
Genetic Analysis of Floral Symmetry in Van Gogh's Sunflowers Reveals Independent Recruitment of Genes in the Asteraceae
The genetic basis of floral symmetry is a topic of great interest because of its effect on pollinator behavior and, consequently, plant diversification. The Asteraceae, which is the largest family of flowering plants, is an ideal system in which to study this trait, as many species within the family exhibit a compound inflorescence containing both bilaterally symmetric (i.e., zygomorphic) and radially symmetric (i.e., actinomorphic) florets. In sunflower and related species, the inflorescence is composed of a single whorl of ray florets surrounding multiple whorls of disc florets. We show that in double-flowered (dbl) sunflower mutants (in which disc florets develop bilateral symmetry), such as those captured by Vincent van Gogh in his famous nineteenth-century sunflower paintings, an insertion into the promoter region of a CYCLOIDEA (CYC)-like gene (HaCYC2c) that is normally expressed specifically in WT rays is instead expressed throughout the inflorescence, presumably resulting in the observed loss of actinomorphy. This same gene is mutated in two independent tubular-rayed (tub) mutants, though these mutations involve apparently recent transposon insertions, resulting in little or no expression and radialization of the normally zygomorphic ray florets. Interestingly, a phylogenetic analysis of CYC-like genes from across the family suggests that different paralogs of this fascinating gene family have been independently recruited to specify zygomorphy in different species within the Asteraceae.
Published in the journal:
. PLoS Genet 8(3): e32767. doi:10.1371/journal.pgen.1002628
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002628
Summary
The genetic basis of floral symmetry is a topic of great interest because of its effect on pollinator behavior and, consequently, plant diversification. The Asteraceae, which is the largest family of flowering plants, is an ideal system in which to study this trait, as many species within the family exhibit a compound inflorescence containing both bilaterally symmetric (i.e., zygomorphic) and radially symmetric (i.e., actinomorphic) florets. In sunflower and related species, the inflorescence is composed of a single whorl of ray florets surrounding multiple whorls of disc florets. We show that in double-flowered (dbl) sunflower mutants (in which disc florets develop bilateral symmetry), such as those captured by Vincent van Gogh in his famous nineteenth-century sunflower paintings, an insertion into the promoter region of a CYCLOIDEA (CYC)-like gene (HaCYC2c) that is normally expressed specifically in WT rays is instead expressed throughout the inflorescence, presumably resulting in the observed loss of actinomorphy. This same gene is mutated in two independent tubular-rayed (tub) mutants, though these mutations involve apparently recent transposon insertions, resulting in little or no expression and radialization of the normally zygomorphic ray florets. Interestingly, a phylogenetic analysis of CYC-like genes from across the family suggests that different paralogs of this fascinating gene family have been independently recruited to specify zygomorphy in different species within the Asteraceae.
Introduction
The evolution of floral symmetry (i.e., the transition between actinomorphy and zygomorphy) is of great interest to plant biologists due to its apparent effect on plant-pollinator interactions and, as a consequence, rates of speciation [1]–[4]. Actinomorphy (i.e., radial symmetry) is typically considered to be the ancestral state [5], [6], with zygomorphy (i.e., bilateral symmetry) having arisen several times during the evolution of flowering plants [1], [6], [7]. Clades with zygomorphic flowers have been shown to be significantly more speciose than their sister clades with actinomorphic (i.e., radially symmetric) flowers [4], presumably because zygomorphy increases pollinator specificity, thereby setting the stage for the evolution of reproductive isolation. Given the above, it has been suggested that the evolution of zygomorphy has played an important role in plant diversification [6], [8].
The Asteraceae is a particularly interesting family in which to investigate the genetics of floral symmetry. Beyond being generally recognized as the most speciose family of flowering plants [9], [10], a large number of species within this family exhibit a radiate flower head, containing both actinomorphic and zygomorphic florets within the same inflorescence. For example, in sunflower (Helianthus annuus L.), the wild-type (WT) inflorescence is composed of multiple whorls of actinomorphic (disc) florets surrounded by a single whorl of zygomorphic (ray) florets (Figure 1A and 1B; Figure S1). The recent elucidation of phylogenetic relationships amongst the major clades of the Asteraceae [11], [12] suggests that ray florets have evolved more than once during the diversification of this family, with a number of tribes and genera containing mixtures of radiate and discoid taxa. Ray florets have also been shown to increase pollination success in species across the family [13]–[15].

The genetic control of floral symmetry has been investigated in several species (e.g., [16], [17]). This has typically been found to involve CYCLOIDEA (CYC)-like TCP transcription factors and, at least in Antirrhinum, their interplay with MYB-like transcription factors [18]. Within the Asteraceae, CYC-like genes have since been shown to play a role in determining floral symmetry in two different species (Gerbera and Senecio [19], [20]). The CYC-like gene family in sunflower was previously found to be twice the size (at least ten members) of that of any other species that had been investigated to date (but see [21] for a more recent report of a similarly large number of CYC-like genes in the Dipsacaceae), and members of this gene family are known to have experienced positive selection and expression divergence following their duplication within the sunflower genome [22].
Sunflower mutants that show alterations in floral symmetry have been previously described (e.g., [23]–[26]), and provide an opportunity to investigate the genetic basis of this trait. For example, in double-flowered (dbl) mutants, the normally actinomorphic disc florets are elongated and vary from strongly zygomorphic towards the outside, to weakly zygomorphic towards the center of the inflorescence (Figure 1C and 1D; Figure S1), more or less reminiscent of WT ray florets. WT rays are sterile whereas WT discs are male and female fertile. In contrast some ray-like disc florets in the dbl mutant do not produce pollen even though anthers are present. In contrast, in tubular-rayed (tub) mutants, the normally zygomorphic ray florets are radialized and contain both stigmas and pollen-producing anthers (Figure 1E and 1F; Figure S1). Interestingly, the dbl mutants bear a strong resemblance to the phenotype captured in Vincent van Gogh's famous 19th century sunflower paintings (Figure 1G), which have become a mainstay of van Gogh exhibits worldwide.
To further our understanding of the genetics and evolution of floral symmetry in the Asteraceae, we investigated the relationship between members of the CYC-like gene family and the aforementioned floral mutants in sunflower. Upon discovering that both the historically-important dbl phenotype as well as the tub phenotype are conditioned by independent mutations in the same member of the sunflower CYC-like gene family, we performed a phylogenetic analysis of CYC-like genes from across the breadth of the family. The results of this analysis suggest that different members of this fascinating gene family have been independently recruited to specify zygomorphy in species across the Asteraceae.
Results
Performing controlled crosses with double-flowered sunflower cultivars is difficult due to reduced pollen production and difficulty in accessing the stigmas. Therefore, we initiated our investigation of floral symmetry in sunflower by crossing a ‘weak’ (i.e., intermediate) dbl individual (cultivar Primrose) to a WT line (cultivar NMS373) (Figure 2). F1 plants exhibited either weak dbl or WT phenotypes in a ratio not significantly different from 1∶1 (χ2 = 0.67, df = 1, P = 0.414), consistent with the effects of a single gene with codominant alleles. Three weak dbl F1 plants were selfed, and scoring of the progeny as WT, weak dbl, or fully dbl (hereafter fully dbl plants are referred to simply as dbl) revealed 1∶2∶1 (WT∶weak dbl∶dbl) segregation in all three families (all Bonferroni-adjusted P>0.05; Fisher's combined probability P = 0.09). Because modifier loci for other dbl lines have been reported [24], and because the phenotypic boundary between weak dbl and dbl plants is not always discrete, these families were also tested against a 3∶1 ([dbl+weak dbl]∶WT) ratio and did not differ significantly from the expectation (P>0.05; Fisher's combined probability P = 0.21).

Self-pollination of four of the WT F1 plants revealed a novel phenotype (tub; tubular-ray florets; Figure 1E and 1F) amongst the resulting progeny. In this case, the phenotypic ratios were not significantly different from 3∶1 (WT∶tub) (all P>0.05; Fisher's combined probability P = 0.10), consistent with the effects of a single gene with a recessive, mutant allele. Selfing of the tub plants resulted in only tub offspring. Moreover, no double-flowered plants with tubular ray florets were observed in any of the F2 families, suggesting that the dbl and tub phenotypes are allelic, or due to the effects of tightly linked genes. This view is supported by the observation that crosses between dbl and tub plants resulted in weak dbl offspring, and that self-pollination of two of these individuals resulted in a 3∶1 ([dbl+weak dbl]∶tub) segregation ratio (both P>0.05). The weak dbl phenotype therefore appears to correspond to dbl/WT (or dbl/tub) heterozygotes.
Genetic mapping in these Primrose×NMS373 F2 families (see Materials and Methods) revealed that both traits map to the same region of sunflower linkage group nine, coincident with the position of three CYCLOIDEA-like (CYC) genes mapped in an earlier study (HaCYC2b, HaCYC2c, and HaCYC2e; [22]). One of the three CYC-like genes (HaCYC2c) showed sequence polymorphism in this population, and exhibited complete cosegregation with the tub phenotype in two WT∶tub populations. All three genes showed sequence polymorphism in a second Primrose×WT population (WT cultivar Moulin Rouge) and, upon mapping, were shown to cosegregate with each other and with the dbl phenotype.
Because of their role in determining floral symmetry in other species, these CYC-like genes are good candidates for being involved in specifying the zygomorphy of WT ray florets. Previous analyses demonstrated that all three CYC-like genes are expressed in floral tissues [22]; however, one of them (HaCYC2c) exhibits ray-specific expression while the other two (HaCYC2b and HaCYC2e) are expressed across multiple floral tissues including rays, discs, ovules and stigmas [22].
Sequencing from the WT parent (NMS373), as well as true-breeding dbl and tub lines, revealed that HaCYC2b and HaCYC2e have identical, uninterrupted coding sequences in all three types, suggesting that these genes are not responsible for the observed phenotypes. In contrast, the sequences of HaCYC2c from both the dbl and tub lines (alleles HaCYC2c-dbl and HaCYC2c-tub, respectively) contained a 999 bp insertion upstream of the start codon, and the HaCYC2c-tub allele contained an additional 1190 bp insertion in the coding region (Figure 3).
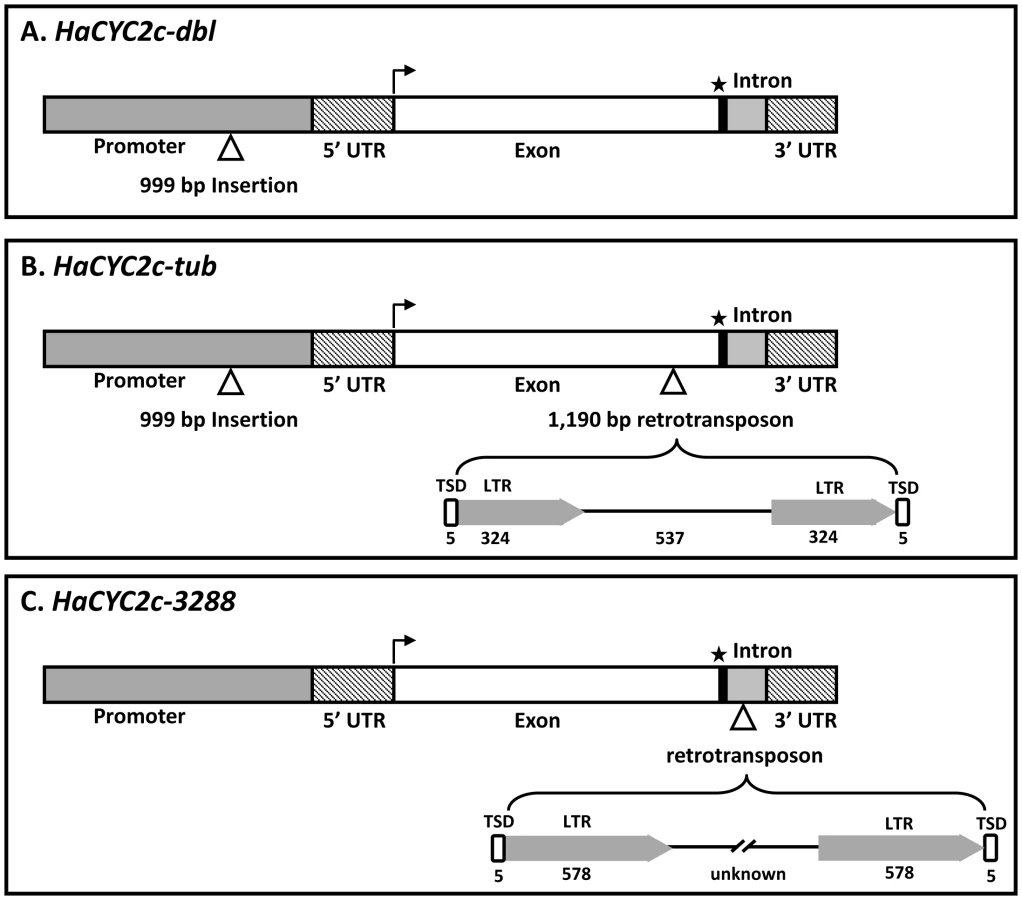
The 999 bp upstream insertion showed no hallmarks of being a transposon or other mobile DNA element. In contrast, inspection of the sequence of the 1190 bp insertion in HaCYC2c-tub revealed a 5 bp target-site duplication (TSD), the presence of identical 324 bp long terminal repeats (LTRs), a primer binding site, and a polypurine tract, suggesting that this insertion is a terminal repeat retrotransposon in miniature (TRIM) [27]. Both mutations in HaCYC2c affect gene expression, causing a deviation from the WT ray-specific expression of HaCYC2c, as follows. In the dbl flower head, HaCYC2c is expressed in all florets across the inflorescence (i.e., in both disc and ray florets), whereas reduced HaCYC2c expression was detected across the head in tub mutants (Figure 4A). It thus appears that the 999 bp insertion affects a ‘ray-floret-specific’ element in the promoter region of HaCYC2c, as evidenced by the expression across all floret types in both the dbl and tub lines. The TRIM insertion apparently reduces expression of HaCYC2c in tub mutants, and also results in the production of a premature stop codon, presumably preventing its WT function. In contrast, expression patterns for HaCYC2b were generally similar across genotypes (Figure 4B) and, while HaCYC2e showed some expression variation across genotypes (Figure 4C), there was no clear evidence of disrupted gene expression resulting in the observed mutant phenotypes. For example, inner and central discs (ID and CD) showed low expression (similar to WT) in dbl and sdbl mutants despite their ray-like appearance, and the Primrose and Ames3288 tub mutants (see below for details on Ames3288) did not show consistent changes relative to WT. These findings suggest that CYC2c is required for zygomorphy in normal WT ray florets, with ectopic expression in disc florets (i.e., HaCYC2c-dbl) causing them to become zygomorphic in dbl mutants, and greatly reduced expression coupled with a truncated mRNA (i.e., HaCYC2c-tub) causing a loss of zygomorphy in tub mutants.

To further investigate this possibility, HaCYC2b, 2c, and 2e were sequenced from a second line with tubular ray florets (USDA accession Ames3288). Whilst HaCYC2b and 2e sequences were identical between Ames3288 and the Primrose tub mutant line (and WT), HaCYC2c contained a unique mutation (HaCYC2c-3288; Figure 3C). In this second tubular-rayed line, a putative retrotransposon (identified on the basis of a 5 bp TSD and 578 bp identical LTRs) had inserted 55 bp downstream of the stop codon, interrupting the intron in the 3′ untranslated region (UTR), and resulting in an almost complete loss of expression (Figure 4A; see below for additional discussion). When this line was crossed with individuals homozygous for the Primrose-derived HaCYC2c-tub mutant allele, all resulting offspring (at least five from each of four independent crosses) exhibited tubular rays (i.e., there was no complementation), presumably due to non-complementary knock-out mutations at HaCYC2c. We therefore conclude that the mutations are indeed allelic, and that mutations in the HaCYC2c gene are responsible for the dbl and tub phenotypes.
HaCYC2c was also sequenced from three other double-flowered sunflower lines. Two of these, Sungold Tall and Chrysanthemoides [24], both harbor the HaCYC2c-dbl allele with the upstream insertion. The third, Teddy-bear, was found to be heterozygous for the same alleles that were present in the original cv. Primrose individual (i.e., HaCYC2c-dbl and HaCYC2c-tub). When additional Teddy-bear plants were grown and self-pollinated, we observed some individuals with the dbl phenotype and others with tubular ray florets. Hence, the HaCYC2c-dbl and HaCYC2c-tub alleles appear to be segregating in this line in the same manner as in Primrose. Expression of HaCYC2c in Chrysanthemoides follows the same overall pattern as Primrose-dbl, confirming our observation of mis-expression of this mutant allele in a different genetic background (Figure 4A).
In order to further examine whether the three insertions (HaCYC2c-dbl, HaCYC2c-3288 and HaCYC2c-tub) are responsible for the mutant phenotypes, we carried out a polymerase chain reaction (PCR) screen of a diverse collection of 108 sunflower lines that exhibit WT floral morphology (see Materials and Methods). This screen revealed that all three insertions are indeed unique to the respective mutant lines (i.e., they were never observed in WT lines), providing further evidence that the HaCYC2c gene plays a critical role in proper floret development, with mis-expression and loss of function of this gene giving rise to the dbl and tub phenotypes, respectively.
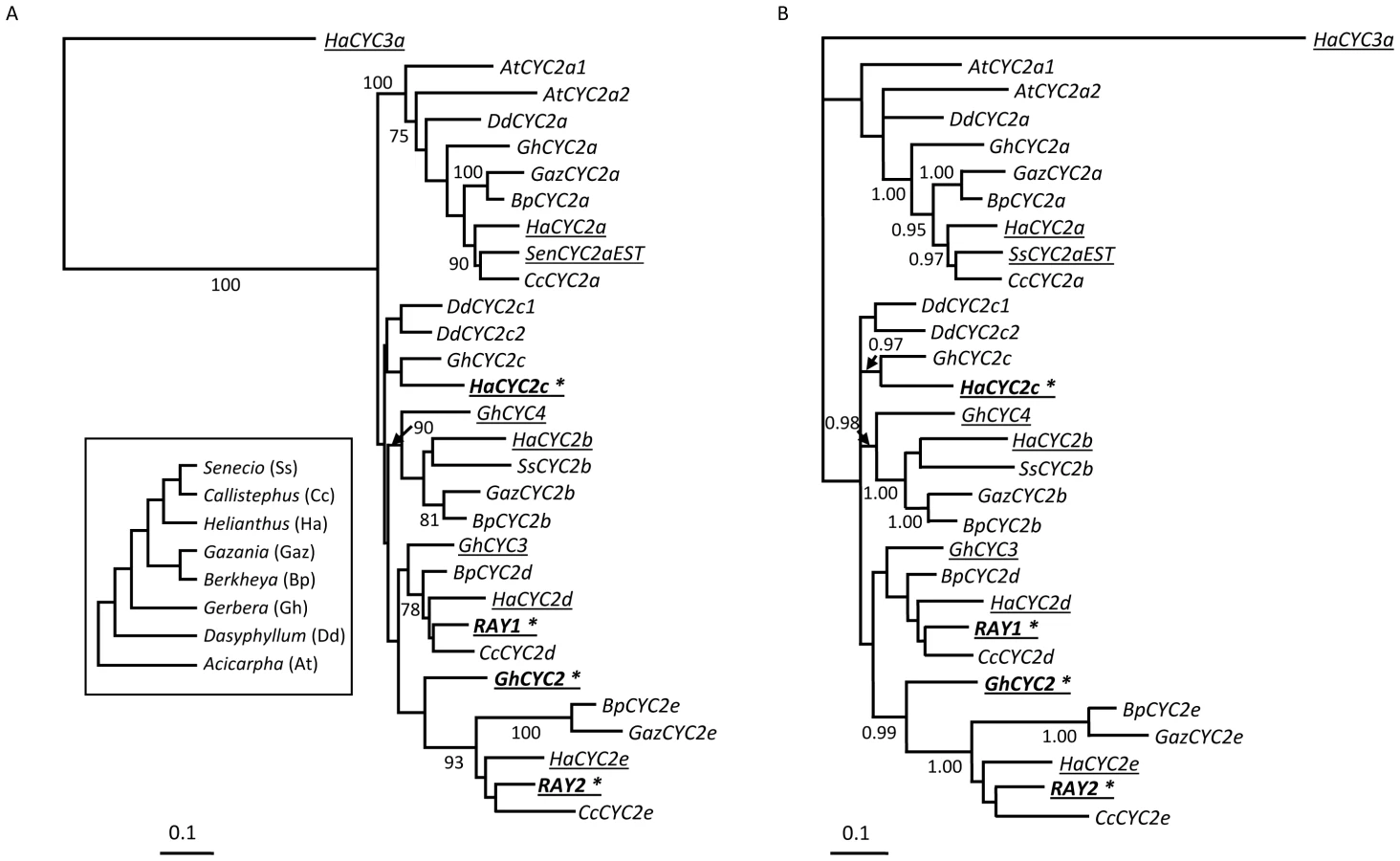
To better understand the diversification of the CYC-like gene family within the Asteraceae, CYC2-like genes were isolated from other radiate members of the family, a basal species with actinomorphic and zygomorphic florets and a member of the sister family, the Calyceraceae, with only actinomorphic flowers (see Materials and Methods), and gene trees were constructed (Figure 5). The focus here was on the CYC2-like subfamily because it is CYC2 genes (as opposed to CYC1 or CYC3) that are responsible for specifying zygomorphy in a wide range of species [3], [16]–[20]. The gene trees suggest that the sunflower HaCYC2c gene, which is responsible for specifying ray floret formation, is paralogous to (i.e., not the direct ortholog of) the Senecio and Gerbera CYC-like genes that have previously been shown to influence floral symmetry in these other members of the Asteraceae (Figure 5). The Gerbera locus controlling floral symmetry is GhCYC2, which is not orthologous to HaCYC2c; rather, this gene is part of the HaCYC2e-like clade, along with one of the two Senecio floral symmetry genes, RAY2. Although there is only weak support for some branches in these trees, the second Senecio symmetry gene, RAY1, also falls outside of the well-supported HaCYC2c clade, grouping instead with HaCYC2d.
Discussion
The evolution of floral zygomorphy has been proposed as a key innovation in angiosperm evolution, with elevated divergence rates thought to be the result of adaptation to specialized pollinators [1], [2], [28]. As noted above, zygomorphy has evolved on multiple occasions, and the role of CYCLOIDEA-like genes in specifying zygomorphy has been implicated in a number of these cases [3], [16]–[20]. The connection between CYC-like genes, floral symmetry, and pollination syndromes is further evidenced by instances in which changes in CYC gene number and/or expression patterns correlate with alterations of floral symmetry [29]–[34], and at least one case in which the loss of CYC-like genes (and their downstream targets) caused a reversion from zygomorphic to actinomorphic flowers and a concomitant change in pollination syndrome from insect to wind pollination [35].
In sunflower, which has both actinomorphic and zygomorphic florets within the same inflorescence (Figure 1), our results indicate that HaCYC2c is necessary for floral zygomorphy, with individuals that are homozygous for a loss-of-function mutation (either due to a premature stop codon or to a reduction/loss of expression of HaCYC2c) exhibiting actinomorphic ray florets. Moreover, the historically important double-flowered mutation, which was captured by van Gogh in his famous late-19th century sunflower paintings, appears to be conditioned by a different mutation in the same gene. In this case, ectopic expression of HaCYC2c across the entire inflorescence results in the transformation of normally actinomorphic disc florets into zygomorphic ray-like florets.
It has previously been shown that CYCLOIDEA-like genes that control zygomorphy act in a dorsal-specific manner, repressing cell growth in this region and allowing outgrowth of the ventral petals [16], [17]. The phenotypes of the tub and dbl mutants are therefore in line with this mode of gene action. That is, loss of function (tub) causes radialization of the ray florets, whereas expression in disc florets (dbl) results in ventralization. It is interesting that, in the tub lines, the outer florets were not simply radialized ray florets; rather, they were effectively transformed into elongated disc florets, as indicated by the presence of stigmas and pollen-producing anthers. This result, along with the observed transformation of disc florets in dbl mutants, suggests that HaCYC2c is responsible for specifying ray vs. disc floret identity, and not simply required for zygomorphy of the ray florets – though the situation is clearly complex, as the radialized ray florets still exhibit elongation and the zygomorphic disc florets in dbl mutants still produce anthers and stigmas.
Given that all four of the dbl lines investigated herein carried the same mutant allele (HaCYC2-dbl), it seems likely that this mutation arose just once, and has been incorporated into multiple cultivars because it produces a desirable floral morphology. In contrast, sunflower lines that exhibit tubular ray florets appear to have arisen at least twice. In one case (HaCYC2c-tub), a transposon insertion into the coding region of this gene appears to have resulted in reduced expression and production of a premature stop codon, resulting in radialization of the ray florets. The second case (HaCYC2c-3288) involves the insertion of a different transposon into an intron in the 3′ UTR of this gene, resulting in a loss of gene expression and radialization of the ray florets. In fact, previous studies have revealed that introns in UTRs can play a vital role in gene regulation [36], [37]; hence, the interruption of an intron in a UTR has the potential to disrupt gene expression, as appears to be the case here.
It is noteworthy that the HaCYC2c-dbl and HaCYC2c-tub alleles both contain the 999 bp upstream insertion resulting in a loss of ray-specific expression, and differ only by the presence (HaCYC2c-tub) or absence (HaCYC2c-dbl) of the TRIM insertion into the coding region (Figure 3). This finding implies that the HaCYC2c-tub allele is derived from HaCYC2c-dbl, and further suggests that the family of elements to which the CYC2c TRIM insertion belongs may still be active in the sunflower genome. The apparent recency of this insertion is further evidenced by the fact that the element in question appears to be intact, complete with identical LTR sequences. In contrast, the HaCYC2c-3288 allele appears to be derived from a WT HaCYC2c allele. The high level of sequence similarity between the coding regions of WT alleles and HaCYC2c-3288, along with the identical LTR sequences, likewise suggest that this was an evolutionarily recent insertion event.
We previously suggested [22] that the large number of CYC-like genes in sunflower was due in part to a whole genome duplication event at the base of the Heliantheae (i.e., the sunflower subfamily) [38]. These duplications are, however, clearly shared with other members of the family outside the Heliantheae, suggesting that the radiation of this gene family occurred earlier in the evolution of the Asteraceae. Moreover, the apparent absence of CYC2b, c, d, and e genes from Acicarpha and CYC2b, d, and e genes from Dasyphyllum (Figure 5) suggests that some members of this gene family may have been lost in certain lineages, or that some duplications occurred since the split between Gerbera and Dasyphyllum, giving rise to the 2b, d, e clade. A more thorough investigation of the genetics of floral symmetry in the basal members of the family is clearly warranted.
Interestingly, inspection of the CYC2-like gene tree from across the Asteraceae (Figure 5) reveals that the sunflower HaCYC2c gene is not the direct ortholog of the CYC2 genes that have been shown to be responsible for specifying zygomorphy in two other members of the Asteraceae (i.e., Senecio and Gerbera). Rather, HaCYC2c appears to be paralogous to these genes, suggesting that the evolution of zygomorphy within the Asteraceae, which is thought to have occurred multiple times [11], [39], involved the parallel co-option of different members of the same gene family for an analogous function.
It is possible that other, closely related CYC-like genes have functions similar to HaCYC2c. In this context, the role of HaCYC2e is of particular interest because: (1) HaCYC2e and HaCYC2c both co-segregate with the mutant phenotypes, and (2) HaCYC2e appears to be the true sunflower ortholog of RAY2 and GhCYC2 (Figure 5b), which are known to influence zygomorphy in Senecio and Gerbera. In this light, it is noteworthy that HaCYC2e is much more broadly expressed than HaCYC2c in the WT inflorescence ([22] and Figure 4; i.e., the former is expressed in all floral tissues examined, albeit at low levels near the center of the disc, while the latter is ray-specific), and that HaCYC2e expression patterns did not clearly correlate with the mutant phenotypes. Moreover, the upstream and coding sequences of HaCYC2e were identical between WT, dbl, and tub individuals, whereas for HaCYC2c, mutations that gave rise to altered expression patterns and/or predicted protein sequences were observed. Nonetheless, a better understanding of the possible role of HaCYC2e in floral development in sunflower awaits further investigation. We can, however, conclude that HaCYC2c is a key transcription factor in the developmental pathway resulting in the development of zygomorphic ray florets in sunflower.
The parallel evolution of adaptive traits has been documented many times [40]. In some cases, such parallel phenotypic changes have been shown to result from parallel molecular changes [41], [42]. In other cases, however, it has been shown that parallel phenotypic evolution results from the evolution of different genes to perform the same function, suggesting that independent gene co-option may be an important mechanism for the origin of evolutionary novelty [43]–[46]. In contrast to the co-option of unrelated genes/proteins to fill the same functional role in independent evolutionary lineages, the co-option of different, but related genes (i.e., those with ancestral similarity, such as different members of the same gene family) has been less well documented. It has, however, recently been shown that O2-transporting hemoglobins have evolved independently from different members of the globin gene superfamily in jawed vs. jawless vertebrates [47]. While CYC2-like genes have been implicated in controlling zygomorphy in several plant species (reviewed in [48]), our work provides evidence that subfunctionalization of different paralogs within this gene family has resulted in the independent evolution of an analogous adaptive trait in evolutionarily-independent lineages.
Materials and Methods
Plant Material and Crosses
Most of the sunflower (Helianthus annuus L.) lines used in this research were obtained from the USDA North Central Regional Plant Introduction Station (NCRPIS; Ames, IA). The exceptions were Chrysanthemoides [24], which was kindly provided by Dr. Claudio Pugliesi (Università di Pisa, Italy) and Moulin Rouge, which was obtained from Johnny's Selected Seeds (Winslow, Maine, USA). USDA Plant Introduction (PI) numbers for the wild-type parent and mutant lines are as follows: NMS373 (PI 597362), Primrose (PI 490320), Ames3288 (PI 650394), Teddy-bear (PI 650838), Sungold Tall (PI 490322). The 108 WT lines that were screened for insertions in HaCYC2c (see below) are listed in Table S1. When crosses were made, florets were emasculated and heads were bagged pre - and post-pollination to prevent pollen contamination. Individuals were scored as wild-type (WT), weak double-flowered (weak dbl), fully double-flowered (dbl), or tubular-rayed (tub) based on the morphology of their florets.
Genetic Mapping
Microsatellite markers showing linkage with the three phenotypes (i.e., WT vs. dbl; WT vs. tub) were first identified by extracting DNA from individuals of the three phenotypes using MagAttract 96 or DNeasy DNA extraction kits (Qiagen, Valencia, CA, USA), bulking equal amounts of DNA from ten individuals per phenotype (WT, dbl, and tub) and genotyping the bulked samples using multiplexed polymerase chain reaction (PCR) amplification [49]. Alleles were visualized using GeneMapper (Applied Biosystems, Carlsbad, California, USA). Both phenotypes showed an association with marker ORS844 on linkage group (LG) nine, so additional markers (ORS1265, ZVG39, CRT250, and ORS176; also from ref. 49) from this LG were genotyped in 96 F2 progeny from a WT×dbl cross and 192 F2 progeny from two WT×tub crosses, all of which were derived from the original NMS 373×Primrose cross (Figure 2). Genetic mapping was carried out using Mapmaker 3.0 (Lander et al. 1987; Lincoln et al. 1992) following ref. 22. The HaCYC2c-tub insertion was also mapped in the two Primrose×NMS373 F2 families segregating for WT∶tub phenotypes (n = 192 F2 plants) and complete co-segregation between the HaCYC2c-tub allele and the tub phenotype was observed.
Because the alleles of both HaCYC2b and HaCYC2e were identical in the parents of the aforementioned mapping populations, these genes could not be re-mapped in these crosses. Therefore, single nucleotide polymorphism (SNP) markers were developed for all three loci in an independent WT (Moulin Rouge)×Primrose mapping population (n = 195 F2 plants) where they showed sequence differences. Loci were PCR amplified in 5 µl reaction volumes containing 2 ng genomic DNA, 1 µL 1× LightCycler 480 Genotyping Master mix (Roche Diagnostics, Indianapolis, IN, USA), 1.0 µM of excess primer, 0.5 µM of limiting primer, and 0.2 µM of both the sensor and anchor probes (Table S2). PCR was performed in the Light Cycler 480 (Roche) for 45 cycles with 10 sec 95°C, 15 sec at 55°C, and 20 sec at 72°C. The final melting cycle was performed by raising the temperature to 95°C for 3 min, lowering the temperature to 40°C for 3 min, and increasing the temperature to 85°C with continuous fluorescent acquisition 5 times/degree. The fluorescence signal (F) was plotted in real time against temperature (T) to produce melting curves for each sample, and the melting curves were converted to negative derivative curves of fluorescence with respect to temperature (-dF/dT) by the LightCycler Data Analysis software (Roche).
DNA Sequencing and Allele Screening
HaCYC2-like genes were PCR-amplified (see primer sequences in Table S2) and sequenced using previously established protocols (e.g., ref. 50). Insertions in CYC2c were found in the mutant genotypes that were initially screened (Primrose and Ames3288; Genbank accession numbers HQ891026–HQ891029). HaCYC2c was also sequenced from three additional dbl mutants (Sungold Tall, Chrysanthemoides, and Teddy-bear; Genbank accession numbers JF489909–JF489913). Primer pairs (Table S2) specific to all three insertions (for each insertion, one primer was placed inside the insertion, with the other being placed in the gene itself) were then used to PCR amplify and screen a diverse panel of 108 WT cultivated sunflower lines [51] for presence or absence of each insertion. DNA extraction was carried out as above and PCR carried out as previously described [50]. Amplicons were visualized via agarose gel electrophoresis and staining with ethidium bromide. Presence or absence of a PCR product indicated presence/absence of an insertion. A positive control PCR was carried out for each individual using another pair of primers specific to HaCYC2c to protect against the possibility of false negatives.
Expression Analyses
Gene expression analyses were carried out using quantitative reverse-transcriptase (qRT)-PCR on RNA isolated from WT (NMS373), dbl, tub, Ames3288 and Chrysanthemoides individuals. Briefly, petal tissue was collected for each genotype/floret type combination at a stage when the sunflower head was fully open. Tissues were collected from four different floret types, namely: ray florets, outer discs, intermediate discs (halfway between the outer and central florets), and central disc florets. RNA was extracted using the guanidium isothiocyanate method followed by isolation using the RNeasy Mini Kit (Qiagen, Valencia, CA) using previously established protocols [22]. Synthesis of cDNA was performed using 500 ng of total RNA after removal of genomic DNA using Qiagen on-column DNase treatment. ImProm II reverse transcriptase (Promega Corporation, WI, USA) and oligo dT (15) primers were used to perform reverse transcription in a 30 µl reaction volume. The cDNA was diluted with 80 µl of water for all gene expression analyses. All the qRT-PCR analyses were performed using an Eppendorf realplex2 real-time PCR system (Eppendorf, Hauppauge, NY) with primers in Table S2. The reaction conditions were as follows: 50°C for 2 min; 95°C for 10 min; 40 cycles of 95°C (15 s), 59°C (20 s) and 68°C (30 s). Melt-curve analyses were performed after the PCR. A single distinct peak was observed for both the target (HaCYC2b, 2c, and 2e) and control (actin) genes indicating the specific amplification of a single product. Relative expression (scaled against WT ray florets) was calculated using the Pfaffl method [52].
Gene Discovery and Phylogenetic Analysis
Degenerate PCR primers were employed as previously described [22] to amplify the central portion (i.e., the region between the TCP and R domains) of the CYC-like genes from other members of the Asteraceae (Berkheya purpurea [subfamily Cichorioideae], Callistephus chinensis [Asteroideae], Dasyphyllum diacanthoides [Barnadesioideae], Gazania hybrida [Cichorioideae], Gerbera hybrida cultivar [Mutisioideae], Senecio squalidus [Asteroideae]) as well as Acicarpha tribuloides (Calyceraceae; outgroup). Seeds of most of these species were obtained from Chileflora (www.chileflora.com), Chiltern Seeds (www.chilternseeds.co.uk), or the USDA (see above). The exceptions were Senecio (seed from Richard Abbott, University of St. Andrews) and Acicarpha (leaf material from Leigh Johnson, Brigham Young University). PCR, cloning, and sequencing were carried out as before [50]. These species were chosen to represent a broad cross-section of the family. However, we focused on radiate species where possible. The exceptions are Dasyphyllum, with an inflorescence made up of actinomorphic disc and zygomorphic bilabiate florets, and Acicarpha, which approximates an inflorescence with only actinomorphic flowers.
The CYC-like gene family in the Asteraceae is made up of three subfamilies, and the focus here was on CYC2-like genes. Therefore, CYC clones from the above reactions that showed similarity to CYC2 genes from Helianthus were further characterized via genome walking (as described previously [22]) to obtain the entire TCP and R domains. A nucleotide alignment was next obtained for the conserved TCP and R domains as well as the intervening region using ClustalW2 [53] followed by manual adjustment such that indels were in multiples of three. Maximum likelihood analysis was then carried out using PhyML [54] with 100 bootstrap replicates and Mr. Bayes [55] to produce gene trees.
Accession Numbers
Sequence data have been deposited into the Genbank DNA database (www.ncbi.nlm.nih.gov/genbank) under accession numbers HQ891026–HQ891029 and JF489909–JF489913 (HaCYC2c), JF489906–JF489908, JQ594983 (HaCYC2b), JF489914–JF489916, JQ594982 (HaCYC2e), and JF299240–JF299257 (Asteraceae CYC2 sequences).
Supporting Information
Zdroje
1. DonoghueMJReeRHBaumDA 1998 Phylogeny and the evolution of flower symmetry in the Asteridae. Trends Plant Sci 3 311 317
2. EndressPK 2001 Evolution of floral symmetry. Curr Opin Plant Biol 4 86 91
3. PrestonJCHilemanLC 2009 Developmental genetics of floral symmetry evolution. Trends Plant Sci 14 147 154
4. SargentRD 2004 Floral symmetry affects speciation rates in angiosperms. Proc R Soc Lond B Biol Sci 271 603 608
5. TakhtajanA 1969 Flowering Plants: Origin and Dispersal. Oliver and Boyd, Edinburgh
6. DilcherD 2000 Toward a new synthesis: Major evolutionary trends in the angiosperm fossil record. Proc Nat Acad Sci USA 97 7030 7036
7. WesterkampCClaßen-BockhoffR 2007 Bilabiate flowers: The ultimate response to bees? Ann Bot 100 361 374
8. CubasP 2004 Floral zygomorphy, the recurring evolution of a successful trait. Bioessays 26 1175 1184
9. FunkVABayerRJKeeleySChanRWatsonL 2005 Everywhere but Antarctica: Using a supertree to understand the diversity and distribution of the Compositae. Biol Skr 55 343 373
10. StevensPF 2001 onwards Angiosperm Phylogeny Website. Version 8, June 2007. http://www.mobot.org/MOBOT/research/APweb/
11. PaneroJLFunkVA 2008 The value of sampling anomalous taxa in phylogenetic studies: Major clades of the Asteraceae revealed. Mol Phylogenet Evol 47 757 782
12. FunkVASusannaAStuessyTFBayerRJ 2009 Systematics, Evolution, and Biogeography of Compositae. International Association for Plant Taxonomy (Vienna)
13. AnderssonS 2008 Pollinator and nonpollinator selection on ray morphology in Leucanthemum vulgare (oxeye daisy, Asteraceae). Am J Bot 95 1072 1078
14. NielsenLRPhilippMSiegismundHR 2002 Selective advantage of ray florets in Scalesia affinis and S. pedunculata (Asteraceae), two endemic species from the Galapagos. Evol Ecol 16 139 153
15. StuessyTFSpoonerDMEvansKA 1986 Adaptive significance of ray corollas in Helianthus grosseserratus (Compositae). Am Midl Nat 115 191 197
16. FengXZZhaoZTianZXuSLuoY 2006 Control of petal shape and floral zygomorphy in Lotus japonicus. Proc Nat Acad Sci USA 103 4970 4975
17. LuoDCarpenterRVincentCCopseyLCoenE 1996 Origin of floral asymmetry in Antirrhinum. Nature 383 794 799
18. CorleySBCarpenterRCopseyLCoenE 2005 Floral asymmetry involves an interplay between TCP and MYB transcription factors in Antirrhinum. Proc Nat Acad Sci USA 102 5068 50734
19. BroholmSKTähtiharjuSLaitinenRAEAlbertVATeeriTH 2008 A TCP domain transcription factor controls flower type specification along the radial axis of the Gerbera (Asteraceae) inflorescence. Proc Nat Acad Sci USA 105 9117 9122
20. KimMCuiM-LCubasPGilliesALeeK 2008 Regulatory genes control a key morphological and ecological trait transferred between species. Science 322 1116 1119
21. CarlsonSEHowarthDGDonoghueMJ 2011 Diversification of CYCLOIDEA-like genes in Dipsacaceae (Dipsacales): implications for the evolution of capitulum inflorescences. BMC Evolutionary Biology, 11 325
22. ChapmanMALeebens-MackJBurkeJM 2008 Positive selection and expression divergence following gene duplication in the sunflower CYCLOIDEA gene family. Mol Biol Evol 25 1260 1273
23. CockerellTDA 1915 Specific and varietal characters in annual sunflowers. Am Nat 49 609 622
24. FambriniMBertiniDPugliesiC 2003 The genetic basis of a mutation that alters the floral symmetry in sunflower. Ann Appl Biol 143 341 347
25. FambriniMMichelottiVPugliesiC 2007 The unstable tubular ray flower allele of sunflower: inheritance of the reversion to wild-type. Plant Breeding 126 548 550
26. FickGN 1976 Genetics of floral color and morphology in sunflowers. J Hered 67 227 230
27. WitteC-PLeQHBureauTKiumarA 2001 Terminal-repeat retrotransposons in miniature (TRIM) are involved in restructuring plant genomes. Proc Nat Acad Sci USA 98 13778 13783
28. NealPRDafniAGiurfaM 1998 Floral symmetry and its role in plant-pollinator systems: terminology, distribution, and hypotheses. Annu Rev Ecol Syst 29 345 373
29. BartlettMESpechtCD 2011 Changes in expression pattern of the teosinte branched1-like genes in the Zingiberales provide a mechanism for evolutionary shifts in symmetry across the order. Am J Bot 98 227 243
30. CiterneHLPenningtonRTCronkQCB 2006 An apparent reversal in floral symmetry in the legume Cadia is a homeotic transformation. Proc Nat Acad Sci USA 103 12017 12020
31. HilemanLCKramerEMBaumDA 2003 Differential regulation of symmetry genes and the evolution of floral morphologies. Proc Nat Acad Sci USA 100 12814 12819
32. ZhangWKramerEMDavisCC 2010 Floral symmetry genes and the origin and maintenance of zygomorphy in a plant-pollinator mutualism. Proc Nat Acad Sci USA 100 6388 6393
33. ZhouXRWangYZSmithJFChenRJ 2008 Altered expression patterns of TCP and MYB genes relating to the floral developmental transition from initial zygomorphy to actinomorphy in Bournea (Gesneriaceae). New Phytol 178 532 543
34. HowarthDGDonoghueMJ 2005 Duplications in CYC-like genes from Dipsacales correlate with floral form. Int J Plant Sci 166 357 370
35. PrestonJCMartinezCCHilemanLC 2011 Gradual disintegration of the floral symmetry gene network is implicated in the evolution of a wind-pollination syndrome. Proc Nat Acad Sci USA 108 2343 2348
36. HughesTA 2006 Regulation of gene expression by alternative untranslated regions. Trends Genet 22 119 122
37. PesoleGMignoneFGissiCGrilloGLicciulliF 2001 Structural and functional features of eukaryotic mRNA untranslated regions. Gene 276 73 81
38. BarkerMSKaneNCMatvienkoMKozikAMichelmoreRW 2008 Multiple paleopolyploidizations during the evolution of the Compositae reveal parallel patterns of duplicate gene retention after millions of years. Mol Biol Evol 25 2445 2555
39. TankDCDonoghueMJ 2010 Phylogeny and phylogenetic nomenclature of the Campanulidae based on an expanded sample of genes and taxa. Syst Bot 35 425 441
40. WoodTBurkeJMRiesebergLH 2005 Parallel genotypic adaptation: when evolution repeats itself. Genetica 123 157 170
41. ColosimoPFHosemannKEBalabhadraSVillarealGDicksonM 2005 Widespread parallel evolution in sticklebacks by repeated fixation ofectodysplasin alleles. Science 307 1928 1933
42. McCrackenKGBargerCPBulgarellaMJohnsonKPSonsthagenSA 2009 Parallel evolution in the major haemoglobin genes of eight species of Andean waterfowl. Mol Ecol 18 3992 4005
43. GanforninaMDSanchezD 1999 Generation of evolutionary novelty by functional shift. Bioessays 21 432 439
44. TrueJRCarrollSB 2002 Gene co-option in physiological and morphological evolution. Annu Rev Cell Dev Biol 18 53 80
45. ChenLBDeVriesALChengCHC 1997 Convergent evolution of antifreeze glycoproteins in Antarctic notothenioid fish and Arctic cod. Proc Nat Acad Sci USA 94 3817 3822
46. SutherlandTDYoungJHWeismanSHayashiCYMerrittDJ 2010 Insect Silk: One Name, Many Materials. Annu Rev Entomol 55 171 188
47. HoffmannFGOpazoJCStorzJF 2010 Gene cooption and convergent evolution of oxygen transport hemoglobins in jawed and jawless vertebrates. Proc Nat Acad Sci USA 107 14274
48. RosinFMKramerEM 2010 Old dogs, new tricks: Regulatory evolution in conserved genetic modules leads to novel morphologies in plants. Dev Biol 332 25 35
49. TangSKishoreVKKnappSJ 2003 PCR-multiplexes for a genome-wide framework of simple sequence repeat marker loci in cultivated sunflower. Theor Appl Genet 107 6 19
50. ChapmanMAPashleyCHWenzlerJHvalaJTangS 2008 A genomic scan for selection reveals candidates for genes involved in the evolution of cultivated sunflower (Helianthus annuus). Plant Cell 20 2931 2945
51. MandelJDechaineJMarekLBurkeJ 2011 Genetic diversity and population structure in cultivated sunflower and a comparison to its wild progenitor, Helianthus annuus L. Theor Appl Genet 123 693 704
52. PfafflMW 2001 A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 29 2002 2007
53. ThompsonJDHigginsDGGibsonTJ 1994 Clustal W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22 4673 4680
54. GuindonSGascuelO 2003 A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol 52 696 704
55. RonquistFHuelsenbeckJP 2003 MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19 1572 1574
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2012 Číslo 3
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- PIF4–Mediated Activation of Expression Integrates Temperature into the Auxin Pathway in Regulating Hypocotyl Growth
- Metabolic Profiling of a Mapping Population Exposes New Insights in the Regulation of Seed Metabolism and Seed, Fruit, and Plant Relations
- A Splice Site Variant in the Bovine Gene Compromises Growth and Regulation of the Inflammatory Response
- Comprehensive Research Synopsis and Systematic Meta-Analyses in Parkinson's Disease Genetics: The PDGene Database
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy