
Regulation of Budding Yeast Mating-Type Switching Donor Preference by the FHA Domain of Fkh1
During Saccharomyces cerevisiae mating-type switching, an HO endonuclease-induced double-strand break (DSB) at MAT is repaired by recombining with one of two donors, HMLα or HMRa, located at opposite ends of chromosome III. MATa cells preferentially recombine with HMLα; this decision depends on the Recombination Enhancer (RE), located about 17 kb to the right of HML. In MATα cells, HML is rarely used and RE is bound by the MATα2-Mcm1 corepressor, which prevents the binding of other proteins to RE. In contrast, in MATa cells, RE is bound by multiple copies of Fkh1 and a single copy of Swi4/Swi6. We report here that, when RE is replaced with four LexA operators in MATa cells, 95% of cells use HMR for repair, but expression of a LexA-Fkh1 fusion protein strongly increases HML usage. A LexA-Fkh1 truncation, containing only Fkh1's phosphothreonine-binding FHA domain, restores HML usage to 90%. A LexA-FHA-R80A mutant lacking phosphothreonine binding fails to increase HML usage. The LexA-FHA fusion protein associates with chromatin in a 10-kb interval surrounding the HO cleavage site at MAT, but only after DSB induction. This association occurs even in a donorless strain lacking HML. We propose that the FHA domain of Fkh1 regulates donor preference by physically interacting with phosphorylated threonine residues created on proteins bound near the DSB, thus positioning HML close to the DSB at MAT. Donor preference is independent of Mec1/ATR and Tel1/ATM checkpoint protein kinases but partially depends on casein kinase II. RE stimulates the strand invasion step of interchromosomal recombination even for non-MAT sequences. We also find that when RE binds to the region near the DSB at MATa then Mec1 and Tel1 checkpoint kinases are not only able to phosphorylate histone H2A (γ-H2AX) around the DSB but can also promote γ-H2AX spreading around the RE region.
Published in the journal:
. PLoS Genet 8(4): e32767. doi:10.1371/journal.pgen.1002630
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002630
Summary
During Saccharomyces cerevisiae mating-type switching, an HO endonuclease-induced double-strand break (DSB) at MAT is repaired by recombining with one of two donors, HMLα or HMRa, located at opposite ends of chromosome III. MATa cells preferentially recombine with HMLα; this decision depends on the Recombination Enhancer (RE), located about 17 kb to the right of HML. In MATα cells, HML is rarely used and RE is bound by the MATα2-Mcm1 corepressor, which prevents the binding of other proteins to RE. In contrast, in MATa cells, RE is bound by multiple copies of Fkh1 and a single copy of Swi4/Swi6. We report here that, when RE is replaced with four LexA operators in MATa cells, 95% of cells use HMR for repair, but expression of a LexA-Fkh1 fusion protein strongly increases HML usage. A LexA-Fkh1 truncation, containing only Fkh1's phosphothreonine-binding FHA domain, restores HML usage to 90%. A LexA-FHA-R80A mutant lacking phosphothreonine binding fails to increase HML usage. The LexA-FHA fusion protein associates with chromatin in a 10-kb interval surrounding the HO cleavage site at MAT, but only after DSB induction. This association occurs even in a donorless strain lacking HML. We propose that the FHA domain of Fkh1 regulates donor preference by physically interacting with phosphorylated threonine residues created on proteins bound near the DSB, thus positioning HML close to the DSB at MAT. Donor preference is independent of Mec1/ATR and Tel1/ATM checkpoint protein kinases but partially depends on casein kinase II. RE stimulates the strand invasion step of interchromosomal recombination even for non-MAT sequences. We also find that when RE binds to the region near the DSB at MATa then Mec1 and Tel1 checkpoint kinases are not only able to phosphorylate histone H2A (γ-H2AX) around the DSB but can also promote γ-H2AX spreading around the RE region.
Introduction
Saccharomyces mating-type switching occurs through a DSB-initiated intrachromosomal gene conversion event at MAT, using one of two donors on chromosome III, HML and HMR (Figure 1A) [1]–[3]. Switching is initiated by expression of the site-specific HO endonuclease that cleaves only one site in the yeast genome, MATa or MATα. The unexpressed mating-type genes in HMLα and HMRa also contain HO cleavage sites, but they are not cut because these regions are heterochromatic [4]–[6]. Although either HMLα or HMRa can be used to repair a DSB at MAT, there is a strong mating type-dependent preference for the choice of the two donors. In MATa cells, HMLα is preferentially chosen for repair, about 85–90% of the time, whereas MATα cells strongly prefer HMRa, about 95% [3], [7]–[9]. Donor preference is not altered if the mating-type genes encoded in the Y region are changed, e.g. if HMR carries Yα instead of Ya or if HML is replaced with HMR [7], [8].

Donor preference in MATa depends on an approximately 275-bp Recombination Enhancer (RE), located 17 kb to the right of HML [10], [11]. One important aspect of donor preference is that MATa cells activate a large (∼40 kb) region near the left end of chromosome III, so that a donor within this region is strongly preferred over HMR [8]. RE is responsible for this activation along the entire left arm of chromosome III [11], [12]. Donor preference does not change if the cis-acting silencer sequences around HML or HMR are removed [13]. In addition, RE is not limited to the special features of MAT switching. If a leu2 allele is inserted in place of HML, its success in recombining with a different leu2 allele, either near MAT or even on another chromosome, is 20–30 times higher in MATa than in MATα and is RE-dependent [8], [12].
RE is “portable;” that is, it will work in other chromosome contexts. When HML, HMR and MATa are all inserted on chromosome V, HML usage increases significantly when RE is inserted nearby [12]. In addition, in MATa cells where RE promotes HML, the usage of HMR can be markedly increased by placing a second RE near HMR [11], [12].
In MATα cells, RE is inactivated by binding of the Matα2-Mcm1 repressor complex, which leads to formation of highly organized nucleosomes covering the RE region but not extending into adjacent gene regions [8], [10], [14]. In MATa cells, RE exhibits several nuclease hypersensitive sites when Mcm1 binds RE in the absence of the Matα2 protein (which is not expressed in MATa cells). In addition to the Matα2-Mcm1 operator region, RE is composed of several evolutionarily conserved chromatin domains [14], several of which were shown to contain putative binding sites for the Fkh1 transcription factor [15]. A conserved SCB (Swi4/Swi6 cell cycle box) is also present in Region C of RE [16]. Both Fkh1 and Swi4/Swi6 regulate donor preference by binding to RE in MATa cells [15]–[17]. Despite the presence of these transcription factors, there are no open reading frames adjacent to RE, although there is an adjacent noncoding RNA [18]. The DNA repair proteins Ku70 and Ku80 have a small effect on MATa donor preference that may be caused by the role of these proteins in localizing HML to the nuclear periphery [19]. Deleting the Chl1 helicase also causes a small reduction of MATa donor preference without affecting MATα choice [16], [20].
Despite the identification of several proteins that bind to RE, it is still not clear how RE regulates donor preference. Previously we showed that RE could be deleted and replaced with small modules derived from RE. Notably 4 tandem copies of a 22-bp sequence containing a putative Fkh1 binding site were sufficient to increase HML usage to >60% (where the use of HML in REΔ is 5%); this increased preference for HML is abolished in fkh1Δ [15]. To further explore the mechanism of RE regulation, we replaced RE with four LexA operators and found that a LexA-Fkh1 fusion strongly promotes HML usage. Using this system, we dissect Fkh1 and find out that RE activity depends on the phosphothreonine binding motif of the FHA domain of Fkh1 and not on its forkhead domain. We show that LexA-FHAFkh1 becomes associated with the chromatin surrounding the MAT only after DSB induction. This interaction is seen even in a donorless strain, demonstrating that the FHA-mediated regulation is a break-dependent but repair-independent process. MATa donor preference is partially dependent on casein kinase II but not on two checkpoint kinases, Mec1 and Tel1. We propose that the FHAFkh1 domain regulates donor preference by physically interacting with phosphorylated threonines on histones or other bound proteins surrounding the DSB during mating-type switch.
Results
Measuring Donor Preference by Southern Blot or a PCR-Based Assay
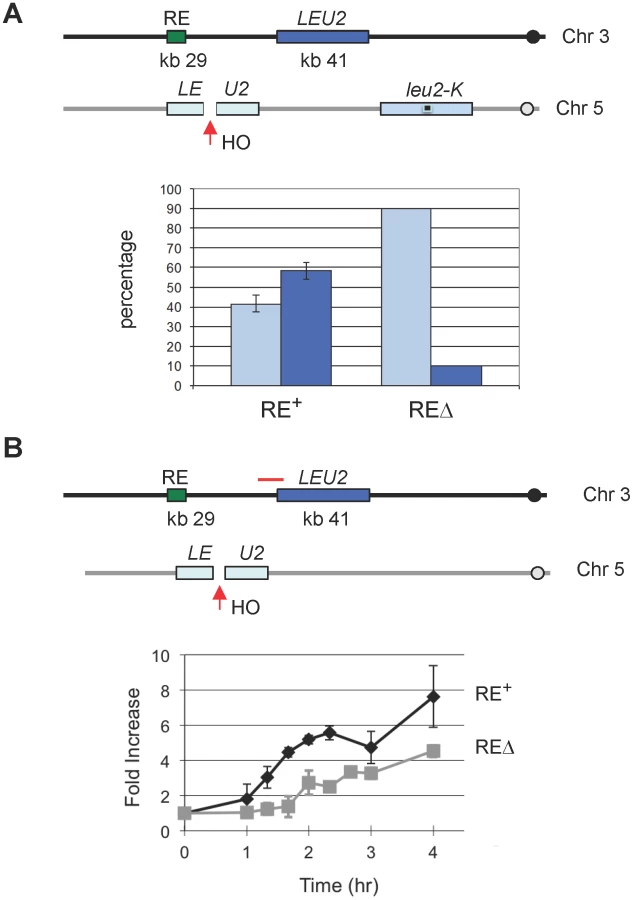
All strains in this study are derived from XW652 [11], which carries HMLα, MATa and HMRα-B on chromosome III (Figure 1A). HMRα-B contains a single base pair change that creates a BamHI site [8]. After galactose-induced expression of HO, MATa can be repaired to MATα or MATα-B, using HMLα or HMRα-B, respectively. Following HO induction for 60 min, HO expression was repressed by the addition of 2% dextrose and the ratio of switching to MATα or MATα-B was checked after 24 h. Donor preference could be measured either by Southern blot [8] or by a PCR-based assay in which the combination of MATα or MATα-B PCR products is digested with BamHI (Figure 1B). PCR-based assay showed 85% usage of HMLα for XW652 but ≤10% for RE-deleted XW676 (Figure 1C).
A LexA System to Study the Regulation of Donor Preference
Fkh1 is involved in the regulation of donor preference through direct interaction with RE [15], [16]. To further explore the role of Fkh1, we constructed a strain ECY406 by replacing RE with four LexA operators (Figure 2A). In an otherwise wild type background, HML usage in ECY406 was less than 5% as expected for a deletion of RE (Figure 2B). We then constructed a plasmid pEC16 that constitutively expresses a LexA-Fkh1 fusion protein from an ADH1 promoter of pAT4 [21]. The LexA-Fkh1 sequences from pEC16 were stably integrated at the arg5,6 locus of ECY406 to generate a new strain ECY457 (Figure 2A). Expression of LexA-Fkh1 in ECY457 was able to up-regulate donor preference to around 32% presumably by binding to four LexA operators replacing RE (Figure 2B), whereas the use of HML was less than 5% when LexA alone was expressed (data not shown). This result demonstrates that regulation of donor preference by Fkh1 does not require the binding of Mcm1 or Swi4/Swi6 to their specific sites in the normal RE sequences. We noted further that the Fkh1 moiety in the LexA-Fkh1 fusion remained functional even with normal RE, as it could complement a fkh1Δ mutant in YJL017 by up-regulating donor preference to 68% (Figure 2C).

The Phosphothreonine-Binding FHA Domain of Fkh1 Is Responsible for Donor Preference
Fkh1 contains two conserved domains: a forkhead-associated (FHA) and a forkhead DNA binding domain (Figure 3A) [22], [23]. To understand roles of different domains of Fkh1 in the regulation of donor preference, we prepared three plasmid constructs by fusing LexA of pAT4 with different regions of Fkh1: pJL4 for LexA-FHA (aa 1–230 of Fkh1), pJL5 for LexA-interdomain (aa 163–302), and pJL6 for LexA-forkhead (aa 231–484) (Figure 3A). The LexA fused sequences from these plasmids were integrated at arg5,6 locus of ECY406 to generate strains YJL019, YJL020, and YJL021, respectively (Figure 3A). These three strains and ECY457 (Figure 2A) all have a wild-type Fkh1, which is not functional in donor preference because Fkh1 cannot bind to REΔ::LexABD4. Southern blots revealed that only YJL019 could re-establish donor preference to 90%, whereas YJL020 and YJL021 failed to increase HML usage (Figure 3B). This result suggests that the FHA domain may play a critical role in the regulation of donor preference.

We noted that donor preference regulated by LexA-FHAFkh1 (90% donor preference for YJL019; Figure 3B) was much higher than that by LexA-Fkh1 (32% donor preference for ECY457; Figure 2B). We suggest two possible explanations for this difference. First, two DNA binding domains (LexA and the forkhead DNA binding domain) are present in LexA-Fkh1, whereas only one (LexA) is present in LexA-FHAFkh1. Therefore, the LexA-Fkh1 fusion protein likely binds multiple sites in yeast genome, which could mean that less fusion protein is available for regulating donor preference. In contrast, because there is only one DNA binding domain for LexA-FHAFkh1, all fusion protein will be available for donor preference regulation. A second possible reason is that the FHAFkh1 domain is more exposed in LexA-FHAFkh1 than in LexA-Fkh1 when both fusion proteins bind to four LexA operators replacing RE. The presence of a forkhead domain in LexA-Fkh1 could interfere with regulation of the FHAFkh1 domain in donor preference, whereas this kind of interference is not present in LexA-FHAFkh1.
The FHA (forkhead-associated) domain is a small protein module that can preferentially bind to phosphothreonine residues on proteins [22], [24], [25]. FHA domains have been found in a wide range of proteins, such as kinases, phosphatases and transcription factors [23], [26]. To confirm that the FHAFkh1 domain was responsible for increasing HML usage, LexA-FHA-R80A from pJL8 was integrated into the arg5,6 locus of ECY406 to generate a strain YJL094 (Figure 3A). Preferential usage of HML was completely abolished using LexA-FHA-R80A (Figure 3C), which carried a non-functional FHA domain [22], [23]. Thus, the phosphothreonine-binding motif of the FHA domain plays a critical role in the regulation of donor preference.
The FHA Domain of Fkh1 Physically Interacts with the MAT Region after DSB Induction
We employed Chromatin Immunoprecipitation (ChIP) to ask if LexA-FHAFkh1 could associate with the region around MAT before or after induction of a DSB. Using an anti-LexA antibody, we showed that LexA-fused FHAFkh1 physically interacted with the MAT region after DSB induction in a strain lacking HML and HMR (Figure 4A), so that DSBs could not be repaired by homologous recombination. We observed a >10-fold increase in ChIP signals within about 5 kb on either side of the HO cleavage site at the MAT, whereas no significant signal could be detected using primer pairs that amplify regions further away from the HO site (Figure 4B). Therefore, the LexA-FHAFkh1 fusion protein physically interacted with the DSB-cut MAT through a repair-independent mechanism, which suggests that LexA-FHAFkh1 or RE can be used to stimulate recombination between any two homologous sequences in budding yeast [11].

We note that the localization of LexA-FHAFkh1 binding is quite different from the roughly 50-kb phosphorylation of histone H2A-S129 (γ-H2AX) on either side of the DSB [27], [28], although it is similar to a second damage-induced modification, which is the casein kinase II-dependent phosphorylation of H4-S1 (Figure 4D) [29].
Given that RE activity was completely abolished in the strain YJL094 carrying LexA-FHA-R80A (Figure 3C), it was not unexpected that the ChIP assay showed no physical interaction between LexA-FHA-R80A and the MAT region after DSB induction (Figure 4C); However, the LexA-FHA-R80A fusion protein still strongly associated with REΔ::LexABD4 likely due to the presence of the LexA domain (Figure 4C). These data strongly support the idea that the FHA domain of Fkh1 regulates donor preference by physically interacting with the MAT region during mating-type switch, and these interactions fully depend on the phosphothreonine binding motif of the FHAFkh1 domain.
RE Accelerates the Rate of DSB–Induced Ectopic Recombination for Non-MAT Sequences
Because the FHAFkh1 domain regulates donor preference via a repair-independent but break-dependent mechanism, it suggests that FHAFkh1 domain or RE can be used to facilitate recombination between any homologous sequences in yeast genome. Previously we showed that RE stimulated leu2 heteroallele spontaneous recombination when one of the alleles was situated in place of HML [11]. In that case, the nature and position of the initiating DNA lesions were unknown. Here we integrated a leu2::HOcs construct at can1 locus on chromosome V, so HO-induced DSBs can recombine with a LEU2 locus placed near RE on chromosome III (Figure 5) [30]. In one assay, LEU2 on chromosome III could be used as a donor to repair HO-induced DSBs on chromosome V in competition with a leu2-K donor inserted at ura3, which is 85 kb from the leu2::HOcs (Figure 5A). The leu2-K allele was created by ablating KpnI site in LEU2 [31]. As shown in Figure 5A, the proportion of repair events using the interchromosomal donor was more than 50% when RE was present but fell to less than 10% when RE was deleted. In a second assay, the LEU2 on chromosome III was the only possible donor for DSB repair. This construct allowed us to ask whether RE stimulated recombination by facilitating the earliest step, the search for homology by Rad51 recombinase bound to the resected end of the DSB. We measured the time at which Rad51 became associated with the donor (i.e. when strand invasion had occurred) by a ChIP assay analogous to that used to assay strand invasion kinetics during MAT switching [32], [33]. As seen in Figure 5B, the kinetics of Rad51 association with the LEU2 donor was significantly faster when RE was present. The presence of RE also assured that the proportion of cells that completed repair was 72% compared to 37% when RE was deleted. The percentage of completed repair was determined by comparing survival on galactose plates with that on dextrose plates where HO was not induced.
γ-H2AX Formation at RE in MATa Cells Provides Additional Evidence of Direct RE-to-MAT Contact
γ-H2AX rapidly forms around the site of a DSB, dependent on either Mec1 or Tel1 checkpoint protein kinase [27], [28]. If RE bound to regions around the DSB, would γ-H2AX also form around RE region? To address this question, we used ChIP with anti-γ-H2AX antibody to examine the phosphorylation of histones around RE following initiation of a DSB. γ-H2AX formed over a large domain around MAT following the induction of a DSB within 15–60 min (Figure 6A). Surprisingly, γ-H2AX also appeared around RE at 1 hr after HO induction in MATa cells. As predicted, there was no similar modification around RE in MATα cells, where RE is repressed (Figure 6B). Moreover, the kinetics of γ-H2AX modification around RE was slower than around MAT, consistent with the idea that RE first had to be recruited to the DSB before this modification could take place (Figure 6A, 6C). Finally, by using both mec1Δ sml1Δ and tel1Δ derivatives of JKM139, we showed that either checkpoint kinase was capable of carrying out γ-H2AX modification around RE (Figure 6D). These data provide additional supporting evidence of a direct RE-to-MAT contact after DSB induction and support the model that the binding of RE to MAT is the basis of bringing HML into close proximity. In addition, these data show for the first time that a region not suffering a DSB can be modified by both checkpoint kinases if this region is brought close to the DSB site.

Are Histones the Target of FHAFkh1 Domain in Donor Preference Regulation?
Our data strongly argue that the FHA domain of Fkh1, clustered at the normal RE or REΔ::LexABD4, interacts with phosphorylated residues in the region surrounding the DSB. The most obvious candidates are histones that are phosphorylated after DSB induction, including H4-S1 [29] and histone H2A-S129 (γ-H2AX). The possibility that H4-S1 could be involved was made more attractive by our finding that this modification is confined to the first 10 kb around a DSB, much more restricted than γ-H2AX (Figure 4D). We constructed a strain YJL102, carrying the h4-S1A in HHF2 and deleted for HHF1; however this alteration had no effect on donor preference (Figure 7A). In addition, phosphorylation of H2A-S122, H2A-T126 and H2A-S129 have been implicated after MMS-induced DNA damage [34]. To test these H2A modifications, we constructed a strain YJL121 by deleting endogenous HTA1-HTB1 and HTA2-HTB2 and complementing by a plasmid carrying hta1-S122A-T126A-S129A-HTB1, but donor preference was not affected (Figure 7A).

We have directly tested whether post-translational modifications of the N-terminal tail of histones H3 or H4 are implicated in donor preference. In addition to H4-S1, several other sites have been reported to be phosphorylated during the cell cycle, such as H3-T3, H3-S10 and H3-S28 [35]–[37], which might also be targets for modification after a chromosome break. In particular, we constructed a strain YJK340, in which HHF1-HHT1 was deleted with NAT. Then, the remaining copy of H3 gene was modified to carry a deletion of the first 32 amino acids or HHF2 was modified to lack the first 16 amino acids of histone H4. We found that the H3 tail deletion unsilenced HML but not HMR (i.e. cells became non-mating by expressing both HMLα and MATa); hence we replaced the Yα sequences at HML with HPH as previously described [13]. This modification also deleted part of the HO cleavage site at HML, so only MATa would be cleaved by HO. When HO was induced at MAT, there was no change in donor preference, as 73 of the 82 (89%) switched products were derived from hml::HPH and only 9 (11%) were MATα-B, derived from HMRα-B. In the case of the H4 N-terminal truncation, both HMLα and HMRα-B become unsilenced [38]; thus to look at donor preference, we replaced HML's Yα with HPH and HMR's Yα-B with KAN [13]. Among 39 colonies that switched from MATa, 34 (87%) used hml::HPH whereas only 5 (13%) recombined with hmr::KAN. Therefore, deleting the tail of histone H3 or H4 had no effect on donor preference in MATa cells.
Fkh1-Directed Regulation Depends on Casein Kinase II, but Not on Mec1/Tel1
Although Mec1 and Tel1 can phosphorylate histone H2A in the region surrounding RE when it is brought in conjunction with MAT, these checkpoint kinases are not responsible for promoting MAT donor preference. We constructed a strain YJL054 (mec1Δ tel1Δ sml1Δ) derived from XW652. We noted that because Mec1 or Tel1 was required for efficient clipping of the Ya tail to enable the completion of switching to MATα or MATα-B [39], there was a strong reduction in the switching efficiency (data not shown), but the proportion of MATα to MATα-B was unaltered in YJL054 (Figure 7B). This conclusion that Mec1 and Tel1 are not involved in the regulation of donor preference was further supported by our data that donor preference was not altered in YJL121, in which histone H2A-S129 was mutated to alanine (Figure 7A).
Casein kinase II phosphorylates serine 1 (S1) of histone H4 after exposure to MMS - and phleomycin-induced DSBs [29] and after HO-induced DSBs (Figure 4D). Casein kinase II is required for cell cycle progression in budding yeast and essential for cell viability [40]. We constructed a strain lacking the chromosomal CKA1 and CKA2 genes but carrying a pRS315 plasmid with a temperature-sensitive cka2-8 allele (Figure 7B). Cells were grown overnight at the permissive temperature of 25°C and then shifted to the restrictive temperature of 37°C. Inactivation of Cka2 leads to 43% use of HML (YJL119) compared to 87% donor preference in control YJL019 (Figure 7B). This result indicates that casein kinase II activity is required for Fkh1-dependent regulation of donor preference. Because the N-terminal truncation of H4 (including H4-S1) has no effect on HML usage, it is likely that casein kinase II phosphorylates some other targets, on a histone or another protein, which is involved in donor preference regulation. However, the fact that 43% donor preference is still significantly higher than 10% observed in RE-deleted strains (Figure 1C) suggests that multiple kinases may be involved in the regulation of donor preference.
Discussion
We have shown that the phosphothreonine binding motif of the FHA domain of Fkh1 plays a critical role in the regulation of donor preference (Figure 3). A strong physical association between the FHAFkh1 domain bound at the RE region and MAT is readily seen, but only after a DSB is induced. This interaction is independent of the presence of an adjacent homologous HML donor (Figure 4). Conversely, the region surrounding RE can be phosphorylated by Mec1 and Tel1 kinases only after DSB induction in MATa but not in MATα strains (Figure 6), again suggesting that these regions can come into physical contact when there is a DSB at MAT and RE is active.
RE's activity does not depend on any of the special features of MAT switching such as HML or HMR silencing [13] or HO cleavage [11], [15]. Consequently RE is able to improve the use of an ectopic donor in repairing a DSB on a different chromosome. Normally, a DSB will be preferentially repaired by a donor on the same chromosome in competition with an ectopic donor, but if the ectopic donor is located near RE, more than half of gene conversions use the interchromosomal donor (Figure 5A). Although our data and those from others show that HML is not constitutively much closer to MATa than HMR is (i.e. in the absence of HO cleavage) [41]–[43], the data we present here suggest that such a reorganization will occur after a DSB is created.
Taken together, our data suggest a simple model for RE action (Figure 7C). After the induction of a DSB, casein kinase II and possibly other kinases modify some proteins bound near the DSB. These modifications, most likely phosphothreonines, are clustered near the DSB and can be bound by FHAFkh1 domains tethered at RE. This binding effectively tethers HML within about 20 kb of the DSB whereas HMR remains 100 kb away. Thermodynamic considerations argue that this close proximity is sufficient to explain why HML should be used 90% of the time for DSB repair in MATa cells [13]. This model also explains how RE can act over a long region of the left arm of chromosome III [8], although with diminishing effect [12], by this tethering mechanism.
The model we propose argues that RE should be portable and able to stimulate the use of any homologous donor in a DSB repair mechanism. Our previous work has shown that RE is portable, as it is able to activate HML use when both are inserted on chromosome V [12]. Moreover, if a copy of RE is inserted near HMR in a MATa strain that also has RE near HML, then HMR usage is increased to about 50% (E.C., S.-Y. Tay and J.E.H., unpublished). The ectopic recombination experiment presented here shows that RE can act efficiently on non-MAT sequences for DSB repair (Figure 5A).
We note that we have previously shown that RE could stimulate spontaneous recombination between leu2 heteroalleles when one of them was located close to the RE [11], [12]. The results we report here suggest that a large proportion of spontaneous recombination events may be triggered by DSBs or that the same phosphorylated protein attracting the attention of RE during DSB repair also accumulates at the lesions that stimulate spontaneous recombination.
At present, we have not yet identified the phosphothreonine target for the FHA domain of Fkh1. We have ruled out a number of candidates, including γ-H2AX, N-terminal tails of histones H3 and H4, as well as Mre11 and Sae2, two proteins involved in DSB end-binding and initiating 5′ to 3′ resection (C.-S. L., J.E.H., unpublished observations). Studies using peptide libraries and immunoprecipitation of the FHAFkh1 domain after DSB induction are underway.
Aparicio group has recently made the intriguing finding that Fkh1 and Fkh2 proteins play a key role in the activation and clustering of early origins of replication in budding yeast [44]. This regulation involves a cis-acting association of these two forkhead proteins with proteins at origins. It will be interesting to ask if the FHA domain of Fkh1 plays an important role in this regulation.
Another important finding emerging from our work is that two DNA damage checkpoint kinases, Mec1/ATR and Tel1/ATM, can act to phosphorylate distant DNA sequences when they are tethered in the vicinity of the DSB. As shown in Figure 6, the γ-H2AX modification spreads around the RE region, but with significantly delayed kinetics compared with the modification around MAT, consistent with the idea that RE has to first recognize and bind to phosphorylated residues in the vicinity of the DSB at MAT. How these checkpoint kinases act on their target sequences is not yet firmly established. Mammalian ATM has been shown to be activated by intermolecular autophosphorylation and dimer exchange, which would suggest that activated ATM would initially form a “cloud” of activated kinases around the site where the kinases were associated with the DSB ends [45], [46]. In the case of Tel1/ATM, the association with the DSB is via its association with the MRX/MRN proteins [47], [48]; in the case of Mec1/ATR, by its association its partner protein Ddc2/ATRIP with RPA bound to ssDNA at the resected DSB end [49], [50]. In budding yeast, the spreading of γ-H2AX from the DSB site is consistent with that the tethered kinases interact with phosphorylating histones on the adjacent chromosomal segment in a manner, which is similar to the contact of chromosomal regions as measured in chromosome conformation capture experiments [51]. Spreading of γ-H2AX further along the chromosome occurs more slowly and apparently depends on the continuing 5′ to 3′ resection of the DSB ends, generating ssDNA, as it depends only on Mec1 [27], [28]. Here we show that histones in another distant chromosomal region, brought into proximity with the DSB by RE, can also be efficiently phosphorylated – and by both Mec1 and Tel1. This result is different from the slow addition of γ-H2AX to regions further from the DSB, which depends on continuing 5′ to 3′ resection of the DSB ends and can only be performed by Mec1 [27], [28]. We have also observed γ-H2AX spreading onto a different chromosome during the ectopic recombinational repair of a DSB, when these two regions are brought together by Rad51-mediated strand invasion (K.L. and J.E.H., unpublished observations).
Materials and Methods
Yeast Strains
All strains except when noted were derived from strain XW652 (ho ade3::GAL::HO HMLα RE MATa HMRα-B ura3-52 lys5 leu2-3,112 trp1::hisG) carrying a galactose-inducible HO endonuclease integrated at the ADE3 locus [11]. Strains are pre-cultured in YP-lactate medium until cell density reaches about 5∼8×106 per ml. Galactose induction is performed for 1 hour and stopped by the addition of 2% dextrose.
Construction of ECY406 (Figure 2A): Four LexA operators are amplified from pSH18-34 [52] using primers BglIILexAU (5′-cga cga gat cta tac ata tcc ata tct aat ctt acc-3′) and BglIILexAL (5′-gct gca gat ctc taa tcg cat tat cat ccc tcg a-3′). Then PCR products were digested with BglII and subcloned into the BamHI site of pKS58 to generate pEC15. The SphI-digested pEC15 (marked with “LEU2”) was transformed into XW676 (ho ade3::GAL::HO HMLα REΔ::URA3 MATa HMRα-B ade1 leu2 trp1 ura3-52) to replace REΔ::URA3 with four LexA operators to generate a strain ECY405. Then, REΔ::LexABD4-LEU2 from ECY405 was replaced with REΔ::LexABD4-KAN to generate a strain ECY406 (Figure 2A) via transformation using PCR fragments amplified from pJH1894 with primers leu2KanU (5′-gag aac ttc tag tat atc cac ata cct aat att att gcc tta tta aaa atc agc tga agc ttc gta cgc-3′) and leu2KanL (5′-tac gtc gta agg ccg ttt ctg aca gag taa aat tct tga ggg aac ttt cag cat agg cca cta gtg gat ctg-3′).
ECY457 (Figure 2A) is constructed by transforming ECY406 with PCR fragment arg5,6::LexA-Fkh1 obtained with primers pAT4UII (5′-atg cca tct gct agc tta ctc gtc tcg aca aag aga ctt aac gct tcc aaa ttc cat ttt gta att tcg tgt cg-3′) and pAT4LII (5′-tca gac acc aat aat ttt att ttc agg gat acc agc ata ctc tcc ata aca agg gaa caa aag ctg gag c-3′) on the plasmid pEC16. Using a similar strategy, PCR products arg5,6::LexA-FHA (from pJL4), arg5,6::LexA-interdomain (from pJL5), arg5,6::LexA-forkhead (from pJL6) and arg5,6::LexA-FHA-R80A (from pJL8) are transformed into ECY406 to generate YJL019, YJL020, YJL021 and YJL094, respectively (Figure 3A).
YJL084 was made by transforming YJL019 (Figure 3B) with BamHI digested pJH1250 to delete HML using the URA3 marker. YJL110 (Figure 4A) is made by transforming YJL084 with BsaI-digested pJH2039 to delete HMR using the NAT marker.
Yeast strains with H3 or H4 N-terminal truncation were constructed by sequential transformations of JKM139 [32]. The HHF1-HHT1 allele of JKM139 was first knocked out by NAT-MX cassette to generate a strain YJK340 (ho ade3::GAL::HO hmlΔ::ADE1 RE MATa hmrΔ::ADE1 ade1 leu2-3-112 lys5 ura3-52 trp1::hisG hhf1-hht1Δ::NAT). Then, YJK340 was transformed with linearized plasmid carrying hht2-hhf2 mutant alleles linked to URA3 marker to replace endogenous HHT2-HHF2 allele. HHT2 was modified to lack the first 32 amino acids of histone H3 or HHF2 was modified to lack the first 16 amino acids of histone H4. To prepare for mating-type switching assay, HMLα and HMRα-B from XW652 were crossed into a yeast strain with H3 or H4 N-terminal truncation.
Measure Donor Preference via Southern Blot or a PCR-Based Assay
All strains except when noted in this study are derived from XW652 (ho ade3::GAL::HO HMLα RE MATa HMRα-B ura3-52 lys5 leu2-3,112 trp1::hisG) [11]. The C→A change at position 658 of Yα creates a BamHI restriction site (HMRα-B), which is absent in HMLα [8]. Donor preference (HML usage) is calculated using the formula (MATα/(MATα+MATα-B) for all XW652 derived strains (Figure 1A). The measurement of donor preference via Southern blot was described previously [8]. Southern signals were quantified using ImageQuant V1.2 (Molecular Dynamics).
Because there is only 1-bp difference between two repaired products (MATα and MATα-B), we have developed a PCR-based assay to measure donor preference. The presumption is that PCR amplification efficiency is almost identical for MATα and MATα-B because there is only 1-bp difference [8]. Around 10 ng of genomic DNA isolated from galactose-induced colonies will be used for PCR amplification. Two primers Yalpha105F (5′-gcc cac ttc taa gct gat ttc aat ctc tcc-3′) and MATdist-4R (5′-cct gtt ctt agc ttg tac cag agg-3′) can only amplify MATα or MATα-B, but not MATa, HMLα or HMRα-B due to sequence specificities of these two primers (Figure 1B). Although amplified PCR products are the mixture of MATα-B and MATα, only one 1470-bp band can be visualized on DNA agarose gel prior to digestion. PCR products are then purified and subsequently digested with BamHI. The digested PCR products will be checked on DNA agarose gel. MATα product will remain as the 1470-bp band, whereas MATα-B product is digested into two smaller bands with different sizes (550-bp and 920-bp) (Figure 1C). Donor preference is determined by comparing intensities of MATα and MATα-B after the agarose gel is stained with ethidium bromide.
Plasmid Constructs
To study if Fkh1 can regulate donor preference in our LexA system, we construct a LexA-Fkh1 fusion plasmid (pEC16) carrying the coding sequence of Fkh1. Fkh1 coding sequence is PCR amplified from XW652 genomic DNA using primers XmaIFkh1U (5′-tcg cga ccc ggg gat ccg tat gtc tgt tac cag tag gg-3′) and PstIFkh1L (5′-gca cga cct gca gtc aac tca gag agg aat tgt tca cg-3′). The amplified PCR product is digested with XmaI and PstI and then subcloned into a pre-digested pAT4 [21] to generate the plasmid pEC16.
To address different roles of Fkh1 domains in the regulation of donor preference, three regions of Fkh1 are subcloned into pAT4 (Figure 3A). The FHA domain of Fkh1 is amplified via PCR using primers XmaIFkh1U and PstIFkh1-690L (5′-gca cga cct gca gta ggt ggt cca gct gtt gta atc g-3′). The interdomain region is amplified using primers XmaIFkh1-487U (5′-tcg cga ccc ggg gat cgg tgt gca aat gat ctt tat at-3′) and PstIFkh1-906L (5′-gca cga cct gca gga tat atc tgt ttt cat cca gc-3′). The forkhead domain is amplified using primers XmaIFkh1-691U (5′-tcg cga ccc ggg gat cca cac ccc att atc gtc atc at-3′) and PstIFkh1L. These PCR products are then digested with XmaI and PstI, and subcloned into a pre-digested pAT4, to generate three fusion plasmids pJL4, pJL5 and pJL6, respectively.
Site-Directed Mutagenesis of pJL4
Quickchange Multi Site-Directed Mutagenesis Kit (Catalog # 200515, Stratagene, La Jolla, CA) was used to mutate the FHA domain of pJL4. Two primers Fkh1-Arg80 (5′-tta gaa gtt acc att ggt gcg aac aca gac agc ttg aac-3′) and pAT4-940R (5′-ctt tgc cag aca aga aca ccg cat-3′) were used to synthesize mutant strand from pJL4. Fkh1-Arg80 shares two-base mismatches with Fkh1 and pAT4-940R perfectly matches pJL4. The mutated plasmid pJL8 (pLexA-FHA-R80A) was confirmed by direct sequencing.
Chromatin Immunoprecipitation (ChIP)
Procedures for ChIP analysis were described previously [15]. Rabbit anti-LexA polyclonal antibody (Catalog no. 39184) used in ChIP assay is purchased from “Active Motif” company (Carlsbad, CA). LexA ChIP signals are quantified with real-time PCR using a Chromo 4 machine from MJ Research. The linearity of PCR signals is monitored with r-square value of a calibration curve, which is prepared using a series of dilutions of the 0 hr input sample. IP signal is determined by comparing to the calibration curve, and then normalized to the IP signal of a control locus CEN8. PCR primer sequences around the MAT, RE and the ectopic leu2::HOcs are available on request.
Zdroje
1. HaberJE 1998 Mating-type gene switching in Saccharomyces cerevisiae. Annu Rev Genet 32 561 599
2. HaberJE 2012 Mating-type genes and MAT switching in Saccharomyces cerevisiae. Yeast Book Genetics Society of America In press
3. KlarAJHicksJBStrathernJN 1982 Directionality of yeast mating-type interconversion. Cell 28 551 561
4. LooSRineJ 1994 Silencers and domains of generalized repression. Science 264 1768 1771
5. RavindraAWeissKSimpsonRT 1999 High-resolution structural analysis of chromatin at specific loci: Saccharomyces cerevisiae silent mating-type locus HMRa. Mol Cell Biol 19 7944 7950
6. WeissKSimpsonRT 1998 High-resolution structural analysis of chromatin at specific loci: Saccharomyces cerevisiae silent mating type locus HMLalpha. Mol Cell Biol 18 5392 5403
7. WeilerKSBroachJR 1992 Donor locus selection during Saccharomyces cerevisiae mating type interconversion responds to distant regulatory signals. Genetics 132 929 942
8. WuXHaberJE 1995 MATa donor preference in yeast mating-type switching: activation of a large chromosomal region for recombination. Genes Dev 9 1922 1932
9. WuXMooreJKHaberJE 1996 Mechanism of MAT alpha donor preference during mating-type switching of Saccharomyces cerevisiae. Mol Cell Biol 16 657 668
10. WuCWeissKYangCHarrisMATyeBK 1998 Mcm1 regulates donor preference controlled by the recombination enhancer in Saccharomyces mating-type switching. Genes Dev 12 1726 1737
11. WuXHaberJE 1996 A 700 bp cis-acting region controls mating-type dependent recombination along the entire left arm of yeast chromosome III. Cell 87 277 285
12. CoicERichardGFHaberJE 2006 Saccharomyces cerevisiae donor preference during mating-type switching is dependent on chromosome architecture and organization. Genetics 173 1197 1206
13. CoicEMartinJRyuTTaySYKondevJ 2011 Dynamics of homology searching during gene conversion in Saccharomyces cerevisiae revealed by donor competition. Genetics 189 1225 1233
14. WeissKSimpsonRT 1997 Cell type-specific chromatin organization of the region that governs directionality of yeast mating type switching. Embo J 16 4352 4360
15. SunKCoicEZhouZDurrensPHaberJE 2002 Saccharomyces forkhead protein Fkh1 regulates donor preference during mating-type switching through the recombination enhancer. Genes Dev 16 2085 2096
16. CoicESunKWuCHaberJE 2006 Cell cycle-dependent regulation of Saccharomyces cerevisiae donor preference during mating-type switching by SBF (Swi4/Swi6) and Fkh1. Mol Cell Biol 26 5470 5480
17. ErcanSReeseJCWorkmanJLSimpsonRT 2005 Yeast recombination enhancer is stimulated by transcription activation. Mol Cell Biol 25 7976 7987
18. SzetoLFafaliosMKZhongHVershonAKBroachJR 1997 Alpha2p controls donor preference during mating type interconversion in yeast by inactivating a recombinational enhancer of chromosome III. Genes Dev 11 1899 1911
19. BystrickyKVan AttikumHMontielMDDionVGehlenL 2009 Regulation of nuclear positioning and dynamics of the silent mating type loci by the yeast Ku70/Ku80 complex. Mol Cell Biol 29 835 848
20. WeilerKSSzetoLBroachJR 1995 Mutations affecting donor preference during mating type interconversion in Saccharomyces cerevisiae. Genetics 139 1495 1510
21. TaddeiAHedigerFNeumannFRBauerCGasserSM 2004 Separation of silencing from perinuclear anchoring functions in yeast Ku80, Sir4 and Esc1 proteins. Embo J 23 1301 1312
22. DurocherDHenckelJFershtARJacksonSP 1999 The FHA domain is a modular phosphopeptide recognition motif. Mol Cell 4 387 394
23. DurocherDJacksonSP 2002 The FHA domain. FEBS Lett 513 58 66
24. LiJSmithGPWalkerJC 1999 Kinase interaction domain of kinase-associated protein phosphatase, a phosphoprotein-binding domain. Proc Natl Acad Sci U S A 96 7821 7826
25. LiaoHByeonIJTsaiMD 1999 Structure and function of a new phosphopeptide-binding domain containing the FHA2 of Rad53. J Mol Biol 294 1041 1049
26. LiJLeeGIVan DorenSRWalkerJC 2000 The FHA domain mediates phosphoprotein interactions. J Cell Sci 113 4143 4149
27. KimJAKruhlakMDotiwalaFNussenzweigAHaberJE 2007 Heterochromatin is refractory to gamma-H2AX modification in yeast and mammals. J Cell Biol 178 209 218
28. ShroffRArbel-EdenAPilchDIraGBonnerWM 2004 Distribution and dynamics of chromatin modification induced by a defined DNA double-strand break. Curr Biol 14 1703 1711
29. CheungWLTurnerFBKrishnamoorthyTWolnerBAhnSH 2005 Phosphorylation of histone H4 serine 1 during DNA damage requires casein kinase II in S. cerevisiae. Curr Biol 15 656 660
30. JainSSugawaraNLydeardJVazeMTanguy Le GacN 2009 A recombination execution checkpoint regulates the choice of homologous recombination pathway during DNA double-strand break repair. Genes Dev 23 291 303
31. LichtenMBortsRHHaberJE 1987 Meiotic gene conversion and crossing over between dispersed homologous sequences occurs frequently in Saccharomyces cerevisiae. Genetics 115 233 246
32. SugawaraNWangXHaberJE 2003 In vivo roles of Rad52, Rad54, and Rad55 proteins in Rad51-mediated recombination. Mol Cell 12 209 219
33. WolnerBvan KomenSSungPPetersonCL 2003 Recruitment of the recombinational repair machinery to a DNA double-strand break in yeast. Mol Cell 12 221 232
34. MooreJDYazganOAtaianYKrebsJE 2007 Diverse roles for histone H2A modifications in DNA damage response pathways in yeast. Genetics 176 15 25
35. HendzelMJWeiYManciniMAVan HooserARanalliT 1997 Mitosis-specific phosphorylation of histone H3 initiates primarily within pericentromeric heterochromatin during G2 and spreads in an ordered fashion coincident with mitotic chromosome condensation. Chromosoma 106 348 360
36. GotoHTomonoYAjiroKKosakoHFujitaM 1999 Identification of a novel phosphorylation site on histone H3 coupled with mitotic chromosome condensation. J Biol Chem 274 25543 25549
37. DaiJHigginsJM 2005 Haspin: a mitotic histone kinase required for metaphase chromosome alignment. Cell cycle 4 665 668
38. KaynePSKimUJHanMMullenJRYoshizakiF 1988 Extremely conserved histone H4 N terminus is dispensable for growth but essential for repressing the silent mating loci in yeast. Cell 55 27 39
39. TohGWSugawaraNDongJTothRLeeSE 2010 Mec1/Tel1-dependent phosphorylation of Slx4 stimulates Rad1-Rad10-dependent cleavage of non-homologous DNA tails. DNA repair 9 718 726
40. HannaDERethinaswamyAGloverCV 1995 Casein kinase II is required for cell cycle progression during G1 and G2/M in Saccharomyces cerevisiae. J Biol Chem 270 25905 25914
41. BressanDAVazquezJHaberJE 2004 Mating type-dependent constraints on the mobility of the left arm of yeast chromosome III. J Cell Biol 164 361 371
42. DekkerJRippeKDekkerMKlecknerN 2002 Capturing chromosome conformation. Science 295 1306 1311
43. MieleABystrickyKDekkerJ 2009 Yeast silent mating type loci form heterochromatic clusters through silencer protein-dependent long-range interactions. PLoS Genet 5 e1000478 doi:10.1371/journal.pgen.1000478
44. KnottSRPeaceJMOstrowAZGanYRexAE 2012 Forkhead transcription factors establish origin timing and long-range clustering in S. cerevisiae. Cell 148 99 111
45. BakkenistCJKastanMB 2003 DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature 421 499 506
46. BonillaCYMeloJAToczyskiDP 2008 Colocalization of sensors is sufficient to activate the DNA damage checkpoint in the absence of damage. Mol Cell 30 267 276
47. LeeJHPaullTT 2005 ATM activation by DNA double-strand breaks through the Mre11-Rad50-Nbs1 complex. Science 308 551 554
48. NakadaDMatsumotoKSugimotoK 2003 ATM-related Tel1 associates with double-strand breaks through an Xrs2-dependent mechanism. Genes Dev 17 1957 1962
49. CimprichKACortezD 2008 ATR: an essential regulator of genome integrity. Nat Rev Mol Cell Biol 9 616 627
50. ZouLElledgeSJ 2003 Sensing DNA damage through ATRIP recognition of RPA-ssDNA complexes. Science 300 1542 1548
51. DekkerJ 2008 Mapping in vivo chromatin interactions in yeast suggests an extended chromatin fiber with regional variation in compaction. J Biol Chem 283 34532 34540
52. EstojakJBrentRGolemisEA 1995 Correlation of two-hybrid affinity data with in vitro measurements. Mol Cell Biol 15 5820 5829
53. LeeSEMooreJKHolmesAUmezuKKolodnerRD 1998 Saccharomyces Ku70, mre11/rad50 and RPA proteins regulate adaptation to G2/M arrest after DNA damage. Cell 94 399 409
54. PaquesFLeungWYHaberJE 1998 Expansions and contractions in a tandem repeat induced by double-strand break repair. Mol Cell Biol 18 2045 2054
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2012 Číslo 4
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- A Coordinated Interdependent Protein Circuitry Stabilizes the Kinetochore Ensemble to Protect CENP-A in the Human Pathogenic Yeast
- Coordinate Regulation of Lipid Metabolism by Novel Nuclear Receptor Partnerships
- Defective Membrane Remodeling in Neuromuscular Diseases: Insights from Animal Models
- Formation of Rigid, Non-Flight Forewings (Elytra) of a Beetle Requires Two Major Cuticular Proteins
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy