
Evoluce a evoluční teorie pro lékaře
VI. Evo-devo
(evoluční vývojová biologie)
Evolution and evolutionary theory for physicians. VI. Evo-devo (evolutionary developmental biology)
Evolutionary developmental biology, informally evo-devo, is a biological discipline that compares developmental processes of different life forms. It deals with origin and evolution of the embryonic development, developmental plasticity in evolution, how ecology impacts the development and evolutionary change and basis of homoplasy and homology. Evo-devo completes and changes neodarwinian synthesis due to a new view on genetics of evolutionary adaptation. Basic research of evo-devo produces fundamental pieces of knowledge and future therapy of complex diseases eg. Alzheimer’s disease and malignant tumours.
Key words:
Evo-devo, modularity, evolutionary adaptation.
Autoři:
F. Koukolík
Působiště autorů:
Primář: MUDr. František Koukolík DrSc
; Národní referenční laboratoř prionových chorob
; Fakultní Thomayerova nemocnice s poliklinikou, Praha
; Oddělení patologie a molekulární medicíny
Vyšlo v časopise:
Prakt. Lék. 2010; 90(6): 327-332
Kategorie:
Editorial
Souhrn
Evoluční vývojová biologie, neformálně evo-devo, je biologická disciplina porovnávající vývojové procesy u různých forem života. Zabývá se původem a evolucí embryonálního vývoje, vývojovou plasticitou, ekologickým ovlivněním vývoje a evolučních změn, jakož i podkladem homologie a homoplazie. Evo-devo doplňuje a mění neodarwinovskou syntézu novým pohledem na genetiku evoluční adaptace. Základní výzkum evo-devo přináší klíčové poznatky pro pochopení a budoucí léčení komplexních onemocnění, například Alzheimerovy nemoci a maligních nádorů.
Klíčová slova:
Evo-devo, modularita, evoluční adaptace.
Úvod
Jestliže genomika změnila moderní syntézu evoluční teorie (13) v řadě zásadních směrů (přehled 9), mění ji evoluční vývojová biologie (evolutionary developmental biology, evolution of development, neformálně, nicméně velmi rozšířeně evo-devo) pravděpodobně stejně výrazně. Je dobré mít na mysli, že jde o soubor změn, nikoli změnu jedinou a že jde o svým způsobem bouřlivý proces, nikoli kanonizovaný stav. Protichůdná tvrzení a spory jsou proto samozřejmou součástí, jako je tomu v průběhu vývoje jakéhokoli vědeckého oboru. V textu se vyskytují biologické pojmy s nimiž lékaři běžně nepřicházejí do styku. Součástí je proto jejich stručný abecední slovník (Slovník na str. 331).
Charakteristika evo-devo
Evo-devo je součást biologie porovnávající vývojové procesy různých živočichů a rostlin ve snaze určit jak mechanismus vývoje, tak jejich vzájemné vývojové vztahy. Klíčovými pojmy a vývojovými směry poznávání evo-devo jsou:
1. modularita.
Živočišná i rostlinná těla a metabolické cykly jsou uspořádány modulárně, jednotlivé části se opakují, příkladem jsou prsty nebo žebra. Evo-devo hledá genetické a evoluční základy rozčlenění embryí do jednotlivých modulů a důvody následného částečně nezávislého vývoje modulů. Pojem modulus je zde užíván obecně ve smyslu jednotky obsahující nějaké podjednotky. Ke studiu embryií byl vyvinut standardní systém (15).
2. Některé genové produkty fungujíjako přepínače, jiné jsou difundující signály.
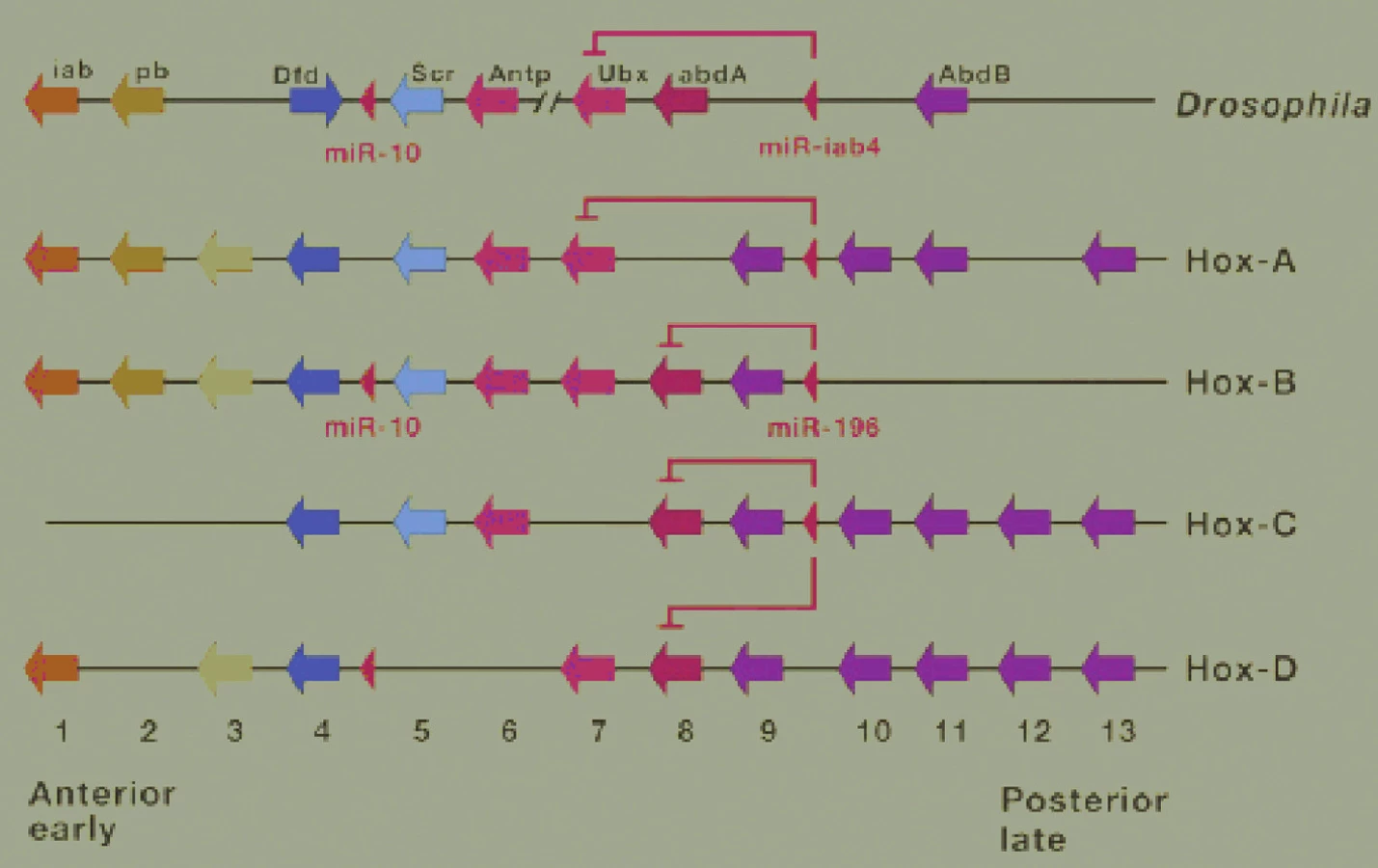
Geny kódují strukturální proteiny, které tvoří například cytoskelet, stejně jako funkční proteiny, enzymy, řídící biochemické procesy. Dlouhou dobu převládal názor, podle kterého byly organismy reflexí svých genů. Podle tohoto názoru byly proměny existujících a vznik nových metabolických drah důsledkem mutací. V r. 1961 byl u bakterie Escherichia coli objeven gen fungující na základě „zapnutí“ (nebo „přepnutí“) signálem z prostředí (12). Následně byly u živočichů objeveny Hox geny (obr. 1, 2), které jsou přepínače funkce jiných genů a jejichž činnost je ovlivňována morfogeny činnými podobně jako zevní signály ovlivňující činnost genu Escherichií.

To byl velký objev:
Dokazoval, že činnost genů je možné výběrově zapnout nebo vypnout, takže geny nejsou činné stále. Nadto se zjistilo, že tak odlišné organismy jako jsou octomilky a lidé užívají v průběhu embryogeneze stejné geny – ale odlišně je regulují.
3. Podoby organismů mohou být ovlivněny mutacemi v promotorových oblastech genů.
To jsou úseky DNA, na něž se váží produkty některých genů a kontrolují činnost stejných nebo jiných genů. Zásadním evolučním objevem tedy je, že rozdíly mezi jednotlivými druhy, řády nebo kmeny života jsou podmíněny rozdíly v časové a prostorové expresi vývojově konzervovaných genů. Velké evoluční změny jsou tedy více důsledkem změn regulace činnosti konzervovaných genů, než by byly dány evolucí nových genů.
4. Fenotypy organismů nemusí být plně určovány svými genotypy, podílí se vlivy prostředí. Předmětem výzkumu evo-devo je proto fenotypická plasticita.
Evo-devo a evoluční syntéza
Vztah evo-devo a evoluční teorie shrnuli v nedávné době Carroll (4), Mueller (11) a De Robertis (6).
Carrollův výklad (4), vysvětlující které geny a jaký typ změn jejich sekvencí odpovídají za vývoj morfologické rozmanitosti říká v osmi tézích, že se morfologicky odlišné podoby života vyvíjejí v důsledku změn exprese konzervovaných proteinů a že tyto změny jsou z valné části výsledkem mutací cis-regulačních sekvencí pleiotropních vývojových regulačních locusů a cílových genů uvnitř rozsáhlých sítí, které kontrolují.
Osm Carrollových principů zní:
- Většina proteinů
regulujících vývoj se podílí na mnohotných nezávislých
vývojových procesech a na
formování a uspořádání
morfologicky odlišných tělesných struktur. Stačí si uvědomit
odlišnost morfologie a ekologie
larvální a dospělé
podoby octomilky Drosophila melanogaster:
je větší, než je odlišnost dospělé podoby jakéhokoli jiného
páru sesterských druhů. Přitom jsou stovky genových produktů
utvářejících larvální a dospělou
podobu co do sekvencí identické.
To samé platí pro většinu signálních proteinů a transkripčních faktorů, součástí takzvané „genetické skřínky na nářadí“ (genetic toolkit, dále jen „skřínka“) řídících vývoj daného živočicha. Kupříkladu protein označovaný Sonic hedgehog řídí u kuřat kromě mnoha jiného jak vývoj mozečku, tak vývoj počtu prstů (10). Proteiny (stejně jako geny) ovlivňující větší počet „cílů“ se označují jako pleiotropní. Vliv většiny proteinů řídících vývoj je mozaikově pleiotropní, čímž se označuje jejich působení nezávisle na druzích buněk, zárodečných vrstvách, orgánech nebo vývojových stádiích.
To má evoluční důsledky: jestliže stejný protein ovlivňuje vývoj odlišných částí těla, lze soudit, že může ovlivňovat rozdíly stejných částí těla různých druhů. Mutace, které by v konečném důsledku jejich činnost poškozovaly, jsou tedy evolučně omezené. Jestliže stejný gen ovlivňuje tak rozdílné znaky, pak se v průběhu evoluce mohou funkce genů měnit, aniž by se geny zdvojovaly. - Morfologicky odlišné a vývojově vzdálené taxony sdílejí podobné „genetické nástrojové skřínky“ a geny utvářející těla. Důkazem je existence Hox genů a jejich hnízd u většiny živočišných kmenů (obr. 1, 2). Předpokládá se, že nositelem poměrně celistvé „skřínky“ byl poslední společný předek bilateralií, žijící v prekambrijském údobí.
- Řada živočišných proteinů odpovídajících „skřínce“ je často in vivo funkčně ekvivalentních, jedny tedy mohou nahradit druhé bez ohledu na více než jednu miliardu let nezávislého vývoje v odlišných živočišných liniích. Z toho plyne, že se biochemické vlastnosti těchto proteinů a jejich interakce například s receptory nebo kofaktory měnily v průběhu velmi dlouhé doby jen málo. Známým příkladem je schopnost myšího Pax-6 proteinu, jenž se u myší podílí na vývoji oka, indukovat vývoj ommatidií, což jsou jednotky složeného oka u octomilek.
- Vzájemně podobné skupiny genů a hluboce konzervované genetické regulace odpovídají za vznik a vývoj například očí, končetin nebo srdcí různých živočichů. Tyto orgány jsou u živočichů vzájemně odlišných kmenů natolik odlišné, že dlouho převládala představa jejich vzájemně nezávislého vývoje.
- V porovnání se zdvojováním jiných genových rodin je zdvojování některých základních genových rodin „skřínky“ v průběhu vývoje a rozčleňování živočichů vzácné. Z toho plyne, že zdvojování genů není nutná součást morfologických inovací. Naopak: selekce může jít proti jejich zdvojování.
- Morfologická rozmanitost souvisí se změnami prostorové regulace „skříňkových“ genů a genů, které jsou jimi regulovány.
- Typickým znakem pleitropických „skřínkových“ lokusů jsou velké, složité a modulární cis-regulační oblasti. Například Eyless je lokus klíčový nejen pro vývoj oka, ale i pro správný vývoj centrálního nervového systému. Sekvence vyžadované pro jeho expresi zabírají nejméně 7 kilobází nekódující DNA ve vztahu ke genu 5’a 3’ a uvnitř jednoho intronu. Existuje nejméně šest cis-regulačních elementů řídících expresi Eyless v různém prostorovém uspořádání (1) – například v oku a v různých lalocích vyvíjejícího se embryonálního, larválního a dospělého mozku.
- jednotlivé regulační proteiny kontrolují mnoho desítek až stovek cis-regulačních elementů (CRE) cílových genů
Carroll (2, 3, 4) z pokladů těchto osmi tézí vyvozuje cis-regulační hypotézu:
Mutace cis-regulačních elementů existujících genů považuje za častější a průběžný zdroj morfologických variací, než je evoluce genů nových. Má za to, že změny sekvencí CRE jsou pro evoluční divergenci znaků a genových regulací mezi populacemi, druhy a taxony dostačující. Evoluce CRE je nutná pro změnu zapojení (rewiring) regulačních sítí. Zdvojování genů a změny kódování podle jeho názoru ke změně zapojení nedostačují. Variace a divergence CRE je zjistitelná v daleko kratších časových údobích a taxonomických vzdálenostech než jsou funkční rozdíly v transkripčních faktorech a nová zdvojení genů ve „skřínce“.
Vývoj CRE se může dít:
- kooptováním vstupů nových transkripčních faktorů mutacemi existujících CRE,
- kooptováním mobilních (transposable) elementů, coby nových CRE,
- ztrátou transkripčních vstupů v existujících CRE,
- remodelací CRE.
Podle Carrollova souhrnu (4) se tedy morfologické formy vyvíjejí díky alteraci funkčně konzervovaných proteinů, přičemž jsou tyto změny hlavně podmíněny mutacemi cis-regulačních oblastí mozaikově pleiotropních vývojových regulačních genů a cílových genů ve velkých regulačních sítích, které jsou jimi kontrolovány.
S Carrollovým názorem, jenž je svým způsobem v evoluční teorii revolucí a je souhrnem jeho dosavadní mnohaleté práce, polemizují Hoekstra a Coyne (7). Podle jejich názoru není Carrollova domněnka tvrdící, že se morfologické adaptace vyvíjejí genetickými mechanismy, které by se odlišovaly od mechanismů týkající se adaptací funkčních a dalších znaků, dobře podložena teoreticky, ani empiricky. Některé podoby evoluce proteinů se mohou vyhnout záporným důsledkům pleiotropie, tedy ovlivnění většího počtu „cílů“ současně, právě genovou duplikací.
Podle těchto kritiků dokládají genomické studie adaptace v oblastech kódujících proteiny včetně transkripčních faktorů, jen málo studií zkoumalo regulační oblasti. Studie cílené na jednotlivé lokusy vypovídají spíše o ztrátě znaků než o jejich zisku, žádná nedokládá vyvinutou regulační oblast. Podle Hoekstra a Coyne (7) je cis-regulační teorie nezralá, adaptace a speciace jsou podmíněny kombinacemi cis-regulačních a strukturálních mutací, podstatný evoluční podíl mají mít strukturální mutace.
Mueller (11) ve filozoficky laděném přehledu popisuje rozdíl nedarwinovské syntézy a evo-devo pojmy evolvabilita, emergence a inherence.
Evolvabilita
je pojem, jenž by se dal převést do češtiny například souslovím „soubor vývojových možností“. Je definována jako intrinsický, niterný potenciál dané linie vytvářet fenotypické variace. Evolvabilitu je možné v současnosti zkoumat jako vývojovou plasticitu orgánových systémů a jejich odpovědi na proměny prostředí. Příkladem jsou studie rostlin, motýlů nebo brouků (přehled 11).
Emergence,
doslovně objevování se, doplňuje nedarwinovskou teorii adaptace zavedením nedeterministických vývojových a environmentálních faktorů, odpovídajících za původ evolučních inovací.
Zcela filosofickým pojmeminherencemá na mysli niterně dané, bytostné, neodmyslitelné vlastnosti vyvíjejícího se systému:
Genotypická evoluce není dána jen genovou regulační evolucí. Specifické vlastnosti tkání a biologických forem vznikající buněčnými interakcemi nejsou deterministicky zakódovány v genomu.
Živočišné kmeny a kambrijská exploze života
De Robertis (6) považuje za klíčovou otázku, kterou musí evo-devo vyřešit, stavbu a funkci nejstarších společných předků bilaterálních živočichů, Urbilateralií (pra-bilaterálií). Má zato, že tato hypotetická, předprvohorní podoba živočišného života byla již značně složitá.Vlastnila většinu základních vývojových genetických drah.
Dalším základním problémem je vysvětlení, jak se genetické sítě přítomné v tomto archetypálním prapředkovi proměňovaly do dnešní rozmanitosti života. V současnosti totiž existuje asi 35 živočišných kmenů s rozdílnými plány těl (14). Téměř 30 z nich jsou bilateralia, tradičně dělená na protostomia a deuterostomia (prvoústé a druhoústé).
U protostomií (dále dělených na dvě skupiny, viz níže) se vyvíjí ústní otvor v blízkosti blastopory, to je otvor jímž komunikuje gastrula, rané embryonální vývojové stadium, se vnějškem; kromě toho mají ventrální nervovou trubici a dorzální „mozkové“ ganglion.
U deuterostomií se blastopora vyvíjí v blízkosti anu; kromě toho mají dorzální centrální nervový systém. Fosilie dospělých bilateralií se objevují v průběhu takzvané kambrijské exploze života (535–525 Ma před současností), předpokládá se však, že bilateralia jsou vývojově starší. Všechny plány těl 35 současných živočišných kmenů současnosti byly přítomny před 525 Ma.
Otázkou je, proč se od té doby neobjevily nové živočišné kmeny (5). Zajímavá ekologická odpověď na tuto otázku je hypotéza Země – sněhové koule:
V rozmezí doby 750–550 Ma před současností prošla Země několika masivními zaledněními. Předpokládá se možnost částečného nebo úplného promrznutí oceánů (8). Živočišné populace přežívaly v oblastech mělké vody snad ohřívané vulkanickou hydrothermální aktivitou. Chlad znamenal pro řadu populací geneticky „zúžené hrdlo lahve“ – přežívaly jen malé, geneticky relativně homogenní, efektivní, to znamená rozmnožování schopné, populace. Jakmile došlo k oteplení, rozšířili se živočiši, kteří tento selekční tlak přežili, opět do oceánů. S sebou nesli coby adaptaci tělesný plán. Efektivní způsob rychlé adaptace je ztráta genů, což mohlo být podkladem vzniku současných živočišných kmenů.
Jestliže se najde u lidí, nebo u kteréhokoli jiného strunatce gen, který se najde u ecdysozoa nebo lophotrochozoa, to jsou dvě skupiny prvoústých (příkladem prvních jsou členovci, příkladem druhých měkkýši), pak lze předpokládat, že ho urbilateralia měla také.
Lze soudit, že některé z těchto genů některé živočišné kmeny v průběhu vývoje ztratily, což umožnilo rychlou evoluční proměnu (6).


Genetika evoluční adaptace
Jestliže je „skřínka“ tak vývojově konzervovaná, jak je možná taková druhová rozmanitost?
V současnosti ji vysvětlují čtyři typy adaptivních mutací (přehled 7):
Svrchu podrobněji probrané cis-regulační mutace, o nichž se předpokládá, že právě jejich variace jsou podkladem většiny novinek v uspořádání živočišných těl.
Strukturální mutace. Klasickým příkladem je změna jediné aminokyseliny v globinu, která zvyšuje afinitu hemoglobinu pro kyslík u hus migrujících ve velkých výškách (Perutz, cit. 7).
Zdvojení genů. Duplikáty mohou být užity pro novou funkci aniž by původní ztratily.
Genové delece, což jsou velmi efektivní způsoby rychlé adaptace na novou ekologickou niku (15). Příkladem jsou jeskynní živočiši, kteří se stali albíny a ztratili oči.
Závěr
Diagnostická i terapeutická medicína současnosti začíná být a v krátké době bude ve velkém rozsahu molekulární medicínou, založenou na genomice a proteomice. Základní výzkum evo-devo přináší v současnosti poznatky, které lékařům často připadají příliš biologické a obecné s malým dopadem na každodennost. V důsledcích by to byl neprozíravý postoj, protože každý, kdo jen zcela rámcově sleduje odbornou literaturu vidí, jak se doslova před očima mění pochopení tak rozšířených chorob, jako jsou například Alzheimerova nemoc nebo zhoubné nádory právě díky základnímu biologickému výzkumu prováděnému často na modelech, které připadají lékařům vzdálené.
Změna medicínského myšlení probíhá zlomově. Stačí se jen krátce podívat do fascinující databáze KEGG (Koyto Encyclopedia of Genes and Genomes; http://www.genome.jp/kegg/).
MUDr. František Koukolík, DrSc.
Oddělení
patologie a molekulární
medicíny
Národní
referenční laboratoř prionových
chorob
Fakultní Thomayerova nemocnice s poliklinikou
Vídeňská
800
140
59 Praha 4 Krč
E-mail:
frantisek.koukolik@ftn.cz
SLOVNÍK
Analogie viz homologie
Cis-regulace
V molekulární biologii pojem označuje „působení z totožné molekuly“, to znamená intramolekulární. Cis-regulační element je úsek DNA nebo RNA řídící expresi genů umístěných ve stejné molekule DNA. Latinský pojem cis znamená „na stejné straně“. Protikladem je trans-regulace označující působení z odlišné molekuly, to znamená intermolekulární. Trans-regulační elementy jsou difundující faktory, obvykle proteiny, které mohou ovlivňovat expresi vzdálených genů. Například transkripční faktor regulující činnost genu na chromozomu 6 může být sám přepisován z genu na chromozomu 11
„Genetická skřínka na nářadí“ (genetic toolkit, developmental-genetic toolkit)
„Genetickou skřínku na nářadí“ tvoří geny, jejichž produkty kontrolují vývoj mnohobuněčných živočichů. Rozdíly v „užití nástrojů“ ovlivňují plán těla, počet, identitu a uspořádaní částí těla. „Skřínka“ je vývojově vysoce konzervovaná – její obsah je u nejrůznějších živočišných kmenů velmi podobný. Většina genů skřínky jsou součásti signálních drah, kódují transkripční faktory, proteiny buněčné adheze, povrchové buněčné receptory a morfogeny. Funkce genů je ve vysoké korelaci s jejich časovou a prostorovou expresí. Jeden z nejdůležitějších nástrojů skřínky je hnízdo, respektive komplex Hox genů (Hox gene cluster).
Fenotypická plasticita
Fenotypická plasticita je schopnost organismu měnit svůj fenotyp úměrně proměnám prostředí
Homeobox
Homeobox je sekvence DNA, součást genů regulujících utváření, neboli morfogenezu živočichů, hub i rostlin. Zjednodušeně lze říci, že homeobox geny jsou architekti, respektive konstruktéři plánu těl. Homeobox je tvořen přibližně 180 páry bází („písmen“) DNA. Homeobox geny kódují transkripční faktory jejichž činnost se dá přirovnat k přepínačům spouštějícím činnost dalších genů.
U lidí jsou homeobox geny ve čtyřech hnízdech: součástí chromozomu 7 je HOXA. Součástí chromozomu 17 jsou HOXB. Součástí chromozomu 12 jsou HOXC, součástí chromozomu 2 jsou HOXD. Názvy mají synonyma (obr 1, 2).
Mutace homeobox genů jsou příčinou nápadných fenotypických změn, příkladem jsou nohy rostoucí na hlavě místo tykadel, což je důsledek defektu jediného genu (mutace se jmenuje Antennapedia).
Homologie
V evoluční biologii označuje pojem homologie jakoukoli podobnost mezi znaky organismu, které vznikly na základě sdíleného předka. Příkladem evoluční homologie jsou přední končetiny savců. Příkladem vývojovéhomologie jsou varlata a ovaria: obě struktury vznikají v průběhu embryonálního vývoje ze stejné tkáně.
V genetice označuje pojem homologie podobnost sekvencí (pořadí „písmen“, neboli bází DNA), ty obvykle bývají podobné, nikoli totožné. Podobné mohou být sekvence DNA v jedné buňce stejně jako sekvence DNA zjišťované u různých organismů. Předpokládá se, že úseky DNA s podobnými sekvencemi sdílejí společného předka. Homologní chromozomy mají stejné geny a podobné sekvence. Páry homologních chromozomů se vzájemně řadí v průběhu meiosy, neboli redukčního dělení, jímž vznikají pohlavní buňky.
Homologie je opakem analogie popisující dvě struktury vykonávající stejné funkce podobnými mechanismy, jejichž vznik byl vzájemně odlišný (žert říká, že homologické jsou lidské ruce a koňská kopyta, analogická jsou křídla čmeláka a letadla). Podobné struktury tedy mohou vznikat zcela odlišnými způsoby, tomu se říká konvergentní evoluce, nebo mohou být homologické. Lidské ruce a netopýří křídla lze považovat jak za homologické, tak za analogické: přední končetiny společného předka ptáků a netopýrů se vyvinuly do orgánů s podobnou funkcí nezávisle.
Homologní chromozomy
Homologní chromozomy jsou neidentické chromozomy vytvářející páry v průběhu meiosy. S výjimkou pohlavních chromozomů jsou jejich sekvence v celé délce podobné, obvykle obsahují stejné genové sekvence. Variace genů homologních chromozomů vytváří jejich odlišné podoby – alely.
Homoplasie
Podobnost ve znacích druhů, které nemají společného předka. Homoplasie je výsledkem konvergentní evoluce (viz homologie). Příkladem konvergentní evoluce je oko hlavonožců, savců a žahavců.
Konzervace
V obecné biologii se za konzervované sekvence považují identické nebo podobné sekvence DNA, RNA, proteinové, včetně jejich struktury stejně jako sekvence karbohydrátových polymerů, a to mezi jednotlivými druhy (to jsou ortologní sekvence) a/nebo mezi různými molekulami vznikajícími uvnitř téhož organismu (to jsou paralogní sekvence). Čím je sekvence ve fylogenetickém kmeni hlouběji (blíž k poslednímu univerzálnímu společnému předkovi), tím více je konzervovaná. Mutace ve vysoce konzervované oblasti vede k neživotným formám nebo formám vyloučeným přírodním výběrem. Podobnost mezi molekulami se vyjadřuje v procentech identity nebo podobnosti. Nemá se užívat pojem procenta homologie. Podobnost zejména krátkých sekvencí však může vzniknout náhodně. Sekvence mohou být podobné také proto, že byly selektovány k vazbě na transkripční faktor.
Lokus
Lokus je počeštěné anglické, původně latinské slovo locus (pl. loci) označující v genetice umístění genu nebo genetické sekvence v chromozomu. Pojem lokus se často užívá pro označení chromozomální oblasti v níž se předpokládá existence hledaného genu.
Modularita
Mnohé organismy jsou uspořádány modulárně, a to jak anatomicky, tak metabolicky. Anatomické moduly jsou tvořeny segmenty nebo orgány. Příkladem metabolického modulu je citrátový cyklus.
Morfogen
Látka, která kontroluje způsob vývoje tkání a umístění různých druhů specializovaných buněk ve tkáni. Šíří se z lokálního zdroje a ve vyvíjející se tkáni vytváří koncentrační gradient. Ve vývojové biologii je morfogen označení signální molekuly ovlivňující přímo buňky: výsledkem je typ buněčné odpovědi závislé na koncentraci morfogenu. Příkladem morfogenu je epidermální růstový faktor.
Ortologie
Homologní sekvence jsou ortologní, jestliže je oddělila speciace, vznik nových druhů. Jestliže se jeden druh rozštěpil na dva odlišné druhy, pak se divergentní kopie téhož genu u obou odlišných druhů nazývají ortologní. Jinak řečeno: ortologní geny různých druhů pocházejí ze společného předka.
Paralogie
Homologní sekvence jsou paralogické, jestliže je oddělila genová duplikace: jestliže se gen zdvojil a zaujímá ve stejném genomu dvě místa, jsou obě kopie paralogické. Příkladem vývojové paralogie jsou geny kódující hemoglobin a myoglobin.
Promotor
Promotor je v genetice oblast DNA, která umožňuje spustit transkripci nějakého genu. Promotory jsou obvykle v blízkosti genů jejichž činnost řídí, na stejném vlákně a „proti proudu“ (upstream). Promotory jsou vazebná místa pro RNA polymerázu a pro proteiny označované jako transkripční faktory. U eukaryot je k ovlivnění promotoru nutných přinejmenším sedm rozmanitých faktorů. Promotory ovlivňují úroveň transkripce daného genu společně s dalšími regulačními oblastmi, které transkripci posilují (enhancers), tlumí (silencers), nebo tlumí možnost vazby signálu na promotor (insulators). Promotory jsou místa, jejichž prostřednictvím ovlivňují činnost genů například endokrinní signály nebo signály zevního prostředí, a to konec konců adaptivně. Mutace promotoru stejně jako mutace vedoucí k odchylkám transkripčních faktorů nebo transkripčních ko-aktivátorů je mechanismem řady onemocnění včetně zhoubných nádorů. Evoluční význam promotorů je podle některých autorů větší než změny kódujících sekvencí.
Trans-regulace viz cis-regulace
Transkripční faktor
Transkripce je kopírování DNA pomocí enzymu RNA polymerázy do poslíčkové (messenger, m) RNA. Transkripční faktor je protein, jenž se váže na specifickou sekvenci bází DNA, a tím kontroluje transkripci genetické informace z DNA do mRNA. Transkripční faktor může působit samostatně nebo v komplexu s dalšími proteiny a působit jako aktivátor transkripce nebo ji naopak blokovat. Transkripční faktor je definován jednou nebo více doménami, které se váží na specifickou sekvenci DNA sousedící s geny jejichž činnost řídí. Řada proteinů, které rovněž ovlivňují činnost genů (například kinázy, metylázy apod.), avšak nemají doménu vážící se na DNA se proto za transkripční faktor nepovažují.
Urbilateralia
Ur-bilateralita (ur něm. původní) jsou hypotetický poslední společný předek kladu (klad je vývojová větev) všech bilaterálně symetrických živočichů. Udává se, že nejstarší obecně akceptovaný bilaterální organismus je 555 Ma stará Kimberella(obr. 3A,B),předpokládá se, že jsou vývojově starší. Stopy bilateralií jsou datovány z konce ediacarské periody, tedy z období předcházejícího kambriu, počátku prvohor.
Zdroje
1. Adachi, Y., Hauck, B., Clements, J. et al. Conserved cis-regulated modules mediate complex neural expression paterns of the eyless gene in the Drosophila brain. Mech. Dev. 2003, 120, p. 113-1126.
2. Carroll, S.B. Homeiotic genes and the evolution of arthropods and chordates. Nature 1995, 376, p. 479-485.
3. Carroll, S.B. The making of the fittest. New York: WW. Norton and Company, 2006.
4. Carroll, S.B. Evo-devo and an expanding evolutionary synthesis. A genetic theory of morphological evolution. Cell 2008, 134, p. 25-36.
5. Davidson, E.H., Erwin, D.H. Gene regulatory network and the evolution of animal body plans. Science 2006, 311, p. 796-800.
6. De Robertis, E.M. Evo-devo: variation on ancestral themes. Cell 2008, 132, p.185-195.
7. Hoekstra, H.E., Coyne, J.A. The locus of evolution: evo-devo and the genetics of adaptation. Evolution Int. J. Org. Evolution 2007, 61, p. 996-1016.
8. Hyde, W.T., Crowley, T.J., Baum, S.K. et al. Neoproterozoic „snowball Earth“ simulations with a coupled climate / ice sheet model. Nature 2000, 405, p. 425-429.
9. Koukolík, F. Evoluce a evoluční teorie pro lékaře. V. Darwinovy teorie evoluce. Prakt. lék., 2010; 90, s 268–273.
10. McMahon, A.P., Ingham, O.W., Tabin, C.J. Developmental roles and clinical significance of hedge hog signalling Curr. Top. Dev. Biol. 2003, 53, p. 1-114.
11. Mueller, G.B. Evo-devo as a discipline. In: Minelli A, Fusco G. (eds.). Evolving pathways: key themes in evolutionary developmental biology. Cambridge: Cambridge University Press 2008, p. 3-29.
12. Monod, J., Jacob, F. Teleonomic mechanisms in cellular metabolism, growth, and differentiation”. Cold Spring Harbor symposia on quantitative biology 1961, 26, p. 389–401.
13. Tax, S., Callender, C. (eds.) Evolution after Darwin; the University of Chicago Centennial. Chicago: University of Chicago Press, 1960.
14. Valentine, J.W. On the origin of phyla. Chicago: University of Chicago Press, 2004.
15. Wapinski, I., Pfeffer, A., Friedman, N. et al. Natural history and evolutionary principles of gene duplication in fungi. Nature 2007, 449, p. 54-61.
16. Werneburg, I. A standard system to study vertebrate embryos. PLoS ONE 2009; 4(6): e5887. doi:10.1371/journal.pone.0005887.
Štítky
Praktické lékařství pro děti a dorost Praktické lékařství pro dospěléČlánek vyšel v časopise
Praktický lékař

2010 Číslo 6
- Efektivita kartáčku Sonicare For Kids u dětí předškolního věku
- Naděje budí časná diagnostika Parkinsonovy choroby založená na pachu kůže
- Pacient se bude cítit komfortně pouze při správně zvolené absorpční pomůcce
- Na výběru inkontinenčních pomůcek záleží − ale jak se mezi nimi neztratit?
Nejčtenější v tomto čísle
- Nežádoucí účinky nesteroidních antiflogistik
- Mezi objektivním a subjektivním, stres pacienta, psychosomatika
- Supervize jako prevence syndromu vyhoření u zdravotníků
-
Miniportréty slavných českých lékařů
Gynekolog a porodník profesor MUDr. Josef Jerie
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy