
Evoluce a evoluční teorie pro lékaře
III. O vzniku života
Evolution and evolutionary theory for physicians.
III. The origin of life
Considerations about the origins of life should have some basis on the notion of what life is. There are three types of scientific definitions of life:
1. biological,
2. biophysical, and
3. attempts at a theory of general life systems.
Difficulties exist with all of them. A description of the characteristics of life is therefore perhaps better. An English acronym PICERAS
– program,
– improvisation,
– compartmentalization,
– energy,
– regeneration,
– adaptability,
– seclusion,
Koshland’s seven pillars of life, is a good working description. Abiogenesis, the emergence of life in nonliving environment is an open question. The rise of simple organic monomers, the building blocks of future life, is explained by a spectrum of theories from Oparin’s coacervate theory and the Miller-Urey experiment to speculations and experiments taking into account minerals, crystals, clays, zeolites and radioactive beaches to outer space. The journey from monomers to polymers to protocells unravels a number of theories which could be divided into two main groups: “gene first” and “metabolism first” and their hybrid forms, the Lipid world theory and Kauffman’s autocatalytic networks theory included.
Key words:
definition of the life, abiogenesis, monomers, polymers, protocells.
Autoři:
F. Koukolík
Působiště autorů:
Primář: MUDr. František Koukolík, DrSc.
; Národní referenční laboratoř prionových chorob
; Fakultní Thomayerova nemocnice s poliklinikou, Praha
; Oddělení patologie a molekulární medicíny
Vyšlo v časopise:
Prakt. Lék. 2010; 90(3): 135-140
Kategorie:
Editorial
Souhrn
Úvahy o vzniku života by se měly opírat o představu, co život je. Existují tři druhy vědeckých definic života:
1. biologické,
2. biofyzikální,
3. pokusy o obecnou teorii živých systémů.
Se všemi jsou obtíže. Proto je lepší popis vlastností života, které vystihuje anglický akronym PICERAS, Koshlandových sedm pilířů života, převedený do češtiny pojmy:
– program,
– improvizace,
– kompartmentalizace,
– energie,
– regenerace,
– adaptabilita,
– izolace.
Abiogeneze, vznik života v neživém prostředí, je otevřená otázka. Vznik jednoduchých organických monomerů, stavebních kamenů budoucího života vysvětluje spektrum teorií počínaje Oparinovou hypotézou, přes Millerův-Ureův experiment, po úvahy a experimenty beroucí v potaz minerály, krystaly, jíl, zeolity, radioaktivní pláž, až po kosmický prostor. Cestu od monomerů přes polymery k primitivním buňkám vysvětluje větší počet teorií ve dvou skupinách, které lze nazvat „první byl genetický kód“ a „první byl metabolismus“ a jejich smíšené formy, včetně teorie světa lipidů a Kaufmanovy teorie autokatalytických sítí.
Klíčová slova:
definice života, abiogeneze, monomery, polymery, primitivní buňky.
Úvod
Roku 1871 v dopisu Josephu Hookerovi Charles Darwin napsal: „Často se říká, že v současnosti existují všechny podmínky pro první vytvoření živého organismu, podmínky, které mohly existovat vždy. Jestliže (ouvej, a jak velké je ono „jestliže“) si však dokážeme představit nějaký malý teplý rybník se všemi druhy amonných a fosfátových solí a současně s přítomností světla, tepla a elektřiny, takže se v něm chemicky zformovala proteinová sloučenina schopná prodělat složitější změny, byla by v současnosti taková matérie okamžitě pozřena, nebo absorbována, což by se v době před vznikem živých bytostí nestalo.“
Uvedený odstavec je často citován
- jako Darwinova představa o abiogenezi, neboli vzniku života z neživota,
- jako jeho představa o mechanismu této změny,
- jako jeho názor, že ke vzniku života mohlo dojít jen jednou.
Dalších 140 let poznávání ukázalo, o jak složitou problematiku se jedná.
Evoluční teorie se zabývá vývojem života, nikoli jeho vznikem. Chceme-li se však zabývat vývojem života, neujdeme otázce, jak život vznikl. Pokusím se proto popsat některé současné vědecké odpovědi. Plyne z nich, že vznik života je otevřená, velmi složitá otázka, z čehož na druhé straně neplyne, že by šlo o otázku neřešitelnou. Následující velmi stručný a neúplný přehled, jenž se snaží pokud možno vyhnout jazyku chemie, který bývá lékařům méně srozumitelný, je nutné považovat za pouhý vstup do problematiky.
Definice života
Úvahy o evoluci a vzniku života by se měly opírat o představu co život je. Existují tři druhy vědeckých definic života:
- biologické, které budou lékařům pravděpodobně nejbližší,
- biofyzikální, obvykle vyžadující osvěžení některých základních školních fyzikálních fakt,
- pokusy o obecnou teorii živých systémů, jejichž nároky na znalosti a představivost bývají pro lékaře nejvyšší snad pro jejich obecnost a určitou znalost matematiky. Důsledkem je, že mnohým zatím připadají sice zajímavé, nicméně vágní. Nadto jedna z nich, Huttonova (8) a Lovelockova (17, 18) hypotéza Gaia, tvrdící, že je naše planeta jediný superorganismus ovlivňující prostředí ve jménu vlastního přežití, má postmoderně mystický přídech.
Biologické definice života
Výhodou stručnosti je snadná zapamatovatelnost, nevýhodou může být nejasnost užitých pojmů. Stručné biologické definice života říkají:
- Život je samostatný chemický systém schopný prodělávat darwinovskou evoluci, což je definice užívaná americkou vesmírnou agenturou NASA. Doslovná autorská definice zní: „Život je to, co se vyvíjí v darwinovském smyslu“ (10).
- Život je síť nižších záporných zpětných vazeb, neboli regulačních mechanismů, které jsou podřízeny vyšším, pozitivním zpětným vazbám, ty jsou schopné expanze a reprodukce (13).
- Živé bytosti jsou systémy odpovídající na změny ve své prostředí a ve svém nitru tak, že napomáhají vlastnímu pokračování (38).
Ke každé z nich lze najít nepřehledný počet věcných i méně věcných polemických komentářů. Pokusy o dosažení obecně přijaté definice života v současnosti připomínají snahu o nalezení El Dorada.
Lepší než definice je proto popis vlastností života.
Život charakterizuje
- homeostáza (přesnější pojem by mohl být homeodynamika, protože nejde o statický jev),
- organizace,
- metabolismus,
- růst,
- adaptace,
- odpověď na podněty, a
- reprodukce.
Výstižný anglický akronym vlastností života je PICERAS.
- Program,
- Improvisation,
- Compartmentalization,
- Energy,
- Regeneration,
- Adaptability,
- Seclusion (překládám pojmem izolace),
jehož autor, Daniel Koshland Jr. (14) mluví o sedmi pilířích života. Pojmem pilíř má na mysli termodynamické (P1) a kinetické principy, které jsou podkladem stavby a funkce živých bytostí.
Program
je organizovaný plán popisující jak složky živých systémů, tak jejich interakce. Nositelem programu většiny podob života na Zemi je DNA replikující se s případnými malými změnami z jedné generace do generace další.
Improvizace
umožňuje adaptaci na proměny na proměny prostředí. Příkladem mechanismu improvizace je mutace a selekce.
Kompartmentalizace,
doslovně rozčlenění na úseky, popisuje oddělení organismů od prostředí a rozdělení součástí organismů do menších částí, orgánů, tkání, buněk a buněčných organel. Lidské tělo tvoří 350 různých druhů buněk obvykle obsahujících jádro s jadérkem, mitochondrie, ribosomy, lysosomy, Golgiho aparát…Rozdělení živých systémů do menších oddílů umožňuje centralizaci a specializaci jednotlivých funkcí včetně reakční kinetiky substrátů a enzymů.
Energie
je čtvrtým pilířem. Organismy jsou termodynamicky otevřené systémy. Jejich chod vyžaduje společně s látkou a informací dodávku energie. Dodávka energie kompenzuje entropii. Hlavním, nikoli jediným energetickým zdrojem je Slunce.
Regenerace
kompenzuje daň za život – život není věc, ale proces – jímž je trvalé opotřebovávání buněk a jejich složek.
Adaptabilita
je molekulárně viděno soubor zpětnovazebných i dopředných (feedforward) reakcí umožňujících rychlou odpověď na proměny zevního prostředí. Příkladem adaptability může být reflexní stažení ruky, která se dotkne horkého objektu nebo zesílení a tvrdnutí kůže na pracující dlani. Improvizace, která je podobou adaptability, umožňuje odpovědi pomalé.
Izolace
Koshland přirovnává význam tohoto pojmu k izolaci elektrických drátů zabraňující krátkému spojení, znemožňuje, aby členy jednoho metabolického řetězu ovlivňovaly katalyzátory jiného metabolického řetězu.
* * *
Biofyzikálních definic života, vymezujících život jako otevřený systém schopný snižovat vnitřní entropii na vrub okolí (33) a definic plynoucích ze vznikající obecné teorie živých systémů, se dotknu v závěrečné stati nazvané Darwinovská revoluce.
Spontánní vznik života
Do roku 1859 panovalo předsvědčení o spontánním vzniku života, například myší ze zapařeného sena. Francesco Redi roku 1668 dokázal, že se muší larvy z masa nelíhnou, zabrání-li se mouchám klást vajíčka. Lazzaro Spallanzani r. 1778 prokázal, že množení mikroorganismů lze zabránit převařením nálevu. Počínaje rokem 1859 doložil Louis Pasteur geniálními, jednoduchými a elegantními experimenty v řadě variant, že mikroorganismy ve sterilním prostředí spontánně nevznikají. Pasteur, chemik a mikrobiolog, užil nádobu obsahující bujón. Nádoba měla úzké, dlouhé, prohnuté hrdlo podobné labutímu krku. Jestliže byl bujón převařený a díky tvaru hrdla nepřišel do styku se vzduchem, nezkalil se (obr. 1).

Nejstarší stopy života
Stáří Země bylo radiometricky určeno na přibližně 4,6 miliardy let. První oceány se objevily po 200 milionech let. Povrchová teplota byla kolem 100 °C. V hadejském eonu pojmenovaném podle starořeckého boha podsvětí, 4,1–3,8 miliardy let před současností, život na Zemi nebyl.
Přepokládá se, že nejstarší nespornou podobou života jsou stromatolity, v současnosti polokulovité útvary tvořené mineralizovanými mikroorganismy (obr. 2), nejstarším je kolem 3 miliard let.

Stromatolitům musely předcházet primitivnější vývojové podoby. Stáří jejich sporných podob připomínajících primitivní verzi dnešních sinic se odhaduje kolem 3,5–3,8 miliardy let (obr. 3) (32).
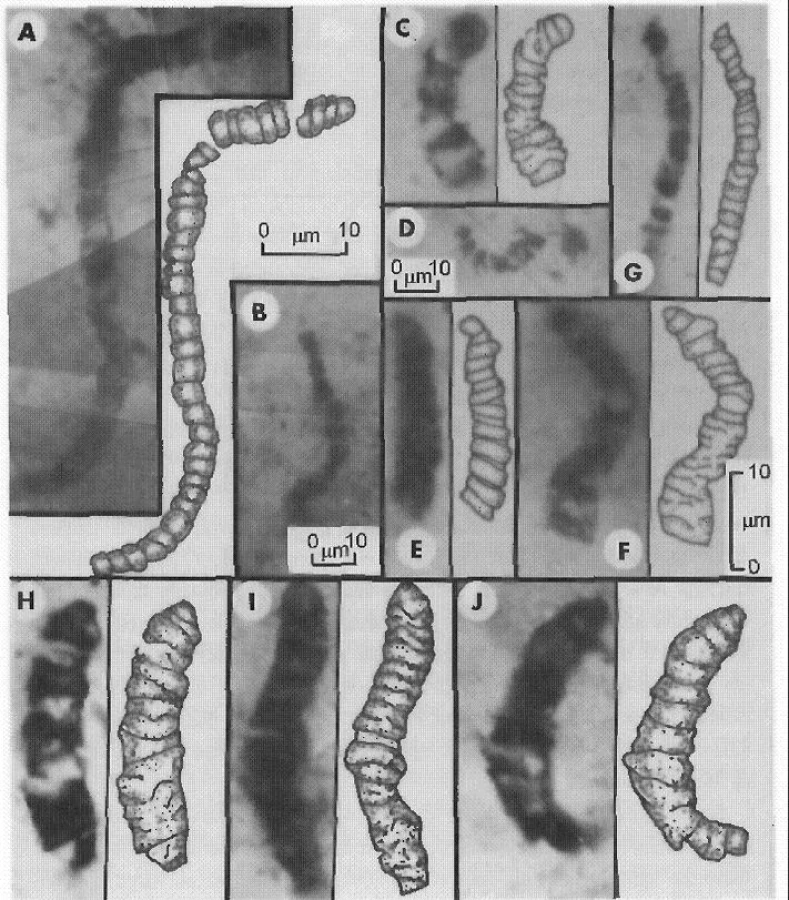
Na nejstarší podoby života lze soudit i z přítomnosti fosilních izotopů uhlíku. Izotop C12 se akumuluje v organismech: poměr C12/C13 je v rostlinách vyšší než v okolí, v býložravcích vyšší než v rostlinách, v masožravcích vyšší než v býložravcích. Nejstarší stopa ukazující z tohoto směru výzkumu na možnost existence života pochází z ostrova v blízkosti Grónska, stáří je 3,85 miliardy let (30).
Vznik jednoduchých organických molekul
Oparin (1924/1952; 24) a nezávisle na něm Haldane (1929; 7) došli k názoru, že život vznikl z neživé hmoty v praoceánu, v „prvotní polévce“ organických molekul. Předpokládali, že tehdejší atmosféra byla redukující, velmi chudá na kyslík, zdrojem energie bylo Slunce. Oparinovu-Haldaneovu hypotézu testoval Millerův-Ureyův experiment publikovaný na dvou stránkách časopisu Science r. 1953. Experiment napodobil praoceán a užil předpokládanou redukující atmosféru Země tvořenou vodou, metanem, čpavkem a vodíkem. Atmosférou probíhaly elektrické výboje simulující blesky. Po týdnu byly v systému nalezeny organické sloučeniny včetně aminokyselin (23).
Experiment měl předchůdce i následovníky, byl proveden v mnoha různých podobách. Dvě z nich Miller nepublikoval. Analýza dochovaného obsahu nádob z nepublikované druhé varianty experimentu, doložila 22 aminokyselin místo původně nalezených pěti (obr. 4) (9).
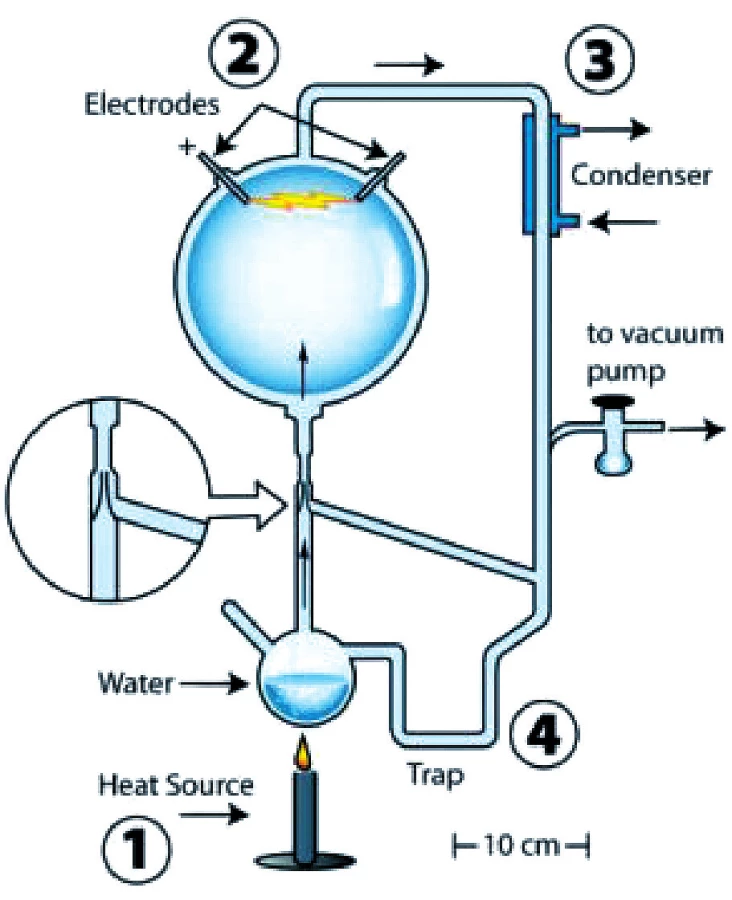
V průběhu těchto experimentů byly vytvořeny kromě látek, jež dnešní život neužívá, všechny monomery, které užívá, s výjimkou ribózy, adeninu a guaninu. Monomery jsou malé molekuly, příkladem je glukóza, které jsou s to spojovat se s dalšími monomery a vytvořit tak delší molekulu, polymer, příkladem je škrob a celulóza.
Výsledky Millerova-Ureyova experimentu doložily možnost vzniku monomerů z jednoduchých sloučenin. Výsledky jsou však problematické:
- Je otázkou, zda bylo složení atmosféry takové, jaké experiment užil. Současný výzkum má za to, že byla méně reaktivní, tvořil ji převážně dusík a oxid uhličitý.
- Existence „prvotní polévky“ nebyla doložena.
- Polymery, například RNA a proteiny, které jsou základem života, při experimentu nevznikly.
- V podmínkách Millerova-Ureyova experimentu se polymery rozpadají.
Další studie doložily vznik některých klíčových monomerů střídáním mrazu a tání. Ve směsi amonného iontu a kyanidu ponechané v lednici v rozmezí let 1972–1997 vzniklo sedm druhů aminokyselin a jedenáct druhů bazí nukleových kyselin. Jestliže směs obsahující prekurzory monomerů klíčových pro vznik života zmrzne, reakce proběhne i v ředěném roztoku, což bylo doloženo pro vznik purinů jakož i pyrimidinů cytosinu a uracilu. Chladné prostředí umožňuje vznik všech bází RNA i v prostředí, kde je koncentrace prekurzorů nízká. Důkazem by byl jejich nález na Marsu, případně na Titanu a Evropě (4).
Minerály, krystaly, jíl, zeolity, kosmický prostor a radioaktivní pláž
Existuje značný počet teoretických možností vzniku biogenních monomerů. Jedním ze studovaných témat je vztah jejich vzniku a minerálních látek. Skaliska mohla poskytovat jezírkům vody, která vznikala na pobřeží příbojem, stínění před UV zářením a odparem. Mnoho minerálních formací má na povrchu trhliny a póry. Obojí zvyšuje pravděpodobnost kondenzace vznikajících monomerů a vzniku polymerů samoorganizací.
Prostředím umožňujícím vznik složitějších molekul mohly být některé jíly a zeolity, to jsou mikroporézní aluminosilikátové minerály adsorbující velká množství vody (34), případně krystaly (16, 2). Na vzniku klíčových monomerů života se mohly podílet uhlovodíky z hloubi zemského pláště (6). Překvapivě působil objev většího počtu různých druhů organických látek v kosmickém prostoru, například v některých mlhovinách, takže zdroj některých základních stavebních kamenů života včetně polycyklických aromatických uhlovodíků může být i zde (15, 28). Adam (1) se domnívá že gravitace podmíněná blízkostí Měsíce v raném vývojovém období Země mohla koncentrovat těžší radioaktivní prvky, například uran, v čáře přílivu. Radioaktivita mohla být energetickým zdrojem pro tvorbu organických molekul z acetonitrilu ve vodě. Modelování říká, že touto cestou mohly simultánně vzniknout aminokyseliny, cukry i fosfáty.
Chiralita
Základní stavební kameny života charakterizuje chiralita: nukleové kyseliny jsou pravotočivé, aminokyseliny jsou levotočivé.
Levotočivá forma alaninu zjištěná v Murchinsonově meteoritu je dvojnásobně častější než forma pravotočivá, levotočivá forma glutamátu je častější než forma pravotočivá trojnásobně. Možné je, že cirkulárně polarizované světlo tvořící 17 % hvězdného záření v protoplanetárním disku ničilo pravotočivý enantiomer častěji než levotočivý. Řada autorit má za to, že uvedená chiralita vznikla zprvu náhodně, jakmile se však ustavila, byla nadále selektována (21, 22).
Tvorba polymerů z monomerů a vznik autoreplikace
Jestli existuje teoreticky velký počet možností vzniku monomerů, které jsou stavebními kameny života, pak je složitější otázkou vznik polymerů, buněčných membrán, genetického kódu, proteinů a metabolických řetězů.
Za opuštěnou představu se považují Foxovy experimenty z počátku druhé poloviny minulého století, které prokazovaly, že aminokyseliny mohou spontánně tvořit malé peptidy, jež společně s aminokyselinami vytvářejí sférické membrány pojmenované proteinoidní mikrosféry s vlastnostmi připomínajícími vlastnosti života.
Schusterova - Eigenova hypotéza (1979; 31) hypercyklického přechodu od molekulárního chaosu prebiotické polévky do autoreplikace byla rovněž opuštěna. V průběhu hypercyklu měl systém ukládající informace katalyzovat vznik dalšího informačního systému až do vzniku „druhu“ jenž zahájí darwinovskou evoluci. Je pravda, že RNA může za jistých okolností katalyzovat vlastní chemické reakce, nicméně tyto reakce její řetěz obvykle zkracují. Prodlužování je vzácné, vzniklé úseky nejsou s to kódovat vznik proteinu. Ke vzniku RNA jsou kromě toho nutné nukleotidy, které v podmínkách Millerova-Ureyova pokusu nevznikají.
Extremofily
Odhaduje se, že přibližně polovinu biomasy Země mohou tvořit podzemní mikroorganismy, zejména Archea, ty mohou být stejně staré, případně starší než bakterie a žijí až do hloubky 5 km. Mikrobiální společenství byla zjištěna v čedičových a dalších minerálních formacích pod mořským dnem (35). Tyto podoby života existují ve vysokém tlaku a teplotě, bez slunečního světla. Jejich metabolismus je pomalý, formy žijící v horninách a ledu mohou být inaktivní tisíce let.
Úvahy o vzniku života rozšířil objev nečekaných podob života v blízkosti podmořských sopečných průduchů (angl. vent), někdy zvaných podle vzhledu „černé kuřáky“. Tlak v okolí podmořských průduchů je 500–2 000 atmosfér, teplota 200–300 °C. Předpokládá se, že mořské dno mohlo představovat v průběhu hadejského eonu a bombardování Země asteroidy lepší ochranu vznikajícího života než zemský povrch.
Zajímavá teorie chemoautotrofního vzniku života ve vulkanickém světě železa a síry předpokládá vznik nejranějších podob života v tomto prostředí (36). Hypotetický pionýrský organismus autora této teorie byl tvořen anorganickou substrukturou a organickou superstrukturou. Anorganickou substrukturu tvořily sloučeniny železa, kobaltu, niklu. Měly katalytické karbonylové, sulfidické a další ligandy. Organická superstruktura vznikala fixací uhlíku. Energetickým zdrojem byl redukční potenciál vulkanických exhalací. Metabolismus pionýrských podob života se reprodukoval autokatalytickou zpětnou vazbou. Následoval vznik genetické mašinérie a buněk. Raná fáze evoluce končila vznikem tří hlavních domén života:
- Bacteria,
- Archea, a
- Eukarya.
Ty se vyvinuly z rychle se vyvíjející populace primitivních buněk. Klíčové je, že podmořské sopečné průduchy jsou zdrojem řady prvků a organických sloučenin, které se dostávají do vody. Mohou být podporou života nezávislého na fotosyntéze a být kolébkou symbiotického života, jehož součástí jsou lithoautotrofní mikroorganismy užívající chemickou energii pro podporu mnohobuněčných podob života. Je možné, že současný mikrobiální život kolem sopečných průduchů má fyziologické charakteristiky podobající se nejranějším mikrobiálním formám života na Zemi (20).
Existují dobré důvody pro předpoklad, že v tomto prostředí se vyvinuly první metabolické dráhy vedoucí k syntéze purinů a pyrimidinů, základních stavebních kamenů nukleových kyselin (19).
Od organických molekul k primitivním buňkám
Domněnky vysvětlující cestu od organických molekul k primitivním buňkám zčasti připomínají úvahy o slepici a vejci: objevily se jako první nukleové kyseliny a s nimi genetický kód, a/nebo se jako první objevily složité seřetězené metabolické dráhy?
„Ribosvětem“, RNA světem, bývá nazývána představa života založeného na RNA, jenž předcházel životu založenému na DNA a proteinech. O možnosti existence ribosvěta (11) uvažoval v roce 1963 Rich a roku 1968 Woese, přičítá se však Gilbertovi (5).
V RNA světě, který mohl mít větší počet podob, byla nositelkou všech klíčových funkcí života – kalatýzy, dědičnosti, rekombinace a evoluce – RNA spolu se svými kofaktory. Ale i tak velký zastánce ribosvěta jakým byl Orgel, (25) si plně uvědomova úskalí této hypotézy:
„Jestliže svět RNA předcházel původ syntézy proteinů, pak lze o chemických procesech, které byly podkladem vzniku života, zjistit ze studia proteinových enzymových mechanismů jen málo…jestliže RNA svět vznikl na primitivní Zemi de novo, pak vztyčil mezi biochemii a prebiotickou chemii téměř neprostupnou překážku.“
Experimentálně se sice podařilo doložit polymerizaci ribonukleotidů do RNA, je však nejasné jak vznikaly ribonukleotidy. Selektivní syntéza ribózy je obtížná. Powner et al. (27) však zjistili, že aktivované pyrimidinové ribonukleotidy mohou vytvářet krátké řetězy odlišnou chemickou cestou. Výchozím materiálem pro syntézu jsou kyanoacetylen, glykoaldehyd, glyceraldehyd a anorganický fosfát, což jsou látky, které se v prebiotickém stavu vyskytovat mohly.
Současná transkripce je „přepis“ genetické informace z DNA do RNA. Současná translace je „překlad“ genetické informace z RNA do pořadí aminokyselin.
Pravděpodobně nejtěžším problém všech teorií vzniku života je původ translace. Vysokou mírou translační přesnosti nelze docílit bez složitých podob RNA a proteinů, přitom proteiny nelze vytvořit bez přesného translačního systému. S tím souvisí otázka vzniku genetického kódu. Wolf a Koonin (37) mají za to, že původ translačního systému lze hledat v oddělených souborech segmentů RNA, které se replikovaly, jinými slovy řečeno v „ribosvětě“ obsahující ribozymy. Evoluce nepředvídá, takže translační systém se nemohl v RNA světě vyvinout jako výsledek selekce proteinové syntézy. Podle Wolfa a Koonina (37) spíše jde o vedlejší evoluční produkt, funkci původně selektovanou k jinému účelu.
Translační systém je tedy exaptací – jinak řečeno funkcí původně určenou jiné činností, podobně jako je tomu se čtením a psaním, k nimž využíváme funkční systémy mozku určené rovněž zcela odlišným funkcím. Evoluční proces jehož výsledkem je translace začala selekcí ve směru ribozymů vážících biogenní aminokyseliny:
Prvním krokem byla vazba aminokyselin na ribozym, což zvýšilo jeho katalytickou aktivitu. Takto stimulovaný ribozym se vyvinul do peptidové ligázy, která je předchůdcem větší ribozomální podjednotky. Výsledkem byl vznik zprvu unikátního peptidu, jenž aktivoval původní ribozym a snad i další ribozymy v souboru.
Dalším krokem byl vznik primitivních t-RNA, původně selektovaných pro akumulaci aminokyselin, následně pro nabídku aminokyselin peptidové lipáze.
V následném kroku dokázala peptidová ligáza tvořit soubory peptidů při užití exogenní RNA. Ta byla templátem pro komplementární vazbu primitivních proteoribozomů. Výsledkem byl vznik stále delších peptidů – prvotních bílkovin. Tím byl vývoj translace dokončen.
Modely vzniku života, které mají za to, že vznik metabolismu předcházel vzniku „nahých genů“, lze datovat od Oparinovy koacervátové teorie, přes výše zmíněnou Wächterhäuserovu teorii síra-železo k modelům Christiana de Duveho, založeným na thiosterech.
Značným problémem abiogeneze je termodynamická rovnováha mezi aminokyselinami a peptidy – směřuje k aminokyselinám. Řešením by mohly být „reaktivní“ polyfosfáty vznikající polymerizací monofosfátových iontů. Polyfosfáty mohly odpovídat za polymerizaci aminokyselin do bílkovin (3). Zdrojem „reaktivních“ polyfosátů mohly být meteority (26).
Autokatalyzátory jsou látky katalyzující tvorbu sebe samotných. Tím mají vlastnosti jednoduchých molekulárních replikátorů. Vznik života coby soustavy autokatalytických chemických sítí razí v pozoruhodné a diskutované teorii Kauffman, jehož poměrně přístupná publikace vyšla i v českém překladu (12).
Teorie lipidového světa má na mysli složení buněčných membrán a předpokládá, že se první replikátory podobaly lipidům. Lipidová tělíska měla umožňovat kompartmentalizaci – rozdělení ukládané informace a její oddělení od prostředí (29).

MUDr. František Koukolík, DrSc.
Oddělení
patologie a molekulární
medicíny
Národní
referenční laboratoř prionových
chorob
Fakultní Thomayerova nemocnice s poliklinikou
Vídeňská
800, 140
59 Praha 4 Krč
E-mail:
frantisek.koukolik@ftn.cz
Zdroje
1. Adam, Z. Actinides and life’s origin. Astrobiology 2007, 7, p. 852-872.
2. Arrhenius, G.O. Crystals and life. Helvetia Chimia Acta 2003, 86, p. 1569-1586.
3. Brown, M.R., Kornberg, A. Inorganic polyphosphate in the origin and survival of species. Proc. Natl. Acad. Sci. USA 2004, 101(46), p. 16085-16087.
4. Cleaves, H.J., Nelson, K.E., Miller, S.L. The prebiotic synthesis of pyrimidines in frozen solution. Naturwissenchaften 2006, 93, p. 228-231.
5. Gilbert, W. The RNA world. Nature 1986, 319, p. 319.
6. Gold, T. The deep, hot biosphere. Proc. Natl. Acad. Sci. USA 1992, 89, p. 6045–6049.
7. Haldane, J.B.S The origin of life. The Rationalist Annual for the Year 1929, p. 3-10.
8. Hutton, J. 1785 [on-line]. Dostupné na http://en.wikipedia.org/wiki/James_Hutton.
9. Johnson, A.P., Cleaves, H.J., Dworkin, J.P. et al. The Miller volcanic spark discharge experiment. Science 2008, 322, p. 404.
10. Joyce, G.F. The RNA world: The life before DNA and protein [on-line]. Dostupné na http://ntrs.nasa.gov/archive/nasa/casi.ntrs.nasa.gov/19980211165_1998078054.pdf.
11. Joyce, G.F. Recreating the RNA world in the laboratory [on-line]. Dostupné na http://online. itp.ucsb.edu/online/evonet07/joyce/.
12. Kauffman, S. Čtvrtý zákon. Cesty k obecné biologii. Praha: Paseka, 2004.
13. Korzienewski, B. Cybernetic foundations of the definition of life. J. Theor. Biol. 2001, 209, p. 275-286.
14. Koshland. D.E. Jr. The seven pillars of life. Science 2002, 295, p. 2215-2216.
15. Lazcano, A., Oró, J., Miller, S.J. Primitive earth environments: organic syntheses and the origin and early evolution of life. Precambrian Res. 1983, 20, p. 259-282.
16. Lepland, A., van Zuilen, M., Arrhenius, G. et al. Questioning the evidence for Earth’s earliest life - Akilia revisited. Geology 2005, 33, p. 77-79.
17. Lovelock, J.E. A physical basis for life detection experiments. Nature 1965, 207, p. 568-570.
18. Lovelock, J.E. Gaia – a new look at life on earth. Oxford: Oxford University Press, 1979.
19. Martin, W., Rusell, M.J. On the origin of biochemistry at an alkaline hydrothermal vent. Phil. Trans. R. Soc. B. 2007, 362, p. 1887-1925.
20. Martin, W., Baross, J., Kelley, D. et al. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, p. 805-814.
21. Meierhenrich, U.W., Munoz Caro, G.M., Bredehoeft, J.H. et al. Identification of diamino acids in the Murchison meteorite. Proc. Natl. Acad. Sci. USA,2004, 101, p. 9281-9286.
22. Meierhenrich, U.W. Amino acids and the asymmetry of life. Heidelberg, Berlin, New York: Springer, 2008.
23. Miller, S.L. A production of amino acids under possible primitive Earth conditions. Science 1953, 117, p. 529-530.
24. Oparin, A.I. The origin of life. New York: Dover Publications, 1952.
25. Orgel, L.E. Prebiotic chemistry and the origin of the RNA world. Crit. Rev. Biochem. Mol. Biol. 2004, 39, p. 99-123.
26. Pasek, M.A. Rethinking early Earth phoshorus geochemistry. Proc. Natl. Acad. Sci. USA 2008, 105, p. 853-858.
27. Powner. M.W., Gerland, B., Sutherland, J.D. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 2009, 459, p. 239-242.
28. Rhee, Y.M., Lee, T.J., Gudipatti, M.S., et al. Charged polycyclic aromatic hydrocarbon clusters and the galactic extended red emission. Proc. Natl. Acad. Sci. USA 2007, 104, p. 5274-5278.
29. Segré. D., Ben-Eli, D., Dealer, D.W. et al. The lipid world. Orig. Life. Evol. Biosph. 2001, 31, p. 119-145.
30. Schidlowski, M. Carbon isotopes as biogeochemical recorders of life over 3.8 Ga of Earth history: evolution of a concept. Precambrian Res. 2001, 106, p. 117-134.
31. Schuster, P., Eigen, M. The hypercycle: a principle of natural self-organization. Berlin: Springer Verlag, 1979.
32. Schopf, W. Fossil evidence of archean life. Phil. Trans. R. Soc. B. 2006, 361, p. 869-885. Dostupné na http://physwww.mcmaster.ca/~higgsp/3D03/SchopfArchaeanLife.pdf.
33. Schroedinger, E. What is life? The Physical Aspect of the Living Cell. Based on lectures delivered under the auspices of the Dublin Institute for Advanced Studies at Trinity College, Dublin, in February 1943. [on-line]. Dostupné na http://whatislife.stanford.edu/Homepage/LoCo_files/What-is-Life.pdf.
34. Smith, J.V. Atmospheric weathering and silica-coated feldspar: Analogy with zeolite molecular sieves, granite weathering, soil formation, ornamental slabs, and ceramics. Proc. Natl. Acad. Sci. USA 1998, 95, p. 3366-3369.
35. Stevensa, T. Lithoautotrophy in the subsurface. FEMS Microbiol. Rev. 2006, 20, p. 327-337.
36. Wächtershäuser, G. From volcanic origins of chemoautotrophic life to bacteria, archaea and eukarya. Phil. Tran. R. Soc. B. 2006, 361, p. 1787-1808.
37. Wolf, Y.I., Koonin, E.V. On the origin of the translation system end the genetic code in the RNA world by means of natural selection, exaptation and subfunctionalization. Biol Direct 2007; 2 : 14. Published online 2007 May 31. doi: 10.1186/1745-6150-2-14.
38. Witzany, G. The Logos of the bios. 2. Bio-communication. Helsinki: Umweb, 2007.
Štítky
Praktické lékařství pro děti a dorost Praktické lékařství pro dospěléČlánek vyšel v časopise
Praktický lékař

2010 Číslo 3
- Efektivita kartáčku Sonicare For Kids u dětí předškolního věku
- Naděje budí časná diagnostika Parkinsonovy choroby založená na pachu kůže
- Pacient se bude cítit komfortně pouze při správně zvolené absorpční pomůcce
- Na výběru inkontinenčních pomůcek záleží − ale jak se mezi nimi neztratit?
Nejčtenější v tomto čísle
- Nozokomiální nákazy
- Komorbidita migrény a epilepsie
- Bazocelulárny karcinóm kože – najvýznamnejšie rizikové a prognostické parametre ochorenia v klinickej praxi
- Aseptická meningitida jako projev primoinfekce HIV
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy