
Initiator Elements Function to Determine the Activity State of BX-C Enhancers
A >300 kb cis-regulatory region is required for the proper expression of the three bithorax complex (BX-C) homeotic genes. Based on genetic and transgenic analysis, a model has been proposed in which the numerous BX-C cis-regulatory elements are spatially restricted through the activation or repression of parasegment-specific chromatin domains. Particular early embryonic enhancers, called initiators, have been proposed to control this complex process. Here, in order to better understand the process of domain activation, we have undertaken a systematic in situ dissection of the iab-6 cis-regulatory domain using a new method, called InSIRT. Using this method, we create and genetically characterize mutations affecting iab-6 function, including mutations specifically modifying the iab-6 initiator. Through our mutagenesis of the iab-6 initiator, we provide strong evidence that initiators function not to directly control homeotic gene expression but rather as domain control centers to determine the activity state of the enhancers and silencers within a cis-regulatory domain.
Published in the journal:
. PLoS Genet 6(12): e32767. doi:10.1371/journal.pgen.1001260
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1001260
Summary
A >300 kb cis-regulatory region is required for the proper expression of the three bithorax complex (BX-C) homeotic genes. Based on genetic and transgenic analysis, a model has been proposed in which the numerous BX-C cis-regulatory elements are spatially restricted through the activation or repression of parasegment-specific chromatin domains. Particular early embryonic enhancers, called initiators, have been proposed to control this complex process. Here, in order to better understand the process of domain activation, we have undertaken a systematic in situ dissection of the iab-6 cis-regulatory domain using a new method, called InSIRT. Using this method, we create and genetically characterize mutations affecting iab-6 function, including mutations specifically modifying the iab-6 initiator. Through our mutagenesis of the iab-6 initiator, we provide strong evidence that initiators function not to directly control homeotic gene expression but rather as domain control centers to determine the activity state of the enhancers and silencers within a cis-regulatory domain.
Introduction
The Drosophila bithorax complex (BX-C) is one of two homeotic gene clusters in the fly and is responsible for determining the segmental identity of the posterior thoracic segment and each of the fly abdominal segments [1], [2]. It does this by using a >300 kb cis-regulatory region to control the parasegement-specific expression of the three BX-C homeotic genes: Ubx, abd-A and Abd-B (for review, see [3]).
Through the early genetic analysis of the BX-C, it was shown that its cis-regulatory sequences can be divided into nine parasegment-specific chromosomal domains (abx/bx, bxd/pbx, and iab-2 through iab-8), where each domain controls the activation of one of the three BX-C homeotic genes in a pattern appropriate for that parasegment [4]–[8]. Since their identification, these domains have been dissected using transgenic reporter assays to identify individual regulatory elements capable of modifying reporter gene expression. Among the elements identified were early embryonic enhancers (initiators), cell-type-specific enhancers, silencers and insulators [9]–[22]. Interestingly, although homeotic gene expression is restricted along the antero-postero (A–P) axis, many of the elements identified by transgenic analysis do not control reporter gene expression in an A–P restricted manner. These findings, when combined with the early genetic data suggest a model in which the cis-regulatory elements of the BX-C are controlled through the activation or repression of parasegment-specific chromatin domains [4], [23]–[24].
According to this model, the BX-C functions through multiple layers of control. First, there are the enhancers that directly activate homeotic gene expression in a pattern appropriate for a specific parasegment. Based on the genetic data, these enhancers are known to be grouped in a way where all the enhancers required to produce a PS-specific pattern of homeotic gene expression are clustered into domains within the BX-C sequence. However, although these enhancers produce a pattern of homeotic gene expression appropriate for a specific parasegment, in transgenic assays, they are not restricted along the A–P axis, and are only restricted to specific cell-types [9], [12], [22], [25].
The second layer of control comes from Polycomb-response element silencers (PREs). These silencers are thought to turn off the clusters of enhancers in parasegments where they are not needed, via modification of the local chromatin structure around the enhancers (for reviews [26]–[28]). Once again, however, like the cell-type-specific enhancers, by themselves, PREs do not seem to sense positional information and can silence genes regardless of A–P position [29].
Domain boundary elements form a third layer of BX-C control. Each of the PS-specific enhancer clusters seems to be flanked by boundary elements, required to keep each cluster separate and autonomous from other clusters. In situ, loss of a domain boundary causes the fusion of PS-specific domains, resulting in mutant phenotypes, where the affected segments displays phenotypes characteristic of the other [18], [22], [30]–[31] (for review see [32]). In transgenic assays, these elements have been shown to behave as insulators, blocking both positive and negative effects of cis-regulatory elements on reporter gene activity [13]–[14], [17]–[18], [21]. However, the presence of boundary elements cannot explain the A–P restriction of the BX-C regulatory elements, for, as with the enhancers and silencers, when taken out of the BX-C, boundary elements do not seem to have an A–P restricted activity.
So then, how do non-restricted regulatory elements control homeotic gene expression in an A–P position-dependent manner? If we are to assume that the reporter gene assays represent a reasonable estimate of the activity of the various regulatory elements in a domain, then there must be some other element in each domain that coordinates the activity of these elements across the A–P axis. Special early embryonic enhancers, called initiators, are the prime candidates to perform this function [9]–[12], [17]–[18] [20], [22]. As mentioned above, the activity of most of the elements isolated from the BX-C is not restricted along the A-P axis. In this respect, however, initiator elements are exceptional. In transgenic assays, these elements behave as early embryonic enhancers that activate reporter gene expression in a pattern along the A-P axis, consistent with the activity of the domain from which it was isolated. For example, the initiator identified from the iab-5 domain, which controls Abd-B expression in PS10/A5, activates reporter gene expression in PS10/A5 and more posterior segments in a pair-rule fashion [12]. Because similar elements were found in many PS-specific domains and these elements were the only elements discovered in the BX-C capable of reading the early parasegmental address set up by the maternal, gap and pair-rule gene products, it was hypothesized that initiators would act as the primary switches to determine if a domain was active or silenced. Unfortunately, although initiators are thought to play such a key role in BX-C gene regulation, their actual role has never been directly tested in vivo due to the lack of appropriate mutations and the difficulties in performing homologous recombination in Drosophila.
Thus, in order to explore the function of initiators and other regulatory elements in vivo, we developed a method to streamline the homologous recombination process for rapid, precise, and systematic mutagenesis. Using this method, called InSIRT (In Situ Integration for Repeated Targeting), we have created twenty new mutations in the iab-6 region of the BX-C, including mutations that directly test the role of the initiator in BX-C gene regulation (Figure 1 and Table 1).



Results/Discussion
Creation of an attP integration site in the BX-C
To study the cis-regulatory elements regulating BX-C homeotic gene expression within their natural chromosomal environment, we sought to design a method that could be used to rapidly and repeatedly target the BX-C for site-specific mutagenesis. Because this method is related to the SIRT method [33], we named it InSIRT for In Situ Integration for Repeated Targeting. Figure 2 provides a rough schematic of this method as used here in the BX-C. In short, homologous recombination is used to replace a genomic region of interest with an entry site (attP) recognized by the φC31-bacteriophage integrase [34]–[36]. Once a region of the genome is replaced by an attP site, a DNA fragment corresponding to the deleted region can be systematically mutagenized in vitro and reinserted into its normal chromosomal location by φC31 integration. As φC31 integration is a relatively fast process (by genetic standards), InSIRT allows site-specific mutagenesis of actual genes to be accomplished within the timeframe required to create a simple transgenic fly. For our experiments, we replaced a 19.3 kb region of the BX-C, roughly corresponding to the iab-6 cis-regulatory domain, with a 255bp φC31 integrase attP site (Figure 1 and Figure 2; note that the previously identified IAB5 initiator fragment [12] and the Fab-7 boundary element [30]–[31] are left intact by the deletion, while the area presumed to be the Fab-6 boundary is removed [22]).

Removal of cis-regulatory domains in the BX-C typically results in the homeotic transformations of posterior segments towards anterior segments. The segments transformed depend on the cis-regulatory domains removed and are transformed towards the last more-anterior segment whose cis-regulatory domain is intact. For example, deletion of the iab-6 cis-regulatory domain should result in the transformation of segment A6 (whose development in specified by iab-6) into A5 (whose development is controlled by iab-5). As we were attempting to delete only iab-6 in our deletion, we thus expected flies homozygous for our 19.3 kb deletion to display a typical iab-6 mutant phenotype. However, this was not the case. Flies homozygous for our 19.3 kb deletion have both their A5 and A6 segments transformed towards A4, indicating that both iab-5 and iab-6 activity are affected by our deletion (Figure 3). This can be clearly seen on the adult cuticle. Most of the segments of the adult fly abdomen can be identified independent of their position, by distinct cuticular features. For example, the wild type male sixth segment (A6) is distinguished from other segments by having a darkly pigmented tergite, covered in a distinctive pattern of tiny hairs, called trichomes, and is devoid of sternite bristles. Meanwhile, the fifth segment (A5) displays a similar darkly pigmented tergite that is uniformly covered with trichomes, and has a sternite with numerous bristles (Figure 3A). Flies homozygous for our deletion display an A4-like pigmentation pattern on both the male fifth and sixth abdominal tergites (Figure 3C). Also, the A6 sternite, normally devoid of bristles, displays numerous bristle like the A4 or A5 sternite. Based on these phenotypes, we named our deletion iab-5,6CI.

Although the adult cuticular phenotypes indicate that iab-5 function is affected in iab-5,6CI, this inactivation is incomplete and only some PS10/A5 phenotypes are affected. For example, in the embryonic CNS, the PS10/A5 Abd-B expression pattern is normal, indicating that in embryos, iab-5 is still active (compare Figure 3B with Figure 3D). Also, while iab-5 null mutants are sterile, iab-5,6CI mutants are fertile. Based on these results, we believe that iab-5,6CI removes an adult cuticle enhancer from iab-5, while leaving the rest of the iab-5 cis-regulatory domain intact. iab-6 function, as would be expected of the deletion we created, seems to be universally affected, as both the adult cuticle, and the embryonic CNS staining are affected (Figure 3C, 3D).
As a control for the InSIRT method, we first decided to reintegrate the 19.3 kb fragment removed in iab-5,6CI. As expected, the reintegrated line, iab-5,6 rescue, reverts all phenotypes associated with iab-5,6CI and demonstrates the feasibility of our approach (Figure 3E, 3F).
To begin our dissection of the cis-regulatory elements in the iab-6 domain, we created a series of overlapping deletions spanning the 19.3 kb iab-5,6CI region (Figure 1 and Table 1) and examined their resulting phenotypes on the adult cuticle and embryonic CNS. We will first discuss deletions affecting the iab-6 initiator.
The iab-6 initiator is necessary, but not sufficient to control Abd-B expression in PS11/A6
Previously, we identified a 2.8 kb element from iab-6 that displayed the characteristics of an initiator in a transgenic reporter assay. Accordingly, this 2.8 kb fragment was shown to be able to drive the early expression of a lacZ reporter in a spatially restricted, pair-rule pattern, from PS11/A6 [22]. Unfortunately, as with other initiators, its precise function in the BX-C was never investigated genetically. We thus created a mutation that removes this 2.8 kb initiator fragment from the iab-6 domain. The resulting mutant is named, iab-61 (see Figure 1B). According to the domain model, initiators should act as switches to turn on all of the enhancers in a domain. Thus, removal of the initiator from iab-6 should result in a complete loss of iab-6 function. This is, in fact, what we see. In flies homozygous for this deletion, A6 seems to be completely transformed into A5 (Figure 4A). This can be clearly seen on the adult cuticle where the 6th sternite takes on the shape and bristle pattern characteristic of A5. The transformation can also be seen in the embryonic CNS, where the Abd-B expression pattern in PS11/A6 is replaced by the pattern normally found in PS10/A5 (Figure 4C).

In as much as the 2.8kb fragment had not been further dissected in the reporter gene assay, we decided to further narrow down the initiator by integrating smaller deletions from within the 2.8 kb fragment. In order to select these deletions, we first applied a bioinformatic approach (fly_Ahab: http://gaspard.bio.nyu.edu/fly_ahab.html) to identify regions in the iab-6 domain that correspond to binding sites for known Drosophila transcription factors. Using this approach, we identified a potential cis-regulatory module in an ∼900 bp region of the iab-6 initiator. This module contains the predicted binding sites for the Kruppel, Caudal and Hunchback proteins, and is supported by recent genome-wide ChIP analysis (http://genome.ucsc.edu). As previous work had indicated that initiators sense A-P position through the binding of the maternal, gap and pair-rule gene products [10], [12], [17], [20], [37]–[41], we decided to test if this ∼900 bp region is required for iab-6 function in the BX-C. To do this, we created three overlapping deletions, iab-62, iab-63, and iab-64, that each deletes this 900 bp region (Figure 1B). As flies homozygous for any of these three mutations display the same phenotype, we will concentrate on the smallest of these deletions, iab-64, which deletes only a 927 bp sequence corresponding to the identified cis-regulatory module. Flies homozygous for iab-64 show a complete loss of iab-6 function, like that seen in iab-61 flies. The extent of the transformation is corroborated by the tergite trichome pattern (Figure 4E) and the embryonic Abd-B expression pattern in the CNS, where the PS11/A6-specific pattern is replaced by the pattern normally found in PS10/A5 (Figure 4F). Based on these results, we conclude that this 927 bp fragment is absolutely necessary for iab-6 activation of Abd-B in PS11/A6. The fact that a deletion of the initiator is capable of completely removing iab-6 activity in the epidermis and the CNS, is consistent with the idea that the initiator functions as a switch to turn on all of the regulatory elements in a cis-regulatory domain. However, these results would also be consistent with the initiator being the sole positive regulatory element in A6/PS11. To rule out this possibility, we created another mutation that removes much of the iab-6 cis-regulatory region but leaves intact the 2.8 kb initiator fragment. This mutation, called iab-68 (Figure 1), also shows a strong loss of iab-6 function (Figure 4G). Thus, although the iab-6 initiator is critical for iab-6 function, it is not sufficient for iab-6 activity.
One important point to note regarding iab-68 is that the loss-of-function (LOF) phenotype is slightly weaker than in iab-64. The difference between these two mutants can be seen when examining the trichome pattern on the transformed A6. While the iab-64 mutation displays a transformed A6 with an A5-like trichome pattern (uniformly covered, Figure 4E), the iab-68 trichome pattern still resembles that of a wild-type A6 (Figure 4H). This suggests that although iab-68 removes most of the iab-6 sequence, there is still some functionality left in the domain. This is an important finding because it supports a prediction of the domain model, which suggests that removal of cell-type specific enhancers would affect individual (or grouped) characteristic, while removal of initiator elements would affect all characteristics. To test this idea more directly, we next performed an initiator swapping experiment.
Initiators act as domain control centers for cis-regulatory domains
The domain model suggests that initiators act as switches to turn on (or off) the various regulatory elements present in a domain. In the simplest state, this would mean that initiators would not participate directly in driving homeotic gene expression, but would simply coordinate the activity of cell/tissue-type specific enhancers along the A-P axis. If this were true, then we hypothesized that we should be able to transform a segment into another, simply by turning on the cell/tissue-type specific enhancers of domain in an area where they are normally off. Using our InSIRT method, we could do this by exchanging initiators from different domains.
For these experiments, we chose to remove the 927 bp iab-6 initiator and replace it with the molecularly identified initiator from iab-5. The iab-5 initiator is defined as a 1 kb DNA fragment (called IAB5) that, when cloned in front of the Ubx-lacZ reporter gene, activates strong β-galactosidase in a pair-rule fashion from PS10 [12]. Therefore, if the domain model is correct, by replacing the iab-6 initiator with that of iab-5, we should be able to activate the enhancers required for PS11/A6 development in PS10/A5. As the difference in the expression pattern of Abd-B between PS10 and PS11 can be summarized as PS11 having a higher level of Abd-B expression than PS10 (and in more cells), we would expect ectopic activation of iab-6 to be epistatic to the activity of iab-5. In other words, we expected to see a posterior transformation of A5 into A6.
As predicted by the domain model, the swapping of the iab-6 initiator with that of iab-5 results in a dominant A5 to A6 transformation that is stronger in homozygous flies. This type of posterior-directed abdominal transformation in the BX-C is typically called a Frontabdominal (Fab) transformation. For this reason, we have named this new mutation, Fab-6IAB5. Figure 5C shows an abdominal cuticle of a Fab-6IAB5 homozygous male. In this fly, we observe an A6-like lack of A5 sternite bristles, an A6-like trichome pattern on the A5 tergite and a PS11/A6-like Abd-B pattern of expression in PS10/A5 (Figure 5F). The fact that A5 appears to be a copy of A6 suggests that everything required for the patterning of A6 is still present in the modified iab-6 domain, but that these elements have simply been activated one segment too-anterior. These findings strongly support the model in which initiators function, not as enhancers to directly control homeotic gene expression, but rather as domain control centers to turn on the other cis-regulatory elements in a domain.

Interestingly, in the Fab-6IAB5 mutation, A6 seems to be unaffected by the IAB5 swap. As mentioned above, the IAB5 fragment drives reporter gene expression in a pair-rule manner from PS10/A5 (i.e. not in PS11) [12]. In fact, reporter transgenes carrying the entire iab-5 domain are still only capable of driving reporter gene expression in a pair-rule fashion [22]. If this assay reflects IAB5 activity in vivo, then what turns on iab-6 in PS11/A6? Currently, we do not have a completely satisfying answer to this question. However, previous genetic studies tell us that the iab-5 cis-regulatory domain is indeed capable of working in PS11/A6. In iab-6 mutants, for example, A6 is transformed into A5 [22], [42]. Thus, it seems clear that our interpretation of IAB5 activity from the transgenic reporter assay, oversimplifies IAB5 function. This is perhaps not too surprising, as the reporter gene assays were designed to simply test if a DNA fragment is capable acting as an enhancer. From our experiments, however, it seems that initiators may have a more complex function that is not reflected in the transgene assay.
Mutations affecting Fab-6 boundary function
During our dissection of the iab-6 domain, we also created a number of deletions affecting Fab-6 boundary function. Boundaries function to keep domains autonomous. Based on what was observed in other boundary deletions, we know that boundaries prevent both ectopic activation of posterior domains by elements present in anterior domains (initiators), as well as, prevent anterior domains from being silenced by posterior silencing elements (PREs) [18], [31]. The removal of a boundary element, therefore, results in a mixed transformation, where clones of cells in anterior segments become transformed towards cells of more-posterior segments, and clones of cells from posterior segments become transformed towards cells of more-anterior segments. Using our series of deletions, we have narrowed down the Fab-6 boundary to an ∼650 bp region of the BX-C.
Previous work from our lab [22] genetically mapped the Fab-6 boundary to an ∼4.5 kb region of the BX-C between the distal (relative to the Abd-B transcription unit) breakpoints of the iab-6IH (3R:12712604) and Fab-6,71 (3R:12708067) mutations. By using deletions nested on the Abd-B-distal side of the iab-5,6CI mutation, we were able to quickly narrow down the location of the Fab-6 boundary further. The first deletion we will speak about, iab-5Δ1, removes an ∼1.2 kb region from the distal side of iab-5,6CI but displays no visible phenotype. This indicates that neither the iab-5 cuticle enhancer (see above), nor the Fab-6 boundary element is removed by this deletion. On the other hand, two bigger deletions, removing ∼3.2 kb and ∼8 kb respectively (Fab-61 and Fab-62), show mixed anteriorizing (LOF) and posteriorizing (GOF) transformations of A5 and A6 towards A4 or A6. An example of this (Fab-62) can be seen in Figure 6A where we see a loss of bristles on the A5 sternite (indicative of a posterior-directed transformation of A5 to A6), and a loss of pigmentation on the A5 tergite (indicative of an anterior-directed transformation of A5 to A4). Meanwhile, in the A6 segment of each mutant, we see a gain of bristles on the sternite and a loss of pigmentation on the tergite, both indicative of an anterior-directed transformation (probably towards A4). Although very similar, we must note that the Fab-62 mutation displays a slightly stronger GOF phenotype than Fab-61 (data not shown). Consistent with this finding, a PRE has recently been mapped to the region differentiating the Fab-61 and Fab-62 mutations [43]. Thus, if Fab-62 functions like other boundary mutations, the enhanced Fab-6 GOF phenotype is probably caused by deleting this silencing element and shifting the balance between GOF and LOF phenotypes.
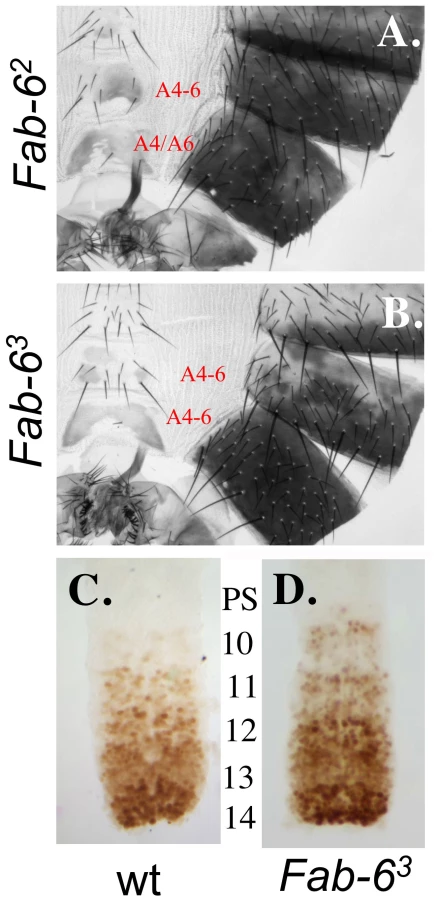
The smallest mutation we created that displays an Fab-6 phenotype is Fab-63 which deletes an ∼2 kb span of DNA between the proximal breakpoints of iab-5Δ1 (3R:12706585) and Fab-61 (3R:12708661). As expected Fab-63 displays a phenotype similar to Fab-61 and Fab-62 (Figure 6B) with mixed gain - and loss-of-function phenotypes. Meanwhile, a deletion spanning the PRE region that differs between Fab-61 and Fab-62 shows no phenotype (iab-6Δ7, Figure 1), suggesting that the critical elements required for Fab-6 function are all contained in Fab-63. Thus, when combining our new data with those of the past, we can now narrow down the Fab-6 boundary to an ∼650 bp region, spanning from the proximal breakpoint of Fab-6,71 (3R:12708067) to the distal breakpoint of iab-6Δ7 (3R:12708714). Consistent with this mapping, it has recently been shown that this region contains binding sites for the insulator protein dCTCF [44], and that a fragment containing these dCTCF binding sites blocks enhancer-promoter interactions in an insulator reporter assay [45].
Additional mutations in the iab-6 region
Other deletions created for this study are presented here, solely for the purpose of completeness. These additional deletions are depicted in Figure 1 and their phenotypes are summarized in Table 1. Although we will not discuss these mutants in detail, we would like to point out two key issues surrounding these mutations. First, all deletions removing the initiator fragment show a loss of iab-6 function comparable to that seen in iab-64. This finding is in agreement with the domain model and our data, which suggests that the initiator is absolutely required for activating iab-6 function in the cuticle and CNS. Second, we have also isolated a number of mutations that show no noticeable iab-6 phenotype. These mutations, iab-6Δ7, iab-6Δ5 and iab-6Δ6, remove a total of ∼9.8 kb of iab-6 sequence without dramatically changing the morphology of the adult cuticle or modifying the Abd-B expression pattern in the embryonic CNS (data not shown). Does this mean that these sequences are without function? Absolutely not. In fact, we know that the region deleted in iab-6Δ7 probably contains a PRE, whose deletion produces a phenotype when combined with the deletion of the Fab-6 boundary. Also, as we have scanned only a small fraction of the possible developmental pathways in which Abd-B is involved, we believe that it is very likely that these other regions contain cell-type specific enhancers controlling Abd-B expression in other tissues than the CNS and the cuticle. Obviously, now that we have the ability to manipulate regions of the BX-C at a base-pair level, we now require equally precise methods to monitor potential phenotypic changes.
The initiator and a hierarchical structure to gene regulation
For more than twenty years, much of the work on the BX-C has proceeded on the assumption that the BX-C cis-regulatory regions control homeotic gene expression through a multilayered, hierarchical process, summarized in the domain model. Key to this model was the idea that there existed specialized switch elements to control the activity state of the entire domain. Based on transgenic assays, these switch elements were thought to be special early embryonic enhancers, often called initiators. Although through the years, we, and others, have generated many results consistent with this model, we were never able to directly test initiator function in situ, due to experimental difficulties. Thus, a key prediction of the domain model went untested for decades. Here, we have finally provided the data confirming the key role of the initiator in domain activation.
Besides being important for studies on BX-C gene regulation, our findings highlight the possibility of having elements whose sole function may be to control the activity state of other elements. Although we still do not understand how this is accomplished mechanistically, we believe that it is probably through modifying the local chromatin environment around the enhancers. In fact, taking into account the enhancer activity of initiators in transgenic constructs, we are left with an intriguing and testable model for initiator action. As mentioned above, initiators were first isolated as early embryonic enhancers that turned on reporter gene expression in an A-P restricted manner. It has been known for years that the cis-regulatory sequences of the BX-C are transcribed in a parasegment-specific manner where transcripts from each cis-regulatory domain are expressed along the A-P axis in correspondence to where a domain is expected to be active [46]–[49]. In other experiments it has been shown that forced transcription across PREs in the BX-C can prevent Pc-dependent silencing, and hence, activate a domain [50]–[52]. Thus, by combining these findings, it is possible to imagine a causal relationship between the initiator and transcription, and the transcription of a domain and domain activation. Accordingly, we propose that initiators might act as enhancers, responding to gap and pair-rule gene products to activate transcription from promoters within the cis-regulatory domains. In doing so, they would indirectly activate homeotic gene expression by preventing the Pc silencing of homeotic gene enhancers. Using the tools developed here, we are now in the process of testing this model.
Another question that we can now address using InSIRT is whether or not initiators are required later in development. Thus far, we have been discussing initiators as only functioning early in development. It is still possible, however, that initiators are constantly required for domain activation or that they play a later role in the regulation of homeotic gene expression. Although the initiator being constantly required to keep a domain active cannot be ruled out, based on our current understanding of initiator function, we do not believe this to be the case. Perhaps the strongest evidence supporting this belief comes from transgenic assays. In transgenic assays, initiator fragments seem to respond to the maternal, gap and pair-rule gene products. Upon disappearance of the early expression pattern of these proteins, initiator activity in transgenes often disappears, or produces a pattern of expression not restricted along the A-P axis (if not paired with a PRE/maintenance element). Based on this, we believe that their role in coordinating the activity of a domain is probably limited to early development. However, this does not mean that all initiators would have no activity outside of initiation phase, only that their function in domain initiation would be limited to early embryogenesis. As mentioned above, the pattern of reporter gene expression driven by some initiator fragments degenerates into a cell-type specific enhancer-like pattern later in development. Therefore, it may be possible that DNA fragments with initiator function may also contain cell-type specific enhancers.
InSIRT as a method to study gene expression
The transgenic reporter assay has played an important role in shaping our understanding of eukaryotic gene expression [53]. Its advantages stem from its speed and the cleanliness of the approach in isolating cis-regulatory elements away from competing or obfuscating signals. To gain these advantages, reporter assays must make a number of critical assumptions. First, they must assume that an activity performed by an element in the reporter assay, is the activity performed by the element in vivo. Second, they must assume that critical transcriptional activities can be tested using the molecular construct devised. And third, they must assume that it is through the addition of these cis-regulatory activities that controlled gene expression is achieved. However, these assumptions are not always correct. Although in the study of the BX-C, transgenes have been extremely useful in estimating the activity of elements, our work on the initiator and our previous work on boundary elements [54] highlight how sometimes the activity seen in transgenes assays only represents a portion of an element's activity in vivo.
InSIRT is a complementary approach. Relative to the transgenic approach, InSIRT has one main advantage: it tests for changes on biologically relevant targets without necessarily simplifying or assuming an activity. This is key when trying to understand unusual regulatory elements, like initiators. Furthermore, this advantage can be achieved with only a small penalty in time, as, once the homologous recombination has been performed, InSIRT mutagenesis takes only as much time as establishing a single transgenic line. Because transgenic approaches often require the analysis of multiple lines to control for genomic position effects, this penalty is further reduced. Thus, we believe that InSIRT offers scientists a powerful new tool that can be used in combination with classical transgenic methods to better study gene regulation.
Materials and Methods
Fly methods and phenotypes
All crosses, and cuticle preparations were performed using standard Drosophila methods. Abd-B antibody staining was performed as in [22]. Abd-B monoclonal antibody was purchased from the “Developmental Studies Hybridoma Bank” at the University of Iowa. Injection experiments were performed using cleaned DNA preparations (Qiagen) and injected into the iab-5,6CI flies stocks containing an X chromosome expressing the φC31 integrase enzyme under the control of the vasa promoter [36] see http://www.frontiers-in-genetics.org/flyc31/).
Phenotypes depicted are representative of the genotypes shown. As some of the boundary phenotypes seem to be clonal in nature, there is an occasional variance in the exact number of bristles and the exact pattern of trichomes. We have, therefore, attempted to choose an average representative cuticle for display. Otherwise, the phenotypes can be considered 100% penetrant.
Homologous recombination
Creation of a donor vector for homologous recombination: An AscI-NotI fragment containing the yellow reporter gene flanked by the two loxP sites, and a 255 bp attP integration site was cloned into pW25 digested with AscI-NotI to create the pY25 plasmid. Homology regions of ∼4 kb were created by PCR using the following primer pairs: IAB7-AvrII: CCTAGGCGGCGAACAGTAGGGAAG and Fab7-AscI: CAGC-AAAAATCGTAAAAAAG, and IAB5-NotI: GCGGCCGCGGTCAGTAAACGG-GTCCC and IAB5-SpHI: GCATGCACTGGCGACATTTCTC. These homology regions were then cloned into the pY25 vector in the AvrII and AscI sites or the NotI and SphI sites respectively. The resulting P-element vector, Py-del iab-6, was injected into yw flies and transformants were isolated as yellow+ flies. Homologous recombination was performed using two independent transformants and the ends-out homologous recombination method of Gong and Golic [34].
Potential homologous recombinants were isolated based on their yellow pigmentation limited to the segments posterior and including A5 (as a result of being in iab-5). Genomic southern blots, however showed that all identified events were aborted recombination events in the BX-C. As aborted events happened on each side of the targeted region, we were able to generate the final “planned” deletion by recombining two chromosomes each having recombined properly at one homology region. This recombination was mediated by the Cre recombinase at the loxP sites left behind after removal of the yellow reporter gene. The final chromosome, iab-5,6CI, was verified by genomic southern blot and sequencing.
Generation of integration vectors
A base vector containing the 19.3 kb area deleted in the iab-5,6CI deletion, a 288 bp attB sequence, a yellow reporter, and one loxP site (called KsY-iab6H) was generated using gap-repair recombineering ([55], [56] see Figure 1B). For this, ∼500 bp PCR products were generated to both the iab-5 and iab-6 regions (each starting at the breakpoints of the iab-5,6CI deletion) using the following primers:
Iab-5 N: ATAAGAATGCGGCCGCGGTGCGTTTCCATTT-TCCCTAGG
Iab-5 new (+PmeI): CTCACCATAGAGCACCACGTTTAAACGTCGT-CCGGAAATGGCAACCAG
Iab-6: CTTTGCCAGCTTTTGCCACTCGTCC
Iab-6P(+PmeI): GGTTGCCATTTCCGGACGACGTTTAAACGGTG-AAGGCGCG-AAACTGTGA.
These two products were linked using overlap PCR. This 1 kb fragment was then cloned into a vector containing a 288 bp attB sequence, a yellow reporter, and one loxP site (in that order), resulting in the plasmid, Ks-Y attB-loxP.
The Ks-Y attB-loxP plasmid was digested with PmeI and used in recombineering experiments to capture the 19.3 kb region deleted in the iab-5,6CI deletion. All recombineering procedures were performed as in ([55], [56]. Basically, the digested plasmid was transformed by electroporation into heat-shock induced competent cells of the EL350 recombineering bacterial strain that was previously transformed with the Abd-B region-containing BAC, BACR24L1 (BACR24L18, GenBank: AC095018). The resulting plasmid is called KsY-iab6H.
The KsY-iab6H was then modified using recombineering. For the initiator deletions, we generated PCR fragments containing a Kanamycin selector gene surrounded by two FRT sites, using primers containing 5′ leaders with 50 bp of homology to the region flanking the sequences to be deleted. The FRT-kanamycin-FRT DNA template for PCR came from pGEM1-K7-FRT-Kan-FRT (kindly provided by François Spitz). For the modification of KsY-iab6H by recombineering, PCR targeting fragments (containing an FRT-Kan-FRT cassette) were generated using the primers listed in Table 2.
For the recombineering of the iab-6/iab-5 initiator swap, the iab-5 initiator was amplified by PCR using the following primers:
s-sub-iab5 5′ATGGCGCGCCGGAGGCGGCAAATGCACAAAG3′
as-sub-iab5 5′ATGGCGCGCCTACTACGCCGATTCTGCTGG3′
Two fragments (of 1.7 kb and 1.3 kb, respectively), each homologous to one side of the iab-6 initiator were amplified by PCR using the following primers:
sub1-AscI: 5′ATGGCGCGCCCAGATTTCTGGAATGGTTAGAAAAT-ATTAAAGG3′
sub2-NotI: 5′ACTCGCGGCCGCTCGGAAACATCAAAGCATCAGCA-AC3′,
sub3-AscI: 5′ATGGCGCGCCGCAGTAAGTTAATATATTTTATAC-TCC3′
sub4-NotI: 5′ACTCGCGGCCGCAGAGAAATATATTCTTTGGCAG-CGAGC3′.
The IAB5 initiator was cloned between these regions of homology using the AscI restriction site to create the vector, Target iab5(+). An FRT-Kan-FRT cassette was amplified by PCR using the following primers:
5′ SnaBI: 5′ACATGGAAAACAACAGTTTCAATCAGGTCATGTAC-CTAATAAATGTATACGAATACAAGCTTGGGCTGCAGG3′
3′ SnaBI 5′AGCTTACATTTTGATAGCTTAAGTGGATGTTTCAAGGA-ATTTATATATACCTCGCCCGGGGATCCTCTAGAG3′
This cassette was then cloned into a unique SnaBI site within the 1.7 kb homology domain (245 bp from the IAB5 initiator) of Target iab5(+), to make Target iab5(+) Kan FRT. A NotI fragment containing the two homology regions, the IAB5 initiator and the Kan-FRT cassette was then used to recombineer the iab-5 swap integration vector. The recombineering was otherwise performed as above.
Upon recombineering on the KsI-iab6H plasmid, the plasmids carrying the designed deletion were selected on Kan plates. The Kanamycin cassette was then removed using a bacterial strain expressing the flipase enzyme under an inducible arabinose promoter (EL 250; [55].
Zdroje
1. LewisEB
1978 A gene complex controlling segmentation in Drosophila. Nature 276 565 570
2. Sanchez-HerreroE
VernosI
MarcoR
MorataG
1985 Genetic organization of Drosophila bithorax complex. Nature 313 108 113
3. MaedaRK
KarchF
2006 The ABC of the BX-C: the bithorax complex explained. Development 133 1413 1422
4. PeiferM
KarchF
BenderW
1987 The bithorax complex: control of segmental identity. Genes Dev 1 891 898
5. Sanchez-HerreroE
1991 Control of the expression of the bithorax complex genes abdominal-A and abdominal-B by cis-regulatory regions in Drosophila embryos. Development 111 437 449
6. CelnikerSE
SharmaS
KeelanDJ
LewisEB
1990 The molecular genetics of the bithorax complex of Drosophila: cis - regulation in the Abdominal-B domain. Embo J 9 4277 4286
7. KarchF
WeiffenbachB
PeiferM
BenderW
DuncanI
1985 The abdominal region of the bithorax complex. Cell 43 81 96
8. BenderW
AkamM
KarchF
BeachyPA
PeiferM
1983 Molecular genetics of the bithorax complex in Drosophila melanogaster. Science 221 23 29
9. SimonJ
PeiferM
BenderW
O'ConnorM
1990 Regulatory elements of the bithorax complex that control expression along the anterior-posterior axis. Embo J 9 3945 3956 Issn: 0261-4189
10. QianS
CapovillaM
PirrottaV
1991 The bx region enhancer, a distant cis-control element of the Drosophila Ubx gene and its regulation by hunchback and other segmentation genes. Embo J 10 1415 1425
11. MullerJ
BienzM
1992 Sharp anterior boundary of homeotic gene expression conferred by the fushi tarazu protein. Embo J 11 3653 3661 Issn: 0261-4189
12. BusturiaA
BienzM
1993 Silencers in abdominal-B, a homeotic Drosophila gene. Embo J 12 1415 1425 Issn: 0261-4189
13. ZhouJ
BaroloS
SzymanskiP
LevineM
1996 The Fab-7 element of the bithorax complex attenuates enhancer-promoter interactions in the Drosophila embryo. Genes Dev 10 3195 3201
14. HagstromK
MullerM
SchedlP
1996 Fab-7 functions as a chromatin domain boundary to ensure proper segment specification by the Drosophila bithorax complex. Genes Dev 10 3202 3215
15. HagstromK
MullerM
SchedlP
1997 A Polycomb and GAGA dependent silencer adjoins the Fab-7 boundary in the Drosophila bithorax comple. Genetics 146 1365 1380
16. FritschC
BrownJL
KassisJA
MullerJ
1999 The DNA-binding Polycomb group protein Pleiohomeotic mediates silencing of a Drosophila homeotic gene. Development 126 3905 3913
17. ZhouJ
AsheH
BurksC
LevineM
1999 Characterization of the transvection mediating region of the abdominal - B locus in Drosophila. Development 126 3057 3065
18. BargesS
MihalyJ
GalloniM
HagstromK
MüllerM
2000 The Fab-8 boundary defines the distal limit of the bithorax complex iab-7 domain and insulates iab-7 from initiation elements and a PRE in the adjacent iab-8 domain. Development 127 779 790
19. HorardB
TatoutC
PouxS
PirrottaV
2000 Structure of a polycomb response element and in vitro binding of polycomb group complexes containing GAGA factor. Mol Cell Biol 20 3187 3197
20. ShimellMJ
PetersonAJ
BurrJ
SimonJA
O'ConnorMB
2000 Functional analysis of repressor binding sites in the iab-2 regulatory region of the abdominal-A homeotic gene. Dev Biol 218 38 52
21. GruzdevaN
KyrchanovaO
ParshikovA
KullyevA
GeorgievP
2005 The Mcp Element from the bithorax Complex Contains an Insulator That Is Capable of Pairwise Interactions and Can Facilitate Enhancer-Promoter Communication. Mol Cell Biol 25 3682 3689
22. MihalyJ
BargesS
SiposL
MaedaR
CleardF
2006 Dissecting the regulatory landscape of the Abd-B gene of the bithorax complex. Development 133 2983 2993
23. MaedaRK
KarchF
2009 The bithorax complex of Drosophila an exceptional Hox cluster. Curr Top Dev Biol 88 1 33
24. MaedaRK
KarchF
2010 Cis-regulation in the Drosophila bithorax Complex. Adv Exp Med Biol 689 17 40
25. PirrottaV
ChanCS
McCabeD
QianS
1995 Distinct parasegmental and imaginal enhancers and the establishment of the expression pattern of the Ubx gene. Genetics 141 1439 1450
26. MullerJ
KassisJA
2006 Polycomb response elements and targeting of Polycomb group proteins in Drosophila. Curr Opin Genet Dev 16 476 484
27. RingroseL
ParoR
2007 Polycomb/Trithorax response elements and epigenetic memory of cell identity. Development 134 223 232
28. SchwartzYB
PirrottaV
2007 Polycomb silencing mechanisms and the management of genomic programmes. Nat Rev Genet 8 9 22
29. ChiangA
O'ConnorMB
ParoR
SimonJ
BenderW
1995 Discrete Polycomb-binding sites in each parasegmental domain of the bithorax complex. Development 121 1681 1689 Issn: 0950-1991
30. GyurkovicsH
GauszJ
KummerJ
KarchF
1990 A new homeotic mutation in the Drosophila bithorax complex removes a boundary separating two domains of regulation. Embo J 9 2579 2585 Issn 0261-4189
31. MihalyJ
HoggaI
GauszJ
GyurkovicsH
KarchF
1997 In situ dissection of the Fab-7 region of the bithorax complex into a chromatin domain boundary and a Polycomb-response element. Development 124 1809 1820
32. MaedaRK
KarchF
2007 Making connections: boundaries and insulators in Drosophila. Curr Opin Genet Dev 17 394 399
33. GaoG
McMahonC
ChenJ
RongYS
2008 A powerful method combining homologous recombination and site-specific recombination for targeted mutagenesis in Drosophila. Proc Natl Acad Sci U S A 105 13999 14004
34. GongWJ
GolicKG
2003 Ends-out, or replacement, gene targeting in Drosophila. Proc Natl Acad Sci U S A 100 2556 2561
35. GrothAC
CalosMP
2004 Phage integrases: biology and applications. J Mol Biol 335 667 678
36. BischofJ
MaedaRK
HedigerM
KarchF
BaslerK
2007 An optimized transgenesis system for Drosophila using germ-line-specific φC31 integrases. Proc Natl Acad Sci U S A 104 3312 3317
37. WhiteRA
LehmannR
1986 A gap gene, hunchback, regulates the spatial expression of Ultrabithorax. Cell 47 311 321
38. IrishVF
Martinez-AriasA
AkamM
1989 Spatial regulation of the Antennapedia and Ultrabithorax homeotic genes during Drosophila early development. Embo J 8 1527 1537
39. ZhangCC
MullerJ
HochM
JackleH
BienzM
1991 Target sequences for hunchback in a control region conferring Ultrabithorax expression boundaries. Development 113 1171 1179 Issn: 0950-1991
40. ShimellMJ
SimonJ
BenderW
O'ConnorMB
1994 Enhancer point mutation results in a homeotic transformation in Drosophila. Science 264 968 971 Issn: 0036-8075
41. CasaresF
Sanchez HerreroE
1995 Regulation of the infraabdominal regions of the bithorax complex of Drosophila by gap genes. Development 121 1855 1866 Issn: 0950-1991
42. CrosbyMA
LundquistEA
TautvydasRM
JohnsonJJ
1993 The 3′ regulatory region of the Abdominal-B gene: genetic analysis supports a model of reiterated and interchangeable regulatory elements. Genetics 134 809 824 Issn: 0016-6731
43. Perez-LluchS
CuarteroS
AzorinF
EspinasML
2008 Characterization of new regulatory elements within the Drosophila bithorax complex. Nucleic Acids Res 36 6926 6933
44. HolohanEE
KwongC
AdryanB
BartkuhnM
HeroldM
2007 CTCF genomic binding sites in Drosophila and the organisation of the bithorax complex. PLoS Genet 3 e112 doi:10.1371/journal.pgen.0030112
45. SmithST
WickramasingheP
OlsonA
LoukinovD
LinL
2009 Genome wide ChIP-chip analyses reveal important roles for CTCF in Drosophila genome organization. Dev Biol 328 518 528
46. BaeE
CalhounVC
LevineM
LewisEB
DrewellRA
2002 Characterization of the intergenic RNA profile at abdominal-A and Abdominal-B in the Drosophila bithorax complex. Proc Natl Acad Sci U S A 99 16847 16852
47. CumberledgeS
ZaratzianA
SakonjuS
1990 Characterization of two RNAs transcribed from the cis-regulatory region of the abd-A domain within the Drosophila bithorax complex. Proc Natl Acad Sci U S A 87 3259 3263
48. LipshitzHD
PeattieDA
HognessDS
1987 Novel transcripts from the Ultrabithorax domain of the bithorax complex. Genes Dev 1 307 322
49. Sanchez-HerreroE
AkamM
1989 Spatially ordered transcription of regulatory DNA in the bithorax complex of Drosophila. Development 107 321 329
50. BenderW
FitzgeraldDP
2002 Transcription activates repressed domains in the Drosophila bithorax complex. Development 129 4923 4930
51. HoggaI
KarchF
2002 Transcription through the iab-7 cis-regulatory domain of the bithorax complex interferes with maintenance of Polycomb-mediated silencing. Development 129 4915 4922
52. SchmittS
PrestelM
ParoR
2005 Intergenic transcription through a polycomb group response element counteracts silencing. Genes Dev 19 697 708
53. SmallS
KrautR
HoeyT
WarriorR
LevineM
1991 Transcriptional regulation of a pair-rule stripe in Drosophila. Genes Dev 5 827 839
54. CleardF
MoshkinY
KarchF
MaedaRK
2006 Probing long-distance regulatory interactions in the Drosophila melanogaster bithorax complex using Dam identification. Nat Genet 38 931 935
55. LeeEC
YuD
Martinez de VelascoJ
TessarolloL
SwingDA
2001 A highly efficient Escherichia coli-based chromosome engineering system adapted for recombinogenic targeting and subcloning of BAC DNA. Genomics 73 56 65
56. LiuP
JenkinsNA
CopelandNG
2003 A highly efficient recombineering-based method for generating conditional knockout mutations. Genome Res 13 476 484
57. LewisEB
1985 Regulation of the genes of the bithorax complex in Drosophila. Cold Spring Harb Symp Quant Biol 50 155 164
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2010 Číslo 12
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Functional Comparison of Innate Immune Signaling Pathways in Primates
- Expression of Linear and Novel Circular Forms of an -Associated Non-Coding RNA Correlates with Atherosclerosis Risk
- Genome-Wide Interrogation of Mammalian Stem Cell Fate Determinants by Nested Chromosome Deletions
- Histone H2A C-Terminus Regulates Chromatin Dynamics, Remodeling, and Histone H1 Binding
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy