
The Functions of Mediator in Support a Role in Shaping Species-Specific Gene Expression
The Mediator complex is an essential co-regulator of RNA polymerase II that is conserved throughout eukaryotes. Here we present the first study of Mediator in the pathogenic fungus Candida albicans. We focused on the Middle domain subunit Med31, the Head domain subunit Med20, and Srb9/Med13 from the Kinase domain. The C. albicans Mediator shares some roles with model yeasts Saccharomyces cerevisiae and Schizosaccharomyces pombe, such as functions in the response to certain stresses and the role of Med31 in the expression of genes regulated by the activator Ace2. The C. albicans Mediator also has additional roles in the transcription of genes associated with virulence, for example genes related to morphogenesis and gene families enriched in pathogens, such as the ALS adhesins. Consistently, Med31, Med20, and Srb9/Med13 contribute to key virulence attributes of C. albicans, filamentation, and biofilm formation; and ALS1 is a biologically relevant target of Med31 for development of biofilms. Furthermore, Med31 affects virulence of C. albicans in the worm infection model. We present evidence that the roles of Med31 and Srb9/Med13 in the expression of the genes encoding cell wall adhesins are different between S. cerevisiae and C. albicans: they are repressors of the FLO genes in S. cerevisiae and are activators of the ALS genes in C. albicans. This suggests that Mediator subunits regulate adhesion in a distinct manner between these two distantly related fungal species.
Published in the journal:
. PLoS Genet 8(4): e32767. doi:10.1371/journal.pgen.1002613
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002613
Summary
The Mediator complex is an essential co-regulator of RNA polymerase II that is conserved throughout eukaryotes. Here we present the first study of Mediator in the pathogenic fungus Candida albicans. We focused on the Middle domain subunit Med31, the Head domain subunit Med20, and Srb9/Med13 from the Kinase domain. The C. albicans Mediator shares some roles with model yeasts Saccharomyces cerevisiae and Schizosaccharomyces pombe, such as functions in the response to certain stresses and the role of Med31 in the expression of genes regulated by the activator Ace2. The C. albicans Mediator also has additional roles in the transcription of genes associated with virulence, for example genes related to morphogenesis and gene families enriched in pathogens, such as the ALS adhesins. Consistently, Med31, Med20, and Srb9/Med13 contribute to key virulence attributes of C. albicans, filamentation, and biofilm formation; and ALS1 is a biologically relevant target of Med31 for development of biofilms. Furthermore, Med31 affects virulence of C. albicans in the worm infection model. We present evidence that the roles of Med31 and Srb9/Med13 in the expression of the genes encoding cell wall adhesins are different between S. cerevisiae and C. albicans: they are repressors of the FLO genes in S. cerevisiae and are activators of the ALS genes in C. albicans. This suggests that Mediator subunits regulate adhesion in a distinct manner between these two distantly related fungal species.
Introduction
The transcription factor complex Mediator is associated with RNA polymerase II and it has essential roles in transcription ([1], reviewed in [2]). The yeast Mediator is composed of 25 subunits, which are structurally and functionally organized into four modules [3]–[8]. The core complex is comprised of the Head, Middle and Tail domains [3]–[6]. A fourth, Kinase domain is associated with Mediator under some conditions ([9]–[11]; reviewed in [2]). The core Mediator has a positive role in transcription, while the Kinase domain mainly functions in repression [2].
The roles of Mediator in transcription are complex [2], [12]. Mediator interacts with gene-specific transcription factors and RNA polymerase II and mediates polymerase-activator interactions and formation of the pre-initiation complex (reviewed in [2], [12], [13]). In addition to activated transcription, Mediator also stimulates basal transcription [1], [14], [15]. Further proposed roles for Mediator are in post-initiation steps [12], [16]–[19], re-initiation during multiple rounds of transcription [20] and regulation of chromatin structure [12], [21], [22]. Two recent reports showed an additional role for the core Mediator in sub-telomeric gene silencing [23], [24]. In addition to these versatile roles in gene transcription, Mediator also appears to be a central “integrative hub” for the regulation of gene expression by physiological signals [12]. Examples from yeast include regulation of the Kinase domain by the Ras/PKA pathway via phosphorylation of the Srb9/Med13 subunit [25], and control over the expression of iron-responsive genes by an interplay between the Tail subunit Med2, which has a positive role, and the Kinase domain that phosphorylates Med2 to inhibit its function [7], [26].
The multisubunit Mediator complex emerged early in the evolution of eukaryotes, and the versatility of its functions and its role as an integrative platform for cell physiology could have contributed to the shaping of gene expression programs in different species, for adaptation to specific environments and life styles [27]. Fungi represent an excellent model system for exploring these questions. A comparative analysis in model yeasts Saccharomyces cerevisiae and Schizosaccharomyces pombe showed remarkable conservation of the roles of Mediator in spite of the fact that these two yeasts are highly divergent [28]. The conserved functions include a broad role in stress responses, and specific, distinct roles of the Mediator domain in the regulation of cell wall dynamics and cell morphology. The Head and Middle domains of Mediator are required for the expression of the cytokinesis genes under the control of the transcription factor Ace2 [23], [28], while the Kinase domain represses transcription of the cell wall adhesins [7], [9], [25], [28], [29]. Furthermore, studies of the Srb11/Ssn8 cyclin subunit of the Mediator Kinase domain in human and plant fungal pathogens (Cryptococcus neoformans, Candida albicans, Fusarium vertisilloides and Fusarium gramineaurum) suggest conserved roles in the repression of nutrient responsive functions and genes required for the production of toxins and pigments, as well as a conserved role in stress responses and regulation of cell wall integrity [9], [30]–[37]. Moreover, in Candida glabrata, the Mediator Tail subunit Med15/Gal11 plays a conserved role with S. cerevisiae in drug resistance mediated by the transcription factor Pdr1 [38], [39].
Here we report the first study of Mediator functions in the human pathogen C. albicans. We show that the C. albicans Mediator has some conserved functions with S. cerevisiae and Schizo. pombe, but also has additional roles in the expression of virulence-related genes, most notably the ALS adhesins. Phenotypic analysis showed roles for Mediator subunits in phenotypes of C. albicans associated with pathogenesis – filamentous growth and biofilm formation. Our data presented here and previous reports [25], [40] show that control of the cell wall adhesins by Mediator subunits Med31 and Srb9/Med13 differs between S. cerevisiae and C. albicans, suggesting distinct Mediator-dependent control of adhesion in these two yeast species.
Results
Roles for the C. albicans Mediator Middle domain subunit Med31 in the expression of genes related to morphogenesis, host–pathogen interactions, and pathogen-specific gene families
To start delineating the function of the Mediator complex in C. albicans, we made homozygous deletion mutants in the Middle domain subunit Med31. In C. albicans, Med31 is encoded by orf19.1429 and it displays 48.2% and 39.80% sequence identity with its S. cerevisiae and Schizo. pombe orthologs respectively. Transcriptome-wide profiles of the med31ΔΔ mutant were obtained and compared to those of a complemented med31ΔΔ+MED31 strain. Routine manipulations during mutant strain construction can result in gross chromosomal rearrangements, such as aneuploidies [41], which could profoundly affect the results of transcriptome analysis. Inspection of the med31ΔΔ transcriptional profile in the chromosomal context did not reveal a colour distribution associated with aneuploidies (Figure S1A), indicating that the med31ΔΔ mutant has the same chromosomal structure as the complemented strain. Additionally, gene sets representing 50 kb fragments were included in the Gene Set Enrichment Analysis (see below), and this analysis also did not reveal any gross chromosomal alterations (data not shown).
In agreement with a general role for Med31 in gene expression, and consistent with data from S. cerevisiae and Schizo. pombe [7], [28], [42], [43], 7.8% of the genome (510 genes) was differentially expressed in the absence of MED31 (cut-off of 1.5 fold, p<0.05, Dataset S1). Out of the genes differentially expressed in med31ΔΔ cells, 61.7% (315) were down-regulated and 38.2% (195) were up-regulated. This is consistent with a predominantly positive role of Med31 in transcription, and is in line with reports in S. cerevisiae and Schizo. pombe [7], [28], [42], [43]. To reveal the cellular pathways regulated by Med31 in C. albicans, we performed Gene Set Enrichment Analysis (GSEA) [44], [45]. GSEA compares a list from the transcript profile of interest created by ranking all of the genes according to the change in their expression (in this case that of a med31ΔΔ mutant) to a predefined gene set, and asks if a specific gene set is enriched in the top (up-regulated genes) or the bottom (down-regulated genes) of the ranked list [44], [45]. A ranked list of genes from the transcript profile of med31ΔΔ cells was compared to a custom database of 8123 gene sets (http://candida2.bri.nrc.ca/andre/GSEA/index.cfm; Sellam and Nantel, submitted) constructed using GO annotations and protein interaction data from CGD (PMID: 19808938), SGD (http://www.yeastgenome.org) and BioGRID [46], most currently published C. albicans transcriptional profiling and ChIP-CHIP experiments, our own TF motif database (PMID: 18342603), and S. cerevisiae genetic-association data (PMID: 20093466). Since profiles can exhibit correlations with hundreds of overlapping gene sets, significantly enriched gene sets (p<0.005, FDR<25%) were further organized and visualized using the Cytoscape: Enrichment Map plug-in (PMCID: PMC2981572), which produces networks of gene sets that share significant overlaps with each other (Figure 1A shows the most prominent networks of genes; the complete network is shown in Figure S2 where the details can be visualised by using the “zoom in” function in the pdf document). Figure 1B shows examples of enrichment plots for selected gene sets. The complete GSEA output can be found at http://dl.dropbox.com/u/7211133/Med31%20GSEA%20Results.zip.

GSEA detected enrichment for nucleolar functions, rRNA and ribosome biogenesis genes, and genes involved in nucleotide biosynthesis in the set of genes down-regulated in the med31ΔΔ mutant, while genes required for mitochondrial function were up-regulated (Figure 1A and 1B). Enrichment was also found in gene sets important for virulence-promoting function in C. albicans. Those include genes differentially expressed during C. albicans-host interactions with mouse macrophages [47], reconstituted human oral epithelial cells [48] and polymorphonuclear leukocytes [49], as well as genes differentially expressed in conditions which alter cellular morphogenesis, such as the induction of hyphal growth [50], [51], mutations in the Ras-cAMP morphogenesis pathway (ras1 and cdc35/cyc1) [52], and inhibition of cell cycle progression that causes pronounced polarised growth (treatment with hydroxyurea or down-regulation of the polo-like kinase CDC5) [53] (Figure 1A and 1B). Genes expressed at the G1-S and S-G2 transition of the cell cycle [54] were up-regulated in the med31ΔΔ mutant, as were those required for membrane and cell wall biosynthesis (Figure 1A and 1B). Genes required for cytokinesis were down-regulated (as shown by the black arrow in Figure 1A, and in the enrichment plot in Figure 1B).
Modulation of several gene sets enriched in the med31ΔΔ mutant, for example down-regulation of genes required for protein synthesis and up-regulation of those required for cell wall biogenesis, is part of a more general stress response in C. albicans [55]. Mediator has been previously implicated in stress responses in yeasts [7], [28], and it is therefore possible that some of the differences in the med31ΔΔ transcriptome are due to activation of stress responses upon loss of Med31 function. However, our analysis indicates that this is unlikely to be the cause for much of the differential gene expression in the mutant. There was little correlation between the med31ΔΔ transcriptional profile and our large database of transcriptional profiles produced from stressed cells when analysed by GSEA (of note, profile to profiles comparisons such as those done by GSEA tend to produce the strongest correlations and therefore if a correlation existed it is very likely that it would have been detected by GSEA). We further used scatter plots to directly compare the med31ΔΔ profile with stressful conditions, such as osmotic or oxidative stress, and these comparisons confirmed lack of extensive correlation between the med31ΔΔ transcriptome and differential gene expression upon stress (Figure S1B).
Analysis of gene ontology terms using the GO term finder tool at the Candida Genome database and the genes up - or down-regulated by at least 1.5 fold in med31ΔΔ cells (see Dataset S1) confirmed that genes related to morphogenesis, mitochondrial function and the cell wall were differentially expressed (Table 1 and Dataset S2). Interestingly, several central regulators of filamentous differentiation, such as the transcription factors Tec1, Efg1, Cph1 and Nrg1, were amongst the down-regulated genes, as were six out of the eight genes from the FGR6 (Filamentous Growth Regulator) family located in the RB2 repeat sequence (Dataset S2). The FGR6 family is one of the gene families found to be enriched in pathogenic yeast species, and specifically expanded in C. albicans [56]. While GSEA scored the cell wall gene set as up-regulated, we noticed that there were also several genes in this group that appeared at the bottom of the list, in the down-regulated group. In fact, another Candida-specific gene family expanded in pathogens was down-regulated in the med31ΔΔ mutant, that encoding the ALS cell wall adhesins [56] (Table 2). The major C. albicans adhesin ALS1 was one of the most down-regulated genes in the mutant (5 fold down-regulation, Table 2 and Dataset S1). ALS5 and ALS6 were also down-regulated (Table 2), but of note, these genes are expected to be expressed at very low levels in the wild type. Additionally, several other genes encoding cell wall proteins were down-regulated in the mutant, as were genes necessary for cell wall construction and remodelling, in particular those required for cytokinesis and regulated by the transcription factor Ace2 (e.g. the chitinase CHT3 and the endoglucanase ENG1) [57] (Table 2; notably GSEA also scored the cytokinesis genes as down-regulated and this is shown in Figure 1). The existence of several down-regulated cell wall genes indicates that the up-regulation of genes with roles in cell wall integrity that is detected in the med31ΔΔ mutant (Figure 1 and Table 2) likely reflects a compensatory feedback regulation due to a defective cell wall structure in the absence of Med31. That med31ΔΔ mutants have altered cell walls is supported by phenotypic analysis demonstrating changes in sensitivity to the cell wall targeting drugs congo red and calcofluor white (Table 3).

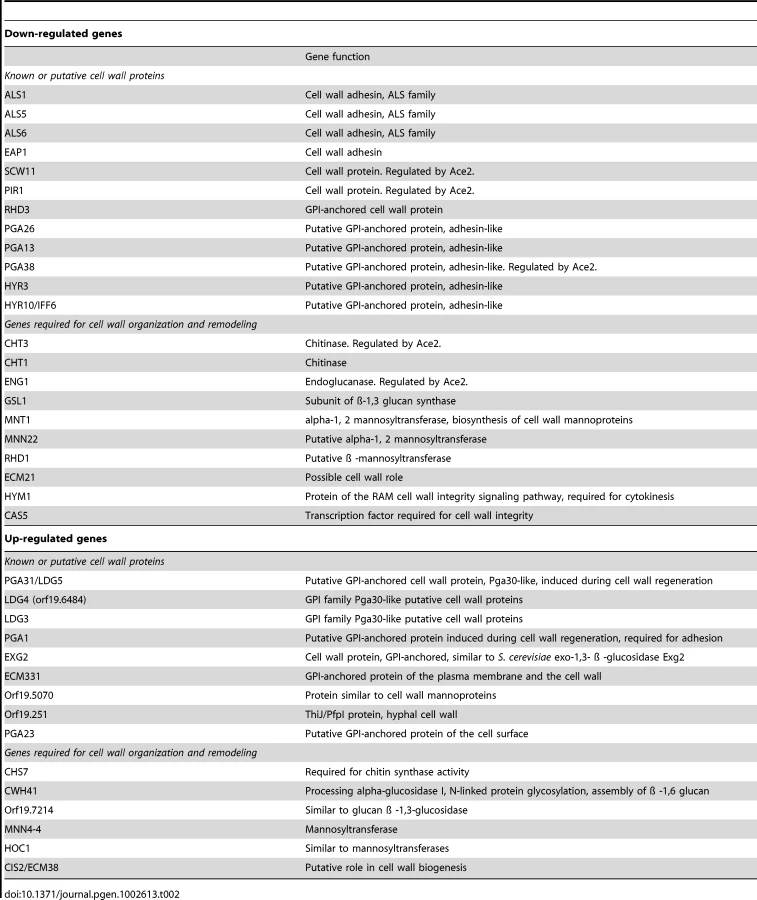

In conclusion, the transcriptome analysis indicated a broad role for Med31 in cell physiology in C. albicans, with functions in morphogenesis and cell cycle progression, growth and metabolism, cell wall integrity, the expression of the cytokinesis genes under the control of the transcription factor Ace2 and those regulated by the interaction of C. albicans with host cells. Finally, two gene families enriched in pathogenic yeasts, the FGR family of filamentous growth regulators and the ALS adhesins, required Med31 for wild type expression levels.
Regulation of Ace2-dependent genes and cell wall adhesins by Med31 in yeast and hyphal growth
Down-regulation of Ace2 target genes in the absence of Med31 in C. albicans is in agreement with a role for Med31 and other Mediator subunits in Ace2-dependent gene expression that is conserved between C. albicans, Schizo. pombe and S. cerevisiae (this study and [28]). Comparing more broadly the genes affected in med31ΔΔ cells with those reported to be differentially expressed in the C. albicans ace2 mutant [57] revealed that differential expression of 35 genes is shared between these two transcription factors (Table S1). Genes involved in cytokinesis and cell wall functions were predominant in the shared “down-regulated” group, whereas mitochondrial biogenesis genes were predominant in the shared “up-regulated” group (Table S1). This analysis suggests that the functions of Med31 in cell wall integrity and metabolism are mediated, at least in part, by Ace2-dependent roles.
To explore this further, we used quantitative PCR (qPCR) to directly compare the expression levels of candidate genes in the med31ΔΔ and ace2ΔΔ mutants (Figure 2A). Under yeast growth conditions (as was done for the transcriptome analysis) the mRNA levels for the chitinase CHT3 and the cell wall proteins PIR1, EAP1 and ALS1 were reduced in both med31ΔΔ and ace2ΔΔ cells (albeit to a different degree). CHT3 and PIR1 have been previously reported as Ace2-targets, while ALS1 and EAP1 were not [57]. The expression of the transcription factor TYE7 was reduced in med31ΔΔ cells (consistent with the microarray data), but not in the ace2ΔΔ mutant.

In C. albicans, the expression of cell wall proteins is activated upon filamentous differentiation [50], [51], [58]–[61]. This includes ALS1 and other ALS and non-ALS adhesins. Therefore, we next tested if Med31 was required for the expression of the ALS1, and two hypha-specific adhesins ALS3 and HWP1, during filamentous growth. For these experiments cells were grown in filament-inducing Spider media at 37°C. The mRNA levels of both ALS1 and ALS3 were down-regulated in med31ΔΔ cells (Figure 2A). Ace2 was not required for ALS1 expression during hyphal growth, while the levels of ALS3 were lower in cells lacking Ace2, but the effect was less than in the absence of Med31 (Figure 2A). HWP1 was up-regulated in both mutants (1.8–3 fold) (Figure 2A). Collectively, the qPCR analysis suggests that Med31 and Ace2 have common, but also independent roles in the expression of the cytokinesis genes and the cell wall adhesins during yeast and hyphal growth.
Given that we found novel cell wall protein targets that require Ace2 for wild type expression (ALS1, ALS3 and EAP1), we searched the upstream regulatory regions of these genes for putative Ace2 binding sites as defined in [57]. There are three Ace2-binding motifs within 1.5 kb upstream of the start codon for ALS3 (Figure 2B). No motifs that strictly conform to the consensus sequence were found in the promoters of ALS1 and EAP1, although variant motifs could be found (data not shown). The motifs in the ALS3 promoter included one at −467 bp, which is in a region found to be essential for ALS3 activation in hyphae (the so-called A1 region) [59]. This suggests that ALS3 could be a direct target of Ace2.
Phenotypic analysis shows roles for Med31 in cytokinesis, morphogenesis, and virulence
We next performed phenotypic analysis of the C. albicans med31ΔΔ mutant to address the biological relevance of the observed changes in gene expression. Cells lacking Med31 displayed a cytokinesis defect (Figure 3A), consistent with a role in Ace2-dependent gene expression. ∼40% of cells from med31ΔΔ cultures showed a cell-chain phenotype typical of mutants that cannot undergo cytokinesis. This phenotype was observed in two independently constructed homozygous deletion mutants and was partially complemented by re-introduction of a wild type copy of MED31 into the mutant genome (Figure 3A and 3B).

The med31ΔΔ mutant also displayed phenotypes consistent with altered cell membrane and cell wall integrity that were suggested by transcriptome analysis. The mutant was sensitive to formamide, SDS, the sterol-binding antifungal drug nystatin, DMSO and growth at 16°C, all phenotypes consistent with defective membrane structure. The mutant was also sensitive to the cell wall-targeting drug congo red, but more resistant to the chitin-binding dye calcofluor white (Table 3 and Figure S4). Furthermore, the med31ΔΔ mutant was sensitive to oxidative and salt stress and ethanol (Table 3 and Figure S4). Some of these phenotypes are also observed in S. cerevisiae and Schizo. pombe med31 mutants, suggesting conserved roles [28], [43]. We also tested the C. albicans ace2ΔΔ mutant side by side with med31ΔΔ for tolerance to the various compounds (Table 3 and Figure S4). Resistance to calcoflour white was also observed in the ace2ΔΔ mutant (Table 3 and [62]), as was a mild sensitivity to nystatin and growth at 16°C, indicating that these phenotypes could be due to the role of Med31 in Ace2-dependent gene expression. However, the other sensitivities of the med31ΔΔ mutant were not shared by the ace2ΔΔ mutant, and are therefore unrelated to Ace2-dependent phenotypes.
The transcriptome analysis indicated a role for Med31 in cellular morphogenesis and we therefore tested whether Med31 was necessary for the yeast-to-hypha morphogenetic switch in response to a variety of inducers in vitro. The med31ΔΔ mutant was unable to produce filaments on solid Spider and M199 media, or on plates containing N-acetylglucosamine (Figure 3C). The mutant was also compromised for filamentation in liquid media, however to a lesser extent than on plates (Figure S5). med31ΔΔ cells could filament in response to serum and in M199 media in culture, but with delayed kinetics and with a proportion of cells remaining in yeast form (Figure S5). In Spider media the mutant had a more pronounced defect, and even after 7 h cells were still largely in yeast form (Figure S5). However, after prolonged incubation of 12 h, filaments were observed in this medium also (data not shown). To address whether the role for Med31 in filamentous growth would be important in a disease context, we further tested the ability of med31ΔΔ cells to filament in vivo, using the C. albicans-Caenorhabditis elegans infection model [63], [64]. This is a well-established host-pathogen system, which recapitulates key elements of disease as seen in vertebrates, most notably, the requirement for filamentous growth [63]. Only 16.5% of the worms infected with the mutant developed filaments after 3 days compared to 49% for worms infected with wild type C. albicans or the complemented strain (Figure 4A and 4B). Moreover, for those med31ΔΔ-infected worms that did develop filaments, there was a significant delay in filamentation, there were many fewer filaments per worm compared to the wild type or complemented strains, and the filaments were much shorter (Figure 4A; a similar phenotype was observed even after 7 days of infection, Figure S6). We also evaluated the pathogenic potential of the med31ΔΔ mutant in the worm. The kinetics of killing of the worm by C. albicans was delayed in worms infected with the med31ΔΔ mutant compared to those infected with the wild type (p<0.02 for all experiments performed) (Figure 4C). This result supports a role for Med31 in C. albicans virulence.

Med31 is necessary for biofilm formation, and ALS1 is a relevant gene target for this phenotype
We next sought to show that the regulation of the adhesins by Med31 was biologically relevant. To that end, we tested the ability of the med31ΔΔ mutant to form biofilms, as both ALS1 and ALS3 are key adhesins for biofilm formation by C. albicans [65]. Biofilm formation was tested in vitro, on serum-coated silicone disks as previously described [66]. In the absence of Med31, biofilms were severely compromised in both density and depth, as determined by confocal scanning laser microscopy (Figure 5A). Scanning electron microscopy (SEM) showed a similar defect (Figure S7). Quantitative analysis confirmed the phenotype observed by microscopy (Figure 5B). The med31ΔΔ mutant displayed a strong defect at the earliest time point of 90 minutes (adherence stage). The med31ΔΔ mutant grew somewhat slower than the wild type in planktonic conditions (Figures S4 and S8), however at the 90 minutes time point in the biofilm formation assay we did not observe significant cell growth for any of the strains, including the wild type (data not shown), strongly suggesting that the defect in the med31ΔΔ mutant is due to an adherence defect and not to the observed slower growth.

Rescue by over-expression of target genes has been previously used as a strategy for identification of transcription factor targets relevant for biofilm formation (for example in the C. albicans bcr1 mutant [65]). We employed a similar strategy to test whether ALS1 is a relevant Med31 target gene for biofilm formation. The idea is that, if lower expression of ALS1 in the med31ΔΔ mutant is contributing to the biofilm defect, its ectopic expression under a constitutive promoter should, at least in part, rescue biofilm formation by med31ΔΔ cells. As shown in Figure 5B, expression of ALS1 in the med31ΔΔ mutant under the constitutive TEF1 promoter led to a substantial rescue of the biofilm defect at all time points, including the early adherence and initiation stages. Similar rescue was observed in three independent med31ΔΔ+TEF1-ALS1 clones. Expression of TEF1-ALS1 in the wild type strain did not significantly change biofilm biomass (data not shown). As an independent confirmation that the biofilm formation defect of the med31ΔΔ mutant is not due to slower growth, we tested whether introduction of the TEF1-ALS1 construct was rescuing the growth defect of the med31ΔΔ mutant. We did not observe rescue of the mutant growth defect by TEF1-ALS1 (Figure S8), although biofilm formation was rescued (Figure 5B). This confirms that the biofilm defect is due to lower adherence and not slower growth. Collectively, these results suggest that ALS1 is a biologically relevant gene target of Med31 for biofilm formation by C. albicans.
The Mediator Head domain subunit Med20 and Srb9/Med13 from the Kinase domain also contribute to morphogenesis, biofilm formation, and adhesin gene expression in C. albicans
To address more generally the roles of Mediator in C. albicans, we constructed mutants in the Med20 subunit from the Head domain (orf19.2711.1) and the Kinase domain subunit Srb9/Med13 (orf19.1452; for simplicity the mutant is indicated as srb9ΔΔ in the figures) [27]. In agreement with a general role for Mediator in adhesin gene expression, both MED20 and SRB9/MED13 were required for the expression of ALS1 and ALS3 in hyphae, and SRB9/MED13 was further required for wild type transcript levels of HWP1 (Figure 6A). Consistent with the effects on adhesins, both MED20 and SRB9/MED13 were necessary for wild type biofilm formation (Figure 6B). In regards to filamentous growth, we observed a similar trend as with the med31ΔΔ cells: filamentous growth was severely compromised in med20ΔΔ and srb9/med13ΔΔ mutants on plates (Figure 6C), while in liquid media the effects were much less pronounced (Figure 6D). In liquid media, med20ΔΔ showed a mild defect with a larger proportion of pseudohyphae (for example see the 3 and 5 h time points in serum), while srb9/med13ΔΔ behaved like the wild type (Figure 6D). To address the similarities and differences between the Mediator subunits more broadly, we analysed whether Med31-dependent genes during yeast growth were also dependent on Med20 and Srb9/Med13 for their expression. EAP1 was down-regulated in med20ΔΔ cells, ALS1 was down-regulated in both mutants, and CHT3 was only affected in srb9/med13ΔΔ cells (Figure 7A). The med20ΔΔ mutant did not display a cytokinesis defect, consistent with wild type CHT3 levels (Figure 7B). The srb9/med13ΔΔ cells were slightly elongated and a fraction (between 20–40%) formed what appeared similar to pseudohyphae, perhaps consistent with lower CHT3 levels (Figure 7B). The med20ΔΔ and srb9/med13ΔΔ mutants shared some (but not all) sensitivities to antifungal compounds and stress conditions with the med31ΔΔ strain (Table 3), and some of these sensitivities are conserved with what has been reported for the homologous mutations in model yeasts [28].


The roles of Med31 in adhesion are different between S. cerevisiae and C. albicans
In the model yeast S. cerevisiae Mediator has been implicated in the repression of the FLO cell wall adhesins, due to the inhibitory functions of the Kinase domain subunits (including Srb9/Med13) and the Tail domain component Sin4/Med16 [25], [40]. Repression of the cell wall adhesins by the Mediator Kinase domain has also been reported in Schizo. pombe [28]. Our data in C. albicans showed that Srb9/Med13 is a positive regulator of the genes encoding adhesins (Figure 6), thus indicating that the roles of this Mediator Kinase domain subunit in adhesion are different in C. albicans. To probe this notion further, we tested the role of Med31 in adhesion in S. cerevisiae using the Σ1278b strain background, which expresses the FLO11 adhesin and is capable of adhesion-dependent phenotypes such as biofilm formation on polystyrene [67]. The wrinkled colony morphology in the med31Δ mutant was enhanced compared to the wild type, a phenotype that is indicative of higher FLO11 levels (Figure 8A). Indeed, the expression of FLO11 was significantly up-regulated in the absence of MED31 (40 fold), and consistently, the mutant was hyper-adherent in the S. cerevisiae biofilm model (Figure 8B and 8C). The levels of the other FLO family members, which are silent in the wild type, were not up-regulated under the conditions assayed (data not shown). These results are consistent with a repressive role for Med31 in the expression of FLO11 and biofilm formation in S. cerevisiae.

Discussion
So far, most comparative analysis of transcription factor function in fungi have focused on DNA binding gene-specific transcription factors, and only very few studies have addressed the functions of transcriptional co-regulator complexes, such as Mediator. With Mediator, conserved roles have been reported for the core complex and the Kinase domain, including distinct roles of the sub-domains in cell wall dynamics shared by the divergent species S. cerevisiae and Schizo. pombe [9], [23], [25], [28], [29]. Our study is the first comprehensive analysis of Mediator components in the pathogen C. albicans. We identified conserved functions for Mediator subunits in Ace2-dependent gene expression and stress responsive phenotypes with the model yeasts, but also novel roles in the expression of genes related to virulence attributes of C. albicans. In particular, our study uncovered roles in the expression of the cell wall adhesins and functions in biofilm formation, which are shared between the Middle domain subunit Med31, the Head domain subunit Med20 and Srb9/Med13 from the Kinase domain.
Overlapping and specific roles for the Mediator subunits in C. albicans
Our data in C. albicans is consistent with reports in model yeasts in that Mediator subunits show overlapping, but also some specific roles [7], [28], [43]. In S. cerevisiae and Schizo. pombe, mutants in the Head and Middle domain subunits tend to correlate in regards to gene expression and cellular phenotypes [7], [28], [43]. We also find this for the C. albicans med31ΔΔ and med20ΔΔ mutants. For example, the expression of the cell wall adhesins ALS1, ALS3, EAP1 is down-regulated in the absence of either MED31 or MED20, both mutants display biofilm defects and a stronger filamentation defect on plates than in liquid media, and they share sensitivities to compounds such as formamide, DMSO, SDS, ethanol and congo red (Table 3). However, our data also supports differences in the roles of Med20 and Med31: Med20 is not required for the expression of the Med31-regulated genes CHT3, PIR1 and TYE7, it shows a milder biofilm formation and filamentous growth defect than Med31, and there are differences in the sensitivities to some compounds between the two mutants. Shared but also distinct roles of Med20 and Med31 are supported by results in S. cerevisiae showing a 0.41 Pearson correlation coefficient between the transcript profiles form the two mutants [43]. In Schizo. pombe, in addition to Med31, Med20 and other Head domain subunits also control the expression of Ace2-dependent genes and the mutants show defects in cell separation [28]. Deletion of MED20 in C. albicans did not result in lower expression of CHT3 or a cytokinesis defect (Figure 7). Together with Med18 and the C-terminal domain of Med8, Med20 forms a structural and functional sub-complex within the Head domain, the Med8C/18/20 submodule [28], [68], [69]. Genetic analysis in Schizo. pombe supports the idea that Med18 can compensate for the loss of Med20 [28]. It is therefore possible that Med18 would need to be inactivated in C. albicans to uncover the roles of the Mediator Head domain in Ace2-dependent transcription and cytokinesis.
In contrast to the core Mediator complex which functions predominantly in transcriptional activation, the Kinase domain is mainly a repressor of transcription and the phenotypes of the Kinase mutants in model yeasts tend not to correlate with mutants in the core Mediator (for example see [7], [28]). Our data in C. albicans supports this, as the med31ΔΔ and med20ΔΔ mutants were more similar to each other in terms of gene expression and phenotypes than to the srb9/med13ΔΔ strain. However, the srb9/med13ΔΔ strain shared some phenotypes with med31ΔΔ and med20ΔΔ including the effects on ALS gene expression, biofilm defects and sensitivities to compounds that affect the cell wall, such as congo red and SDS. The Mediator kinase domain consists of four subunits: Srb8/Med12, Srb9/Med13 and the kinase-cyclin pair Srb10/Srb11. The C. albicans mutants in Srb10/Srb11 were in the Kinase collection constructed by Blankenship et al. [35]. Our srb9/med13ΔΔ mutant shares the sensitivity to oxidative stress with cells inactivated for SRB10 or SRB11 (Table 3 and [35]), however the srb10 and srb11 mutants were reported to display wild type biofilm formation [35]. The Kinase domain subunits share many functions, but can also have different roles, in particular positive functions in the transcription of some genes have been reported that are not shared by the whole domain [70], [71]. The data reported here (Figure 6) and in [35] support distinct roles for the C. albicans Mediator Kinase domain subunits in biofilm formation.
The connection between Med31 and Ace2
Our data indicates that in C. albicans the transcriptional activator Ace2 could be modulating a number of Med31-dependent effects on gene expression, in particular transcriptional activation of the cytokinesis genes, the expression of the adhesins EAP1 and ALS3 and the cell wall protein PIR1, and the regulation of genes with mitochondrial functions (Figure 2 and Table S1). The ALS3 promoter contains a putative consensus Ace2 binding site within a region known to be required for activation [59], suggesting it could be a direct target of Ace2. Consistent with shared functions, both ace2ΔΔ and med31ΔΔ mutants display a cytokinesis defect, as well as adherence and biofilm formation phenotypes (this study and [72]). In Schizo. pombe Mediator interacts directly with Ace2, via the Head domain subunit Med8, and therefore Mediator plays a direct role in Ace2-dependent transcription [73]. By analogy to Schizo. pombe, and given that we did not observe a change in transcript levels for ACE2 in the med31ΔΔ mutant that would indicate an indirect effect (Dataset S1), we propose that in C. albicans Ace2 also interacts directly with Mediator for transcriptional activation. Our data also suggests that Med31 and Ace2 have roles in transcription that are independent of each other. Unlike Ace2, which is indispensable for cytokinesis in C. albicans, a milder phenotype is observed in the absence of Med31 (Figure 3B). There is only a partial overlap between genes differentially expressed in ace2 and med31 mutants respectively ([57] and this study, Table S1), and the degree of the effects on gene transcription differs between the two mutants (for example CHT3 is much more affected by inactivation of ACE2 than MED31, while MED31 has a stronger effect on PIR1, Figure 2). ALS1, which we show is a key target of Med31, does not require Ace2 during high transcription in hyphae (Figure 2). As in Schizo. pombe [28], [73], it could be that in C. albicans Mediator subunits additional to Med31 are involved in co-activating Ace2-dependent transcription. Conversely, Med31 certainly acts through additional DNA binding transcription factors, which remain to be identified.
Mediator and the expression of cell wall adhesins in different yeast species
Med31, Med20 and Srb9/Med13 all positively regulate the expression of the ALS1 and ALS3 cell wall adhesins in C. albicans, and Srb9/Med13 is further required for wild type expression of HWP1. Consistent with the transcriptional defects, the mutants are defective for biofilm formation, an adhesin-dependent phenotype [65], . Moreover, our genetic data supports the notion that the regulation of ALS1 by Med31 is biologically relevant for biofilm development. The activators that mediate the effects of Mediator on adhesin transcription remain to be characterised. The results that Med31 is required for Ace2-dependent transcription, and that Ace2 has a role in ALS3 expression suggest a potential role for Ace2 in Med31-dependent regulation of ALS3 (Figure 2). The srb9/med13ΔΔ mutant showed parallels in regards to both morphogenesis and gene expression phenotypes with the mutant inactivated in the activator Bcr1. Both mutants are required for the expression of ALS1, ALS3 and HWP1, and, as was seen for the bcr1 mutant, the srb9/med13ΔΔ mutant is proficient for hyphal growth in liquid media, but shows biofilm defects (Figure 6 and [65], [66]).
The biochemical, genetic and gene expression data in yeast supports the notion that Mediator is universally required for RNA polymerase II-dependent transcription (reviewed in [12]), and we therefore suggest that Mediator is directly involved in the transcription of the adhesin genes. Chromatin immunoprecipitation (ChIP) experiments to address occupancy of the ALS gene promoters by Med31 remained inconclusive, as we observed variability between biological replicates, from minimal to large 20–30 fold enrichments (data not shown). We suspect that this variability could be due to the exact timing of the crosslinking of Med31 to the promoters in respect to the transcriptional activation of the ALS genes in hyphae and/or how uniformly filamentous growth/ALS gene transcription is induced in the population of cells in the culture. ChIP studies in S. cerevisiae have yielded different results between labs in regards to Mediator occupancy, from modest (albeit functionally important) enrichments at some constitutively transcribed genes [75], no enrichment of Mediator subunits on the majority of transcribed genes [76], [77], to detectable enrichment of Mediator subunits upstream of many active, as well as inactive genes, and even in the coding regions of some genes [78]. More prominent enrichment for Mediator subunits is seen on genes that are responsive to stress (e.g. heat shock, or change of carbon source from glucose to galactose) [75]–[77]. Mediator does not bind directly to DNA, which is likely to be a factor in ChIP experiments. It has also been proposed that the interactions of Mediator with promoters could be transient [77]. Moreover, different Mediator subunits can yield different fold enrichments over the background (for example see [76], [77]), and it is therefore possible that a Mediator subunit other than Med31 needs to be assayed to detect consistent Mediator occupancy on promoters in C. albicans. In addition to a direct role for Mediator in adhesin gene expression, an alternative (and not mutually exclusive) possibility is that Mediator regulates the expression of transcription factors, which then in turn regulate the adhesins. The expression of several transcription factors was lower in med31ΔΔ mutants (Table 1, Dataset S2), for example that of EFG1, TEC1 and CPH1, which have been previously shown to regulate the expression of Med31-regulated adhesins ALS1, ALS3 and EAP1 [59], [79].
In S. cerevisiae Med31 is a repressor of the adhesin FLO11 (Figure 8), suggesting different functions for this Mediator subunit in the expression of cell wall adhesion molecules and regulation of adhesion-dependent phenotypes in comparison to C. albicans. In the S. cerevisiae med31Δ mutant, the reminder of the Mediator complex is intact [43], supportive of a specific role for Med31 in the repression of the FLO11 gene that does not result simply from the loss of other repressor subunits from the Mediator complex. Previous reports in S. cerevisiae for Srb9/Med13 also show repression of the FLO adhesins [25], which is again opposite to the activating function for Srb9/Med13 in the expression of ALS1/3 and HWP1 that we observed in C. albicans. Other subunits of the Mediator Kinase domain as well as Sin4/Med16 from the Tail also inhibit expression of the S. cerevisiae FLO genes [25], [40]. In Schizo. pombe, the Mediator Kinase domain subunits Srb10 and Srb8/Med12 also repress the cell wall adhesins, despite the fact that some of the affected adhesins are not related to the FLO genes [28].
While the FLO and ALS gene are not related, there is conservation in terms of the pathways and transcription factors that regulate their expression in S. cerevisiae and C. albicans respectively. Examples include positive regulation by activators Flo8, Ste12/Cph1 and Tec1, and negative regulation by repressors Nrg1, Tup1 and Sfl1 [50], [59], [80]–[83]. Mediator interacts with gene-specific transcription factors to regulate gene expression [76], [84]. It is possible that the key transcription factor(s) through which Med31 and Srb9/Med13 act to regulate the adhesin-encoding genes differ between the two yeasts. How Mediator subunits repress the S. cerevisiae FLO genes is not well understood, but a suggested mechanism involves functional interactions with the repressors Sfl1 and Tup1 [85]. Both Sfl1 and Tup1 are required for repression of the ALS genes in C. albicans [50], [81], but the requirement for specific Mediator subunits in the repression of adhesins by these factors in C. albicans could differ from what has been suggested in S. cerevisiae. Another possibility relates to silencing mechanisms. The FLO genes in S. cerevisiae are subject to position-dependent silencing regulated by histone deacetylases ([86]; reviewed in [87]). The C. albicans adhesins are not known to be regulated by silencing. Mediator has been implicated in the regulation of chromatin structure and gene expression in regions affected by silencing, such as the sub-telomeres [12], [21]–[24]. Based on work in S. cerevisiae, it has been proposed that Mediator contributes to the establishment of a repressive chromatin structure by binding to the silenced regions and influencing the recruitment of the histone deaceytlase Sir2 and the histone acetyltransferase Sas2 and thus the acetylation status of histone H4 K16, a mark of active chromatin [24]. While this has not been tested directly, an interesting possibility is that the regulation of chromatin contributes to the repressive role of the Mediator subunits in the expression of the S. cerevisiae FLO genes. For example, the Tail subunit Sin4/Med16, which is a repressor of FLO11 and of the sub-telomeric FLO1 adhesin [40], [85], contributes to sub-telomeric silencing [24]. Med7 has also been found to affect chromatin structure, by regulating H4K16 acetylation and the presence of Sir2 and Sas2 at the sub-telomeres [24]. Med7 is closely functionally linked with Med31 - the N-terminal part of Med7 together with Med31 forms a structural and functional sub-module of Mediator, Med7N/31 [43]. It will be interesting to study whether differences in silencing mechanisms that operate on the FLO and ALS genes contribute to determining how Mediator subunits control their expression.
Materials and Methods
Yeast strains and growth conditions
The C. albicans strains used in this study are derivatives of BWP17 [88]. The med31ΔΔ, srb9ΔΔ and med20ΔΔ strains were constructed by standard methods based on PCR and homologous recombination, using ARG4 and URA3 as selective markers. The complemented strains were constructed by re-introducing a wild type copy of MED31, SRB9 or MED20 under own promoter and terminator into the HIS1 locus of the respective mutants using the integrative plasmid pDDB78. To make matched HIS1+ mutant strains, an empty pDDB78 vector was integrated into the genome of the respective strains. The ace2 mutant is a homozygous mutant in the BWP17 strain background and was a generous gift from Aaron Mitchell (this strain is also URA3+ ARG4+ HIS1+). The med31ΔΔ +TEF1-ALS1 overexpression strain was constructed using the plasmid pCJN498, as described in [65]. The S. cerevisiae med31Δ mutant was constructed in the ∑1278b strain using the KANMX4 cassette. The wt and flo11Δ mutant of Σ1278b were a generous gift from Todd Reynolds and are described in [67].
Standard growth conditions were YPD (2% glucose, 2% peptone, 1% yeast extract), at 30°C, 200 rpm. For ura− strains the media was supplemented with 80 µg/ml uridine. The mutants were selected using minimal media lacking the appropriate amino acids. The TEF1-ALS1 overexpression strains have a nourseothricin resistance cassette (NAT) and were selected on 400 µg/ml NAT plates (nourseothricin was from Werner Bioagents). The S. cerevisiae mutant was selected on 200 µg/ml G418 plates.
For cell morphology analysis (Figure 3A), cells were classified as being in a chain if 3 or more cells were attached. An average of 200 cells per sample were counted, and the experiments were repeated at least with 3 independent cultures. The average and the standard error are shown in the figure. To observe the mother-bud junctions, cells were stained with calcofluor white (1 mg/ml) for 8 min in the dark, followed by washes in phosphate buffered saline (PBS). Imaging was done using an Olympus IX81 microscope with the Olympus cell∧M software, using the 100× objective with DIC or the DAPI filter for calcofluor white stained cells.
Filamentous growth was tested by dilution of cells from overnight cultures grown to OD600 = 0.1–0.2 into pre-warmed YPD+10% calf serum, Spider media (1% nutrient broth, 1% D-mannitol, 2 g K2HPO4), M199 or N-acetylglucosamine media (9 g NaCl, 6.7 g yeast nitrogen base and 0.56 g N-acetylglucosamine per liter) and incubated at 37°C for the times indicates in the figures. All cell imaging was done using an Olympus IX81 microscope with the Olympus cell∧M software. For testing filamentation on plates, C. albicans strains were re-streaked on plates containing filamentous-growth inducing media. Plates were incubated for up to five days at 37°C and colonies examined and photographed with a stereo dissecting microscope (Olympus SZX 16).
For analysis of sensitivities to various drugs and chemicals, ten fold serial dilutions of cultures from wild type and mutant strains were dropped on control plates, or plates containing the compounds indicated in Figure S4 and Table 3. Plates were incubated at 30°C for three days (unless growth was assessed at 37°C or 16°C), and photographed.
Biofilm assays and imaging
C. albicans biofilms were grown in 96-well microtiter plates or on silicone disks for quantitative or qualitative analysis respectively. Quantitative biofilm assays were performed as described [89], [90]. 100 ml of cultures of C. albicans wild type or mutant strains (107 cells/ml in Spider media) were added to wells and incubated at 37°C with gentle shaking (75 rpm) for 90 min (adhesion phase). Non-adherent cells were discarded and 100 µl of fresh Spider media were added to each of the wells. Biofilms were allowed to develop for a future 4.5 h (6 h in total), 24 h, and 48 h, representing the early, intermediate or mature stage of biofilm development respectively. The medium was replenished after 24 h by aspiration and addition of fresh medium. Biofilm biomass was determined at the different time points using crystal violet staining. Wells containing only Spider medium with no yeast served as negative controls. For qualitative studies, biofilms were formed in vitro on serum-treated silicone disks, which is a well-established system for biofilm analysis [66]. Sterile silicone disks were pretreated with fetal bovine serum (Sigma) overnight at 37°C with gentle shaking (75 rpm). The silicone disks were then washed twice with PBS and transferred to a 12-well plate containing 2 ml of freshly prepared cell suspensions (107 cells/ml in Spider media). The plate was incubated for 1.5 h at 37°C with gentle shaking (75 rpm) to allow the yeast to adhere to the disk surfaces. The silicone disks were then washed in PBS and transferred to a new 12-well plate with Spider media followed by incubation for 48 h at 37°C with shaking at 75 rpm. The established biofilms were examined with SEM or CLSM. For SEM, biofilms were fixed with glutaraldehyde (2.5%, v/v, in 0.1 M cacodylate buffer, pH 7.0) and 1% osmium tetraoxide at room temperature, and dehydrated with gradually increased ethanol (50%, 75%, 95%, 100%, and absolute 100%) and hexamethyldisilazane (HMDS) (50%, 75%, 95%, 100%, and absolute 100%). Samples were coated with gold with a Balzers SCD005 sputter coater and viewed under a Hitachi S570 scanning electron microscope. For CSLM, biofilms were stained with FUN-1 (10 µM, Molecular Probes) and Concanavalin A–Alexafluor488 conjugate (Con A, 25 µg ml−1; Molecular Probes) for 45 min at 37°C. Stained biofilms were observed with a Leica SP5 CLSM, and images were captured and processed using the softwares Leica LAS AF and Amira 5.2.1. All experiments for quantitative analysis were repeated at least three times in triplicate, and qualitative assays were repeated at least 3 times. One-way ANOVA was used to compare the difference in biofilm biomass produced by different C. albicans strains. p values of <0.05 were considered to be statistically significant.
For S. cerevisiae biofilms (Figure 8), overnight cultures were growth in synthetic complete media supplemented with 2% glucose. The cells were then resuspended into 96 well polystyrene plates to an OD600 = 1.0 using synthetic 0.2% glucose media [67]. Adherence was assayed at the time points indicated in the figure using crystal violet staining as described above.
RNA extraction, quantitative PCR, and microarray analysis
RNA was extracted using the hot-phenol method from cultures grown in YPD at 30°C to log phase (OD600 = 1). Following hot-phenol extraction, 100 µg of RNA samples were further purified using the RNAeasy kit and following the manufacturer's instructions. The microarray analysis was performed as described [91], on microarrays spotted with 6459 70-mer oligonucleotides (GEO Platform GPL9818). Data normalization and analysis was conducted in GeneSpring GX version 7.3 (Agilent Technologies). The microarray data set has been deposited in GEO, under accession number GSE31632. Genes that were up - or down-regulated in the med31ΔΔ mutant by 1.5 fold or more (p≤0.05) were selected from a Volcano Plot and considered to be differentially expressed. Gene ontology analysis was performed at the Candida genome database (CGD, candidagenome.org) [92]. GSEA analysis ([93] and Sellam et al, submitted) was performed using the GseaPreranked tool and the weighted enrichment statistics on 6387 (for C. albicans) gene sets each containing 5–500 genes. Statistical significance was estimated from 1000 permutations. Enrichment maps were constructed with Cytoscape 2.8 (http://www.cytoscape.org; [94]) and the Enrichment Map 1.1 plug-in (http://baderlab.org/Software/EnrichmentMap) using the default settings. For quantitative PCR, reverse transcription was performed using the Transcriptor High Fidelity cDNA synthesis kit from Roche. qPCR reactions were prepared using Fast-Start Sybr Green Master (Roche) on an Eppendorf Realplex master cycler and analysed by absolute quantification. The expression levels of the mRNAs were normalized to the level of ACT1 or the GAPDH encoding gene TDH3. Three independent cultures were analyzed, with two technical replicates each. qPCR primers that enable differential amplification of the ALS1 and ALS3 genes were from Green et al [95]. Sequences for all qPCR primers used in this study are listed in Table S2. For analysis of gene expression in YPD, cultures were grown under the same condition as for the microarrays analysis, to mid log phase (OD600 = 1). For assaying expression of genes under hyphal growth, strains were grown in Spider media as described in [65].
Worm infection assays
The worm-C. albicans infection assay was performed as described previously [64]. Briefly, young adult nematodes were allowed to feed for 4 h on lawns of C. albicans grown on solid BHI media (Difco) containing ampicillin (100 µg/ml), kanamycin (50 µg/ml) and streptomycin (200 µg/ml). Worms were washed with M9 media and transferred into wells of a six-well microtiter dish (Corning) containing 2 ml of liquid media (80% M9 and 20% BHI) at 60 to 80 worms per well. The plates were incubated at 25°C, and worms were qualitatively assessed at 24 h intervals for penetrative C. albicans filamentation using a DIC microscope and photographed using an Olympus IX81 microscope with the Olympus cell∧M software. The % of worms with penetrative filamentation was determined from four independent experiments at day 3 of the infection. Means and the standard error were calculated, and the p value was determined using the student t-test. The killing assays (Figure 4C) were performed three times and equivalent results were obtained.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. KimYJBjorklundSLiYSayreMHKornbergRD 1994 A multiprotein mediator of transcriptional activation and its interaction with the C-terminal repeat domain of RNA polymerase II. Cell 77 599 608
2. ConawayRCConawayJW 2011 Function and regulation of the Mediator complex. Curr Opin Genet Dev 21 225 230
3. DotsonMRYuanCXRoederRGMyersLCGustafssonCM 2000 Structural organization of yeast and mammalian mediator complexes. Proc Natl Acad Sci U S A 97 14307 14310
4. DavisJATakagiYKornbergRDAsturiasFA 2002 Structure of the yeast RNA polymerase II holoenzyme: Mediator conformation and polymerase interaction. Mol Cell 10 409 415
5. GuglielmiBvan BerkumNLKlapholzBBijmaTBoubeM 2004 A high resolution protein interaction map of the yeast Mediator complex. Nucleic Acids Res 32 5379 5391
6. CaiGImasakiTTakagiYAsturiasFJ 2009 Mediator structural conservation and implications for the regulation mechanism. Structure 17 559 567
7. van de PeppelJKettelarijNvan BakelHKockelkornTTvan LeenenD 2005 Mediator expression profiling epistasis reveals a signal transduction pathway with antagonistic submodules and highly specific downstream targets. Mol Cell 19 511 522
8. CollinsSRMillerKMMaasNLRoguevAFillinghamJ 2007 Functional dissection of protein complexes involved in yeast chromosome biology using a genetic interaction map. Nature 446 806 810
9. HolstegeFCJenningsEGWyrickJJLeeTIHengartnerCJ 1998 Dissecting the regulatory circuitry of a eukaryotic genome. Cell 95 717 728
10. BorggrefeTDavisRErdjument-BromageHTempstPKornbergRD 2002 A complex of the Srb8, -9, -10, and -11 transcriptional regulatory proteins from yeast. J Biol Chem 277 44202 44207
11. MoXKowenz-LeutzEXuHLeutzA 2004 Ras induces mediator complex exchange on C/EBP beta. Mol Cell 13 241 250
12. MalikSRoederRG 2010 The metazoan Mediator co-activator complex as an integrative hub for transcriptional regulation. Nat Rev Genet 11 761 772
13. BjorklundSGustafssonCM 2005 The yeast Mediator complex and its regulation. Trends Biochem Sci 30 240 244
14. MittlerGKremmerETimmersHTMeisterernstM 2001 Novel critical role of a human Mediator complex for basal RNA polymerase II transcription. EMBO Rep 2 808 813
15. BaekHJMalikSQinJRoederRG 2002 Requirement of TRAP/mediator for both activator-independent and activator-dependent transcription in conjunction with TFIID-associated TAF(II)s. Mol Cell Biol 22 2842 2852
16. WangGBalamotisMAStevensJLYamaguchiYHandaH 2005 Mediator requirement for both recruitment and postrecruitment steps in transcription initiation. Mol Cell 17 683 694
17. MalikSBarreroMJJonesT 2007 Identification of a regulator of transcription elongation as an accessory factor for the human Mediator coactivator. Proc Natl Acad Sci U S A 104 6182 6187
18. GuglielmiBSoutourinaJEsnaultCWernerM 2007 TFIIS elongation factor and Mediator act in conjunction during transcription initiation in vivo. Proc Natl Acad Sci U S A 104 16062 16067
19. DonnerAJEbmeierCCTaatjesDJEspinosaJM 2010 CDK8 is a positive regulator of transcriptional elongation within the serum response network. Nat Struct Mol Biol 17 194 201
20. YudkovskyNRanishJAHahnS 2000 A transcription reinitiation intermediate that is stabilized by activator. Nature 408 225 229
21. BlackJCChoiJELombardoSRCareyM 2006 A mechanism for coordinating chromatin modification and preinitiation complex assembly. Mol Cell 23 809 818
22. DingNZhouHEstevePOChinHGKimS 2008 Mediator links epigenetic silencing of neuronal gene expression with x-linked mental retardation. Mol Cell 31 347 359
23. LenstraTLBenschopJJKimTSchulzeJMBrabersNA 2011 The specificity and topology of chromatin interaction pathways in yeast. Mol Cell 42 536 549
24. ZhuXLiuBCarlstenJOBeveJNystromT 2011 Mediator influences telomeric silencing and cellular life span. Mol Cell Biol 31 2413 2421
25. ChangYWHowardSCHermanPK 2004 The Ras/PKA signaling pathway directly targets the Srb9 protein, a component of the general RNA polymerase II transcription apparatus. Mol Cell 15 107 116
26. HallbergMPolozkovGVHuGZBeveJGustafssonCM 2004 Site-specific Srb10-dependent phosphorylation of the yeast Mediator subunit Med2 regulates gene expression from the 2-microm plasmid. Proc Natl Acad Sci U S A 101 3370 3375
27. BourbonHM 2008 Comparative genomics supports a deep evolutionary origin for the large, four-module transcriptional mediator complex. Nucleic Acids Res 36 3993 4008
28. LinderTRasmussenNNSamuelsenCOChatzidakiEBaraznenokV 2008 Two conserved modules of Schizosaccharomyces pombe Mediator regulate distinct cellular pathways. Nucleic Acids Res 36 2489 2504
29. ChangYWHowardSCBudovskayaYVRineJHermanPK 2001 The rye mutants identify a role for Ssn/Srb proteins of the RNA polymerase II holoenzyme during stationary phase entry in Saccharomyces cerevisiae. Genetics 157 17 26
30. WheelerRTKombeDAgarwalaSDFinkGR 2008 Dynamic, morphotype-specific Candida albicans beta-glucan exposure during infection and drug treatment. PLoS Pathog 4 e1000227 doi:10.1371/journal.ppat.1000227
31. WangLILinYSLiuKHJongAYShenWC 2011 Cryptococcus neoformans Mediator protein Ssn8 negatively regulates diverse physiological processes and is required for virulence. PLoS ONE 6 e19162 doi:10.1371/journal.pone.0019162
32. ShimWBWoloshukCP 2001 Regulation of fumonisin B(1) biosynthesis and conidiation in Fusarium verticillioides by a cyclin-like (C-type) gene, FCC1. Appl Environ Microbiol 67 1607 1612
33. BluhmBHWoloshukCP 2006 Fck1, a C-type cyclin-dependent kinase, interacts with Fcc1 to regulate development and secondary metabolism in Fusarium verticillioides. Fungal Genet Biol 43 146 154
34. ZhouXHeyerCChoiYEMehrabiRXuJR 2010 The CID1 cyclin C-like gene is important for plant infection in Fusarium graminearum. Fungal Genet Biol 47 143 151
35. BlankenshipJRFanningSHamakerJJMitchellAP 2010 An extensive circuitry for cell wall regulation in Candida albicans. PLoS Pathog 6 e1000752 doi:10.1371/journal.ppat.1000752
36. NelsonCGotoSLundKHungWSadowskiI 2003 Srb10/Cdk8 regulates yeast filamentous growth by phosphorylating the transcription factor Ste12. Nature 421 187 190
37. CooperKFMalloryMJStrichR 1999 Oxidative stress-induced destruction of the yeast C-type cyclin Ume3p requires phosphatidylinositol-specific phospholipase C and the 26S proteasome. Mol Cell Biol 19 3338 3348
38. ThakurJKArthanariHYangFPanSJFanX 2008 A nuclear receptor-like pathway regulating multidrug resistance in fungi. Nature 452 604 609
39. PaulSSchmidtJAMoye-RowleyWS 2011 Regulation of the CgPdr1 transcription factor from the pathogen Candida glabrata. Eukaryot Cell 10 187 197
40. FichtnerLSchulzeFBrausGH 2007 Differential Flo8p-dependent regulation of FLO1 and FLO11 for cell-cell and cell-substrate adherence of S. cerevisiae S288c. Mol Microbiol 66 1276 1289
41. ArbourMEppEHoguesHSellamALacroixC 2009 Widespread occurrence of chromosomal aneuploidy following the routine production of Candida albicans mutants. FEMS Yeast Res 9 1070 1077
42. MiklosISzilagyiZWattSZilahiEBattaG 2008 Genomic expression patterns in cell separation mutants of Schizosaccharomyces pombe defective in the genes sep10 (+) and sep15 (+) coding for the Mediator subunits Med31 and Med8. Mol Genet Genomics 279 225 238
43. KoschubsTSeizlMLariviereLKurthFBaumliS 2009 Identification, structure, and functional requirement of the Mediator submodule Med7N/31. EMBO J 28 69 80
44. SubramanianATamayoPMoothaVKMukherjeeSEbertBL 2005 Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A 102 15545 15550
45. MoothaVKLindgrenCMErikssonKFSubramanianASihagS 2003 PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat Genet 34 267 273
46. StarkCBreitkreutzBJRegulyTBoucherLBreitkreutzA 2006 BioGRID: a general repository for interaction datasets. Nucleic Acids Res 34 D535 539
47. MarcilAGadouryCAshJZhangJNantelA 2008 Analysis of PRA1 and its relationship to Candida albicans-macrophage interactions. Infect Immun 76 4345 4358
48. SpieringMJMoranGPChauvelMMaccallumDMHigginsJ 2010 Comparative transcript profiling of Candida albicans and Candida dubliniensis identifies SFL2, a C. albicans gene required for virulence in a reconstituted epithelial infection model. Eukaryot Cell 9 251 265
49. FradinCDe GrootPMacCallumDSchallerMKlisF 2005 Granulocytes govern the transcriptional response, morphology and proliferation of Candida albicans in human blood. Mol Microbiol 56 397 415
50. KadoshDJohnsonAD 2005 Induction of the Candida albicans filamentous growth program by relief of transcriptional repression: a genome-wide analysis. Mol Biol Cell 16 2903 2912
51. NantelADignardDBachewichCHarcusDMarcilA 2002 Transcription profiling of Candida albicans cells undergoing the yeast-to-hyphal transition. Mol Biol Cell 13 3452 3465
52. HarcusDNantelAMarcilARigbyTWhitewayM 2004 Transcription profiling of cyclic AMP signaling in Candida albicans. Mol Biol Cell 15 4490 4499
53. BachewichCNantelAWhitewayM 2005 Cell cycle arrest during S or M phase generates polarized growth via distinct signals in Candida albicans. Mol Microbiol 57 942 959
54. CotePHoguesHWhitewayM 2009 Transcriptional analysis of the Candida albicans cell cycle. Mol Biol Cell 20 3363 3373
55. EnjalbertBNantelAWhitewayM 2003 Stress-induced gene expression in Candida albicans: absence of a general stress response. Mol Biol Cell 14 1460 1467
56. ButlerGRasmussenMDLinMFSantosMASakthikumarS 2009 Evolution of pathogenicity and sexual reproduction in eight Candida genomes. Nature 459 657 662
57. MulhernSMLogueMEButlerG 2006 Candida albicans transcription factor Ace2 regulates metabolism and is required for filamentation in hypoxic conditions. Eukaryot Cell 5 2001 2013
58. HoyerLLGreenCBOhSHZhaoX 2008 Discovering the secrets of the Candida albicans agglutinin-like sequence (ALS) gene family–a sticky pursuit. Med Mycol 46 1 15
59. ArgimonSWishartJALengRMacaskillSMavorA 2007 Developmental regulation of an adhesin gene during cellular morphogenesis in the fungal pathogen Candida albicans. Eukaryot Cell 6 682 692
60. KimSWolyniakMJStaabJFSundstromP 2007 A 368-base-pair cis-acting HWP1 promoter region, HCR, of Candida albicans confers hypha-specific gene regulation and binds architectural transcription factors Nhp6 and Gcf1p. Eukaryot Cell 6 693 709
61. HeilmannCJSorgoAGSiliakusARDekkerHLBrulS 2011 Hyphal induction in the human fungal pathogen Candida albicans reveals a characteristic wall protein profile. Microbiology 157 2297 2307
62. HomannORDeaJNobleSMJohnsonAD 2009 A phenotypic profile of the Candida albicans regulatory network. PLoS Genet 5 e1000783 doi:10.1371/journal.pgen.1000783
63. BregerJFuchsBBAperisGMoyTIAusubelFM 2007 Antifungal chemical compounds identified using a C. elegans pathogenicity assay. PLoS Pathog 3 e18 doi:10.1371/journal.ppat.0030018
64. PelegAYTampakakisEFuchsBBEliopoulosGMMoelleringRCJr 2008 Prokaryote-eukaryote interactions identified by using Caenorhabditis elegans. Proc Natl Acad Sci U S A 105 14585 14590
65. NobileCJAndesDRNettJESmithFJYueF 2006 Critical role of Bcr1-dependent adhesins in C. albicans biofilm formation in vitro and in vivo. PLoS Pathog 2 e63 doi:10.1371/journal.ppat.0020063
66. NobileCJMitchellAP 2005 Regulation of cell-surface genes and biofilm formation by the C. albicans transcription factor Bcr1p. Curr Biol 15 1150 1155
67. ReynoldsTBFinkGR 2001 Bakers' yeast, a model for fungal biofilm formation. Science 291 878 881
68. LariviereLGeigerSHoeppnerSRotherSStrasserK 2006 Structure and TBP binding of the Mediator head subcomplex Med8-Med18-Med20. Nat Struct Mol Biol 13 895 901
69. LariviereLSeizlMvan WageningenSRotherSvan de PaschL 2008 Structure-system correlation identifies a gene regulatory Mediator submodule. Genes Dev 22 872 877
70. CarreraIJanodyFLeedsNDuveauFTreismanJE 2008 Pygopus activates Wingless target gene transcription through the mediator complex subunits Med12 and Med13. Proc Natl Acad Sci U S A 105 6644 6649
71. ShahiPGulshanKNaarAMMoye-RowleyWS 2010 Differential roles of transcriptional mediator subunits in regulation of multidrug resistance gene expression in Saccharomyces cerevisiae. Mol Biol Cell 21 2469 2482
72. KellyMTMacCallumDMClancySDOddsFCBrownAJ 2004 The Candida albicans CaACE2 gene affects morphogenesis, adherence and virulence. Mol Microbiol 53 969 983
73. MehtaSMiklosISipiczkiMSenguptaSSharmaN 2009 The Med8 mediator subunit interacts with the Rpb4 subunit of RNA polymerase II and Ace2 transcriptional activator in Schizosaccharomyces pombe. FEBS Lett 583 3115 3120
74. NobileCJSchneiderHANettJESheppardDCFillerSG 2008 Complementary adhesin function in C. albicans biofilm formation. Curr Biol 18 1017 1024
75. AnsariSAHeQMorseRH 2009 Mediator complex association with constitutively transcribed genes in yeast. Proc Natl Acad Sci U S A 106 16734 16739
76. FanXChouDMStruhlK 2006 Activator-specific recruitment of Mediator in vivo. Nat Struct Mol Biol 13 117 120
77. FanXStruhlK 2009 Where does mediator bind in vivo? PLoS ONE 4 e5029 doi:10.1371/journal.pone.0005029
78. AndrauJCvan de PaschLLijnzaadPBijmaTKoerkampMG 2006 Genome-wide location of the coactivator Mediator: binding without activation and transient Cdk8 interaction on DNA. Mol Cell 22 179 192
79. LiFPalecekSP 2003 EAP1, a Candida albicans gene involved in binding human epithelial cells. Eukaryot Cell 2 1266 1273
80. BrucknerSMoschHU 2012 Choosing the right lifestyle: adhesion and development in Saccharomyces cerevisiae. FEMS Microbiol Rev 36 25 58
81. BauerJWendlandJ 2007 Candida albicans Sfl1 suppresses flocculation and filamentation. Eukaryot Cell 6 1736 1744
82. MuradAMLengPStraffonMWishartJMacaskillS 2001 NRG1 represses yeast-hypha morphogenesis and hypha-specific gene expression in Candida albicans. EMBO J 20 4742 4752
83. CaoFLaneSRanigaPPLuYZhouZ 2006 The Flo8 transcription factor is essential for hyphal development and virulence in Candida albicans. Mol Biol Cell 17 295 307
84. KurasLBorggrefeTKornbergRD 2003 Association of the Mediator complex with enhancers of active genes. Proc Natl Acad Sci U S A 100 13887 13891
85. ConlanRSTzamariasD 2001 Sfl1 functions via the co-repressor Ssn6-Tup1 and the cAMP-dependent protein kinase Tpk2. J Mol Biol 309 1007 1015
86. HalmeABumgarnerSStylesCFinkGR 2004 Genetic and epigenetic regulation of the FLO gene family generates cell-surface variation in yeast. Cell 116 405 415
87. VerstrepenKJFinkGR 2009 Genetic and epigenetic mechanisms underlying cell-surface variability in protozoa and fungi. Annu Rev Genet 43 1 24
88. WilsonRBDavisDMitchellAP 1999 Rapid hypothesis testing with Candida albicans through gene disruption with short homology regions. J Bacteriol 181 1868 1874
89. JinYSamaranayakeLPSamaranayakeYYipHK 2004 Biofilm formation of Candida albicans is variably affected by saliva and dietary sugars. Arch Oral Biol 49 789 798
90. JinYYipHKSamaranayakeYHYauJYSamaranayakeLP 2003 Biofilm-forming ability of Candida albicans is unlikely to contribute to high levels of oral yeast carriage in cases of human immunodeficiency virus infection. J Clin Microbiol 41 2961 2967
91. DagleyMJGentleIEBeilharzTHPettolinoFADjordjevicJT 2011 Cell wall integrity is linked to mitochondria and phospholipid homeostasis in Candida albicans through the activity of the post-transcriptional regulator Ccr4-Pop2. Mol Microbiol 79 968 989
92. SkrzypekMSArnaudMBCostanzoMCInglisDOShahP 2010 New tools at the Candida Genome Database: biochemical pathways and full-text literature search. Nucleic Acids Res 38 D428 D432
93. HusseinBHuangHGloryAOsmaniAKaminskyjS 2011 G1/S transcription factor orthologues Swi4p and Swi6p are important but not essential for cell proliferation and influence hyphal development in the fungal pathogen Candida albicans. Eukaryot Cell 10 384 397
94. SmootMEOnoKRuscheinskiJWangPLIdekerT 2011 Cytoscape 2.8: new features for data integration and network visualization. Bioinformatics 27 431 432
95. GreenCBZhaoXYeaterKMHoyerLL 2005 Construction and real-time RT-PCR validation of Candida albicans PALS-GFP reporter strains and their use in flow cytometry analysis of ALS gene expression in budding and filamenting cells. Microbiology 151 1051 1060
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2012 Číslo 4
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- A Coordinated Interdependent Protein Circuitry Stabilizes the Kinetochore Ensemble to Protect CENP-A in the Human Pathogenic Yeast
- Coordinate Regulation of Lipid Metabolism by Novel Nuclear Receptor Partnerships
- Defective Membrane Remodeling in Neuromuscular Diseases: Insights from Animal Models
- Formation of Rigid, Non-Flight Forewings (Elytra) of a Beetle Requires Two Major Cuticular Proteins
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy