
, a -Antisense Gene of , Encodes a Evolved Protein That Inhibits GSK3β Resulting in the Stabilization of MYCN in Human Neuroblastomas
The rearrangement of pre-existing genes has long been thought of as the major mode of new gene generation. Recently, de novo gene birth from non-genic DNA was found to be an alternative mechanism to generate novel protein-coding genes. However, its functional role in human disease remains largely unknown. Here we show that NCYM, a cis-antisense gene of the MYCN oncogene, initially thought to be a large non-coding RNA, encodes a de novo evolved protein regulating the pathogenesis of human cancers, particularly neuroblastoma. The NCYM gene is evolutionally conserved only in the taxonomic group containing humans and chimpanzees. In primary human neuroblastomas, NCYM is 100% co-amplified and co-expressed with MYCN, and NCYM mRNA expression is associated with poor clinical outcome. MYCN directly transactivates both NCYM and MYCN mRNA, whereas NCYM stabilizes MYCN protein by inhibiting the activity of GSK3β, a kinase that promotes MYCN degradation. In contrast to MYCN transgenic mice, neuroblastomas in MYCN/NCYM double transgenic mice were frequently accompanied by distant metastases, behavior reminiscent of human neuroblastomas with MYCN amplification. The NCYM protein also interacts with GSK3β, thereby stabilizing the MYCN protein in the tumors of the MYCN/NCYM double transgenic mice. Thus, these results suggest that GSK3β inhibition by NCYM stabilizes the MYCN protein both in vitro and in vivo. Furthermore, the survival of MYCN transgenic mice bearing neuroblastoma was improved by treatment with NVP-BEZ235, a dual PI3K/mTOR inhibitor shown to destabilize MYCN via GSK3β activation. In contrast, tumors caused in MYCN/NCYM double transgenic mice showed chemo-resistance to the drug. Collectively, our results show that NCYM is the first de novo evolved protein known to act as an oncopromoting factor in human cancer, and suggest that de novo evolved proteins may functionally characterize human disease.
Published in the journal:
. PLoS Genet 10(1): e32767. doi:10.1371/journal.pgen.1003996
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1003996
Summary
The rearrangement of pre-existing genes has long been thought of as the major mode of new gene generation. Recently, de novo gene birth from non-genic DNA was found to be an alternative mechanism to generate novel protein-coding genes. However, its functional role in human disease remains largely unknown. Here we show that NCYM, a cis-antisense gene of the MYCN oncogene, initially thought to be a large non-coding RNA, encodes a de novo evolved protein regulating the pathogenesis of human cancers, particularly neuroblastoma. The NCYM gene is evolutionally conserved only in the taxonomic group containing humans and chimpanzees. In primary human neuroblastomas, NCYM is 100% co-amplified and co-expressed with MYCN, and NCYM mRNA expression is associated with poor clinical outcome. MYCN directly transactivates both NCYM and MYCN mRNA, whereas NCYM stabilizes MYCN protein by inhibiting the activity of GSK3β, a kinase that promotes MYCN degradation. In contrast to MYCN transgenic mice, neuroblastomas in MYCN/NCYM double transgenic mice were frequently accompanied by distant metastases, behavior reminiscent of human neuroblastomas with MYCN amplification. The NCYM protein also interacts with GSK3β, thereby stabilizing the MYCN protein in the tumors of the MYCN/NCYM double transgenic mice. Thus, these results suggest that GSK3β inhibition by NCYM stabilizes the MYCN protein both in vitro and in vivo. Furthermore, the survival of MYCN transgenic mice bearing neuroblastoma was improved by treatment with NVP-BEZ235, a dual PI3K/mTOR inhibitor shown to destabilize MYCN via GSK3β activation. In contrast, tumors caused in MYCN/NCYM double transgenic mice showed chemo-resistance to the drug. Collectively, our results show that NCYM is the first de novo evolved protein known to act as an oncopromoting factor in human cancer, and suggest that de novo evolved proteins may functionally characterize human disease.
Introduction
Gene evolution has long been thought to arise from pre-existing genes through duplication or rearrangement followed by rapid divergence [1]–[5]. De novo gene birth from non-coding genomic regions has been generally believed to be exceptionally rare [1]. However, recent studies using genome-wide analyses have suggested the presence of a large number of de novo evolved genes in some species [3], [5]–[11], including primates [12]–[17]. Studies in yeast revealed that the proteins produced from de novo genes were not insignificant polypeptides but functional proteins [6], [7] and that de novo gene birth could be a major mechanism of new gene generation [6]. In multicellular organisms, however, the functions of de novo evolved proteins have been poorly characterized [3], [15], and thus their pathophysiological significance has remained elusive. Therefore, it is still unclear whether de novo gene birth is a general mechanism throughout evolution for the creation of functional protein-coding genes.
Neuroblastoma is one of the most common solid tumors in children. It originates from the neuronal precursor cells of the sympathoadrenal lineage of the neural crest [18]. Its clinical behavior is enigmatic; the tumors in patients of less than one year of age often regress spontaneously, whereas the tumors detected in patients over one year of age are usually aggressive and eventually cause the patient's death despite intensive multimodality therapies [18]. The MYCN oncogene is frequently amplified in those tumors that occur in patients who are over one year of age at diagnosis [19], [20]. Transgenic mice expressing human MYCN in sympathoadrenal tissues spontaneously develop neuroblastomas [21], suggesting that MYCN alone can initiate tumorigenesis and promote tumor growth. However, unlike human neuroblastoma, its distant metastasis is infrequent. Furthermore, in human neuroblastomas without MYCN amplification, MYCN mRNA expression levels do not correlate with the prognosis of the patients [22], [23], suggesting that additional events might contribute to the acquisition of increased aggressiveness. We focused on NCYM as a candidate gene that promotes the aggressiveness of MYCN-amplified neuroblastomas. NCYM is a cis-antisense gene of MYCN [24], [25] and is co-amplified with MYCN in human neuroblastoma cells. NCYM is transcribed in the opposite direction to MYCN, starting from intron 1 of the MYCN gene (Figure 1A), and it has remained unclear for a long time whether the gene encodes a functional protein [24], [26]. In this study, we have found that NCYM is indeed a functional protein that regulates MYCN function in human, but not mouse, neuroblastoma.

Results
NCYM is a de novo evolved gene
We first analyzed the genomic sequence of NCYM in various species and found that in humans and chimpanzees the potential NCYM protein is composed of 109 amino acids (Figure 1B, Figure S1). We next searched for paralogs and orthologs of the human NCYM protein among other animals using the Basic Local Alignment Search Tool (BLAST) databases with an E-value threshold of 10−3. We did not find any paralogs, but identified orthologs for a probable NCYM protein in olive baboons, chimpanzees and pigmy chimpanzees. From here on, we focused on the NCYM gene of the hominidae to investigate the function of the protein product. The evolutionary rates between the indicated species suggest that the coding sequence of NCYM gene was exposed to positive selection in humans and chimpanzees (Figure 1C), and the amino acid frequencies in these species were significantly different from the uniform usage of amino acids (P<0.001; Figure S2). We next raised an antibody against the putative human NCYM protein, and identified a 12 to 15 kDa protein in human neuroblastoma cells which mainly localized to nuclei in MYCN-amplified neuroblastoma cells (Figure S3, Figure S4). The NCYM protein was expressed in a variety of normal human tissues, including the neuronal cells of the cerebrum and cerebellum, spermatocytes of the testis, pancreatic cells and also the heart (Figure S5). NCYM was also localized in both the nucleus and cytoplasm in these cells (Figure S5A–D). NCYM was expressed in both primary and metastatic human neuroblastomas (Figure 1D, Figure S5E and F), and was co-expressed with the MYCN protein in cells of human neuroblastomas (Figure 1D and E) and the neuronal cells of the human cerebrum (Figure 1F and G). It was also co-expressed with the MYCN protein in some primary human cancers, including thyroid cancer (Figure S6). Thus, the NCYM protein is a de novo evolved gene product and is endogenously expressed in both normal human tissues and cancers.
Prognostic significance of NCYM expression in human neuroblastoma
We next examined the prognostic significance of NCYM mRNA expression in human neuroblastoma. The NCYM gene was co-amplified with the MYCN gene in all the cell lines and primary neuroblastomas we examined (Figure S7). NCYM expression levels were significantly correlated with that of MYCN in primary neuroblastomas (n = 106, P = 4.69×10−16; Figure 2A) and in the tumors with a single copy of MYCN (n = 86, P = 1.11×10−13; Figure 2B). In addition, high levels of NCYM mRNA expression were significantly associated with unfavorable prognostic factors (P<0.05, Table S1) and a poor outcome (P = 3.70×10−5; Figure 2C), similar to that for MYCN mRNA expression (P<0.05; Table S1 and P = 2.31×10−5; Figure 2D). Interestingly, high levels of NCYM mRNA expression were also significantly correlated with poor outcome in those patients diagnosed at over one year of age without MYCN amplification (n = 45, P = 0.0375; Figure S8A) whereas those of MYCN did not correlate with the prognoses (n = 45, P = 0.144; Figure S8B). Multivariate analysis of 106 primary neuroblastomas showed, as expected, that NCYM mRNA expression is not an independent prognostic factor from expression and amplification of MYCN (Table S2). However, it is an independent prognostic factor from age at diagnosis, stage and TrkA expression.

Positive feedback regulation between NCYM and MYCN
The co-amplification and co-expression of NCYM and MYCN in human primary neuroblastomas prompted us to investigate the functional interaction between NCYM and MYCN. Previously we have reported that MYCN directly targets its own expression in neuroblastoma cell lines [27]. Because the promoter region of the NCYM gene is localized within intron 1 of MYCN (Figure S9A), we examined whether MYCN regulates NCYM transcription. Overexpression of MYCN in human neuroblastoma cells induced NCYM mRNA expression (Figure 3A), whereas shRNA-mediated knockdown of MYCN downregulated endogenous NCYM mRNA levels (Figure 3B). MYC overexpression did not induce either MYCN or NCYM expression (Figure S9B). However, MYCN overexpression did enhance NCYM promoter activity in a dose-dependent manner (Figure 3C), suggesting that MYCN, but not MYC, activates the transcription of NCYM. Putative E-boxes exist in intron 1 of the MYCN gene; however, it is unclear whether they are responsible for this feedback regulation. We therefore generated constructs containing different lengths of the MYCN intron 1 region and performed luciferase assays to identify the MYCN-responsive region (Figure S9C). MYCN enhances its own promoter activity in a dose-dependent manner when co-transfected with reporter plasmids containing the NCYM promoter region (from +1073 to +1312). However, when co-transfected with plasmids without this NCYM promoter region, MYCN positive autoregulation was diminished. Within this region, there is a putative E-box located just 2 base pairs upstream from the transcription start site of the NCYM gene (Figure S10A). We generated constructs containing the NCYM promoter region comprising either a wild-type or a mutant E-box. Overexpression of MYCN enhanced NCYM wild-type promoter activity, but mutation of the E-box diminished its activation (Figure S10C). MYC overexpression did not activate either of the NCYM promoter constructs (Figure S10B and C). Therefore, these results indicate that MYCN enhances NCYM promoter activity in an E-box-dependent manner. MYC, however, is not involved in NCYM transcription.
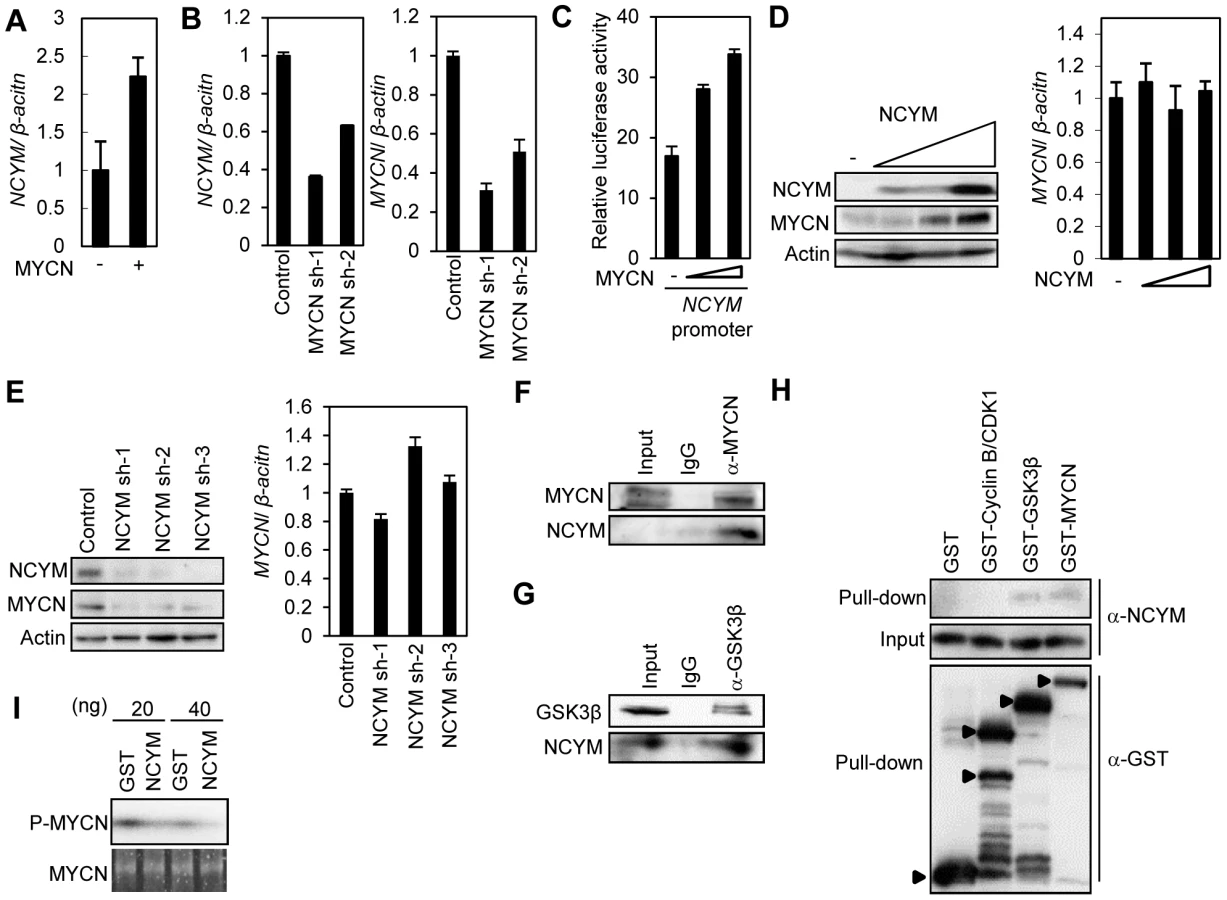
We next investigated the function of NCYM in neuroblastoma cells. NCYM overexpression induced MYCN protein levels (Figure 3D, left panel; Figure S11A), but had no effect on the mRNA levels of MYCN (Figure 3D, right panel; Figure S11A). Consistent with these results, shRNA-mediated knockdown of NCYM significantly downregulated the amount of MYCN protein without affecting the level of MYCN mRNA expression (Figure 3E). In addition, knockdown of NCYM decreased the stability of the MYCN protein (Figure S11B). This NCYM knockdown-mediated destabilization of MYCN could be inhibited using the proteasome inhibitor MG132 (Figure S11C). It is known that the stability of the MYCN protein is regulated by a series of phosphorylation and ubiquitination events that are required for its recognition by the proteasome [28]. CDK1/Cyclin B1 phosphorylates MYCN at serine 62: the mono-phosphorylated MYCN is then recognized by GSK3β and subsequently phosphorylated at threonine 58, leading to its proteasome-dependent protein degradation after an E3-mediated polyubiquitination [28], [29]. Therefore, using immunoprecipitation, we next searched for factors interacting with NCYM that are able to induce MYCN stabilization, and found that NCYM forms a complex with MYCN and GSK3β in CHP134 cells (Figure 3F and G). In addition, purified NCYM was capable of interacting with purified GSK3β and MYCN in vitro (Figure 3H). To examine the effect of NCYM on GSK3β-mediated phosphorylation of MYCN, we performed an in vitro kinase assay (Figure 3I). NCYM protein inhibited the phosphorylation of MYCN. Because the purified NCYM protein is not a substrate of GSK3β (Figure S12), it is unlikely that NCYM competes with MYCN for GSK3β as a substrate. Taken together these results suggest that the NCYM protein inhibits GSK3β-mediated MYCN phosphorylation and stabilizes the MYCN protein in vitro.
It has been reported that MYCN knockdown decreases cell proliferation and induces apoptosis and/or differentiation in MYCN-amplified neuroblastoma cells [30]. Therefore, we next investigated the functional role of NCYM in these cells (Figure S13 and S14). We performed NCYM knockdown in BE (2)-C, CHP134, SK-N-AS and SH-SY5Y human neuroblastoma cells. SK-N-AS and SH-SY5Y cells are MYCN-single copy but have a high expression of MYC, while BE (2)-C and CHP134 are cell lines with MYCN-amplification and hence have a high expression of MYCN and NCYM (Figure S13A). NCYM knockdown did not affect the survival of the MYCN-single neuroblastoma cell lines, but promoted massive apoptosis of the MYCN-amplified neuroblastoma cells (Figure S13B and C). In addition, in BE (2)-C cells, NCYM knockdown was found to inhibit cell proliferation and invasion (Figure S14B and D). These results suggest that NCYM promotes the survival and aggressiveness of MYCN-amplified neuroblastoma cells.
Co-expression of MYCN/NCYM in mice promotes neuroblastoma metastasis
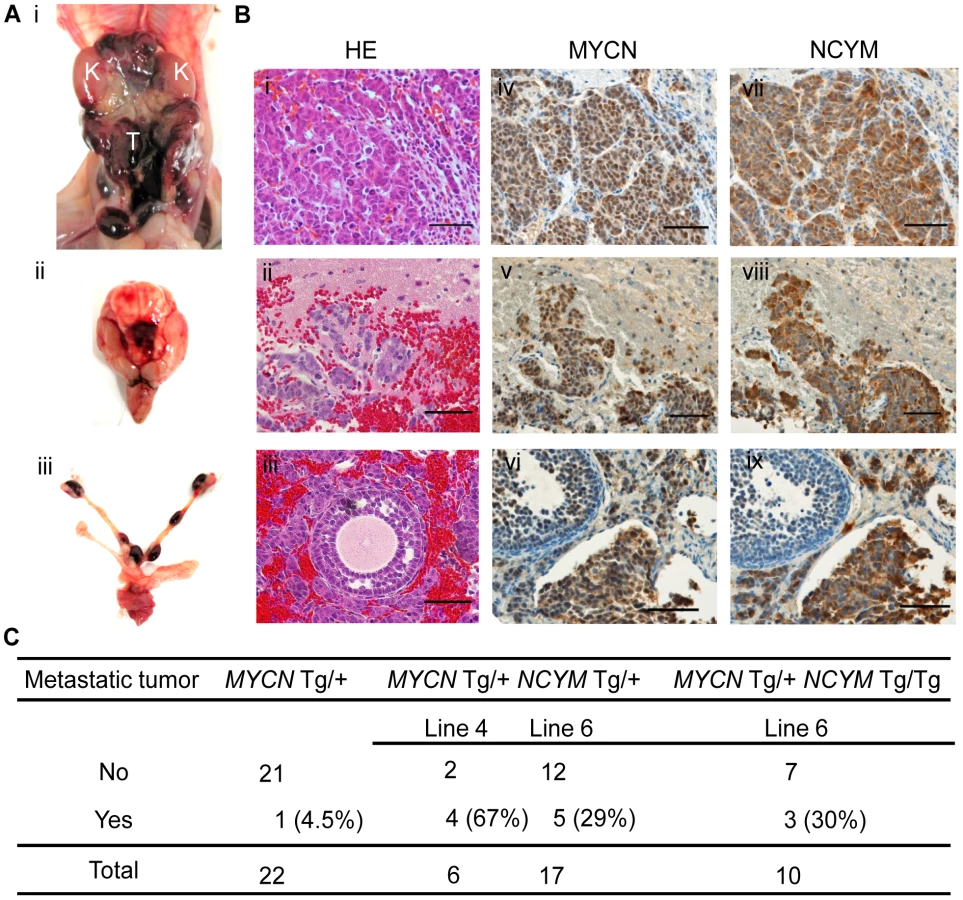
To assess the function of NCYM in vivo, we generated transgenic mice expressing the human NCYM gene under the control of the rat tyrosine hydroxylase (TH) promoter (Figure S15A and B). In addition, we made double transgenic mice carrying both the human MYCN and NCYM genes. NCYM Tg/+ mice were mated with MYCN Tg/+ NCYM Tg/+ mice, and 83 descendants were observed for 200 days (Figure S15C and D). None of the NCYM transgenic mice of the 129+ter/SVJ background developed neuroblastoma (Figure S15D), suggesting that NCYM overexpression alone is not sufficient to initiate neuroblastoma in vivo. Although tumor formation was not accelerated in the MYCN/NCYM double transgenic mice (Figure S15E), the incidence of neuroblastomas with distant metastases was significantly increased in the MYCN/NCYM double transgenic mice (Figure 4, Figure S16, Table S3). The overexpression of the MYCN and NCYM proteins in primary and metastatic tumor cells was confirmed by immunohistochemistry (Figure 4B). In the neuroblastoma tissue of the double transgenic mice, GSK3β was significantly inactivated by phosphorylation at serine 9 (Figure 5A). To investigate the mechanism by which NCYM promotes the phosphorylation of GSK3β, we analyzed the phosphorylation status of the known upstream kinases for GSK3β, AKT [28] and S6K [31]. S6K was highly phosphorylated in the MYCN/NCYM double transgenic mice, whereas AKT was not noticeably activated. The phosphorylation levels of S6K in neuroblastomas from the MYCN/NCYM double transgenic mice were correlated with the expression levels of MYCN and NCYM (Figure 5A, M7-M11). These results suggest that NCYM promotes the phosphorylation of GSK3β via the activation of mTOR-S6K signaling. Furthermore, NCYM co-immunoprecipitated with GSK3β (Figure 5B) and substrates of GSK3β such as MYCN and β-catenin were stabilized in the neuroblastoma tissues induced in MYCN/NCYM transgenic mice (Figure 5A). We next examined the number of apoptotic cells in neuroblastomas from MYCN transgenic mice and MYCN/NCYM double transgenic mice by staining for cleaved caspase-3 (Figure S17). The number of apoptotic tumor cells was significantly decreased in the primary tumors of MYCN/NCYM double transgenic mice, suggesting that NCYM promotes the survival of neuroblastoma cells in vivo.

The tumors which develop in MYCN/NCYM transgenic mice are resistant to PI3K/mTOR inhibition
To examine whether the overexpression of NCYM contributes to the chemosensitivity of neuroblastomas via GSK3β inhibition, we tested the effect of NVP-BEZ235 on the survival of the MYCN/NCYM double transgenic mice. NVP-BEZ235 is a dual inhibitor of both PI3K and mTOR and promotes the degradation of MYCN to effectively reduce tumor burden in the MYCN transgenic mouse via GSK3β activation [32]. As reported, NVP-BEZ235 treatment significantly prolonged the survival duration of the MYCN transgenic mice (P<0.01; Figure 5C). In contrast NVP-BEZ235 did not prolong the survival of the MYCN/NCYM double transgenic mice (P = 0.648; Figure 5D). Thus, the expression of NCYM reduced the efficiency of this drug in vivo.
Discussion
Our results reveal that NCYM, which was initially thought to be a large non-coding RNA transcribed from a cis-antisense gene of human MYCN [26], is actually translated into a functional protein in humans. MYCN is a highly conserved, major oncogene in human cancer. The newly evolved cis-antisense NCYM gene product targets the sense MYCN gene product, influencing its stabilization, which in turn enhances transcription of the NCYM gene. This positive autoregulatory loop may function in primary human neuroblastomas to enhance metastasis as well as drug resistance through stabilization of MYCN and even β-catenin, which are mediated by inhibition of GSK3β (Figure S18). Thus, NCYM is the first de novo evolved gene product shown to function in the development of human neuroblastoma.
NCYM promoted phosphorylation of GSK3β at serine 9 possibly via the activation of mTOR-S6K signaling, that might have led to the constitutive inactivation of GSK3β in vivo. Recently, Schramm et al. reported that MYCN transcriptionally regulates the mTOR pathway, promoting its activation [33]. Thus, MYCN might have enhanced S6K phosphorylation by activating the mTOR pathway in neuroblastomas caused in the double transgenic mice. Previous reports have suggested that neuroblastoma cell lines expressing high levels of MYCN were significantly more sensitive to mTOR inhibitors compared with cell lines expressing low MYCN levels [34]. Furthermore, our study showed that NCYM knockdown significantly induces apoptosis in MYCN-amplified neuroblastoma cells, whereas the effects were marginal in MYCN-single neuroblastoma cells. Therefore, the feedback regulation between mTOR-S6K signaling and MYCN/NCYM may contribute to the survival of MYCN-amplified neuroblastoma cells (Figure S18).
Although NCYM inhibits GSK3β-mediated MYCN phosphorylation in vitro, our data does not rule out the possibility that NCYM may stabilize MYCN in a GSK3β-independent manner. Because NCYM binds directly to MYCN both in vitro and in neuroblastoma cells, this may affect the recruitment of the regulators of MYCN stability. Indeed, we have recently found that the tumor suppressor protein Runx3 directly binds to MYCN in neuroblastoma cells and promotes degradation of MYCN in the ubiquitin–proteasome system dependent manner [35]. Therefore, the binding of NCYM to MYCN itself could affect the interaction of Runx3, or other regulators such as Aurora A [36] with MYCN to induce its stability. Further studies are required to evaluate the role of NCYM-mediated inhibition of GSK3β activity on MYCN stability.
Recent reports have suggested that both mutant ALK [37], [38] and Lin28B [39] promote the growth of neuroblastomas in transgenic mouse models by targeting MYCN for stabilization [37], [38] or overexpression [39]. Among the known regulators of MYCN, NCYM is the only gene that shows 100% co-amplification with MYCN in human primary neuroblastomas. Overexpressed NCYM stabilizes both MYCN and β-catenin, and enhances the generation of neuroblastomas with increased aggressive behavior such as distant metastasis and/or drug resistance, which are characteristics reminiscent of human neuroblastoma. Recently, Valentijn et al. suggested that the activation of the MYCN pathway is a more significant prognostic factor than the expression or amplification of MYCN in primary neuroblastomas [40]. Consistent with this, our results indicate that NCYM expression is associated with poor outcomes in human neuroblastoma regardless of genomic status of the MYCN/NCYM locus. Therefore, we anticipate that the positive auto-regulatory loop formed by MYCN and NCYM may be a promising target for developing novel therapeutic tools against high-risk neuroblastoma.
As suggested in the recent report [37], the concomitant inhibition of apoptosis and/or activation of survival signals may be required for MYCN to induce multiple tumors or metastases in vivo. In this study, we found that NCYM maintains the survival of MYCN-amplified neuroblastoma cells, and that the apoptotic cell number, indicated by cleaved caspase-3, was downregulated in MYCN/NCYM transgenic mice. In addition, GSK3β inhibition contributes to the inhibition of apoptosis in response to treatment with DNA-damaging drugs in neuroblastoma cells [41]. Therefore, the concomitant activation of other GSK3β substrates, such as β-catenin, and mTOR-S6K signaling by NCYM may be involved in the inhibition of apoptosis (Figure S18).
Since the proposals of Ohno and Jacob, the birth of a new gene has been believed to be caused by the duplication or rearrangement of pre-existing genes [1], [2]. The recent advances in whole genome sequencing technology and bioinformatics have identified the presence of de novo proteins; however, their physiological or pathological significance have largely remained unclear [3], [15]. In 2010, Li et al. reported that MDF1 originated de novo from a DNA sequence previously thought to be non-coding in Saccharomyces cerevisiae [7]. MDF1 inhibits mating efficiency by binding MATα2 and promoting vegetative growth. Therefore, while MDF1 was the first reported de novo gene whose protein product function was unveiled in a monad, NCYM may be the first de novo protein whose precise function has been clarified in multicellular organisms, specifically in humans.
In conclusion, NCYM is a de novo evolved protein which acts as an oncopromoting factor in human neuroblastoma. Our results suggest that de novo evolved new gene products may be involved in the functional regulation of human cancers and even other diseases.
Materials and Methods
Evolutionary analyses
DNA sequences of all species were extracted from the UCSC genome browser on the basis of conservation. From the protein-coding regions, we took the conserved block that was annotated as the region corresponding to the NCYM coding sequence, located in exon 3. For intron sequences, we used BLAT [42] to align the NCYM mRNA sequence (NR_026766) to the genome of each species and extracted the unmapped regions in the alignment. We found exactly two unmapped regions for each species except for mouse (and thus did not use the mouse sequence). For intergenic regions, we used multiz [43] alignment across 48 species in the browser and cut out 1000-bp sequences that corresponded to human intergenic regions. The sequences of common ancestors were estimated based on the maximum parsimony principle that led to the minimum number of nucleotide-base changes along the already-known phylogenetic tree of the five primates and mice [16]. For multiple possibilities with the same minimum number, we broke the tie by selecting the nucleotide base of the closest outgroup (e.g., when we had A for human, T for chimpanzee, and T for orangutan, we chose T for the common ancestor of human and chimpanzee). When multiple possibilities still remained (as in common ancestor 1), we considered all the possibilities to be equally probable. We estimated common ancestor sequences only within close species (human, chimpanzee, orangutan, and rhesus macaque). We used BLAST [44] to make an alignment between two translated amino-acid sequences ending at the first terminal codon, and calculated Ka and Ks using the KaKs_Calculator [45] with the ‘gMYN’ method, where Ka and Ks are the rates of non-synonymous and synonymous amino-acid changes, respectively. All pairs of sequences were aligned entirely from the start codon to the terminal codon and did not include any indels, except for the alignment between common ancestor 2 and common ancestor 3, for which we noted ‘frameshift’ instead of the Ka and Ks values.
We measured a bias in the codon frequencies (or amino acid frequencies) through the deviation from the uniform usage of each codon, using the Chi-squared statistic normalized to the number of codons:

Generation of a human NCYM antibody
A polyclonal anti-NCYM antibody was raised in rabbits against a 14-amino acid stretch at the C-terminal region of NCYM (84-LGTRPLDVSSFKLK-97) (Medical and Biological Laboratories, Nagoya, Japan). The specificity of the purified antibody's affinity was assessed by immunoblotting.
Immunohistochemistry
Neuroblastoma tissues obtained from mice were fixed in 4% paraformaldehyde and paraffin-embedded for histological studies. Tissue sections were stained with hematoxylin and eosin (H&E) and examined histologically by pathologists for confirmation of the tumor type. Tissue arrays (FDA808a-1 and FDA808a-2, US Biomax, Rockville, MD, USA) were used for the analyses of NCYM or MYCN expression in normal and tumorous human tissues. For immunohistochemistry, tissue sections were stained with the polyclonal anti-NCYM antibody we generated, an anti-MYCN antibody (Calbiochem, San Diego, CA, USA), and cleaved Caspase-3 (Cell Signaling Technology).
Immunofluorescence
MYCN-amplified human neuroblastoma TGW cells grown on coverslips were fixed with 4% paraformaldehyde in 1× PBS for 20 min at 4°C, permeabilized with 0.1% Triton-X for 20 min at room temperature, and then incubated with 2% BSA and 3% goat serum in PBS for 1 h to reduce nonspecific binding. Immunostaining was performed by incubating cells with the polyclonal anti-NCYM antibody and a monoclonal anti-MYCN antibody (Calbiochem) for 2 h at room temperature in a humidified chamber, followed by incubation with fluorescent-conjugated goat anti-rabbit IgG (diluted 1∶400) or fluorescent-conjugated goat anti-mouse IgG (diluted 1∶400), respectively. The coverslips were washed extensively with PBS, mounted in VECTASHIELD mounting medium with DAPI (Vector Laboratories, Burlingame, CA, USA) and images were captured using a confocal microscope (DMI 4000B, Leica).
Plasmids
We previously made a MYCN-luc (+1312) plasmid that contains the region of MYCN promoter region spanning from −221 to +1312 (where +1 represents the transcription start site) [27]. Luciferase reporter plasmids containing different lengths of the MYCN promoter were generated from MYCN-Luc (+1312) by partial removals of the MYCN promoter region with appropriate restriction enzymes. The MYCN promoter region in MYCN-luc (+1312) was subcloned into the pGL3basic vector or pGL4.17ΔEcoRV EcoRI vector in the opposite direction for generation of NCYM-luc vectors. pGL4.17 ΔEcoRV EcoRI was the luciferase reporter plasmid, where an EcoRV site in pGL4.17 (Promega, Southampton, UK) is replaced with an EcoRI site. NCYM-luc E-box WT and NCYM-luc E-box MT were generated by PCR-based amplification using MYCN-luc (+1312) as a template. Oligonucleotide primers used were as follows: 5′-AACCAGGTTCCCCAATCTTC-3′ (forward) and 5′-ACCACCCCCTGCATCTGCAT-3′ (reverse, NCYM-luc E-box WT) or 5′-ACCACCCCCTGCATCCGCAT-3′ (reverse, NCYM-luc E-box MT). Underlined sequences in the reverse primers indicate the wild-type or mutant E-boxes. The NCYM complementary DNA was introduced into a pcDNA3 expression vector, comprising a FLAG-tag at the 5′ locus of NCYM to generate pcDNA3-FLAG-NCYM. The sequence of the entire NCYM open reading frame was confirmed by sequencing. The FLAG-NCYM cDNA was ligated downstream of the rat TH promoter in the pGEM7z(f+) expression plasmid, which was originally made from a MYCN transgenic construct [21] by excision of the MYCN gene, to generate pGEM7z(f+)-FLAG-NCYM.
Cell culture, infection, transfection, and RNA interference
Human neuroblastoma cell lines SH-SY5Y, SK-N-AS, NLF, IMR32, CHP134, and SK-N-BE were maintained in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS) and antibiotics. Human neuroblastoma cell line BE (2)-C was maintained in a 1∶1 mixture of minimal essential medium (MEM, Gibco by Life technologies, Carlsbad, CA, USA) and Ham's Nutrient Mixture F12 (Gibco) supplemented with 15% heat inactivated fetal bovine serum (FBS) (Gibco) with MEM non-essential amino acids (Gibco) and antibiotics. Mouse neuroblastoma cell line Neuro 2a was maintained in DMEM supplemented with 10% FBS and antibiotics. NLF, IMR32, CHP134, SK-N-BE, and BE (2)-C have amplified MYCN, whereas SH-SY5Y, SK-N-AS, and Neuro 2a are cell lines with a single copy of MYCN. The cells or tissues with a single copy of MYCN have one copy of MYCN gene in a haploid genome. Lentivirus was produced by co-transfecting cDNA or shRNA expression plasmids with pCMVR and pMDG plasmids into HEK293T cells using FuGENE HD reagent (Roche, Mannheim, Germany). The MYCN and NCYM shRNA expression plasmids contained pLKO.1-puro as the backbone (Sigma, St Louis, MO, USA). At 24 and 48 h after transfection, the viral supernatants were collected and mixed with neuroblastoma cells. Other plasmid transfections were done using Lipofectamine 2000 transfection reagent (Invitrogen, Karlsruhe, Germany) according to the manufacturer's instructions. The target sequences of the shRNAs used were as follows: NCYM sh-1 (N-cym1 custom shRNA, Sigma) 5′-tggcaattgcttgtcattaaa-3′, NCYM sh-2 (N-cym 2 custom shRNA, Sigma) 5′-gaggttgctcctgtgtaatta-3′, NCYM sh-3 (N-cym 3 custom shRNA, Sigma) 5′-tcctgtgtaattacgaaagaa-3′, MYCN sh-1 (TRCN0000020694, Sigma) 5′-gccagtattagactggaagtt-3′, MYCN sh-2 (TRCN0000020695, Sigma) 5′-cagcagcagttgctaaagaaa-3′. The control shRNA (SHC002) was purchased from Sigma.
RNA isolation, RT-PCR and quantitative real-time RT-PCR
Total RNA was isolated from the frozen tumor samples and adrenal tissues of transgenic mice with ISOGEN (NIPPON GENE, Tokyo, Japan), and treated with RNase-free DNase I. Total RNA from neuroblastoma cells (CHP134 and SK-N-AS) was prepared using an RNeasy Mini kit (Qiagen, Valencia, CA) following the manufacturer's instruction. cDNA was synthesized using SuperScript II with random primers (Invitrogen). Quantitative real-time RT-PCR (qRT-PCR) using an ABI PRISM 7500 System (Applied Biosystems, Foster City, CA) was carried out using a SYBR green PCR reaction. The primer sets used were as follows: (for clinical experiments using primary neuroblastomas) human MYCN, 5′-ggacaccctgagcgattcag-3′, and 5′-aggaggaacgccgcttct-3′, human NCYM 5′-ccgacagctcaaacacagaca-3′ and 5′ - gtaatggcttctgcgaaaagaaa-3′; (for cellular experiments) human MYCN, 5′-tccatgacagcgctaaacgtt-3′ and 5′ - ggaacacacaaggtgacttcaaca-3′, human NCYM, 5′-cgcccccttaggaacaagac-3′ and 5′ - gcgcccctcttctttcaatt-3′, mouse MYCN, 5′-tcgggacactaaggagcttca-3′ and 5′-ggaatcttggaccggaacaa-3′, mouse GAPDH, 5′-gggaagcccatcaccatct-3′ and 5′-cggcctcaccccatttg-3′. The mRNA levels of each of the genes were standardized by β-actin or GAPDH.
Luciferase assay
SK-N-AS cells were co-transfected with the indicated reporter constructs and the pRL-TK Renilla luciferase cDNA together with increasing amounts of the expression plasmid for MYCN or MYC. Total DNA per transfection was kept constant (510 ng) by adding pcDNA3 (Invitrogen). Forty-eight hours after transfection, firefly and Renilla luciferase activities were measured with a dual-luciferase reporter assay system according to the manufacturer's instructions (Promega).
Immunoblotting
We resolved cell proteins by SDS-PAGE before electro-blotting onto PVDF membranes. We incubated the membranes with the following primary antibodies overnight: anti-NCYM (1∶1000 dilution), anti-MYCN antibody (1∶1000 dilution; Calbiochem and Cell Signaling), anti-Lamin B (1∶1000 dilution; Calbiochem), anti-α-tubulin (1∶1000 dilution; Santa Cruz, CA, USA), anti-GST (1∶1000; Santa Cruz), anti-GSK3β (1∶1000 dilution; Cell Signaling), anti-phospho-GSK3β (S9) (1∶1000 dilution; Cell Signaling), anti-β-catenin (1∶1000 dilution; Cell Signaling), anti-phospho-AKT (S473) (1∶1000 dilution; Cell Signaling), anti-phospho-AKT (S308) (1∶1000 dilution; Cell Signaling), anti-AKT (1∶1000 dilution; Cell Signaling), anti-S6K (1∶1000 dilution; Cell Signaling), anti-phospho-S6K (T389) (1∶1000 dilution; Cell Signaling), and anti-actin (1∶4000 dilution; Sigma). The membranes were then incubated with a horseradish peroxidase-conjugated secondary antibody (anti-rabbit IgG at 1∶2000–1∶4000 dilution or anti-mouse IgG at 1∶2000 dilution; both from Cell Signaling Technology) and the bound proteins were visualized using a chemiluminescence-based detection kit (ECL and ECL pro kit, Amersham, Piscataway, NJ, USA; ImmunoStar LD, Wako).
Immunoprecipitation
Whole lysates prepared from CHP134 cells or tumor tissues were pre-cleared by incubation with protein G-Sepharose beads (Amersham Pharmacia Biotech) for 1 h at 4°C. The supernatant was collected after a brief centrifugation, and incubated with the indicated primary antibodies at 4°C overnight. The immune complexes were precipitated with protein G-Sepharose beads for 1 h at 4°C, and the non-specific bound proteins were removed by washing the beads with lysis buffer five times at 4°C. Different lysis buffers were used for the cell-based experiments (50 mM Tris-HCl pH 8.0, 137 mM NaCl, 2.7 mM KCl, and 1% Triton X) and for the tumor tissues (50 mM Tris-HCl pH 8.0, 1 mM EDTA, 0.2% DOC and 0.2% SDS). The immunoprecipitated proteins were eluted by boiling in Laemmli sample buffer and analyzed by immunoblotting.
Analysis of MYCN stability
CHP134 cells were cultured with 50% lentiviral supernatant for transfection of the indicated shRNA. Forty-eight hours after the transfection, cycloheximide (Sigma) was added to the culture medium at a final concentration of 50 µg/ml and cells were harvested at the indicated time points. For MG132 treatment, 44 h after the transfection, cells were treated with DMSO or 10 µM MG132 for 4 h.
Purification of NCYM protein from bacteria
DH5α cells were transformed with pGEX-4T-NCYM plasmid and cultured in Luria Broth (LB) at 37°C. The expression of the GST-NCYM fusion protein was induced by culturing the cells with 1 mM IPTG for 10 h at 25°C. Cells were collected by centrifugation, dissolved in cell lysis buffer (PBS, 1% TritonX-100, 5 mM EDTA and protease inhibitors), and stored at −80°C. Cell extracts were obtained by thawing the frozen cells, followed by sonication and ultra-centrifugation. After a pulldown with glutathione sepharose 4B beads, the beads were washed five times in cell lysis buffer and once in thrombin buffer containing 50 mM Tris-HCl pH 8.0, 150 mM NaCl, 2.5 mM CaCl2, 5 mM MgCl2, 1 mM DTT. GST-Tag cleavage mediated by thrombin released the full-length NCYM protein from the beads and the thrombin was removed by adding p-aminobenzamidine agarose beads according to the standard protocol. The full length NCYM protein was further purified by filtration using Amicon Ultra-4 (Millipore, Temecula, CA, USA), and dissolved in stock buffer (50 mM Tris-HCl pH 8.0, 150 mM NaCl, 5 mM EDTA, 0.25 mM DTT, 10% sodium azide, 50% glycerol and protease inhibitors) and stored at −20°C. Complete PIC (Roche) was used for protease inhibition.
GST-pulldown assay
For GST-pulldown assay, 0.5 µg of purified NCYM proteins were incubated with 0.5 µg of GST protein or GST-fused CDK1/Cyclin B1 (Signal Chem, Richmond, Canada), GSK3β (Promega) and MYCN (Abnova, Taipei, Taiwan) for 2 h at 4°C. Bound complexes were recovered on the glutathione-sepharose beads, washed with the binding buffer (50 mM Tris-HCl, pH 8.0, 1 mM EDTA, 150 mM NaCl, 0.1% Nonidet P-40 and Complete PIC), boiled in in Laemmli sample buffer and analyzed by immunoblotting.
In vitro kinase assay
For MYCN phosphorylation, two kinase reactions were performed sequentially. The first kinase reactions were performed for 1 h in kinase buffer (40 mM Tris-HCl pH 7.5, 20 mM MgCl2, 0.1 mg/ml BSA, 50 µM DTT) in the presence of 50 µM Ultrapure ATP (Promega), 50 ng of purified MYCN (Abnova), and 40 ng of purified CDK1/Cyclin B1 (Signal Chem) at room temperature. At 1 h, the first reaction solution was mixed with the same volume of kinase buffer containing 100 nM CDK1 inhibitor (CGP74514A, Calbiochem), 4 µCi of [γ-32P] ATP (PerkinElmer), and 20 ng of purified GSK3β with the indicated amounts of purified NCYM or purified GST. The second reaction was performed for 1 h at room temperature. The amount of phosphorylated MYCN was quantified using standard autoradiography. The total amount of MYCN was quantified by using an Oriole Fluorescent Gel stain (Bio-Rad). We also examined whether purified NCYM could be a substrate of GSK3β using the ADP-Glo system (Promega) according to manufacturer's instructions. Reactions were performed for 1 h in kinase buffer (40 mM Tris-HCl pH 7.5, 20 mM MgCl2, 0.1 mg/ml BSA, 50 µM DDT) in the presence of 25 µM Ultrapure ATP (Promega) and 25 ng of purified GSK3β with increasing amounts of NCYM or GST at room temperature. The peptide of human muscle glycogen synthase-1 (YRRAAVPPSPSLSRHSSPHQ(pS)EDEEE) was used as a positive control for the GSK3β substrate. At 1 h, the reaction solutions were mixed and incubated with ADP-Glo reagent for 40 min at room temperature, and the mixture was combined with a kinase detection reagent and allowed to stand for 30 min. The kinase activities were detected using a luminometer (PerkinElmer ARVOX3).
TUNEL staining
The indicated neuroblastoma cells were transfected with the indicated shRNA with 50% lentiviral supernatant. Seventy-two hours after transfection, all cells were collected by centrifugation, attached onto the coverslips by CYTOSPIN 4 (Thermo Fisher Scientific, Wilmington, DE, USA), and fixed in 4% paraformaldehyde for 1 h. Apoptotic cells were detected by using an in situ cell death detection kit (Roche) according to the manufacturer's protocol. The coverslips were mounted with DAPI-containing mounting medium (Vector Laboratories) and observed under a confocal microscope.
Cell viability assay (MTT assay)
Cell viability was quantified by the 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) method. Cells were collected and seeded in 96-well plates at 1×104 cells/ml. After addition of 10 µl of MTT tetrazolium salt (Sigma) solution to each well, the plates were incubated in a CO2 incubator for 60 min. The absorbance of each well was measured using a Dynatech MR5000 plate reader with a test wavelength of 450 nm and a reference wavelength of 630 nm.
Migration and invasion assay
The invasive potential of BE (2)-C cells in vitro was measured by evaluating the number of invading cells using Matrigel-coated trans-well inserts (BD Biosciences) according to the manufacturer's instructions. BE (2)-C cells transfected with the indicated shRNA were seeded onto an insert containing 8 µm pores (BD Biosciences) in a 24-well plate at 1×105 cells/ml. Cells on the lower side of the membrane were fixed with 4% paraformaldehyde and stained using a Diff Quick Staining Kit (Sysmex).
Generation of transgenic mice
All animal experimental procedures used in this study were reviewed and approved by the Committee on the Ethics of Animal Experiments of the Chiba Cancer Center (Permit Number: 12–13). Linearized and purified pGEM7z (f+)-FLAG-NCYM was injected into the pronuclei of fertilized eggs derived from 129/SvJ×C57BL/6J mice. We selected four lines of NCYM transgenic mice according to the level of NCYM expression in adrenal tissues (Figure S15B), and the transgenic mice were backcrossed to 129/SvJ at least 10 times to generate NCYM transgenic mice. To generate MYCN/NCYM double transgenic mice, the NCYM transgenic mice were crossed with MYCN transgenic mice of the 129/SvJ strain. On the basis of breeding schemas, all mice carrying the MYCN transgene were hemizygous. Tail DNA was analyzed for MYCN and NCYM transgenes, and the NCYM transgene copy number was quantified by quantitative genomic PCR. The primer sets used for genotyping were as follows: NCYM, 5′-cgcccccttaggaacaagac-3′ and 5′ - gcgcccctcttctttcaatt-3′, MYCN, 5′-tggaaagcttcttattggtagaaacaa-3′ and 5′-agggatcctttccgccccgttcgttttaa-3′.
Detection of metastatic tumors in mice
If more than one tumor over 2 mm in a diameter separately developed in a different organ, we defined this as the mouse having macroscopic metastatic tumors. In Figure 4C, only the number of mice with macroscopic metastatic tumors was counted. As a preliminary experiment, we used microscopy to detect tumors in the brain, pancreas, spleen, heart, lungs, kidneys and liver in nine mice (MYCN/NCYM double transgenic mice; n = 6, MYCN transgenic mice; n = 3). In addition to macroscopic metastases in the brain, heart, ovary and uterus, we found microscopic metastases in the lungs of MYCN/NCYM double transgenic mice, but the mass of these tumor cells was not large enough to be visible by eye. We also microscopically analyzed the HE-stained bone marrow from the hind legs of 19 mice (MYCN/NCYM double transgenic mice, n = 10; MYCN transgenic mice, n = 9). However, no metastatic tumor cells were found in the bone marrow.
Murine therapy
All mice were genotyped to detect the presence of human MYCN or NCYM transgenes. After weaning, at about 30 days old, MYCN transgenic mice or MYCN/NCYM double transgenic mice were palpated for intra-abdominal tumors every day. Mice of either genotype found with palpable tumors were treated with NVP-BEZ235 (Cayman Chemical, Ann Arbor, MI, USA) (35 mg/kg in PEG300) or vehicle (PEG300, Wako) once daily for 30 days by oral gavage. All mice were monitored until euthanasia was required in accordance with the institutional animal committee.
Tumor specimens
The 106 human neuroblastoma specimens used in the present study were kindly provided by various institutions and hospitals in Japan to the Chiba Cancer Center Neuroblastoma Tissue Bank. Written informed consent was obtained at each institution or hospital. This study was approved by the Chiba Cancer Center Institutional Review Board. Tumors were classified according to the International Neuroblastoma Staging System (INSS): 27 Stage 1, 15 Stage 2, 34 Stage 3, 23 Stage 4, and 7 Stage 4 s. Clinical information including age at diagnosis, tumor origin, Shimada's histology, prognosis and survival duration of each patient was obtained. The patients were treated following the protocols proposed by the Japanese Infantile Neuroblastoma Cooperative Study and the Group for the Treatment of Advanced Neuroblastoma and subjected to survival analysis. Cytogenetic and molecular biological analysis of all tumors was also performed by assessing DNA ploidy, MYCN amplification and TrkA expression, as previously described [46].
Array CGH analysis
Array CGH analysis was conducted using the Human Genome CGH 244K Oligo Microarray Kit (G4411B, Agilent Technologies, Santa Clara, CA, USA). Genomic DNA prepared from primary neuroblastoma tissues or cell lines was labeled with Cy3-dye using a QuickAmp labeling kit. Human placental DNA was labeled with Cy5-dye and used as a reference control. Labeling, hybridization and subsequent data processing by FeatureExtraction and CGH-Analytics software were performed according to the manufacturer's instructions. Relative copy number of the probes surrounding the MYCN and NCYM genomic locus (from DDX1 to FAM49A) were compared in each primary tumor or cell line.
Statistical analysis
Statistical significance was tested as follows: two-group comparison of survival by log-rank test, correlation of gene expression by Pearson's correlation coefficient test or Student's t-test, multivariate analysis for survival by Cox regression model, and the rate of mouse genotype and metastatic tumor occurrence in line 6 was calculated by Chi-square independence test and Mann–Whitney U test, respectively.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. JacobF (1977) Evolution and tinkering. Science 196 : 1161–1166.
2. Ohno S (1970) Evolution by gene duplication. New York, NY, Springer-Verlag.
3. TautzD, Domazet-LosoT (2011) The evolutionary origin of orphan genes. Nat Rev Genet 12 : 692–702.
4. KaessmannH (2010) Origins, evolution, and phenotypic impact of new genes. Genome Res 20 : 1313–1326.
5. KhalturinK, HemmrichG, FrauneS, AugustinR, BoschTC (2009) More than just orphans: are taxonomically-restricted genes important in evolution? Trends Genet 25 : 404–413.
6. CarvunisAR, RollandT, WapinskiI, CalderwoodMA, YildirimMA, et al. (2012) Proto-genes and de novo gene birth. Nature 487 : 370–374.
7. LiD, DongY, JiangY, JiangH, CaiJ, et al. (2010) A de novo originated gene depresses budding yeast mating pathway and is repressed by the protein encoded by its antisense strand. Cell Res 20 : 408–420.
8. BegunDJ, LindforsHA, ThompsonME, HollowayAK (2006) Recently evolved genes identified from Drosophila yakuba and D. erecta accessory gland expressed sequence tags. Genetics 172 : 1675–1681.
9. BegunDJ, LindforsHA, KernAD, JonesCD (2007) Evidence for de novo evolution of testis-expressed genes in the Drosophila yakuba/Drosophila erecta clade. Genetics 176 : 1131–1137.
10. CaiJ, ZhaoR, JiangH, WangW (2008) De novo origination of a new protein-coding gene in Saccharomyces cerevisiae. Genetics 179 : 487–496.
11. ChenST, ChengHC, BarbashDA, YangHP (2007) Evolution of hydra, a recently evolved testis-expressed gene with nine alternative first exons in Drosophila melanogaster. PLoS Genet 3: e107.
12. Toll-RieraM, BoschN, BelloraN, CasteloR, ArmengolL, et al. (2009) Origin of primate orphan genes: a comparative genomics approach. Mol Biol Evol 26 : 603–612.
13. KnowlesDG, McLysaghtA (2009) Recent de novo origin of human protein-coding genes. Genome Res 19 : 1752–1759.
14. LiCY, ZhangY, WangZ, CaoC, ZhangPW, et al. (2010) A human-specific de novo protein-coding gene associated with human brain functions. PLoS Comput Biol 6: e1000734.
15. O'BlenessM, SearlesVB, VarkiA, GagneuxP, SikelaJM (2012) Evolution of genetic and genomic features unique to the human lineage. Nat Rev Genet 13 : 853–866.
16. WuDD, IrwinDM, ZhangYP (2011) De novo origin of human protein-coding genes. PLoS Genet 7: e1002379.
17. XieC, ZhangYE, ChenJY, LiuCJ, ZhouWZ, et al. (2012) Hominoid-specific de novo protein-coding genes originating from long non-coding RNAs. PLoS Genet 8: e1002942.
18. BrodeurGM (2003) Neuroblastoma: biological insights into a clinical enigma. Nat Rev Cancer 3 : 203–216.
19. BrodeurGM, SeegerRC, SchwabM, VarmusHE, BishopJM (1984) Amplification of N-myc in untreated human neuroblastomas correlates with advanced disease stage. Science 224 : 1121–1124.
20. SchwabM, VarmusHE, BishopJM, GrzeschikKH, NaylorSL, et al. (1984) Chromosome localization in normal human cells and neuroblastomas of a gene related to c-myc. Nature 308 : 288–291.
21. WeissWA, AldapeK, MohapatraG, FeuersteinBG, BishopJM (1997) Targeted expression of MYCN causes neuroblastoma in transgenic mice. EMBO J 16 : 2985–2995.
22. CohnSL, LondonWB, HuangD, KatzensteinHM, SalwenHR, et al. (2000) MYCN expression is not prognostic of adverse outcome in advanced-stage neuroblastoma with nonamplified MYCN. J Clin Oncol 18 : 3604–3613.
23. NakagawaraA, ArimaM, AzarCG, ScavardaNJ, BrodeurGM (1992) Inverse relationship between trk expression and N-myc amplification in human neuroblastomas. Cancer Res 52 : 1364–1368.
24. ArmstrongBC, KrystalGW (1992) Isolation and characterization of complementary DNA for N-cym, a gene encoded by the DNA strand opposite to N-myc. Cell Growth Differ 3 : 385–390.
25. KrystalGW, ArmstrongBC, BatteyJF (1990) N-myc mRNA forms an RNA-RNA duplex with endogenous antisense transcripts. Mol Cell Biol 10 : 4180–4191.
26. JacobsJF, van BokhovenH, van LeeuwenFN, Hulsbergen-van de KaaCA, de VriesIJ, et al. (2009) Regulation of MYCN expression in human neuroblastoma cells. BMC Cancer 9 : 239.
27. SuenagaY, KanekoY, MatsumotoD, HossainMS, OzakiT, et al. (2009) Positive auto-regulation of MYCN in human neuroblastoma. Biochem Biophys Res Commun 390 : 21–26.
28. GustafsonWC, WeissWA (2010) Myc proteins as therapeutic targets. Oncogene 29 : 1249–1259.
29. SjostromSK, FinnG, HahnWC, RowitchDH, KenneyAM (2005) The Cdk1 complex plays a prime role in regulating N-myc phosphorylation and turnover in neural precursors. Dev Cell 9 : 327–338.
30. KangJH, RychahouPG, IsholaTA, QiaoJ, EversBM, et al. (2006) MYCN silencing induces differentiation and apoptosis in human neuroblastoma cells. Biochem Biophys Res Commun 351 : 192–197.
31. ZhangHH, LipovskyAI, DibbleCC, SahinM, ManningBD (2006) S6K1 regulates GSK3 under conditions of mTOR-dependent feedback inhibition of Akt. Mol Cell 24 : 185–197.
32. ChantheryYH, GustafsonWC, ItsaraM, PerssonA, HackettCS, et al. (2012) Paracrine signaling through MYCN enhances tumor-vascular interactions in neuroblastoma. Sci Transl Med 4 : 115ra113.
33. SchrammA, KösterJ, MarschallT, MartinM, SchwermerM, et al. (2013) Next-generation RNA sequencing reveals differential expression of MYCN target genes and suggests the mTOR pathway as a promising therapy target in MYCN-amplified neuroblastoma. Int J Cancer 132: E106–115.
34. JohnsenJI, SegerströmL, OrregoA, ElfmanL, HenrikssonM, et al. (2008) Inhibitors of mammalian target of rapamycin downregulate MYCN protein expression and inhibit neuroblastoma growth in vitro and in vivo. Oncogene 27 : 2910–2922.
35. YuF, GaoW, YokochiT, SuenagaY, AndoK, et al. (2013) RUNX3 interacts with MYCN and facilitates protein degradation in neuroblastoma. Oncogene [epub ahead of print].
36. OttoT, HornS, BrockmannM, EilersU, SchüttrumpfL, et al. (2009) Stabilization of N-Myc is a critical function of Aurora A in human neuroblastoma. Cancer Cell 15 : 67–78.
37. BerryT, LutherW, BhatnagarN, JaminY, PoonE, et al. (2012) The ALK(F1174L) mutation potentiates the oncogenic activity of MYCN in neuroblastoma. Cancer Cell 22 : 117–130.
38. HeukampLC, ThorT, SchrammA, De PreterK, KumpsC, et al. (2012) Targeted expression of mutated ALK induces neuroblastoma in transgenic mice. Sci Transl Med 4 : 141ra191.
39. MolenaarJJ, Domingo-FernandezR, EbusME, LindnerS, KosterJ, et al. (2012) LIN28B induces neuroblastoma and enhances MYCN levels via let-7 suppression. Nat Genet 44 : 1199–1206.
40. ValentijnLJ, KosterJ, HaneveldF, AissaRA, van SluisP, et al. (2012) Functional MYCN signature predicts outcome of neuroblastoma irrespective of MYCN amplification. Proc Natl Acad Sci U S A 109 : 19190–19195.
41. LiZ, TanF, ThieleCJ (2007) Inactivation of glycogen synthase kinase-3beta contributes to brain-derived neutrophic factor/TrkB-induced resistance to chemotherapy in neuroblastoma cells. Mol Cancer Ther 6 : 3113–3121.
42. KentWJ (2002) BLAT–the BLAST-like alignment tool. Genome Res 12 : 656–664.
43. BlanchetteM, KentWJ, RiemerC, ElnitskiL, SmitAF, et al. (2004) Aligning multiple genomic sequences with the threaded blockset aligner. Genome Res 14 : 708–715.
44. AltschulSF, MaddenTL, SchafferAA, ZhangJ, ZhangZ, et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25 : 3389–3402.
45. WangD, ZhangY, ZhangZ, ZhuJ, YuJ (2010) KaKs_Calculator 2.0: a toolkit incorporating gamma-series methods and sliding window strategies. Genomics Proteomics Bioinformatics 8 : 77–80.
46. OhiraM, ObaS, NakamuraY, IsogaiE, KanekoS, et al. (2005) Expression profiling using a tumor-specific cDNA microarray predicts the prognosis of intermediate risk neuroblastomas. Cancer Cell 7 : 337–350.
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2014 Číslo 1
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- GATA6 Is a Crucial Regulator of Shh in the Limb Bud
- Large Inverted Duplications in the Human Genome Form via a Fold-Back Mechanism
- Differential Effects of Collagen Prolyl 3-Hydroxylation on Skeletal Tissues
- Affects Plant Architecture by Regulating Local Auxin Biosynthesis
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy