated regulation by assessing downstream FIB-1 expression and rRNA abundance in let-7(n2853) and let-7(n2853); ncl-1(e1942) worms. To this end, expression profiling revealed higher amounts of both FIB-1 (Fig 5H) and ribosomal RNA species (Fig 5H and 5I and S5 Fig) in let-7(n2853) worms grown at 15°C vs. 25°C, in contrast to a lack of discernable differences in the let-7(n2853); ncl-1(e1942) worms between these rearing temperatures (Fig 5H and 5I, and S5 Fig). Such loss of phenotypes in the ncl-1(e1942) background is in agreement with let-7-ncl-1 interaction and functional antagonism. Based on these findings, we hypothesize that the genetic circuit of let-7-ncl-1-fib-1 constitutes a critical determinant in the regulation of nucleolar size and rRNA pool (Fig 6).

Discussion
let-7 is known as a critical regulator of heterochronic development in worms and flies [18, 29]. Our studies outlined for the first time a genetic cascade through which the coordinated actions of let-7 and NCL-1 modulate the expression of a major nucleolar protein FIB-1, thereby fine-tuning the size and function of the nucleolus (Fig 6). This circuit of let-7-ncl-1-fib-1 and nucleolus size may represent an adaptive mechanism that couple cellular protein production capacity to the metabolic state of individual cell types. Interestingly, in a recent genome-wide RNAi-based screening for molecular networks underlying nucleolus size regulation in Drosophila, both brat and fib were identified [4], substantiating the possibility that these factors constitute a conserved core of regulatory network. Moreover, Vogt et al. has demonstrated that nucleolus maturation during early embryonic development in mice is dependent on the pluripotency factor LIN28 [30], which is known as an essential regulator of let-7 biogenesis [19, 20, 31]. Intriguingly, Chan and Slack have also shown that ribosomal protein RPS-14 is able to modulate let-7 function [32], which hints at the possibility for a feedback regulation between let-7 and nucleolar dynamics. Our work thus contributes to these findings by reinforcing the relevance of hierarchical organization of post-transcriptional regulators in the fundamental process of nucleogenesis. As FIB-1 expression in C. elegans is also regulated by the die-1 and let363/TOR pathways [33, 34], our findings further support the notion that intricate integration of multiple mechanisms underpins nucleolus integrity.
NCL-1 is a member of TRIM/RBCC-NHL protein family, which has been implicated in the regulation of tumor suppression, cell growth, and cell differentiation. In Drosophila larval neuroblasts (stem cell-like precursors), the Brat homologue is distributed to only one daughter cell through asymmetric cell division and acts as an inhibitor of its self-renewal through post-transcriptional suppression of Myc expression. In Brat mutant, both daughter cells grow and lead to the formation of larval brain tumor [35]. Similarly, the mammalian homologue TRIM3 has been reported as a tumor suppressor in human glioblastoma (GBM), a highly malignant human brain tumor, through its suppression on Myc [36]. Our study complements these findings on the NCL-1 homologues and further provides significant insight into understanding how microRNA cooperates with TRIM/RBCC-NHL proteins to suppress tumor formation.
Despite the prevalent requirement for proper maintenance of nucleolus size, our data did not exclude the possibility that the NCL-1-dependent control mechanism may have tissue - and developmental stage-specific relevance. First, while elevated FIB-1::GFP expression was robustly observed in the ncl-1 mutant, the extent to which it was up-regulated was varied between cells/tissues. A strong evidence for this phenotype is shown in Fig 3B, in which we observed variation in nucleolar size changes between hyp 9 and hyp 10 cells. Second, and perhaps more intriguingly, even in the absence of putative PUF binding site, loss of ncl-1 led to a prominently up-regulated GFP reporter expression in the head region of the cguEx18 worms (Fig 4B). This observation of differential regulation thus implies that 1) there is additional cis-acting element(s) in the fib-1 3' UTR, through which a yet unknown protein mediates brain-specific expression suppression, and/or 2) NCL-1 may functionally cooperates with other neuronal RNA-binding protein(s) to exert a context-dependent regulation of fib-1. This possibility of a modular organization of NCL-1-based regulatory network, as well as its developmental implications, may be further resolved by genetic screens and/or biochemical characterization of NCL-1-interacting factors.
Materials and Methods
Strains and mutant alleles of C. elegans
N2 Bristol C. elegans (used as a wild-type animal control) and mutant strains were obtained from Caenorhabditis Genetic Center (CGC, Minnesota). Alleles of the mutants are as follows: ncl-1(e1942) III, fib-1(ok2527) V, unc-119(ed3) III and let-7(n2853) X. Transgenic worms generated in this study are listed in S1 Table. Five additional strains for this study were generated by the following crosses: SJL14 ncl-1(e1942); cguIs1, SJL15 ncl-1(e1942); cguIs2, SJL38 ncl-1(e1942); cguIs19, SJL39 let-7(n2853); ncl-1(e1942) and SJL118 ncl-1(e1942); cguEx18. Worms were cultured at 15°C or 20°C on NGM plate (1.7% agar, 2.5 mg/mL peptone, 50 mM KH2PO4 pH 6.0, 25 mM NaCl, 5 mg/mL cholesterol, 1 mM MgSO4, 1 mM CaCl2) with fresh Escherichia coli OP50 as food source [37], and synchronized by the protocol with alkaline hypochlorite treatment and the resulting eggs were seeded onto NGM agar plates [38]. Temperature-sensitive mutants (let-7(n2853) X and SJL39 let-7(n2853); ncl-1(e1942)) were maintained at 15°C and shifted to 25°C at the larval 1 (L1) stage and harvested in L4 stage.
Plasmid constructions
Plasmids containing the genomic region of fib-1, including the promoter and 3' UTR of fib-1, were constructed from the PCR amplified DNA fragment from the C. elegans operon CEOP5428 [39]. The gfp gene was inserted into the last codon of fib-1 open reading frame to obtain a translational reporter, Pfib-1::fib-1::gfp::3' UTR fib-1, which encodes a fusion protein of FIB-1::GFP. Another derivative Pfib-1::fib-1::gfp::3' UTRunc-54 was generated by replacing the fib-1 3' UTR with unc-54 3' UTR sequence, and the construct of Pfib-1::fib-1::gfp::3' UTRfib-1(m) was done by site-direct mutagenesis of PUF binding site. To construct two transcriptional reporters, the 1.0-kb promoter and 3' UTR of ncl-1 were cloned into a vector to then generate Pncl-1::gfp::3' UTRncl-1 and Pncl-1::gfp::3' UTRncl-1(m), which differ in the let-7 presumptive targeting sites.
Worm transformation (microinjection and bombardment)
Germ line transformation by microinjection was performed as described by Mello and Fire [40]. Plasmids at the concentration of 100 ng/μl were injected into young adult N2 worms. An integrated line containing the plasmid of Pfib-1::fib-1::gfp::3' UTR fib-1 in about a hundred copies (determined by RT-qPCR) was first obtained in the wild-type background (designated as SJL1 cguIs1). A male of cguIs1 was then crossed with ncl-1(e1942) hermaphrodites, and GFP positive worms were selected. This was followed by hermaphrodite selfing to generate a homozygote worms [SJL14 ncl-1(e1942); cguIs1]. The same method was used to generate the other integration lines (see S1 Table), whereas strains of SJL6 to SJL12 (S1 Table) were obtained by the bombardment method [41].
RNAi treatment
The RNAi library was obtained from Julie Ahringer's group [42–44]. Bacteria clones producing double-stranded RNA to each target gene were grown in LB broth containing ampicillin and tetracycline for 7 to 8 hrs, and subsequently induced to produce double-stranded RNA by 1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) for 2 hrs. Concentrated bacteria were then seeded on RNAi plates (NGM agar, 1 mM IPTG, 100 mg/ml ampicillin, and 5 mg/ml tetracycline), onto which synchronized L1 - L2 stage worms were placed and cultured for 36 hrs at 25°C. Young adult worms were collected for microscopy, RT-qPCR, and/or Western blot analyses.
Western blot
Protein extracts from embryos or worms at L4 or young adult stage were prepared by sonication and separated on 10% or 15% SDS-PAGE and transferred onto polyvinylidene fluoride (PVDF) membranes. Blocked membranes were then incubated with anti-FIB-1 (1 : 2,000 dilution, Santa Cruz) or anti-Actin (1 : 200,000 dilution, Millipore) antibody overnight at 4°C, and subsequently probed with secondary antibody-horseradish peroxidase conjugate (1 : 5,000 dilution, Sigma). Signals were detected with the ECL Western blot detection system (Thermo Scientific Inc., Waltham, MA).
RT-qPCR
Synchronized worms were collected by washing with M9 buffer and then subjected to sucrose density centrifugation to remove OP50 (E. coli) contamination. Total RNA was isolated from a frozen 1 ml aliquot (100 μl worm pellet dissolved in 1 ml TRIzol) by thawing and vigorous mixing according to the manufacturer’s instructions. The genomic DNA was digested by DNase I (Promega). Reverse transcription reactions were performed with iScript Reverse Transcription Supermix for RT-qPCR (Bio-Rad) with 1 μg of RNA. Fifty ng of cDNA was used for each real-time PCR reaction, which was performed with the iCycler IQ real-time PCR detection system (Bio-Rad). For the quantitative detection of ncl-1, fib-1, act-1, gfp and 26S rRNA transcripts, the following primer pairs were respectively used (the act-1 transcript was simultaneously quantified as an internal control):
Qncl-1F’: 5’CAAATCGGAGGCGAGGGAGT3’
Qncl-1R’: 5’CGGAAGGAAGCGGTAGAGGTA3’
Qfib-1F’: 5’ CGTCGTTGGACCAGAAGGAAT 3’
Qfib-1R’: 5’ CACCGTTGCGAAGGAAGTTTT 3’
Qact-1F’: 5’GTGTGACGACGAGGTTGCCGCTCT3’
Qact-1R’: 5’GGTAAGGATCTTCATGAGGTAATC3’
QgfpF’: 5’ CATTGAAGATGGAAGCGTTC 3’
QgfpR’: 5’ ATAGTTCATCCATGCCATGT 3’
Q26S rRNAF’: 5’ GGAGTGCTTGTCTACTGCGAG 3’
Q26S rRNAR’: 5’ CCTCTGCACAGTCACAAGTG 3’
Northern blot analysis
Synchronized late L4 worms grown at 15°C or 25°C as indicated were homogenized by a bead-beating homogenizer (FastPrep-24, MP Biomedicals) and total RNA was isolated by acid guanidinium thiocyanate-phenol-chloroform extraction [45]. Total RNA was subjected to 1.2% agarose-formaldehyde gel electrophoresis (5 μg/lane) and transferred to a Hybond-N+ membrane (GE Healthcare). DNA probes were generated from PCR products amplified from C. elegans genomic DNA and labeled with 32P-dCTP (Perkin Elmer, PK-BLU513H) by hexamer priming. Primers for generating the ribosomal RNA species and actin probes were performed as described by Voutev et al. [46]. Hybridization was carried out at 55°C in 0.36 M Na2HPO4, 0.14 M NaH2PO4, 1 mM EDTA, 10% SDS, 25% formamide and 0.1 mg/ml salmon sperm DNA. Washes were done at 55°C sequentially in 4× SSPE, 4% SDS and 0.1× SSC, 0.1% SDS. Membranes were exposed to Kodak BioMax MS film.
Light microscopy and quantitative image analysis
To observe the FIB-1::GFP expression, embryos or young adult worms of cguIs1 were pre-stained with WGA 555 (50 μg/ml) (Alexa Fluor 555 conjugate of wheat germ agglutinin, Invitrogen) at room temperature for 30 mins (embryos) or 4 h (worms) and collected by washing 3 times with M9 buffer. They were then mixed with embryos or worms of ncl-1(e1942); cguIs1 in an equal ratio and mounted onto 5% agar pad (worms) or a chamber coverglass (embryo) (Thermo) for image acquisition. Bright field and fluorescence images were captured on an inverted or upright microscopy (Leica DMIRE2 and DM2500) using a 10×/NA 0.3 air immersion objective lens and a cool CCD (CoolSNAP K4, Roper Scientific). In order to distinguish the levels of GFP in the experimental and control embryos or worms under a same fluorescence microscope field, the average fluorescence intensity of different strains in the same images was measured using Metamorph 7.7.10.0 offline (Molecular Devices) and quantitatively determined by using Microsoft Excel software. For visualization of FIB-1::GFP expression and nucleolus size in worms, a upright microscope (Leica DM2500) with high-magnification, differential interference contrast (DIC) and fluorescence channels was used; images (shown in enlarged insets) were captured using a 63×/NA 1.4 oil immersion objective lens and a cool CCD (CoolSNAP K4). Metamorph 7.7.10.0 and Microsoft Excel software were used to measure the nucleolus size.
Deconvolution microscopy
For visualization of GFP signals in the vulva and seam cells, transgenic worms at the L4 stage were paralyzed and mounted onto 5% agar pad for z-series image recording. The DIC and fluorescence signals were collected on a Deltavision deconvolution microscope (PersonalDV, Applied Precision) using a 60×/NA 1.4 oil immersion objective lens and a cool CCD (CoolSNAP HQ2, Roper Scientific). The Metamorph software version 7.7.10.0 offline was used in image analysis.
Time-lapse images recording
Embryos of cguIs1 or ncl-1(e1942); cguIs1 as described above were plated onto a chamber coverglass for image acquisition. Phase contrast and fluorescence images were captured on an inverted microscope (Leica DMIRE2) using a 25×/NA 0.95 water immersion objective lens and an electron multiplying (EM) CCD (iXon ultra 897, Andor Technology). Images were recorded at 30s intervals and converted to pseudo-color using Metamorph software.
Statistical analysis
Statistical analyses were performed with a two-tailed Student’s t-test for independent samples by using GraphPad Prism 5 software. P<0.05 was considered statistically significant.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. Lo SJ, Lee CC, Lai HJ (2006) The nucleolus: reviewing oldies to have new understandings. Cell Res. 16 : 530–538. 16775624
2. Pederson T. (2010) "Compact" nuclear domains: reconsidering the nucleolus. Nucleus 1(5):444–5. doi: 10.4161/nucl.1.5.13056 21326828
3. Powell K. (2015) Thoru Pederson: Spotting novel roles for the nucleolus. J. Cell Biol. 208(4):384–5. doi: 10.1083/jcb.2084pi 25688131
4. Neumuller RA, Gross T, Samsonova AA, Vinayagam A, Buckner M, Founk K, et al. (2013) Conserved regulators of nucleolar size revealed by global phenotypic analyses. Sci. Signal 6:ra70. doi: 10.1126/scisignal.2004145 23962978
5. Tsai RY, Pederson T (2014) Connecting the nucleolus to the cell cycle and human disease. FASEB J. 28(8):3290–6. doi: 10.1096/fj.14-254680 24790035
6. Watanabe-Susaki K, Takada H, Enomoto K, Miwata K, Ishimine H, Intoh A, et al. (2014) Biosynthesis of ribosomal RNA in nucleoli regulates pluripotency and differentiation ability of pluripotent stem cells. Stem Cells 12 : 3099–111.
7. Lee LW, Lee CC, Huang CR, Lo SJ (2012) The nucleolus of Caenorhabditis elegans. J. Biomed. Biotechnol. 2012 : 601274. doi: 10.1155/2012/601274 22577294
8. Hedgecock EM, Herman RK (1995) The ncl-1 gene and genetic mosaics of Caenorhabditis elegans. Genetics 141 : 989–1006. 8582642
9. Frank DJ, Roth MB (1998) ncl-1 is required for the regulation of cell size and ribosomal RNA synthesis in Caenorhabditis elegans. J. Cell Biol. 140 : 1321–1329. 9508766
10. Frank DJ, Edgar BA, Roth MB (2002) The Drosophila melanogaster gene brain tumor negatively regulates cell growth and ribosomal RNA synthesis. Development 129 : 399–407. 11807032
11. Arama E, Dickman D, Kimchie Z, Shearn A, Lev Z (2000) Mutations in the beta-propeller domain of the Drosophila brain tumor (brat) protein induce neoplasm in the larval brain. Oncogene 19 : 3706–3716. 10949924
12. Slack FJ, Ruvkun G (1998) A novel repeat domain that is often associated with RING finger and B-box motifs. Trends Biochem. Sci. 23 : 474–475. 9868369
13. Slack FJ, Basson M, Liu Z, Ambros V, Horvitz HR, Ruvkun G (2000) The lin-41 RBCC gene acts in the C. elegans heterochronic pathway between the let-7 regulatory RNA and the LIN-29 transcription factor. Mol. Cell. 5 : 659–669. 10882102
14. Sonoda J, Wharton RP (2001) Drosophila Brain Tumor is a translational repressor. Genes Dev. 15 : 762–773. 11274060
15. Pi H, Lee LW, Lo S.J. (2009) New insights into polycistronic transcripts in eukaryotes. Chang Gung Med. J. 32 : 494–498. 19840506
16. Marcel V, Ghayad SE, Belin S, Therizols G, Morel AP, Solano-Gonzàlez E, et al. (2013) p53 acts as a safeguard of translational control by regulating fibrillarin and rRNA methylation in cancer. Cancer Cell. 24(3):318–30. doi: 10.1016/j.ccr.2013.08.013 24029231
17. Rodriguez-Corona U, Sobol M, Rodriguez-Zapata LC, Hozak P, Castano E. (2015) Fibrillarin from Archaea to human. Biol. Cell Jun;107(6):159–74. doi: 10.1111/boc.201400077 25772805
18. Ambros V (2003) MicroRNA pathways in flies and worms: growth, death, fat, stress, and timing. Cell 113 : 673–676. 12809598
19. Rybak A, Fuchs H, Smirnova L, Brandt C, Pohl EE, Nitsch R, et al. (2008) A feedback loop comprising lin-28 and let-7 controls pre-let-7 maturation during neural stem-cell commitment. Nat. Cell. Biol. 10 : 987–993. doi: 10.1038/ncb1759 18604195
20. Piskounova E, Polytarchou C, Thornton JE, LaPierre RJ, Pothoulakis C, Hagan JP, et al. (2011) Lin28A and Lin28B inhibit let-7 microRNA biogenesis by distinct mechanisms. Cell 147 : 1066–1079. doi: 10.1016/j.cell.2011.10.039 22118463
21. Bussing I, Slack FJ, Grosshans H (2008) let-7 microRNAs in development, stem cells and cancer. Trends Mol. Med. 14 : 400–409. doi: 10.1016/j.molmed.2008.07.001 18674967
22. Piano F, Schetter AJ, Morton DG, Gunsalus KC, Reinke V, Kim SK, et al. (2002) Gene clustering based on RNAi phenotypes of ovary-enriched genes in C. elegans. Curr Biol. 12(22):1959–64. 12445391
23. Bernstein D, Hook B, Hajarnavis A, Opperman L, Wickens M (2005) Binding specificity and mRNA targets of a C. elegans PUF protein, FBF-1. RNA 11 : 447–458. 15769874
24. Wickens M, Bernstein DS, Kimble J, Parker R (2002) A PUF family portrait: 3'UTR regulation as a way of life. Trends Genet. 18(3):150–7. 11858839
25. Stadler M, Artiles K, Pak J, Fire A (2012) Contributions of mRNA abundance, ribosome loading, and post - or peri-translational effects to temporal repression of C. elegans heterochronic miRNA targets. Genome Res. 12 : 2418–26.
26. Pasquinelli AE, Reinhart BJ, Slack F, Martindale MQ, Kuroda MI, Maller B, et al. (2000) Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 408 : 86–89. 11081512
27. Ambros V (2011) MicroRNAs and developmental timing. Curr. Opin. Genet. Dev. 21 : 511–517. doi: 10.1016/j.gde.2011.04.003 21530229
28. Johnson SM, Lin SY, Slack FJ (2003) The time of appearance of the C. elegans let-7 microRNA is transcriptionally controlled utilizing a temporal regulatory element in its promoter. Dev. Biol. 259 : 364–379. 12871707
29. Reinhart BJ, Slack FJ, Basson M, Pasquinelli AE, Bettinger JC, Rougvie AE, et al. (2000) The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 403 : 901–906. 10706289
30. Vogt EJ, Meglicki M, Hartung KI, Borsuk E, Behr R (2012) Importance of the pluripotency factor LIN28 in the mammalian nucleolus during early embryonic development. Development 139 : 4514–4523. doi: 10.1242/dev.083279 23172912
31. Viswanathan SR, Daley GQ, Gregory RI (2008) Selective blockade of microRNA processing by Lin28. Science 320 : 97–100. doi: 10.1126/science.1154040 18292307
32. Chan SP, Slack FJ (2009) Ribosomal protein RPS-14 modulates let-7 microRNA function in Caenorhabditis elegans Developmental Biology 334(1):152–60. doi: 10.1016/j.ydbio.2009.07.011 19627982
33. Sheaffer KL, Updike DL, Mango SE (2008) The Target of Rapamycin pathway antagonizes pha-4/FoxA to control development and aging. Curr. Biol. 18 : 1355–1364. doi: 10.1016/j.cub.2008.07.097 18804378
34. Goldsmith AD, Sarin S, Lockery S, Hobert O (2010) Developmental control of lateralized neuron size in the nematode Caenorhabditis elegans. Neural Dev. 5 : 33. doi: 10.1186/1749-8104-5-33 21122110
35. Betschinger J, Mechtler K, Knoblich JA. (2006) Asymmetric segregation of the tumor suppressor brat regulates self-renewal in Drosophila neural stem cells. Cell 24;124(6):1241–53.
36. Chen G, Kong J, Tucker-Burden C, Anand M, Rong Y, Rahman F, Moreno CS, et al. (2014) Human Brat ortholog TRIM3 is a tumor suppressor that regulates asymmetric cell division in glioblastoma. Cancer Res. 15; 74(16):4536–48.
37. Brenner S (1974) The genetics of Caenorhabditis elegans. Genetics 77 : 71–94. 4366476
38. Stiernagle T (2006) Maintenance of C. elegans. WormBook 11 : 1–11
39. Lee LW, Lo HW, Lo SJ (2010) Vectors for co-expression of two genes in Caenorhabditis elegans. Gene 455 : 16–21. doi: 10.1016/j.gene.2010.02.001 20149852
40. Mello C, Fire A (1995) DNA transformation. Methods Cell Biol. 48 : 451–482. 8531738
41. Praitis V, Casey E, Collar D, Austin J (2001) Creation of low-copy integrated transgenic lines in Caenorhabditis elegans. Genetics 157 : 1217–1226. 11238406
42. Timmons L, Court DL, Fire A (2001) Ingestion of bacterially expressed dsRNAs can produce specific and potent genetic interference in Caenorhabditis elegans. Gene 263 : 103–112. 11223248
43. Timmons L, and Fire A (1998) Specific interference by ingested dsRNA. Nature 395 : 854. 9804418
44. Fraser AG, Kamath RS, Zipperlen P, Martinez-Campos M, Sohrmann M, Ahringer J. (2000) Functional genomic analysis of C. elegans chromosome I by systematic RNA interference. Nature 408 : 325–330. 11099033
45. Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162 : 156–159. 2440339
46. Voutev R, Killian DJ, Ahn JH, Hubbard EJ (2006) Alterations in ribosome biogenesis cause specific defects in C. elegans hermaphrodite gonadogenesis. Dev. Biol. 298 : 45–58. 16876152
47. Tessarz P, Santos-Rosa H, Robson SC, Sylvestersen KB, Nelson CJ, Nielsen ML, Kouzarides T. (2014) Glutamine methylation in histone H2A is an RNA-polymerase-I-dedicated modification. Nature 505(7484):564–8. doi: 10.1038/nature12819 24352239
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2015 Číslo 10
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Single Strand Annealing Plays a Major Role in RecA-Independent Recombination between Repeated Sequences in the Radioresistant Bacterium
- The Rise and Fall of an Evolutionary Innovation: Contrasting Strategies of Venom Evolution in Ancient and Young Animals
- Genome Wide Identification of SARS-CoV Susceptibility Loci Using the Collaborative Cross
- DCA1 Acts as a Transcriptional Co-activator of DST and Contributes to Drought and Salt Tolerance in Rice
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy