
B Chromosomes Have a Functional Effect on Female Sex Determination in Lake Victoria Cichlid Fishes
The endemic cichlid fishes in Lake Victoria are a model system for speciation through adaptive radiation. Although the evolution of the sex-determination system may also play a role in speciation, little is known about the sex-determination system of Lake Victoria cichlids. To understand the evolution of the sex-determination system in these fish, we performed cytogenetic analysis in 11 cichlid species from Lake Victoria. B chromosomes, which are present in addition to standard chromosomes, were found at a high prevalence rate (85%) in these cichlids. In one species, B chromosomes were female-specific. Cross-breeding using females with and without the B chromosomes demonstrated that the presence of the B chromosomes leads to a female-biased sex ratio in this species. Although B chromosomes were believed to be selfish genetic elements with little effect on phenotype and to lack protein-coding genes, the present study provides evidence that B chromosomes have a functional effect on female sex determination. FISH analysis using a BAC clone containing B chromosome DNA suggested that the B chromosomes are derived from sex chromosomes. Determination of the nucleotide sequences of this clone (104.5 kb) revealed the presence of several protein-coding genes in the B chromosome, suggesting that B chromosomes have the potential to contain functional genes. Because some sex chromosomes in amphibians and arthropods are thought to be derived from B chromosomes, the B chromosomes in Lake Victoria cichlids may represent an evolutionary transition toward the generation of sex chromosomes.
Published in the journal:
. PLoS Genet 7(8): e32767. doi:10.1371/journal.pgen.1002203
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002203
Summary
The endemic cichlid fishes in Lake Victoria are a model system for speciation through adaptive radiation. Although the evolution of the sex-determination system may also play a role in speciation, little is known about the sex-determination system of Lake Victoria cichlids. To understand the evolution of the sex-determination system in these fish, we performed cytogenetic analysis in 11 cichlid species from Lake Victoria. B chromosomes, which are present in addition to standard chromosomes, were found at a high prevalence rate (85%) in these cichlids. In one species, B chromosomes were female-specific. Cross-breeding using females with and without the B chromosomes demonstrated that the presence of the B chromosomes leads to a female-biased sex ratio in this species. Although B chromosomes were believed to be selfish genetic elements with little effect on phenotype and to lack protein-coding genes, the present study provides evidence that B chromosomes have a functional effect on female sex determination. FISH analysis using a BAC clone containing B chromosome DNA suggested that the B chromosomes are derived from sex chromosomes. Determination of the nucleotide sequences of this clone (104.5 kb) revealed the presence of several protein-coding genes in the B chromosome, suggesting that B chromosomes have the potential to contain functional genes. Because some sex chromosomes in amphibians and arthropods are thought to be derived from B chromosomes, the B chromosomes in Lake Victoria cichlids may represent an evolutionary transition toward the generation of sex chromosomes.
Introduction
The species flock of endemic cichlid fishes in Lake Victoria is the largest known example of recent adaptive radiation and has been highlighted as a model system for the genetic study of speciation [1]–[3]. The evolution of the sex-determination system is suggested to drive the speciation of the cichlids because novel sex-determination genes tend to be associated with novel body colors which can drive reproductive isolation [1], [4]. Recent genetic studies suggest that the sex chromosomes of cichlids have turned over rapidly and that the sex-determination locus is different among species and populations [5]–[7]. Among closely related species of Lake Malawi cichlids, which are sister species of Lake Victoria cichlids, two unlinked sex-determination loci were reported [5]–[6]. In a few populations of Lake Malawi cichlids, multiple interacting loci control sex determination [6], suggesting an ongoing transition in the sex chromosomes. In one species of Lake Victoria cichlids, a sex-determination locus was inferred based on the analysis of a sex-linked phenotype [7]. The sex-determination system has been studied, however, in only a few species of Lake Victoria cichlids. Although cytogenetic analysis is important for studying the sex-determination system, it has not been performed in wild populations in Lake Victoria cichlids. However, a cytogenetic analysis of one species of Lake Victoria cichlid obtained from a commercial source revealed the presence of B chromosomes [8].
B chromosomes are chromosomes found in addition to the standard chromosomes (A chromosomes). They occur in many groups of fungi, plants and animals (10 species, >1300 species, and >500 species, respectively) and vary in number among individuals within a population [9]. They are dispensable for the normal life cycle of host individuals [9]. In most cases, the presence of B chromosomes has no effect on the host phenotype or is deleterious when the number per cell increases [10]. Because of their extensive distribution among many organisms, the possibility that B chromosomes have a beneficial effect on their hosts has been argued [10], but there has been no concrete evidence for this effect. Instead, the existence of B chromosomes has been explained by their selfish behavior such as their non-Mendelian inheritance and accumulation in the germ line [11]. B chromosomes are thought to be composed of repetitive sequences and to lack protein-coding genes [9], and they often form a heterochromatic block [10].
Here we analyzed the karyotypes of wild populations of 11 cichlid species in Lake Victoria. Most individuals possessed B chromosomes of varying sizes. In one species, B chromosomes were specific to females. We performed further analysis of the B chromosomes in this species to reveal their function in sex determination.
Results/Discussion
High Prevalence of B Chromosomes in Lake Victoria Cichlids
We collected live individuals from six localities (Figure 1) in Lake Victoria during several expeditions in 2005–2007 and 2009 to prepare the chromosome specimens and to produce breeding lines in the laboratory. We analyzed the karyotypes of wild-caught individuals of 11 cichlid species in Lake Victoria (a total of 51 individuals). The chromosome number varied from 44 to 47 and was not specific to the species or populations analyzed (Table 1). To reveal the inheritance pattern of this chromosome number difference, we analyzed the karyotypes of F1 and F2 generation of Lithochromis rubripinnis Seehausen, Lippitsch and Bouton, 1998 [12] from the Matumbi Island population (Figure 2A–2D; Table S1). The results indicated that chromosome number varied within clutches by the presence of two small chromosomes, designated as B1 (the larger) and B2 (the smaller). These data suggested that the chromosome number in the wild-caught individuals varies based on the number of B chromosomes. Their variations in size make it difficult to identify the B chromosomes precisely by morphology.



B1 and B2 chromosomes of L. rubripinnis were isolated by microdissection and used as probes for FISH analysis. Both of the probes (B1 and B2 probes) painted both B1 and B2 chromosomes but not the other chromosomes (Figure 2E and 2F). Although B1 chromosomes are larger than B2 chromosomes, B2 probe painted whole region of B1 chromosomes. These results indicated that the B1 and B2 chromosomes shared repetitive sequences that are specific to B chromosomes. We performed chromosome painting for the wild-caught Lake Victoria cichlids using the B1 probes. The number of chromosomes painted varied from zero to three, whereas the number of unpainted chromosomes (44) was the same among all individuals (Figure 3; Figure S1; Table 1; Table S2). These results demonstrate that these cichlids possess 44 A chromosomes and 0–3 B chromosomes. B chromosomes were different in size among populations (Figure S2). The B1 probe painted the whole region of all B chromosomes regardless of their size again (Figure S2A), suggesting that B chromosomes are composed of the same B-specific repetitive sequences. B chromosomes were observed in all examined populations at a high prevalence rate (86%; Table 1). The mean number of B chromosomes per individual, however, was low (1.45), suggesting that the accumulation of B chromosomes is restricted. A low prevalence rate of B chromosomes (40%) was reported in breeding individuals of Haplochromis obliquidens obtained from the commercial source [8]. This different prevalence rate raises the possibility that B chromosomes may possess some functions that are not essential for the survival of host individuals but are advantageous in wild populations.
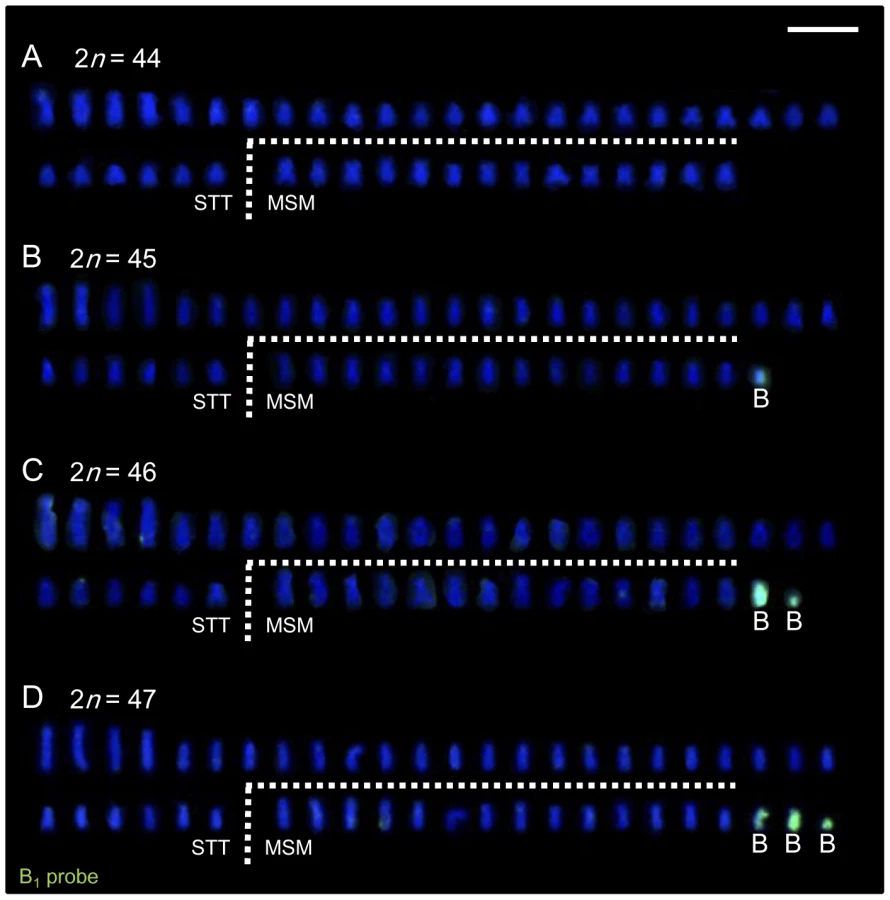
Female-Specific B Chromosomes Have a Functional Effect on Sex Determination
Although B chromosomes were found in both males and females of most other species (Table S2), in wild individuals of L. rubripinnis from the Matumbi island population, all females (N = 4) possessed B chromosomes, whereas males possessed no B chromosomes (N = 2; Figure 4A). We confirmed this female-biased possession of B chromosomes by comparison of karyotypes of males and those of females using F1 and F2 generations in this population. The results showed that almost all females (in a total of N = 24) possessed B chromosomes, whereas no males (in a total of N = 10) possessed B chromosomes (Figure 4A; F1 and F2), indicating that B chromosomes are closely associated with females in this population. We confirmed the absence of B chromosomes in germ cells of F2 males by meiotic analysis (N = 3; Figure S3), indicating that the B chromosomes were specific to females in this population. To reveal the functional effect of B chromosomes on sex determination, we performed cross-breeding experiments between males without B chromosomes and females with different numbers of B chromosomes (0, 1, or 2) and scored the sex ratio of the offspring. The dams without B chromosomes (B−) generated nearly 1∶1 offspring sex ratios (proportion of females: 38% in cross #1 and 50% in #2; Figure 4B). These results indicate that one of the sex determination loci is located on an as yet unknown A chromosome. In contrast, the dams with a B chromosome(s) (B+) generated female-biased sex ratios. The proportion of females in the offspring was 74%, 91%, 79%, and 100% in cross #3 (B = 1 dam), #4 (B = 1 dam), #5 (B = 1 dam), and #6 (B = 2 dam), respectively (Figure 4B). The correlation of the number of B chromosomes in the dam with the proportion of females in the offspring suggests that the presence of female-specific B chromosomes in this species leads to a female-biased sex ratio. Karyotype analysis of the offspring showed ubiquitous distribution of B chromosomes in offspring from a cross with a skewed sex ratio (cross #6; Figure 4A) and an absence of B chromosomes in offspring from a cross with a nearly 1∶1 sex ratio (cross #1; Figure 4A), confirming the effect of the B chromosomes on sex determination. Although we cannot exclude the possibility that the B chromosomes have a male-specific lethal effect, this is unlikely, because we did not observe a higher death rate in the offspring of the B+ dam than in those of the B− dam (Figure 4B, see legend). According to these results, we concluded that B chromosomes have a functional effect on female sex determination. In the offspring of the dam with two B chromosomes (cross #6), four females (29%) possessed two B chromosomes (Figure 4A), indicating non-Mendelian inheritance of the B chromosomes. These observations (i.e., dispensability for host survival and non-Mendelian inheritance) confirmed that these female-specific chromosomes have features that are unique to B chromosomes.

Because all the B chromosomes of several species analyzed here were painted specifically by the B1 probe (Figure 3A–3D; Figure S1), they share sequences derived from the same ancestor. Because males in the other populations and species possessed B chromosomes (Table S2), B chromosomes in males have a different effect on sex determination than do the female-specific B chromosomes in spite of their having a shared ancestry. We cannot determine whether the B chromosomes in females in the other populations and species that we examined here are female specific or not. How extensively such female-specific B chromosomes are distributed in cichlids is currently unknown.
The New Zealand frog [13] possesses W chromosomes, which are functionally very similar to the female-specific B chromosomes described above. This species shows a 0W female/00 male sex-determination system. It additionally has B chromosomes that are not female specific and that share partial DNA components with the W chromosome [14]. The W chromosomes are not B chromosomes because they are indispensable for this frog. We speculate that the univalent W chromosome in the frog species might have differentiated from a B chromosome. In most cases, B chromosomes have similar features to sex chromosomes in terms of their meiotic behavior, morphology, and heterochromatic state [15], suggesting the possibility of their evolutionary relatedness. Y chromosomes may also be derived from B chromosomes in a few species of arthropods [16]–[17]. The female-specific B chromosome in cichlids studied here seems to be in an evolutionary transition from B chromosome to sex chromosome. A similar contribution of B chromosomes to the sex ratio in the characid fish Astyanax scabripinnis was reported [18], but it is still unclear whether there is a direct connection between the presence of a B chromosome and the sex ratio in those fish. The present case differs from the B chromosome contribution to sex determination found in some arthropods with haplodiploid sex determination because the B chromosome is involved in the exclusion of haploid genomes in those species [19].
Protein-Coding Genes in the B Chromosomes
The functional effect of B chromosomes on sex determination in L. rubripinnis suggested the possibility that these chromosomes might have some functional genes. To isolate partial DNA sequences of the B chromosomes, we performed differential screening. We screened B+ genomic DNA library and isolated DNA fragments hybridized with a B+ genomic probe but not a B− genomic probe. This identified a B chromosome–specific repetitive DNA sequence (named Bseq1; Figure S4). We isolated a BAC clone containing Bseq1 DNA (∼128 kb in total; named B-BAC) from the BAC library constructed from Haplochromis chilotes [20]. We analyzed karyotypes of this H. chilotes strain (N = 6) and confirmed that all individuals possessed two B chromosomes, indicating strong possibility for inclusion of B chromosome DNA in this BAC library (data not shown). We also confirmed that the BAC clone DNA is derived from B chromosomes by sequence analysis (see below). We determined 80% of the B-BAC sequence (104.5 kb; 18 contigs). Repetitive sequences occupied 59% of the sequence (Figure 5A; Table S3). Remarkably, we discovered five different protein-coding genes in this BAC clone, each of which is almost identical to its parental gene present in the A chromosomes (Table 2; Table S4). The gene density in this B-BAC was higher (4.5%; Figure 5A) than the gene density reported in the cichlid genome (<4%) [21]. No nonsense mutations were found in these genes (1581 a.a. in total). Although protein-coding genes have been reported in B chromosomes in three other cases, i.e. the fungus Nectria haematococca [22], several Canidae species [23], [24], and the locust Locusta migratoria [25], they have not been thought to have functional significance. The absence of nonsense mutations in the five protein-coding genes in the B chromosomes of cichlids described here might be an indication that the sequence of the B chromosomes has not degenerated from their ancestral sequence, and thus it appears that functional genes have maintained. However, it is possible that their expression might be suppressed by heterochromatin.
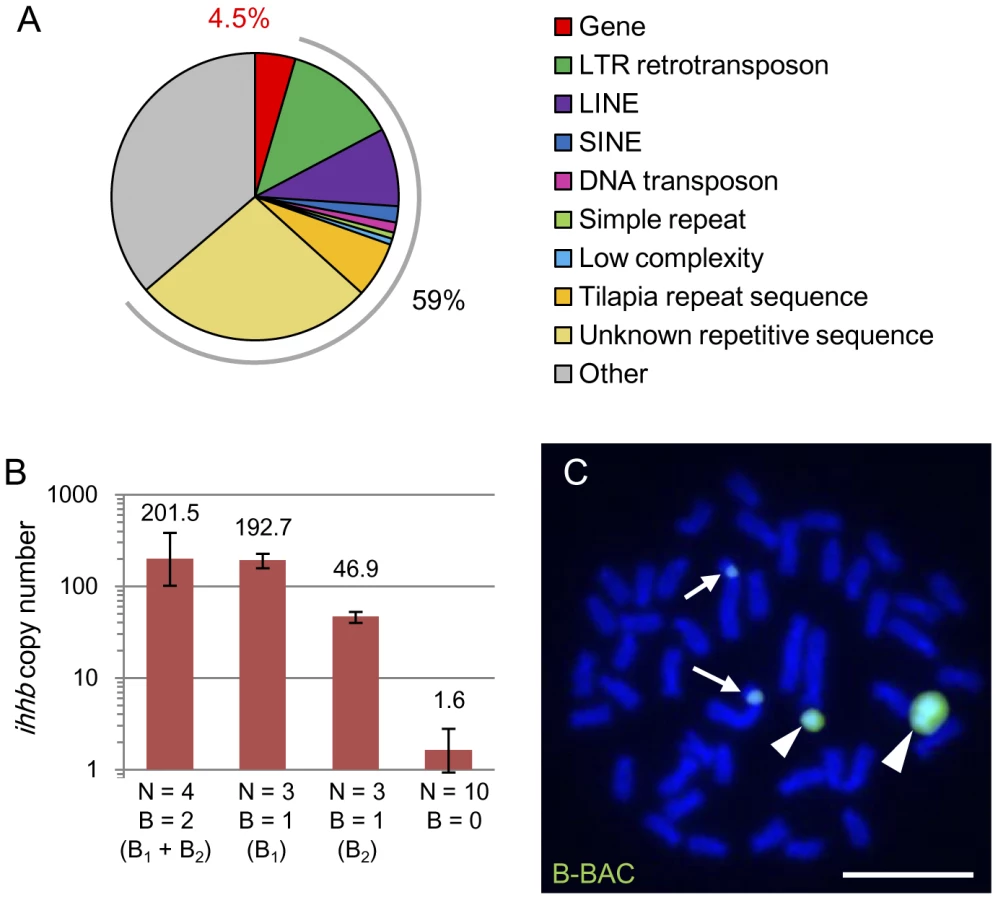

One example of the genes identified in the B chromosome is a morphogenesis-related gene, indian hedgehog b (ihhb). We estimated the copy number of ihhb in the genomes of B+ and B− individuals of L. rubripinnis by quantitative PCR (Figure 5B). The copy numbers of ihhb in the diploid genomes with 2 (B1 and B2), 1 (B1 or B2), and 0 B chromosomes, were estimated at 202, 193, 47, and 1.6, respectively. These results showed that there are >40 copies of the ihhb paralogs on B chromosomes whereas there is a single copy of the ihhb ortholog on the A chromosomes. Sequence analysis provided a tool for distinguishing the paralogs on the B chromosome from the orthologs on the A chromosome. Direct sequencing of ihhb exon 2 and the flanking region (Figure S5; total, 2387 bp) using B+ and B− genomes showed that ihhb orthologs on A chromosomes had a C and T at sites −940 and −88, respectively, in the ihhb region, whereas almost all ihhb paralogs on B chromosomes had a T and C at the same sites (20 individuals each; Figure S5). These results suggested that the ihhb gene was duplicated from an A chromosome to a B chromosome and formed multiple paralogs in B chromosomes. The B-BAC sequence contained the ihhb paralog sequence, indicating that the DNA fragment in the BAC clone was indeed derived from a B chromosome. Phylogenetic analysis of the ihhb regions (Figure S6) confirmed that these paralogs in the B chromosome emerged from their orthologs in the young Lake Victoria cichlid lineage.
The Origin and Evolution of B Chromosomes in Lake Victoria Cichlids
FISH analysis using the B-BAC as a probe for L. rubripinnis showed intense signals on the short arm of the largest chromosome (chromosome 1) as well as on the B chromosomes (Figure 5C), suggesting that chromosome 1 is a strong candidate for the origin of the B chromosomes. Linkage group 3 (LG3) is a sex chromosome in Tilapia, which is a species that is related to Lake Victoria cichlids [26]. Markers for LG3 (GM385 locus and dmrt4 (dmo) gene) [26] were mapped to chromosome 1 in Lake Victoria cichlids (Figure S7). These findings suggest that the sex chromosome in Tilapia corresponds to chromosome 1 in Lake Victoria cichlids and that the sex-determination-related gene might be located on chromosome 1 of Lake Victoria cichlids. It is likely that the B chromosome in Lake Victoria cichlids has evolved from a part of chromosome 1 that contains the sex-determination-related gene and ultimately gained a function for sex determination in some lineages (the model is presented in Figure S8). However, we could not find genes related to sex determination in the B-BAC sequence. Further study of the sequence of the B chromosome is required to identify the genes that influence sex determination.
In this paper, we showed the recent evolution of a sex-determination system driven by female-specific B chromosomes in Lake Victoria cichlid fishes. The evolution of a sex-determination system can resolve sexual conflict [5], [27], [28]. Sexual conflict can arise when sexually antagonistic genes, which are beneficial to one sex and detrimental to the other, are found on autosomes. However, sexual conflict can be resolved if a gene experiencing sexual antagonism evolves linkage with a sex-determination gene. In this way, the evolution of a new sex-determination locus might resolve sexual conflict [27]. In fact, the sexual conflict produced by the orange-blotched body color pattern, which is beneficial to females but detrimental to males, has been resolved by the emergence of a new sex-determination locus in Lake Malawi cichlids, and the appearance of this color pattern is female-specific [5]. The evolution of this female-specific body color pattern possibly causes sexual isolation by male mate choice of this pattern in cichlids [7]. A direct association between the evolution of a new sex chromosome and sexual isolation was reported in sticklebacks [28]. By linking with the newly emerged female-specific sex determination locus, the female-beneficial sexual antagonistic traits such as female preference to male coloration that have generally observed in Lake Victoria cichlids [3] might have evolved rapidly and have driven speciation. It is, therefore, important for the study of speciation via sexual isolation to analyze the recent evolution of the sex-determination system caused by the female-specific B chromosomes that we have described here in Lake Victoria cichlids. Further studies of the molecular components of B chromosomes as well as the function of B chromosomes in wild populations of cichlids will shed light on the molecular mechanism of how and why a novel sex-determination system emerged during the evolution of these fish.
Materials and Methods
Animals
We collected live individuals of 11 species from 6 localities in Lake Victoria (Figure 1) during expeditions in 2005–2007 and 2009. The live fish were shipped to the Tokyo Institute of Technology in Japan for chromosome preparation, extraction of genomic DNA, and cross-breeding. The Malawi cichlid species (Cyrtocara moorii, Fossorochromis rostratus, Tyrannochromis macrostoma and Petrotilapia tridentiger) and Tanganyika cichlid species (Simochromis pleurospilus and Perissodus microlepis) were obtained from traders.
Cross-Breeding
We crossed females of Lithochromis rubripinnis with conspecific males. F1 offspring were allowed to sib-mate to produce the F2 generation. Five F3 families (#1, #3, #4, #5, and #6) were produced by controlled crosses of one F2 male without B chromosomes and five F2 females with zero, one, or two B chromosomes. One F4 family (#2) was produced by a controlled cross of F3 parents without B chromosomes. The sex ratios of these F3 families and the F4 family were scored. Sex ratios were defined as the proportion of males in each clutch and were scored by counting the number of males with breeding coloration as described [29]. All fry exhibited cryptic coloration after hatching, but males begin to display breeding coloration 140 days after their birth [29]. Between 140 to 300 days, all males exhibited breeding color, and sex ratios were scored for each clutch within that time period. We confirmed that sex scored by this method was consistent with the gonadal sex by sacrificing 10 B− males, 10 B+ females and 10B− females and observing their gonads.
Chromosome Preparation and Karyotyping
Chromosome preparation was performed as described [30], [31], with modifications. Chromosomes were prepared from cells of the caudal fin. Caudal fin tissue was cut into small pieces and placed on a collagen-coated dish (IWAKI, Tokyo, Japan). The cells were cultured in Leibovitz's L-15 medium (Invitrogen-GIBCO, Carlsbad, CA) supplemented with 20% fetal bovine serum, 1× antibiotic-antimycotic (PSA; Invitrogen-GIBCO), and 0.1 mg/ml kanamycin sulfate (Meiji Seika, Tokyo, Japan) at 28°C. Non-adherent cells appeared and increased from the caudal fin tissue for 30 days after the initiation of the culture. The cells were harvested after colcemid treatment (0.5 µg/ml) for 2 h, suspended in 0.075 M KCl, fixed three times in 3∶1 methanol/acetic acid, and then dropped onto glass slides and air-dried. More than 30 metaphase spreads for each individual were used for karyotyping. The nomenclature of chromosome morphology as suggested by Levan et al. was used [32], providing for two categories with different arm ratios (r): metacentric-submetacentric (MSM, 1<r≤3) and subtelocentric-telocentric (STT, r>3).
B Chromosome Microdissection and Probe Production
B chromosome microdissection and degenerate oligonucleotide–primed PCR (DOP-PCR) were performed as described [33], [34], with modifications. A single microdissected chromosome fragment, which was sufficient to produce the painting probes, was scraped into a tube. DNA from the scraped chromosome was amplified by first-generation DOP-PCR in a final volume of 15 µl containing 1.5 µl Thermo Sequenase DNA polymerase (GE Healthcare, Chalfont St Giles, UK), 1.5 µl Thermo Sequenase reaction buffer, 0.2 mM dNTPs, and 4 µM primer 6MW (5′-CCGACTCGAGNN NNNNATGTGG-3′). The first-generation DOP-PCR was conducted as follows: 10 min at 95°C; 12 cycles at 94°C for 1 min, 2 min at 30°C, a 6-min transition at 30°C–65°C, and a 3-min extension at 65°C; 30 cycles at 94°C for 1 min, 1 min at 56°C, and 3 min at 72°C; a final extension of 8 min at 72°C. The first-generation DOP-PCR product (3 µl) was used for second-generation DOP-PCR in a final volume of 10 µl. The second-generation DOP-PCR conditions were as follows: 5 min at 95°C; 25 cycles of 94°C for 1 min, 1 min at 56°C, and a 3-min extension at 72°C; a final extension of 8 min. The PCR product of the second-generation DOP-PCR (3 µl) was labeled by the third-generation DOP-PCR in a final volume of 10 µl containing 0.12 nmol/µl digoxigenin-11-dUTP (Roche Diagnostics, Basel, Switzerland). The third-generation DOP-PCR conditions were the same as the second-generation DOP-PCR conditions. The product of the third-generation DOP-PCR was used as the B chromosome probe for painting FISH analysis.
The B chromosome repetitive sequence (Bseq1) was amplified using the primers indicated in Table S5 and subcloned into the pGEM-TA plasmid vector (Promega, Madison, WI) to produce the Bseq1 probe. This clone and the BAC clone were labeled by nick translation with biotin-16-dUTP (Roche Diagnostics).
FISH
FISH analysis was performed as described [30], [31], with modifications. Hybridization was carried out at 37°C overnight. The slides that had been hybridized with the biotin - or digoxigenin-labeled probe were stained with fluorescein-conjugated avidin (Vector Laboratories, Burlingame, CA) or fluorescein-conjugated anti-digoxigenin (Roche Diagnostics), respectively, and stained with 0.25 µg/ml DAPI. FISH images were observed under a fluorescence microscope (Carl Zeiss, Oberkochen, Germany) using the 1 and 17 filter sets.
Measurement of Chromosome Size
The actual size of all chromosomes was measured in five metaphase plates in a single individual using Axio Vision (Carl Zeiss). The mean size of the A chromosomes was calculated for each metaphase plate. The ratio of the size of a B chromosome to the mean size of the A chromosomes in the same cell was defined as the relative size of the B chromosome. We averaged the B chromosome sizes across the five metaphase plates. When there was more than one B chromosome in a single cell, we distinguished them by size.
Meiotic Analysis
We analyzed meiotic chromosomes of three F2 males of L. rubripinnis separately. Testes of a single male were nicked and suspended for 90 min in 1% sodium citrate and were fixed for 5 min in 1∶1 ethanol/acetic acid. The testes were placed into 3∶3∶4 ethanol/acetic acid/distilled water to extract the testicular cells. The cells were refixed three times in 1∶1 ethanol/acetic acid and then dropped onto glass slides and air-dried.
Isolation of the Repetitive Sequence Specific to B Chromosomes
We performed differential screening to isolate the DNA fragments from the B chromosome. We first constructed a B+ genomic library. Using the DNeasy kit (QIAGEN, Venlo, Netherlands), B+ genomic DNA was extracted from Lithochromis rufus (2n = 46), which possesses two large B chromosomes. B+ genomic DNA was partially digested for 15 s with Sau3AI and subsequently subcloned into the pUC19 plasmid vector. We extracted the plasmid DNA from more than 400 clones and chose the plasmids with DNA inserts >500 bp (64 clones).
We separated the chosen plasmid DNAs by 1.5% agarose gel electrophoresis. DNA fragments were transferred from the gels to GeneScreen Plus membranes (Perkin-Elmer, Norwalk, CT) in 0.4 M NaOH and 0.6 M NaCl. Membranes were neutralized in 0.5 M Tris-HC1 (pH 7.0) and 1 M NaCl and then dried. We performed electrophoresis and transfer of the DNA fragments twice using the same amount of DNAs for each plasmid to make two copies of the membranes.
Next, we produced the probe of B+ genomic DNA and B− genomic DNA. B− genomic DNA was extracted from L. rubripinnis (2n = 44, without B chromosome). B+ genomic DNA for the probe was extracted from the same individual as that for the genomic library. One microgram of both B+ and B− genomic DNA was labeled for probes with [α-32P]dCTP using the BcaBEST™ labeling kit (Takara, Tokyo, Japan).
Hybridization was performed at 42°C overnight in a solution of 50% (v/v) formamide, 1 M NaCl, 1% SDS, 2× Denhardt's solution, and 100 pg/ml of labeled probe DNA. We used B+ and B− labeled probes for each membrane. After the hybridization, we washed the membranes and detected the signals. We compared the signal intensity of the membranes hybridized with the B+ probe and with the B− probe. Two clones, including the Bseq1 clone, showed stronger signals in the hybridization with B+ than with B−. We determined the sequences of these clones. The primers were designed according to the sequence of the Bseq1 clone (Bseq1F, Bseq1R; Table S5). This region was amplified by PCR using B+ genomic DNA.
Sequence Analysis of the BAC Clone
A BAC clone (B-BAC) containing Bseq1 was screened and isolated from the Haplochromis chilotes BAC library [20]. B-BAC DNA was digested with BglII, BamHI, HindIII, PstI, XbaI, and SphI. Each of the resultant DNA fragments was subcloned into pUC19. We determined the sequences of the DNA fragments inserted into the plasmid. The flanking sequences were determined by direct sequencing using BAC clone DNA. Repeat masking was performed with RepeatMasker ver. open-3.2.9, with a Teleostei repeat library of database ver. RM-20090604 and the –s (slow and most sensitive) option. Subsequent repeat masking was performed under the same conditions using a tilapia repeat sequence library [35], which contains insufficiently characterized repeat sequences. The sequences in which the repetitive sequences were masked were used for a subsequent NCBI BLAST search (http://blast.ncbi.nlm.nih.gov/Blast.cgi) of the whole-genome shotgun data of five Lake Malawi cichlids (Maylandia zebra, Mchenga conophoros, Melanochromis auratus, Labeotropheus fuelleborni, and Rhamphochromis esox; 21) to identify unknown repetitive elements, because a number of uncharacterized repeats in the B-BAC sequences from H. chilotes were not masked with the RepeatMasker. The partial B-BAC regions that hit at least four different loci in one Lake Malawi cichlid with an E-value of <10−4 over 35 nt were chosen as unknown repetitive sequences. The sequences in which both known and unknown repetitive sequences were masked were used for a subsequent NCBI BLASTX search in the non-redundant protein sequence database of bony fishes with an E-value cutoff of 10−10. To analyze the coding regions of the five protein-coding sequences precisely, we performed a homology search with them in translated B-BAC sequences using Genetyx ver. 6.1.0 (Genetyx, Tokyo, Japan).
Sequence Analysis of ihhb
The primers for the ihhb region are indicated in Table S5 and Figure S5. Two fragments of the ihhb region were defined by the position of the primer pair Bseq1F and ihhbR5 and the pair ihhbF3 and ihhbR8. Each fragment was amplified from the genomic DNA by PCR. We purified the PCR products and determined the sequences using the primers Bseq1F, Bseq1R, ihhbF1, ihhbF2, ihhbF4, ihhbF5, ihhbF6, ihhbR1, ihhbR2, ihhbR3, ihhbR4, ihhbR5, ihhbR7, and ihhbR8. When the sequences included heterogeneous sites, we subcloned the PCR products into the pGEM-TA plasmid vector and determined the sequences of several clones to obtain the sequence information and eliminate PCR errors. Phylogenetic analysis was performed using MEGA4.0 [36]. Phylogenetic trees were obtained by neighbor-joining (NJ), minimum-evolution (ME), and maximum-parsimony (MP) methods with bootstrap tests.
Quantitative Real-Time PCR (qPCR)
The partial fragment of the ihhb region was amplified by PCR using the primers ihhbF3 and ihhbR7 and was cloned into the pGEM-TA plasmid vector for the standard control for calibration of qPCR. qPCR was performed in a 12.5-µl reaction using the Quantitative SYBR Green RT-PCR kit (Applied Biosystems, Foster City, CA). PCR amplification and product detection were conducted using Thermal Cycler Dice (TaKaRa) and the primers ihhbF4 and ihhbR6, which were derived from intron 1 and exon 2 of ihhb, respectively (Figure S5). The sequences of the primers completely matched the primer-annealing sites in all analyzed genomes. The Ct values were calculated by the second-derivative-maximum method. Relative quantification of the samples was calculated by fitting the Ct value to the standard curve of the vector. The copy number for the genomic DNA was calculated using the concentration and length (3508 bp) of the standard plasmid, together with the genomic size of 1.17×109 bp calculated by the reported C-value (1.2 pg) of the Lake Victoria species Haplochromis parvidens [37].
Accession Numbers
The GenBank (http://www.ncbi.nlm.nih.gov/Genbank) accession numbers for DNA sequences discussed in this paper are: AB601473–AB601502.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. KocherTD 2004 Adaptive evolution and explosive speciation: the cichlid fish model. Nat Rev Genet 5 288 298
2. TeraiYSeehausenOSasakiTTakahashiKMizoiriS 2006 Divergent selection on opsins drives incipient speciation in Lake Victoria cichlids. PLoS Biol 4 e433 doi:10.1371/journal.pbio.0040433
3. SeehausenOTeraiYMagalhaesISCarletonKLMrossoHD 2008 Speciation through sensory drive in cichlid fish. Nature 455 620 626
4. LandeRSeehausenOvan AlphenJJM 2001 Mechanisms of rapid sympatric speciation by sex reversal and sexual selection in cichlid fish. Genetica 112–113 435 443
5. RobertsRBSerJRKocherTD 2009 Sexual conflict resolved by invasion of a novel sex determiner in Lake Malawi cichlid fishes. Science 326 998 1001
6. SerJRRobertsRBKocherTD 2009 Multiple interacting loci control sex determination in Lake Malawi cichlid fish. Evolution 64 486 501
7. SeehausenOvan AlphenJJMLandeR 1999 Color polymorphism and sex ratio distortion in a cichlid fish as an incipient stage in sympatric speciation by sexual selection. Ecol Lett 2 367 378
8. PolettoABFerreiraIAMartinsC 2010 The B chromosomes of the African cichlid fish Haplochromis obliquidens harbour 18S rRNA gene copies. BMC Genet 11 1
9. CamachoJPM 2005 B Chromosomes. GregoryTR The Evolution of the Genome Amsterdam Elsevier Academic Rress 223 286
10. JonesRNReesH 1982 B chromosomes London Academic Press 266
11. OstergrenG 1945 Parasitic nature of extra fragment chromosomes. Bot Notiser 2 157 163
12. SeehausenOLippitschEBoutonNZwennesH 1998 Mbipi, the rock-dwelling cichlids of Lake Victoria: description of three new genera and fifteen new species (Teleostei). Ichthyol Explor Freshwaters 9 129 228
13. GreenDM 1988 Cytogenetics of the endemic New Zealand frog, Leiopelma hochstetteri: extraordinary supernumerary chromosome variation and a unique sex-chromosome system. Chromosoma 97 55 70
14. SharbelTFGreenDMHoubenA 1998 B-chromosome origin in the endemic New Zealand frog Leiopelma hochstetteri through sex chromosome devolution. Genome 41 14 22
15. CamachoJPMSharbelTFBeukeboomLW 2000 B-chromosome evolution. Philos Trans R Soc Lond B Biol Sci 355 163 178
16. HacksteinJHPHacksteinPHochstenbachRHauschteck-JungenEBeukeboomLW 1996 Is the Y chromosome of Drosophila an evolved supernumerary chromosome? Bio Essays 18 317 323
17. NokkalaSGrozevaSKuznetsovaVMaryanska-NadachowskaA 2003 The origin of the achiasmatic XY sex chromosome system in Cacopsylla peregrina (Frst.) (Psylloidea, Homoptera). Genetica 119 327 332
18. VicenteVEMoreira-FilhoOCamachoJP 1996 Sex-ratio distortion associated with the presence of a B chromosome in Astyanax scabripinnis (Teleostei, Characidae). Cytogenet Cell Genet 74 70 75
19. WerrenJHStouthamerR 2003 PSR (paternal sex ratio) chromosomes: the ultimate selfish genetic elements. Genetica 117 85 101
20. WatanabeMKobayashiNFujiyamaAOkadaN 2003 Construction of a BAC library for Haplochromis chilotes, a cichlid fish from Lake Victoria. Genes Genet Syst 78 103 105
21. LohYEKatzLSMimsMCKocherTDYiSV 2008 Comparative analysis reveals signatures of differentiation amid genomic polymorphism in Lake Malawi cichlids. Genome Biol 9 R113
22. MiaoVPCovertSFVanEttenHD 1991 A fungal gene for antibiotic resistance on a dispensable (“B”) chromosome. Science 254 1773 1776
23. GraphodatskyASKukekovaAVYudkinDVTrifonovVAVorobievaNV 2005 The proto-oncogene C-KIT maps to canid B-chromosomes. Chromosome Res 13 113 122
24. YudkinDVTrifonovVAKukekovaAVVorobievaNVRubtsovaNV 2007 Mapping of KIT adjacent sequences on canid autosomes and B chromosomes. Cytogenet Genome Res 116 100 103
25. TeruelMCabreroJPerfecttiFCamachoJP 2010 B chromosome ancestry revealed by histone genes in the migratory locust. Chromosoma 119 217 225
26. LeeBLeeWStreelmanTCarletonKLHoweAE 2005 A second-generation genetic linkage map of Tilapia (Oreochromis spp.). Genetics 170 237 244
27. van DoornGSKirkpatrickM 2007 Turnover of sex chromosomes induced by sexual conflict. Nature 449 909 912
28. KitanoJRossJAMoriSKumeMJonesFC 2009 A role for a neo-sex chromosome in stickleback speciation. Nature 461 1079 1083
29. van der SluijsIvan DoorenTJMSeehausenOvan AlphenJJM 2008 A test of fitness consequences of hybridization in sibling species of Lake Victoria cichlid fish. J Evolution Biol 21 480 491
30. MatsudaYChapmanVM 1995 Application of fluorescence in situ hybridization in genome analysis of the mouse. Electrophoresis 16 261 272
31. MatsudaYHaradaYNNatsuume-SakaiSLeeKShiomiT 1992 Location of the mouse complement factor H gene (cfh) by FISH analysis and replication R-banding. Cytogenet Cell Genet 61 282 285
32. LevanAFredgaKSandbergAA 1964 Nomenclature for centromeric position on chromosomes. Hereditas 52 201 220
33. TaguchiTAkimaruKHiraiHHiraiYMwendaJM 2003 A probe generated by chromosome microdissection, useful for analyzing Y chromosome evolution in Old World monkeys. Chromosome Res 11 147 152
34. TaguchiTHiraiYLoVerdePTTominagaAHiraiH 2007 DNA probes for identifying chromosomes 5, 6, and 7 of Schistosoma mansoni. J Parasitol 93 724 726
35. ShirakAGrabherrMPalmaFDLindblad-TohKHulataG 2010 Identification of repetitive elements in the genome of Oreochromis niloticus: tilapia repeat masker. Mar Biotechnol 12 121 125
36. TamuraKDudleyJNeiMKumarS 2007 MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol Biol Evol 24 1596 1599
37. HinegardnerRRosenDE 1972 Cellular DNA content and the evolution of teleostean fishes. Am Nat 106 621 644
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2011 Číslo 8
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- An EMT–Driven Alternative Splicing Program Occurs in Human Breast Cancer and Modulates Cellular Phenotype
- Chromosome Painting Reveals Asynaptic Full Alignment of Homologs and HIM-8–Dependent Remodeling of Chromosome Territories during Meiosis
- A Pre-mRNA–Associating Factor Links Endogenous siRNAs to Chromatin Regulation
- Discovery of Sexual Dimorphisms in Metabolic and Genetic Biomarkers
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy