
Differing Requirements for RAD51 and DMC1 in Meiotic Pairing of Centromeres and Chromosome Arms in
During meiosis homologous chromosomes pair, recombine, and synapse, thus ensuring accurate chromosome segregation and the halving of ploidy necessary for gametogenesis. The processes permitting a chromosome to pair only with its homologue are not fully understood, but successful pairing of homologous chromosomes is tightly linked to recombination. In Arabidopsis thaliana, meiotic prophase of rad51, xrcc3, and rad51C mutants appears normal up to the zygotene/pachytene stage, after which the genome fragments, leading to sterility. To better understand the relationship between recombination and chromosome pairing, we have analysed meiotic chromosome pairing in these and in dmc1 mutant lines. Our data show a differing requirement for these proteins in pairing of centromeric regions and chromosome arms. No homologous pairing of mid-arm or distal regions was observed in rad51, xrcc3, and rad51C mutants. However, homologous centromeres do pair in these mutants and we show that this does depend upon recombination, principally on DMC1. This centromere pairing extends well beyond the heterochromatic centromere region and, surprisingly, does not require XRCC3 and RAD51C. In addition to clarifying and bringing the roles of centromeres in meiotic synapsis to the fore, this analysis thus separates the roles in meiotic synapsis of DMC1 and RAD51 and the meiotic RAD51 paralogs, XRCC3 and RAD51C, with respect to different chromosome domains.
Published in the journal:
. PLoS Genet 8(4): e32767. doi:10.1371/journal.pgen.1002636
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002636
Summary
During meiosis homologous chromosomes pair, recombine, and synapse, thus ensuring accurate chromosome segregation and the halving of ploidy necessary for gametogenesis. The processes permitting a chromosome to pair only with its homologue are not fully understood, but successful pairing of homologous chromosomes is tightly linked to recombination. In Arabidopsis thaliana, meiotic prophase of rad51, xrcc3, and rad51C mutants appears normal up to the zygotene/pachytene stage, after which the genome fragments, leading to sterility. To better understand the relationship between recombination and chromosome pairing, we have analysed meiotic chromosome pairing in these and in dmc1 mutant lines. Our data show a differing requirement for these proteins in pairing of centromeric regions and chromosome arms. No homologous pairing of mid-arm or distal regions was observed in rad51, xrcc3, and rad51C mutants. However, homologous centromeres do pair in these mutants and we show that this does depend upon recombination, principally on DMC1. This centromere pairing extends well beyond the heterochromatic centromere region and, surprisingly, does not require XRCC3 and RAD51C. In addition to clarifying and bringing the roles of centromeres in meiotic synapsis to the fore, this analysis thus separates the roles in meiotic synapsis of DMC1 and RAD51 and the meiotic RAD51 paralogs, XRCC3 and RAD51C, with respect to different chromosome domains.
Introduction
Sexual reproduction involves the fusion of maternal and paternal gametes and this means that the parental genetic complement must be halved in the process of gametogenesis, to avoid it doubling at each generation. This halving of ploidy is carried out by meiosis, the specialised eukaryotic cell division that involves one round of DNA replication followed by two sequential divisions. Errors in segregation of the genetic material during cell division lead to aneuploidy, a well known example of which in humans is Down's Syndrome, caused by trisomy of chromosome 21 [1]. It is thus essential that each daughter cell inherits a full complement of the genetic material and in mitosis this is ensured by centromeric cohesion established at the preceeding S-phase. This mechanism also ensures proper chromosomal segregation during the second meiotic division, however recognition and linking of homologous chromosomes at the first meiotic division is mediated in the majority of eukaryotes by recombination during the first meiotic prophase.
While understanding of the mechanisms of homologous recombination has considerably advanced, the processes permitting chromosomes to recognise and pair with only their homologues remain elusive. Homologue pairing has been shown to depend on a number of mechanisms including homologous recombination, centromere coupling, telomere clustering and the interaction of specific pairing centres with Zinc finger DNA-binding proteins seen in Drosophila melanogaster and Caenorhabditis elegans [2]–[6]. In animals and fungi, clustering of telomeres at the leptotene/zygotene transition into a “bouquet” associated with the microtubule organising center promotes homologue alignment (reviewed by [7]). Evidence from budding yeast shows that Zip1 - and Rec8-dependent centromere coupling, or non-homologous centromere pairing, precedes homologous interactions, which are then stabilised by Spo11-dependent homologous recombination mechanisms [8]–[11]. A recent report has shown that PP4 phosphatase is required to counteract Mec1-dependent phosphorylation of Zip1 and permit this non-homologous centromere coupling [12]. Transition to homologous chromosome interactions leads to co-alignment of homologous chromosome axes in zygotene, and the fully synapsed chomosomes visible at pachytene. In wheat, centromeres cluster premeiotically and further associate in pairs facilitating pairing and recombination in a mechanism dependent on the Ph1 locus [13], [14] and centromere coupling is observed in the Arabidopsis thaliana phs1 mutant [15]. The role of centromeres in meiotic pairing is an active subject of research in many organisms (reviewed by [16]). In this context it is important to note that the structure of centromeric regions differs considerably between species, ranging from 125 bp in Saccharomyces cerevisiae to the highly repeated DNA of up to several megabases in length found in multicellular eukaryotes. In Arabidopsis, centromeric DNA contains long stretches of tandemly repeated DNA sequences, transposons, retrotransposons and rDNA, and this has meant that full DNA sequence analysis of Arabidopsis centromeres has not been completed [17]–[20]; see review by [21].
Meiotic recombination is initiated by the induction of double-strand DNA breaks in the chromosomes by Spo11 during the leptotene stage. Resection at the DSB generates single-stranded DNA overhangs and these load Rad51 and/or Dmc1 protein into a helical nucleofilament, which catalyses invasion of, and synapsis with, an homologous DNA sequence. Repair of DSB in G2 and M-phase mitotic cells, and the majority of DSB in meiotic cells, involves primarily the invasion of the sister chromatid. However during meiosis, a subset of breaks are repaired through recombination with the homologous chromosome, thus establishing the physical linkage necessary to ensure proper chromosomal disjunction at the first meiotic anaphase. How meiotic cells permit synapsis with sister chromosome, rather than the sister chromatid, is a major question in meiosis and much research is currently focussed on the specificities of the Rad51 recombinase and its meiosis-specific paralogue, Dmc1. Dmc1 has been shown to play similar, but not identical roles to Rad51 [22], however, while Rad51 is needed for both meiotic and mitotic recombination, Dmc1 is only required in meiosis [5], [23]. In yeast, current understanding points to action of the Red1/Hop1/Mek1 complex promoting meiotic inter-homolog recombination through phosphorylation of the axial element protein Hop1 [24]–[26]. Also, Hed1 restricts activity of Rad51 nucleofilaments in meiosis by blocking access of Rad54 [27], [28]. These mechanisms attenuate the activity of Rad51 to minimise the use of the sister chromatid and hence favour Dmc1-dependent inter-homolog recombination. Dmc1 plays a key role in inter-homolog recombination in plants, yet the mechanisms through which RAD51 and DMC1 cooperate to promote the homology search and chromosome synapsis are unknown [29]–[31]. Arabidopsis mutants lacking RAD51 or DMC1 are sterile but show normal somatic growth [29], [32]. Both mutants show defects in pairing and synapsis, however while rad51 mutants exhibit extensive chromosome fragmentation at late pachytene, dmc1 mutants are characterised by the presence of intact univalents, showing that DSBs are repaired in a RAD51-dependent manner using the sister chromatid as a template in the absence of DMC1 [33]. The precise roles and mechanisms of action of Dmc1 nonetheless remain poorly understood and some organisms, such as Drosophila or Caenorhabditis elegans, do not possess an apparent Dmc1 ortholog.
The RAD51 paralogue proteins RAD51C and XRCC3 play key roles in the homology search together with RAD51, and have been shown to act as cofactors facilitating loading of RAD51 onto DNA [34]–[39]. The lethality of these mutants in vertebrates has hampered the study in meiosis and the details of their roles in meiotic homologous pairing remain elusive [40]–[42]. In Arabidopsis, xrcc3 and rad51C mutants are viable but extensive chromosome fragmentation in late pachytene results in sterility, a similar phenotype to that of rad51 mutants [43]–[47]. Both the RAD51C and XRCC3 proteins are essential for meiotic recombination and they have both early and late functions during meiotic prophase I [40]–[49]. The specific role(s) of XRCC3 and RAD51C in homologous recombination and how they cooperate with the RAD51 and DMC1 recombinases to achieve efficient homologous chromosome pairing remain subject to debate.
Working with the flowering plant Arabidopsis thaliana, we present here an analysis of the roles of recombination and key recombination proteins in this process, showing the existence of a DMC1-dependent process which stabilises pairing of centromeric regions of homologous chromosomes and that, in contrast, synapsis of chromosome arms requires RAD51 protein and the RAD51 paralogues, RAD51C and XRCC3.
Results
Arabidopsis xrcc3 and rad51C Recombination-Defective Mutants Exhibit Partial Chromosome Alignment and Pairing
Meiosis in Arabidopsis xrcc3 and rad51C mutants progresses up to the zygotene stage and appears to enter pachytene, but do not show full chromosomal synapsis at pachytene and we thus call this meiotic stage zygo-pachytene. Subsequent meiotic stages present fragmented and fused chromosomes and no viable gametes are produced [43]–47,50. The presence of pachytene-like figures and the observation of fragmented bivalents at metaphase I show that some chromosome pairing does occur in these mutants and we have set out to determine the nature of this and its dependence on recombination.
Observation of meiotic figures clearly confirms the presence of partial synapsis at the zygo-pachytene stage in xrcc3 mutants. Chromosome alignment and pairing is visible as short stretches and thick chromosome threads typical of synapsed chromosomes are clearly visible (Figure 1). The thick, synapsed fibres frequently end in bubble structures in which the two chromosome axes are clearly visible (Figure 1B–1F). Immunolocalisation of the synaptonemal complex (SC) central element protein, ZYP1, shows foci and short stretches in the xrcc3 mutant, in agreement with the observed partial synapsis of DAPI-stained chromosomes (Figure S1). Similar ZYP1 staining patterns have been described in Arabidopsis rad51, xrcc3, and rad51C mutants [50]. Transmission electron microscopy studies in rad51 and rad51C mutants also show short sections of synaptonemal complex and thus evidence of partial chromosome synapsis [32], [47].

Pairing of Chromosome Arms Is Dependent on Xrcc3 and Rad51C
These observations prompted us to further characterise the partial synapsis observed in the rad51 paralogue mutants, using fluorescence in situ hybridisation (BAC-FISH) to examine pairing in xrcc3 mutants using a BAC probe (F12C20) recognising a mid-arm region of the long arm of chromosome 2 (Figure 2). As expected, in wild-type meiocytes, two separate signals were observed from leptotene to early zygotene and one signal (or two paired signals) were observed in pachytene figures (Figure 2 and Table 1). In contrast, in xrcc3 mutants two signals were observed in 80 percent of the meiocytes at the zygo-pachytene stage (Figure 2 and Table 1). Similar results were obtained using a second BAC probe (F12K11) probing the distal region of the left-arm (2 Mbp from the telomere) of chromosome 1, with more than 90% of xrcc3 meiocytes showing unpaired chromosomes while 85% of the WT meiocytes were fully paired (Figure 2 and Table 1). BAC-FISH analyses in rad51C meiocytes using F12K11 probe confirmed the absence of pairing observed in the xrcc3 mutant (Table 1). Pairing and synapsis in these regions is thus dependent on RAD51 paralogue-dependent recombination.


Centromeres Pair in Arabidopsis xrcc3 and rad51C Mutants
Centromere pairing is an early event in meiotic chromosome pairing and has been well described in Arabidopsis [51]. Arabidopsis centromeres are unpaired and dispersed during meiotic interphase up to leptotene, cluster at leptotene/zygotene, separate and homologous centromeres then associate in pairs and synapse in zygotene and pachytene. FISH analyses using a 180 bp centromeric repeat-specific probe were used to follow pairing of centromeres through meiotic prophase I in xrcc3 and rad51C mutants (Figure 3). For each stage, meiocytes were placed into one of four classes according to the number of fluorescent signals from the centromeric repeat-specific probe : in type 1, 8–10 centromere signals indicate unpaired centromeres; in type 2 meiocytes, 6–7 signals show partial centromere pairing; type 3 meiocytes have clustered centromeres (1 to 3 signals) and type 4 meiocytes show four or five signals and complete centromere pairing (Figure 3).

In wild - type, virtually all leptotene nuclei were of type 1 (unpaired centromeres; Figure 3A and 3D). Zygotene nuclei were mostly of type 3 or 4, while in pachytene 69% of nuclei were of type 4 and the remainder of type 3, indicative of complete centromere pairing (Figure 3A and 3D). Similarly, in xrcc3 and rad51C mutants, more than 95 percent of the observed leptotene nuclei showed ten centromere signals (type 1 nuclei). The proportion of Type 1 nuclei then declined to reach the five expected signals (type 4) in zygo-pachytene, xrcc3 and rad51C show centromere pairing with 51% and 50% type 4 nuclei, respectively (Figure 3B, 3C, 3E and 3F). These results are in accordance with the centromere coupling reported in the Arabidopsis rad51C mutant [47]. As seen in the frequency distributions, dynamics of centromere pairing in xrcc3 and rad51C is thus similar to WT with, 51%, 50% and 69%, respectively, of the zygo-pachytene nuclei showing 4 or 5 centromeric signals (Figure 3D to 3F). We note, however, that the distribution is a little more spread out in the mutants than in the wild-type (Figure 3D to 3F) and that the proportion of meiocytes of both type 2 and type 3 in xrcc3 and rad51C (47 and 45%, respectively) is higher than in the wild-type (31%). Centromeres thus pair efficiently in xrcc3 and rad51C mutants, in contrast to chromosome arms. Given that the 180 bp DNA repeat region detected by the centromeric FISH probe is found at all Arabidopsis centromeres, the question remains as to whether or not the observed centromeric pairing involves centromeres of homologous chromosomes?
Pairing of Homologous Centromeres and Pericentromeric Chromosomal Regions in xrcc3 and rad51C Mutants
5S rDNA loci are present near to the centromeres of chromosomes 3, 4 and 5 of the Columbia ecotype of Arabidopsis [52]. As a result, fully synapsed homologous chromosomes in pachytene nuclei show 3 distinct 5S rDNA FISH foci (Figure 4). To quantify the extent of homologous or non-homologous centromere coupling, we examined pachytene meiocytes using FISH with both the 180 bp repeat and 5S rDNA probes. Three 5S rDNA foci thus indicate homologous centromere pairing and more than three, non-homologous centromere coupling. Zygo-pachytene figures in the mutants showing more than 5 or less than 3 centromeric signals were excluded from the analysis as centromere pairing was not yet complete in these meiocytes. Meiocytes exhibiting 2 or 3 5S rDNA signals were considered as homologously paired while meiocytes with more than 3 5S rDNA signals and 4/5 centromere signals were considered as non-homologously coupled.

As shown in Figure 4, 80% of xrcc3 (n = 20) and 72% of rad51C (n = 68) meiocytes showed homologous pairing. Although reduced compared to the wild-type (100% paired), homologous centromere pairing does clearly not fully depend on Xrcc3 - and Rad51C-dependent recombination in Arabidopsis.
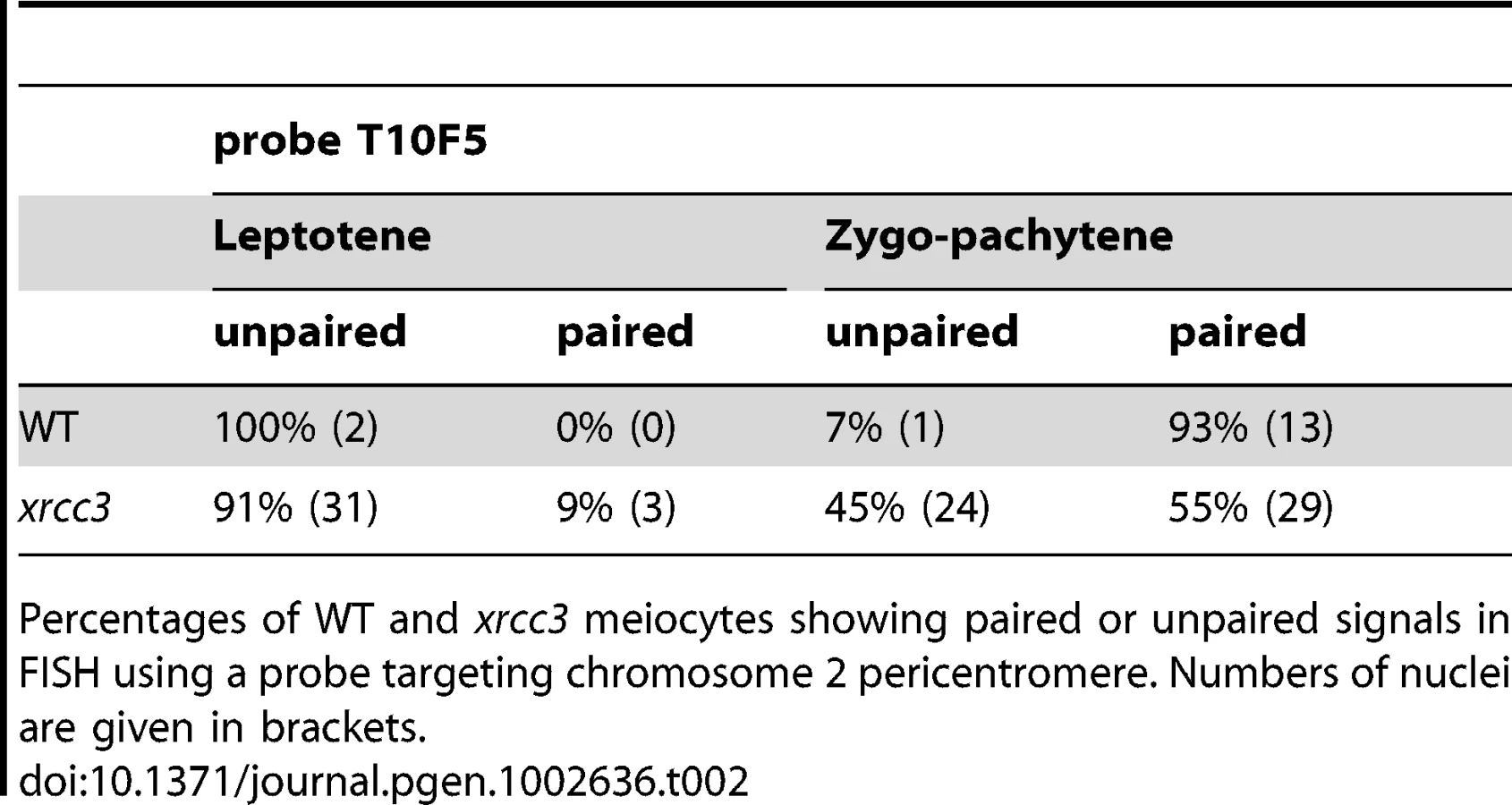
In order to confirm our observations and to exclude the possibility that homologous pairing is restricted only within these repeated regions, we monitored homologous pairing in xrcc3 mutants using BAC-FISH with a unique pericentromeric probe (T10F5) located on the long arm of chromosome 2. Two foci were observed in leptotene and early zygotene in both wild-type and xrcc3 mutants. Beginning at late zygotene and extending through pachytene, pairing proceeds and a single focus is seen in more than 90% of the wild-type meiocytes (Figure 5; Table 2). Homologous pairing was confirmed in xrcc3 mutants, with 55% of zygo-pachytene stage meiocytes showing a single focus (29 of 53 nuclei; Figure 5 and Table 2). We note that these results underestimate the homologous pairing since, in contrast to 5S rDNA analyses, chromosome 2 foci were monitored in all zygo-pachytene nuclei irrespective of the number of centromere foci. These results thus corroborate the 5S-rDNA FISH (above) showing that rad51 paralogue mutants, xrcc3 and rad51C, show homologous centromere pairing. Strikingly, as seen in Figure 5B, this pairing can extend well into the euchromatic regions flanking the centromeric heterochromatin (the BAC probe used in this analysis lies 2 Mbp from the centromere repeat region).

DMC1 Is the Key Recombinase for Efficient Homologous Centromere Pairing
This study was originally undertaken following our observation of meiotic phenotypes of the Arabidopsis xrcc3 and rad51C mutants [44]–[46]. These results have been confirmed and extended by other authors, and strikingly, a similar phenotype is also observed in the rad51 mutant [32], [50], [53]. This raises the possibility that the homologous centromere pairing we observe in Arabidopsis xrcc3 and rad51C mutant plants is also independent of the Rad51 protein. In order to verify this we carried out experiments to determine which recombination pathway is involved in the homologous centromere pairing.
Following the approach used for the Rad51 paralogue mutants, we monitored numbers of centromeric and 5S rDNA foci through meiotic prophase I in rad51, dmc1 and spo11-1 mutants (Figure 6). Complete centromere pairing with 4 or 5 centromeric signals was observed in 51% of the rad51 zygo-pachytene nuclei, a value similar to that observed for the xrcc3 and rad51C mutants (Figure 3). The dmc1, double rad51 dmc1 and spo11-1 mutants also show similar levels of centromere pairing, with 39%, 36% and 46%, respectively (Figure 6A and 6B and Table 3). As noted above for the RAD51 paralogue mutants, the frequency distributions of centromeric foci in the mutants are very similar, although slightly more spread out, than in the wild-type (compare Figure S2 with Figure 3D–3F).


These data clearly suggest the existence of a centromere coupling mechanism that is independent of recombination. To test whether or not this concerns centromeres of homologous chromosomes, we monitored the number of 5S rDNA signals in zygo-pachytene mutant meiocytes. These data show the existence of a SPO11-dependent process being responsable for homologous centromere pairing, with rad51, xrcc3 and rad51C mutants not being significantly different to the WT (Table 4).

That the rad51, xrcc3 and rad51C mutants do not show the SPO11-dependent effect implies that this homologous centromere pairing principally involves DMC1. We thus dissected the relative roles of DMC1 and RAD51 by monitoring numbers of 5S rDNA foci in nuclei showing 4 to 5 centromeric signals (full centromere coupling, see above). The rad51 mutant gives values similar to those reported above for the xrcc3 and rad51C mutants, with 68% (n = 101) of zygo-pachytene figures showing homologous centromere pairing (Figure 6C). In contrast, non-homologous centromere coupling was predominant in dmc1, rad51 dmc1 and spo11-1 mutants, with homologous centromere pairing observed in 43% of dmc1 (n = 14), 41% of rad51 dmc1 (n = 17) and 36% of spo11-1 (n = 36) mutants (Figure 6C). To confirm these data, BAC-FISH analyses were performed in the rad51, dmc1, and rad51 dmc1 mutants. Pericentromeric pairing was analysed using BAC-FISH with two adjacent pericentromeric probes, 1.4 to 1.85 Mbp from the centromere of chromosome 1 (Figure 7). As expected, the majority of rad51 meiocytes with full centromere pairing showed a single BAC-FISH focus and thus homologous centromere pairing (64%, n = 14; Figure 7). In contrast, only 38% of the dmc1 meiocytes analysed (n = 16) displayed homologous pairing and this dropped to 10% in the rad51 dmc1 double mutant (n = 39; Figure 7).

Meiotic pairing of homologous centromeres in Arabidopsis thus depends upon Spo11 induced recombination and primarily requires DMC1, but is largely independent of Rad51 and the Rad51 paralogues.
Discussion
Proper pairing of homologous chromosomes is necessary for proper disjunction at the first meiotic division and thus for the production of functional gametes. This pairing is mediated in the majority of eukaryotes by recombination during the first meiotic prophase and, although the subject of considerable interest, the molecular mechanisms ensuring that chromosomes pair only with their homologues are not fully understood. In the work presented here we have analysed meiotic chromosome pairing in recombination mutants of Arabidopsis and show a differing requirement for these proteins in pairing of pericentromeric regions and chromosomal arms regions during meiotic prophase. We confirm the existence of a non-homologous centromere coupling mechanism in Arabidopsis, independent of the formation of double-strand breaks by SPO11, and RAD51 - and DMC1-dependent recombination. Establishment and stabilisation of pairing of homologous centromeric and pericentromeric regions depends principally upon DMC1, while pairing and synapsis of euchromatic chromosome arms of homologues requires the presence of RAD51 and the RAD51 paralogues, XRCC3 and RAD51C.
Centromere Coupling in Arabidopsis Is Independent of Recombination
Homologous pairing and synapsis is achieved through recognition and proper alignment of homologous chromosomes. With some notable exceptions [54], [55] recombination is clearly required for this, but prior to recombination, different homology-independent interactions have been shown to position and pre-align homologues, thus perhaps preparing the ground for recombination and full homologous synapsis.
The role of telomeres in homologous chromosome positioning and alignment is well established in a number of organisms (reviewed by [7], [56]), although many questions remain concerning the mechanisms involved and the extent to which telomeres participate in pairing is unclear. Of particular relevance here, although Arabidopsis telomeres associate with the nucleolus in meiotic prophase, no telomere “bouquet” is observed [51].
Pairing between non-homologous centromeres, or centromere coupling [11], in early meiotic prophase has been reported in many organisms (reviewed by [16]). In budding yeast, centromere pairing is tightly linked to recombination and initiation of synapsis [9]–[12], [16] and can be sufficient in itself to mediate proper segregation [57]. Non-homologous centromere coupling does not depend upon Spo11-induced recombination, but is dependent on the Zip1 protein. Spo11 and hence initiation of recombination, is however needed for the transition to homologous centromere pairing [10], [11]. Recent results from Drosophila show that centromere clustering initiates SC formation and synapsis of homologous chromosomes in Drosophila female meiosis [58], [59].
In contrast to the pre-meiotic centromere pairing described in onions [60] and wheat [14], Arabidopsis centromeres remain separate until leptotene, when they transiently cluster, separate and initiate pairing during zygotene. Arabidopsis centromere pairing completes during pachytene and is released in diplotene/diakinesis [51]. As summarised in Table 5, meiotic non-homologous centromere coupling has been observed in a number of Arabidopsis mutants affecting meiotic chromosome cohesion, synapsis and recombination, such as the phs1 mutant [15] (full references are given in Table 5). Only SYN1/REC8, a member of the meiotic cohesin complex, has been shown to be required for centromere coupling in plants, with 10 centromeres being visible throughout prophase I in Arabidopsis syn1 meiocytes [61]. Similar but less conclusive results are also seen in the mutant of the Arabidopsis SKP1 homologue, ASK1, implicated in a number of early meiotic nuclear reorganisation events [62], [63].

In this work we show that meiotic prophase I coupling of centromeres of non-homologous chromosomes does occur in Arabidopsis spo11-1, rad51, dmc1 and rad51 dmc1 mutants. Notwithstanding the presence of long repetitive DNA homologies at the different Arabidopsis centromeres (see introduction), this process thus does not depend upon the induction of DNA breaks by SPO11, nor the RAD51/DMC1 recombination machinery in Arabidopsis.
RAD51 and XRCC3 Act Together for Homologous Pairing in Chromosome Arms
Meiosis in Arabidopsis rad51, xrcc3 and rad51C mutants appears normal up to the zygotene/pachytene stage, however no fully synapsed pachytene figures are observed and later stages present dramatic chromosomal fragmentation and fusion (This work and [32], [43]–[47], [50]. Some pairing however occurs in these mutants with clustering of centromeres and pairing of telomeres reported in rad51C and homologous pairing of 5S rDNA loci in xrcc3 mutants (this work) [46], [47]. Similarly, telomere pairing has also been reported in asy1 and atm mutants [51], [64]. A recent study of a hypomorph rad51 Arabidopsis knockdown-mutant provided indications of homologous pairing at metaphase I, with half of the bivalents involving homologous chromosomes and this homologous pairing was abolished in a rad51 dmc1 double mutant [30]. The severe chromosomal fragmentation and fusion observed post zygotene/pachytene means however that some care must be taken with this conclusion. We present here analyses of the roles of recombination in homologous pairing of specific chromosome regions at the zygo-pachytene stage, prior to chromosome fragmentation/fusion. This approach shows that homologous centromeres pair in Arabidopsis rad51, rad51C and xrcc3 mutants. Strikingly, this is not the case for the tested euchromatic chromosome arms. Our results furthermore show that this pairing can extend into the euchromatic pericentromeric regions for at least 2 Mb from the centromere (Figure 5B) The Arabidopsis genome is sequenced and chromosome 2 is estimated at 19,698,400 bp (TAIR10; http://www.arabidopsis.org), which means that the chromosome 2 pairing seen in this figure represents at least 10% of the whole chromosome. To further put this in context, the KEGG database (http://www.genome.jp/kegg-bin/show_organism?org=sce) lists the length of chromosome IV of S. cerevisiae (the largest yeast chromosome) as 1,531,933 bp, considerably less than the 2 Mbp centromere-flanking synapsed region observed here in the Arabidopsis xrcc3 mutant. RAD51, XRCC3 and RAD51C thus act together to promote pairing and synapsis along chromosome arms.
DMC1 Is the Key Recombinase for Efficient Pairing of Homologous Centromeres during Meiosis
In most eukaryotes, including plants, meiotic homologous pairing and synapsis are intimately linked to homologous recombination between homologous chromosomes (reviews by [4], [23], [65]). Both the RAD51 and the DMC1 recombinases are necessary for the normal progression of meiotic recombination, with however differing roles, and these differences are believed to be key to the specificities of meiotic and mitotic recombination. RAD51 is involved in both mitotic and meiotic homologous recombination between sister chromatids, while DMC1 acts to promote recombination with the sister chromosome during meiosis, but has no identified role in mitosis. These differing meiotic roles of RAD51 and DMC1 are seen in the phenotypes of Arabidopsis rad51 and dmc1 mutants : although no bivalents are detected in the dmc1 mutant the genome remains intact, while absence of RAD51 leads to genome fragmentation and sterility [29], [32]. Absence of the RAD51 paralogues XRCC3 and RAD51C results in a similar phenotype to rad51, with fragmentation of the genome following the zygotene/pachytene stage [43], [44], [46], [47]. The earlier loading of DMC1 compared to RAD51 [31] and the presence of only (intact) univalents in dmc1 mutants, implies that DSBs are repaired using the sister chromatid as a template and thus that DMC1 functions to favour interhomolog repair [29], [30], [33], [50]. Similarly, recent work on meiosis in a dmc1 mutant of Tetrahymena has shown the suppression of crossing-over associated with efficient RAD51-dependent repair of DSBs [66]. Studies in yeast led to an asymmetrical strand invasion model in which Dmc1 and Rad51 are loaded on opposite sides of the break [67], [68]. The favoured homology search model is that Dmc1 first invades the homologous partner and that Rad51 acts to stabilise and extend the strand invasion intermediates (see review by [23]).
As described above, we have confirmed that meiotic coupling of non-homologous centromeres in Arabidopsis occurs independently of recombination. This is not however the case with the transition to homologous centromere pairing, which we show to be dependent on the initiation of recombination by SPO11. Surprisingly, pairing of homologous centromeres seen in nearly 70% of meiocytes of rad51, xrcc3 and rad51C mutants, but synapsis does not however extend to chromosome arms. Homologous centromere pairing thus is largely independent of RAD51, XRCC3 and RAD51C. In striking contrast, homologous centromere pairing is significantly reduced in the dmc1 mutant and this effect is even more pronounced in the rad51 dmc1 double mutant (and spo11-1), confirming the primary role of DMC1 and showing a minor role for RAD51. Such a role for RAD51 in supporting DMC1 in Arabidopsis is in accordance with the recent study of the hypomorph rad51-2 allele [30]. Meiotic pairing of homologous centromeres in Arabidopsis is thus dependent on the initiation of recombination by SPO11 and principally promoted by DMC1. Extension and completion of synapsis to chromosome arms is however dependent upon RAD51 (and XRCC3 and RAD51C). Based on these data, we suggest a model of meiotic chromosome pairing in Arabidopsis in which, following recombination-independent non-homologous centromere coupling, homologous chromosome pairing is initiated at centromeres via a DMC1-dependent interhomolog search mechanism and that pairing is further stabilised and extended along chromosome arms through RAD51-dependent homologous recombination (Figure 8).

A Role for Heterochromatin in Meiotic Chromosome Pairing?
The observations of differing requirements for RAD51 - and DMC1-dependent recombination at heterochromatic centromeres and euchromatic chromosome regions raises the question of whether these differences are centromere-specific or whether they reflect differing behaviour of heterochromatin and euchromatic regions in chromosomal synapsis.
The role for heterochromatin in chromosome pairing during Drosophila female achiasmate disjunction has long been established [69], [70] and studies in maize and C. elegans have shown that chromosome pairing is associated with a change in chromatin conformation [71], [72]. In plants, strong evidence for a role of chromatin remodeling in pairing comes from analyses in wheat, where the recognition and pairing of homologous chromosomes at telomeres and centromeres triggers a conformational change in adjacent chromatin that is suggested to further facilitate homologous chromosome recognition and recombination [73]–[75]. Whether such mechanisms also occur in Arabidopsis is unknown. The existence of a centromeric coupling mechanism that is independent of recombination suggests a role for heterochromatin and chromatin structure in the homology search also in Arabidopsis and this may not be restricted to centromeres. In this respect it is interesting to note that in most Arabidopdis ecotypes, cytological observations show the two NORs to be very frequently associated as one large knob [52], [76], [77]. This characteristic association of the nucleolar heterochromatin present on chromosomes 2 and 4 was also observed in all of the mutants studied in this work, indicating that this association is independent of recombination.
Moreover, notwithstanding the non-homologous chromosome pairing observed in Arabidopsis mnd1 and hop2/ahp2 mutants [78]–[81], in ahp2 mutants the NOR-bearing arms of chromosomes 2 and 4 exhibit stabilised homologous pairing and synapsis [80]. These observations strongly suggest a role for heterochromatin in pairing in Arabidopsis. Uncovering the precise role of heterochromatin in meiotic pairing will require further studies such as analysis of mutants with altered heterochromatin configuration or chromatin remodeling capacity.
Materials and Methods
Plant Material and Growth Conditions
All Arabidopsis thaliana plants used in this study were of Columbia ecotype with the exception of dmc1 (Wassilewskija). Arabidopsis xrcc3, rad51C, rad51, dmc1, rad51 dmc1, and spo11-1-2 mutants and PCR genotyping have been described previously [29], [32], [44], [46], [47], [50], [82]. The dmc1 allele used in this experiment (dmc1 and rad51 dmc1 mutants) comes from the Wassilewskija (Ws) background which only has 2 5S rDNA loci, on chromosomes 4 and 5. The DMC1 locus is situated on the same chromosome (III) as the 5S rDNA locus and we verified that the rad51 dmc1 mutant lines used here have 4 rDNA FISH foci in interphase and leptotene nuclei (unpaired chromosomes). Seeds were sown onto soil, stratified in water at 4°C for 2 days and grown in a greenhouse with a 16/8 h light∶dark photoperiod; temperature 23°C; and approximately 60% relative humidity.
Fluorescent In Situ Hybridisation
Chromosome preparation by spreading
Chromosome spreads were prepared according to Ross et al. [77] with the modifications introduced by Fransz et al. [52]. Briefly, whole inflorescences were fixed in absolute ethanol/glacial acetic acid (3∶1) for 3×30 min and stored at 4°C. Immature flower buds were rinsed twice at room temperature in distilled water for 5 min followed by two washes in 1× citrate buffer for 5 min. Buds of appropriate size were selected under a binocular microscope and incubated for 3 hr. 30 mins. on a slide in 100 µl of enzyme mixture (0.3% w/v cellulase (Sigma), 0.3% w/v pectolyase (Sigma) and 0.3% cytohelicase (Sigma) in a moist chamber at 37°C. Each bud was then softened in 15 µl 60% acetic acid on a microscopic slide at 45°C, fixed with ice-cold ethanol/glacial acetic acid (3∶1) and air dried. Slides were then rinsed in Coplin jar for 2 mins. in distilled water, 10 mins. in 4% paraformaldehyde in 1× PBS and finally rinsed for 5 min in distilled water; air-dried and stored at 4°C for further use.
Probe preparation and labelling
The following DNA probes were used: a centromeric specific probe (containing two copies of the 180 bp repeat) [83], a 5S rDNA probe, BAC probes T10F5 (chromosome 2 pericentromere), F12C20 (chromosome 2 long arm), F12K11 (chromosome 1 left-arm), F2J6 (chromosome 1 pericentromere), and T12C22 (chromosome 1 pericentromere). BACs were obtained from the Ohio Arabidopsis Stock Center, ABRC (http://abrc.osu.edu).
DNA from BAC clones was prepared using QIAGEN Large-Construct Kit according to the manufacturer's instructions. BAC probes were labelled with either biotin-dUTP or digoxygenin-dUTP using a Nick-Translation kit (ROCHE) following manufacturer's recommendations. The 180 pb and 5S rDNA probes were labelled by PCR incorporation of dUTP-biotin or dUTP-Digoxygenin according to Lysak et al. (2006). Probes were made with 1 µl of PCR product and/or 3 µl of Nick-Translation product, HB50 buffer to 10 µl and 10 µl Dextran Sulfate 20% in HB50. Probes were used either immediately or stored at −20°C until use. In Situ Hybridisation and fluorescence detection of hybridised probes were performed according to Lysak et al. [84].
Microscopy
All observations were made with a motorised Zeiss AxioImager.Z1 epifluorescence microscope (Carl Zeiss AG, Germany) using a PL Apochromat 100X/1.40 oil objective. Photographs were taken with an AxioCam Mrm camera (Zeiss) and appropriate Zeiss filter sets adapted for the fluorochromes used : filter set 25HE (DAPI), filter set 38HE (Alexa 488) and filter set 43HE (Texas-Red). Captured images were further processed with AxioVision 4.6.2. and Adobe Photoshop CS4 software.
Supporting Information
{kind=link}
{kind=link}
Zdroje
1. HassoldTHallHHuntP 2007 The origin of human aneuploidy: where we have been, where we are going. Hum Mol Genet 16 Spec No. 2 R203 208
2. BozzaCGPawlowskiWP 2008 The cytogenetics of homologous chromosome pairing in meiosis in plants. Cytogenet Genome Res 120 313 319
3. MooreGShawP 2009 Improving the chances of finding the right partner. Curr Opin Genet Dev 19 99 104
4. OsmanKHigginsJDSanchez-MoranEArmstrongSJFranklinFC 2011 Pathways to meiotic recombination in Arabidopsis thaliana. New Phytologist 190 523 544
5. TsaiJHMcKeeBD 2011 Homologous pairing and the role of pairing centers in meiosis. J Cell Sci 124 1955 1963
6. ZetkaM 2009 Homologue pairing, recombination and segregation in Caenorhabditis elegans. Genome Dyn 5 43 55
7. ScherthanH 2007 Telomere attachment and clustering during meiosis. Cell Mol Life Sci 64 117 124
8. BardhanAChuongHDawsonDS 2010 Meiotic cohesin promotes pairing of nonhomologous centromeres in early meiotic prophase. Molecular Biology of the Cell 21 1799 1809
9. ObesoDDawsonDS 2010 Temporal characterization of homology-independent centromere coupling in meiotic prophase. PloS ONE 5 e10336 doi:10.1371/journal.pone.0010336
10. TsubouchiTMacqueenAJRoederGS 2008 Initiation of meiotic chromosome synapsis at centromeres in budding yeast. Genes Dev 22 3217 3226
11. TsubouchiTRoederGS 2005 A synaptonemal complex protein promotes homology-independent centromere coupling. Science 308 870 873
12. FalkJEChanACHoffmannEHochwagenA 2010 A Mec1 - and PP4-dependent checkpoint couples centromere pairing to meiotic recombination. Developmental Cell 19 599 611
13. Martinez-PerezEShawPMooreG 2001 The Ph1 locus is needed to ensure specific somatic and meiotic centromere association. Nature 411 204 207
14. Martinez-PerezEShawPReaderSAragon-AlcaideLMillerT 1999 Homologous chromosome pairing in wheat. J Cell Sci 112 Pt 11 1761 1769
15. RonceretADoutriauxMPGolubovskayaINPawlowskiWP 2009 PHS1 regulates meiotic recombination and homologous chromosome pairing by controlling the transport of RAD50 to the nucleus. Proc Natl Acad Sci U S A 106 20121 20126
16. StewartMNDawsonDS 2008 Changing partners: moving from non-homologous to homologous centromere pairing in meiosis. Trends in Genetics 24 564 573
17. Arabidopsis-Genome-Initiative 2000 Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408 796 815
18. CopenhaverGPNickelKKuromoriTBenitoMIKaulS 1999 Genetic definition and sequence analysis of Arabidopsis centromeres. Science 286 2468 2474
19. MartienssenR 1998 Transposons, DNA methylation and gene control. Trends in genetics : TIG 14 263 264
20. RoundEKFlowersSKRichardsEJ 1997 Arabidopsis thaliana centromere regions: genetic map positions and repetitive DNA structure. Genome Research 7 1045 1053
21. NagakiKWallingJHirschCJiangJMurataM 2009 Structure and evolution of plant centromeres. Progress in Molecular and Subcellular Biology 48 153 179
22. BugreevDVPezzaRJMazinaOMVoloshinONCamerini-OteroRD 2011 The resistance of DMC1 D-loops to dissociation may account for the DMC1 requirement in meiosis. Nature Structural & Molecular Biology 18 56 60
23. NealeMJKeeneyS 2006 Clarifying the mechanics of DNA strand exchange in meiotic recombination. Nature 442 153 158
24. CarballoJAJohnsonALSedgwickSGChaRS 2008 Phosphorylation of the axial element protein Hop1 by Mec1/Tel1 ensures meiotic interhomolog recombination. Cell 132 758 770
25. NiuHWanLBaumgartnerBSchaeferDLoidlJ 2005 Partner choice during meiosis is regulated by Hop1-promoted dimerization of Mek1. Molecular Biology of the Cell 16 5804 5818
26. HoHCBurgessSM 2011 Pch2 acts through Xrs2 and Tel1/ATM to modulate interhomolog bias and checkpoint function during meiosis. PLoS Genet 7 e1002351 doi:10.1371/journal.pgen.1002351
27. BusyginaVSaroDWilliamsGLeungWKSayAF 2011 Novel attributes of Hed1 affect dynamics and activity of the Rad51 presynaptic filament during meiotic recombination. Journal of Biological Chemistry 287 1566 1575
28. BusyginaVSehornMGShiIYTsubouchiHRoederGS 2008 Hed1 regulates Rad51-mediated recombination via a novel mechanism. Genes & Development 22 786 795
29. CouteauFBelzileFHorlowCGrandjeanOVezonD 1999 Random chromosome segregation without meiotic arrest in both male and female meiocytes of a dmc1 mutant of Arabidopsis. Plant Cell 11 1623 1634
30. PradilloMLopezELinaceroRRomeroCCunadoN 2011 Together yes, but not coupled: new insights into the roles of RAD51 and DMC1 in plant meiotic recombination. Plant Journal Online publication. doi: 10.1111/j.1365-1313X.2011.04845.x
31. Sanchez-MoranESantosJLJonesGHFranklinFC 2007 ASY1 mediates AtDMC1-dependent interhomolog recombination during meiosis in Arabidopsis. Genes Dev 21 2220 2233
32. LiWChenCMarkmann-MulischUTimofejevaLSchmelzerE 2004 The Arabidopsis AtRAD51 gene is dispensable for vegetative development but required for meiosis. Proc Natl Acad Sci U S A 101 10596 10601
33. SiaudNDrayEGyIGerardETakvorianN 2004 Brca2 is involved in meiosis in Arabidopsis thaliana as suggested by its interaction with Dmc1. EMBO J 23 1392 1401
34. BadieSLiaoCThanasoulaMBarberPHillMA 2009 RAD51C facilitates checkpoint signaling by promoting CHK2 phosphorylation. J Cell Biol 185 587 600
35. BishopDKEarUBhattacharyyaACalderoneCBeckettM 1998 Xrcc3 is required for assembly of Rad51 complexes in vivo. J Biol Chem 273 21482 21488
36. MassonJYTarsounasMCStasiakAZStasiakAShahR 2001 Identification and purification of two distinct complexes containing the five RAD51 paralogs. Genes Dev 15 3296 3307
37. TarsounasMDaviesAAWestSC 2004 RAD51 localization and activation following DNA damage. Philos Trans R Soc Lond B Biol Sci 359 87 93
38. van VeelenLRCervelliTvan de RaktMWTheilAFEssersJ 2005 Analysis of ionizing radiation-induced foci of DNA damage repair proteins. Mutat Res 574 22 33
39. van VeelenLREssersJvan de RaktMWOdijkHPastinkA 2005 Ionizing radiation-induced foci formation of mammalian Rad51 and Rad54 depends on the Rad51 paralogs, but not on Rad52. Mutat Res 574 34 49
40. KuznetsovSPellegriniMShudaKFernandez-CapetilloOLiuY 2007 RAD51C deficiency in mice results in early prophase I arrest in males and sister chromatid separation at metaphase II in females. The Journal of Cell Biology 176 581 592
41. KuznetsovSGHainesDCMartinBKSharanSK 2009 Loss of Rad51c leads to embryonic lethality and modulation of Trp53-dependent tumorigenesis in mice. Cancer Research 69 863 872
42. SharanSKKuznetsovSG 2007 Resolving RAD51C function in late stages of homologous recombination. Cell Division 2 15
43. AbeKOsakabeKNakayamaSEndoMTagiriA 2005 Arabidopsis RAD51C gene is important for homologous recombination in meiosis and mitosis. Plant Physiol 139 896 908
44. BleuyardJYGallegoMESavignyFWhiteCI 2005 Differing requirements for the Arabidopsis Rad51 paralogs in meiosis and DNA repair. Plant J 41 533 545
45. BleuyardJYGallegoMEWhiteCI 2004 The atspo11-1 mutation rescues atxrcc3 meiotic chromosome fragmentation. Plant Mol Biol 56 217 224
46. BleuyardJYWhiteCI 2004 The Arabidopsis homologue of Xrcc3 plays an essential role in meiosis. EMBO J 23 439 449
47. LiWYangXLinZTimofejevaLXiaoR 2005 The AtRAD51C gene is required for normal meiotic chromosome synapsis and double-stranded break repair in Arabidopsis. Plant Physiol 138 965 976
48. LiuYMassonJYShahRO'ReganPWestSC 2004 RAD51C is required for Holliday junction processing in mammalian cells. Science 303 243 246
49. LiuYTarsounasMO'ReganPWestSC 2007 Role of RAD51C and XRCC3 in genetic recombination and DNA repair. J Biol Chem 282 1973 1979
50. VignardJSiwiecTChelyshevaLVrielynckNGonordF 2007 The interplay of RecA-related proteins and the MND1-HOP2 complex during meiosis in Arabidopsis thaliana. PLoS Genet 3 e176 doi:10.1371/journal.pgen.0030176
51. ArmstrongSJFranklinFCJonesGH 2001 Nucleolus-associated telomere clustering and pairing precede meiotic chromosome synapsis in Arabidopsis thaliana. J Cell Sci 114 4207 4217
52. FranszPArmstrongSAlonso-BlancoCFischerTCTorres-RuizRA 1998 Cytogenetics for the model system Arabidopsis thaliana. Plant J 13 867 876
53. PradilloMSantosJL 2011 The template choice decision in meiosis: is the sister important? Chromosoma 120 447 454
54. DernburgAFMcDonaldKMoulderGBarsteadRDresserM 1998 Meiotic recombination in C. elegans initiates by a conserved mechanism and is dispensable for homologous chromosome synapsis. Cell 94 387 398
55. McKimKSGreen-MarroquinBLSekelskyJJChinGSteinbergC 1998 Meiotic synapsis in the absence of recombination. Science 279 876 878
56. ScherthanH 2001 A bouquet makes ends meet. Nat Rev Mol Cell Biol 2 621 627
57. KempBBoumilRMStewartMNDawsonDS 2004 A role for centromere pairing in meiotic chromosome segregation. Genes Dev 18 1946 1951
58. TakeoSLakeCMMorais-de-SaESunkelCEHawleyRS 2011 Synaptonemal complex-dependent centromeric clustering and the initiation of synapsis in Drosophila oocytes. Curr Biol 21 1845 1851
59. TannetiNSLandyKJoyceEFMcKimKS 2011 A Pathway for Synapsis Initiation during Zygotene in Drosophila Oocytes. Curr Biol 21 1852 1857
60. ChurchKMoensPB 1976 Centromere behavior during interphase and meiotic prophase in Allium fistulosum from 3-D, EM reconstruction. Chromosoma 56 249 263
61. CaiXDongFEdelmannREMakaroffCA 2003 The Arabidopsis SYN1 cohesin protein is required for sister chromatid arm cohesion and homologous chromosome pairing. J Cell Sci 116 2999 3007
62. YangXBoatengKAStrittmatterLBurgessRMakaroffCA 2009 Arabidopsis separase functions beyond the removal of sister chromatid cohesion during meiosis. Plant Physiol 151 323 333
63. YangXTimofejevaLMaHMakaroffCA 2006 The Arabidopsis SKP1 homolog ASK1 controls meiotic chromosome remodeling and release of chromatin from the nuclear membrane and nucleolus. J Cell Sci 119 3754 3763
64. RobertsNYOsmanKArmstrongSJ 2009 Telomere distribution and dynamics in somatic and meiotic nuclei of Arabidopsis thaliana. Cytogenet Genome Res 124 193 201
65. BhallaNDernburgAF 2008 Prelude to a division. Annual Review of Cell and Developmental Biology 24 397 424
66. Howard-TillRALukaszewiczALoidlJ 2011 The recombinases Rad51 and Dmc1 play distinct roles in DNA break repair and recombination partner choice in the meiosis of Tetrahymena. PLoS Genet 7 e1001359 doi:10.1371/journal.pgen.1001359
67. HunterNKlecknerN 2001 The single-end invasion: an asymmetric intermediate at the double-strand break to double-holliday junction transition of meiotic recombination. Cell 106 59 70
68. ShinoharaMGasiorSLBishopDKShinoharaA 2000 Tid1/Rdh54 promotes colocalization of rad51 and dmc1 during meiotic recombination. Proc Natl Acad Sci U S A 97 10814 10819
69. DernburgAFSedatJWHawleyRS 1996 Direct evidence of a role for heterochromatin in meiotic chromosome segregation. Cell 86 135 146
70. KarpenGHLeMHLeH 1996 Centric heterochromatin and the efficiency of achiasmate disjunction in Drosophila female meiosis. Science 273 118 122
71. DaweRKSedatJWAgardDACandeWZ 1994 Meiotic chromosome pairing in maize is associated with a novel chromatin organization. Cell 76 901 912
72. MacQueenAJVilleneuveAM 2001 Nuclear reorganization and homologous chromosome pairing during meiotic prophase require C. elegans chk-2. Genes Dev 15 1674 1687
73. ColasIShawPPrietoPWanousMSpielmeyerW 2008 Effective chromosome pairing requires chromatin remodeling at the onset of meiosis. Proc Natl Acad Sci U S A 105 6075 6080
74. PrietoPMooreGReaderS 2005 Control of conformation changes associated with homologue recognition during meiosis. Theor Appl Genet 111 505 510
75. PrietoPShawPMooreG 2004 Homologue recognition during meiosis is associated with a change in chromatin conformation. Nat Cell Biol 6 906 908
76. PecinkaASchubertVMeisterAKrethGKlatteM 2004 Chromosome territory arrangement and homologous pairing in nuclei of Arabidopsis thaliana are predominantly random except for NOR-bearing chromosomes. Chromosoma 113 258 269
77. RossKJFranszPJonesGH 1996 A light microscopic atlas of meiosis in Arabidopsis thaliana. Chromosome Res 4 507 516
78. KerzendorferCVignardJPedrosa-HarandASiwiecTAkimchevaS 2006 The Arabidopsis thaliana MND1 homologue plays a key role in meiotic homologous pairing, synapsis and recombination. J Cell Sci 119 2486 2496
79. PanoliAPRaviMSebastianJNishalBReddyTV 2006 AtMND1 is required for homologous pairing during meiosis in Arabidopsis. BMC Mol Biol 7 24
80. StronghillPPathanNHaHSupijonoEHasenkampfC 2010 Ahp2 (Hop2) function in Arabidopsis thaliana (Ler) is required for stabilization of close alignment and synaptonemal complex formation except for the two short arms that contain nucleolus organizer regions. Chromosoma 119 443 458
81. SchommerCBevenALawrensonTShawPSablowskiR 2003 AHP2 is required for bivalent formation and for segregation of homologous chromosomes in Arabidopsis meiosis. Plant J 36 1 11
82. GrelonMVezonDGendrotGPelletierG 2001 AtSPO11-1 is necessary for efficient meiotic recombination in plants. EMBO J 20 589 600
83. Martinez-ZapaterJEstelleMSomervilleC 1986 A highly repeated DNA sequence in Arabidopsis thaliana. Molec Gen Genet 204 417 423
84. LysakMFranszPSchubertI 2006 Cytogenetic analyses of Arabidopsis. Methods Mol Biol 323 173 186
85. PawlowskiWPGolubovskayaINTimofejevaLMeeleyRBSheridanWF 2004 Coordination of meiotic recombination, pairing, and synapsis by PHS1. Science 303 89 92
86. BleuyardJYGallegoMEWhiteCI 2004 Meiotic defects in the Arabidopsis rad50 mutant point to conservation of the MRX complex function in early stages of meiotic recombination. Chromosoma 113 197 203
87. YangXBoatengKAYuanLWuSBaskinTI 2011 The radially swollen 4 separase mutation of Arabidopsis thaliana blocks chromosome disjunction and disrupts the radial microtubule system in meiocytes. PLoS ONE 6 e19459 doi:10.1371/journal.pone.0019459
88. BoatengKAYangXDongFOwenHAMakaroffCA 2008 SWI1 is required for meiotic chromosome remodeling events. Mol Plant 1 620 633
89. MercierRArmstrongSJHorlowCJacksonNPMakaroffCA 2003 The meiotic protein SWI1 is required for axial element formation and recombination initiation in Arabidopsis. Development 130 3309 3318
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2012 Číslo 4
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- A Coordinated Interdependent Protein Circuitry Stabilizes the Kinetochore Ensemble to Protect CENP-A in the Human Pathogenic Yeast
- Coordinate Regulation of Lipid Metabolism by Novel Nuclear Receptor Partnerships
- Defective Membrane Remodeling in Neuromuscular Diseases: Insights from Animal Models
- Formation of Rigid, Non-Flight Forewings (Elytra) of a Beetle Requires Two Major Cuticular Proteins
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy