
Transgenerational Inheritance of Diet-Induced Genome Rearrangements in Drosophila
We show that altering the nutritional medium of Drosophila cultures, emulating dietary largess in the wild, increases expression of the high copy-number ribosomal RNA genes and results in rDNA instability and loss. The reduction in gene copy number occurs both in somatic and germ cells, such that altered copy numbers are transmitted to the next generation. Our findings have clear ecological and disease relevance. The reduction of supernumerary ribosomal RNA gene copy number has been previously shown to compromise both epigenetic and genomic stability and alter the expression of hundreds of genes, while rDNA instability itself is a key progression hallmark of some cancers. This study links diet and Insulin/Insulin-like Signaling modulation to changes in the genome and provides an accounting for natural copy number variation of rRNA genes.
Published in the journal:
. PLoS Genet 11(4): e32767. doi:10.1371/journal.pgen.1005148
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1005148
Summary
We show that altering the nutritional medium of Drosophila cultures, emulating dietary largess in the wild, increases expression of the high copy-number ribosomal RNA genes and results in rDNA instability and loss. The reduction in gene copy number occurs both in somatic and germ cells, such that altered copy numbers are transmitted to the next generation. Our findings have clear ecological and disease relevance. The reduction of supernumerary ribosomal RNA gene copy number has been previously shown to compromise both epigenetic and genomic stability and alter the expression of hundreds of genes, while rDNA instability itself is a key progression hallmark of some cancers. This study links diet and Insulin/Insulin-like Signaling modulation to changes in the genome and provides an accounting for natural copy number variation of rRNA genes.
Introduction
It is clear that an organism’s gene expression patterns are responsive to environmental input. Often, this influence is not limited to short-term regulatory changes, but can persist through multiple cell divisions and can, in some cases, be transmitted to offspring. Typically, such changes are identified as “epigenetic” and are thought to be mediated by a variety of chromatin modifications [1–8]. However, because genome stability, particularly of highly-repetitive (e.g., pentameric repeat) sequences or middle-repetitive transposable elements, is modified by silencing involving repressive histone modifications, “epigenetic” perturbations may have both direct and long-term consequences: the former caused by disruption of silencing leading to “epigenetic instability,” and the latter by creating transmissible changes to chromosomes that themselves may affect gene regulation in subsequent generations. This consideration significantly adds to models of epigenetic inheritance that often overlook the ease with which histones and DNA methylation can be modified and the rapid rate at which they are turned over in non-dividing cells [9, 10]. Recent [11–14] and previous [15] findings show epigenetic silencing is unstable even in non-dividing cells, making it a particularly difficult challenge to reconcile models of chromatin (e.g., histone) mediated epigenetic silencing with transgenerational (i.e., mitotic and/or meiotic) inheritance.
35S Ribosomal RNA gene (rDNA) transcription has been a powerful model for understanding the regulatory effects of chromatin modification because evidence suggests identical genes may adopt different stable activity states (expressed versus repressed) even when immediately juxtaposed [16–19]. Transcription from tandem repeated rDNA arrays accounts for approximately 50% of total transcription [20] and is regulated such that only a subset of the redundant copies are active in a given cell type, while the remainder are inactive and are accompanied by chromatin structure consistent with silencing [21, 22]. Consequences of misregulation are severe, in part due to the tandem repeat of identical sequence. Mutations in silencing factors (e.g., gene products of the dcr-2, Su(var)3-9, HP1/Su(var)205, PARP, CTCF, and sir2/sirt loci) result in supernumerary mini - or micro-nucleoli and rDNA copy number reduction [23–27], possibly through increased frequency of intrachromosomal recombination resulting from the repair of transcription-induced damage [28, 29]. The tendency for natural loss and the ability of some rDNA arrays to expand through unknown processes [30–32] contribute to striking variation in rDNA copy number in both wild and laboratory strains [33–36]. This variation, in turn, is a potent genetic modifier of a number of phenomena, including the regulation of ecologically - and metabolically-relevant gene networks, the stability of genome structure and heterochromatin silencing, stress responses, and potentially metabolic function [24, 26, 37–46].
The relationship between rDNA transcriptional activity and rDNA array stability suggests a non-epigenetic mechanism through which the environment might induce heritable and consequential changes in the genome through long-term (i.e., permanent) modulation of genetic variation and epigenetic stability. Although the change may bear the hallmarks of epigenetic regulation (i.e., inducible, heritable, consequential), it may not technically be considered epigenetic because it involves chromosome changes [47]. Nonetheless, because rDNA copy number modulates the stability of epigenetic silencing [24, 26, 43], the origin of rDNA copy number variation is a significant concern to studies of “hidden” regulatory variation, heterochromatin, and epigenetics.
The aim of this study was to identify a natural, ecologically-common, and human health-relevant source of rDNA copy number variation. Expanding on previous work suggesting that an increase in rDNA expression may lead to its loss, we hypothesized that rDNA copy number changes can be induced by modulating diet. In support of our hypothesis, we found that flies raised on high dietary yeast concentration had increased supernumerary nucleoli as larvae, and somatic rDNA copy-number reductions as adults. Similar results were observed in flies expressing a hypermorphic insulin receptor allele, and were reproduced pharmacologically using human insulin in vitro, suggesting that diet-induced rDNA instability is mediated by known nutrient signaling pathways. Drugs that inhibit expression of the rDNA mitigated instability and loss, suggesting the effects were a consequence of expression. Furthermore, adult males fed high-yeast diets produced offspring with fewer Y-linked rDNA copies demonstrating that the effect was transgenerationally heritable. These findings identify diet as a potential source for rDNA variation observed in natural populations and suggest a mechanism through which environmental conditions might result in induced “transgenerational” genome changes.
Results
Previous work from yeast and filamentous fungi, Drosophila, plants, and experimental mammal systems have suggested that rDNA expression increases in response to diet, while other work has shown that derepression of the rDNA results in instability and loss. Together, these imply that natural rDNA variation may be reflective of the nutritional history of a population. Because of the growing awareness of the importance of rDNA instability and copy number variation in phenotypic variation, stress response, and disease, we directly tested our hypothesis that rDNA copy number may be manipulated by affecting diet.
Dietary manipulation
Dietary composition is an easily modified environmental variable in Drosophila laboratory studies and has been shown to influence a variety of complex phenotypes including rRNA transcription and ribosome biogenesis [48–51]; we therefore considered it as a potential source of “natural” rDNA instability and variability. In this study we used two experimental media based on those used in dietary restriction studies [52], consisting of a constant carbon source (5% sucrose) and altered concentrations of nutritional yeast (10%, and 30% w/v). Throughout the study we refer to these as SY10 and SY30 respectively (Sugar-Yeast-w/v). Larvae raised on SY30 developed to pupation approximately 9 hours earlier than their SY10 raised counterparts. Apart from this observation, there were no obvious differences in terms of body mass or survival to adulthood between the two media. To obviate secondary effects on culture conditions based on crowding, and to assure similarity in conditions as much as possible, we controlled density of eggs, larvae, and adults in vials by collecting eggs from petri dishes containing agar made from apple juice, suspending eggs in PBS, and pipetting identical volumes of eggs to SY10, SY30, or standard cornmeal medium.
Dietary manipulation alters rDNA expression and rDNA stability
We fist confirmed that altered diet affected rRNA expression. We could discriminate accumulated mature rRNA products (18S, 28S, 5.8S), for instance bound in ribosomes, from actively-transcribed pre-rRNAs (35S, or 45S in some organisms) by detecting the quantity of cDNA derived from the pre-processed 5’-most sequence of the 35S primary transcript containing the External Transcribed Spacer (ETS). The ETS is constitutively processed during maturation of the pre-rRNA 35S transcript and quickly degraded, and is therefore used to measure de novo rDNA expression [26, 53, 54]. We compared male flies of genotype yellow1 white67c23/Dp(1;Y) y+, P{w = RS5}10B (henceforth Y,10B), upon which we have performed other studies of the ribosomal rDNA [24, 41, 55–58]. We detected an approximately 50% increase in pre-rRNA levels in populations of second instar larvae raised on SY30 compared to Standard media (Fig 1A), confirming the suitability of these media for this study.

We addressed whether an increase in dietary yeast concentration during development would result in rDNA loss. In interphase cells, the rDNA is the genetic location of the cytogenetic Nucleolus Organizing Region (NOR), and thus the foundation of the nucleolus. Even single rRNA genes are capable of forming tiny supernumerary nucleoli [59]. Consequently, nucleolar morphology is sensitive to the overall size, activity level, and integrity of the rDNA arrays. In Drosophila, rDNA damage is readily observed in larval salivary glands. Damage and repair are thought to lead to extrachromosomal circles, which coalesce mini - or micro-nucleoli in non-dividing or post-mitotic cells, forming supernumerary nucleoli. Such “fragmentation” has been observed in flies mutant for “heterochromatin components” [23–26], where it is thought to stem from aberrant intrachromosomal recombination resulting in the formation of extrachromosomal rDNA circles [28, 60]. We performed immunofluorescence on larval salivary gland cells with an anti-fibrillarin antibody to detect the nucleolar dense fibrillar component which, in Drosophila, typically forms a single focus containing both X-linked and Y-linked rDNA arrays. We observed an elevated frequency of multiple nucleoli in SY30-fed larvae compared to SY10-fed larvae (Fig 1B and 1C). Multiple nucleoli (defined as more than one discrete separate fibrillarin focus) were present in 40% ± 24% of the nuclei within single salivary glands dissected from SY30-fed larvae; in contrast multiple nucleoli were observed in 7% ± 6% of salivary gland nuclei from from SY10-fed larvae. The ranges are standard deviations of nucleolar instability rates from independently grown and dissected salivary glands, indicating that populations of genetically-identical individuals raised on different food sources show variation that can be discriminated by their averages.
In dividing cells, acentric extrachromosomal rDNA circles are lost, effectively reducing rDNA copy number and shortening the rDNA array through development. To quantify rDNA loss stemming from diet-influenced extrachromosomal circle formation during development, we used real-time PCR to measure the rDNA abundance of flies raised as larvae on either SY10 or SY30. Relative copy number was quantified using genomic copy number of a tRNA gene as normalization resulting in a DNA-to-DNA proportion [55]. rDNA copy number differences were monitored by comparing the rDNA copy number in freshly eclosed F1 males to that of males taken from the “F0” parental stock (i.e., treated males compared to their fathers and uncles). Male adults were collected within hours of eclosion, thus any diet-influenced changes to rDNA copy number were the result of physiological effects initiated prior to metamorphosis. The rDNA copy number of flies raised on SY10 was indistinguishable from that of their sires, while those raised on SY30 exhibited an average copy number reduction of ~20% (Fig 1D). The standard deviation for these data are derived from independent biological replicate vials from sibling males. The standard deviation of the F0 (not presented on graph, but whose average is defined as 100%) is pooled into the SY10 and SY30 populations.
It has been known that culture conditions affect the extent of position effect variegation (PEV), the somaclonal variation that occurs at new juxtapositions of euchromatin and heterochromatin. Specifically, richer or more abundant food leads to reduced heterochromatin-induced gene silencing and thus higher levels of variegating gene expression [61, 62]. To address whether SY10 and SY30 alter the extent of PEV, we raised isogenic whitemottled-4 (wm4/Y,10B) males on SY10 or SY30. The only source of white+ expression in this genotype is from the whitemottled-4 allele. If SY30 reduced silencing, we expected to see increased eye pigmentation. Three days after eclosion, we decapitated males and extracted pigment from their heads in acidified alcohol. The extent of the suppression of variegation, observed as increased pigment extraction (Fig 1E), accords well with the increased expression as a result of reducing rDNA copy number through molecular means [24, 55] or from wild-caught strains [43, 63].
The quantified loss of rDNA due to SY30 is within the range of natural Y-linked rDNA variation [35, 55, 64], and within the experimental range used to demonstrate heterochromatin changes and gene regulatory variability by us and others [24, 64] on the Y,10B chromosome specifically. Therefore, altered diet could in principle be responsible in entirety or in part for the natural rDNA copy number variance observed in natural populations isolated from the wild.
Diet-induced rDNA loss is independent of R1 and R2 expression
In Saccharomyces, it is thought that rDNA damage and loss is a consequence of a collision between the replication fork and transcriptional machinery at the rDNA array [29]. This model accounts for the transcription-dependent nature of rDNA recombination as well as the relationship between rDNA stability, cell division, calorie-restriction, and aging [27, 50, 65]. Much less is known about the regulation of rDNA stability in multicellular organisms and, therefore, the precise mechanism underlying our observations is unknown.
There is no known analogous system for specific loss in Drosophila (or any of the metazoa), however the unique retrotransposable elements in arthropods provides one possibility. It is possible that loss of rDNA in Drosophila is a direct consequence of activity, for instance derepression of the rDNA leading to mobilization of R1 or R2 retrotransposons, whose mobilization is known to cause single-stranded DNA breaks. This possibility makes two predictions that we tested.
First, we investigated whether consumption of SY30 lead to pronounced derepression of R1 or R2, the retrotransposable elements resident in the Drosophila rDNA. We could not detect significant increases in expression of either when raised on SY10 or SY30 (Fig 2A). Second, we investigated whether R1 - or R2-interrupted 35S rRNA genes were preferentially lost after growth on SY30. We observed decreased R1-interrupted and R2-interrupted rDNA copies, as well as loss of uninterrupted 35S rDNA copies, but neither R1 nor R2 reductions were strongly biased over decreases as a result of I-CreI-induced damage (Fig 2B) [55].

It should be noted that absolute numbers of R1 - and R2 - containing 35S genes are not possible to reliably calculate, nor are our “R1” and “no R1,” or “R2” and “no R2,” numbers necessarily additive, because these data are derived from different priming real-time PCR oligonucleotides, thus do not have the same kinetics of amplification [55]. Nonetheless, comparisons between conditions are valid, and in these cases we see no strong biased loss of R1 or R2 sequences (judged by either R1 - and R2-containing or R1 - and R2-lacking rRNA genes). We cannot specifically quantify rRNA genes with both insertions (although these likely represent a vast minority of rRNA genes [66, 67]), or that are free from either retrotransposable element insertions (likely the vast majority). Together these data argue against an R1 - or R2-related mechanism of chromosome damage and rDNA loss. Work by Cohen and colleagues additionally showed that repetitious gene arrays that do not house such retrotransposons show progressive loss through the lifespan of Drosophila [28], similar to the rDNA and similar to what we observe in SY30 conditions.
However it requires extreme caution in interpreting these results. We know that SY30 causes about 20% reduction in total rDNA copy number, and the rate, extent, and cytological manifestation of this loss indicate that rDNA loss is not of individual rRNA genes. Instead the best inference is that large contiguous blocks of rRNA genes—containing 35S genes interrupted and uninterrupted by R1, R2, or both—are lost as groups. The arrangements of the inserted rDNA genes is thought to vary considerably [18], so recombination, damage-and-repair, or loss events that occur between non-juxtaposed rDNA genes may remove a considerable number of rRNA subtypes (uninterrupted, R1-interrupted, R2-interrupted, and double-interrupted) that themselves were not part of the mechanism of loss.
Altered insulin receptor activity (insulin/insulin-like signaling) affects nucleolar stability
If instability is a consequence of changes in nutrient availability, then we reasoned that modulating known nutrient signaling pathways should produce similar results. In Drosophila, as in Cænorhabditis elegans, mouse, and human, the insulin/insulin-like growth factor signaling (IIS) and TOR signaling networks mediate many cellular responses to nutrient availability, including ribosome biogenesis and rDNA expression [49, 68, 69]. We expressed a constitutively active form of the insulin receptor (InR.R418P) either generally in larvae (using a Ubiquitin promoter to drive GAL4 expression in all cells) or specifically in salivary glands (using the Sgs3 promoter to drive GAL4 expression in larval salivary glands). Because these cells do not undergo cytokinesis, rDNA removed from the chromosomal rDNA array (e.g., as extrachromosomal circles) would not be measurably lost using real-time PCR, thus we used the formation of supernumerary nucleoli as proxy for rDNA instability [23, 25]. In both cases we observed elevated levels of multiple nucleoli (~41% and 28% respectively) (Fig 3A and 3B), indicating that activating the Insulin Receptor was sufficient to induce instability. Raising flies expressing constitutive InR.R418P on food laced with rapamycin mitigated the effect (Fig 3C), suggesting inhibition of rDNA transcription suppressed the nucleolar instability. Expression of an antimorphic allele (InR.K1409A) had no apparent effect (Fig 3D), indicating that either antimorphic receptor expression levels were insufficient to reduce rDNA expression and affect loss rate, or the basal level of nucleolar instability we observe is independent of IIS-mediated rDNA expression.

Acute loss of nucleolar stability can be effected by insulin treatment in vitro
In order to limit our view to acute cell-autonomous effects of nutrient signaling perturbation, we next opted to modulate the activity of these pathways pharmacologically in cultured larval salivary glands, a fully developed, post-mitotic tissue. In this way we could separate developmental defects (as a result of prolonged expression of hypermorphic or antimorphic alleles) from acute defects caused by altered cell physiology.
We dissected larval salivary glands from flies expressing a Fibrillarin-RFP fusion protein [70] whose expression was controlled by a heat shock responsive hsp70 promoter. We did not induce expression with heat shock because sufficient nucleolar RFP was detectable without heat shock. Salivary glands were cultured in Drosophila cell/tissue culture medium for 22–24 hours in the presence (or absence) of recombinant human insulin. Treatment with insulin resulted in increased supernumerary nucleoli in live salivary glands (Fig 4A and 4B). Exposure to either actinomycin D or rapamycin (two drugs which block RNA Polymerase I activity, the former directly and the latter by inhibiting TOR) for two hours prior to insulin addition abrogated the multiple nucleolar morphology (Fig 4C and 4E). This pharmacological suppression of supernumerary nucleoli could not be reproduced if the drug was administered in the last two hours of insulin exposure (Fig 4D and 4F), supporting the interpretation that drug-induced stability of the nucleolus was a consequence of reduced rDNA transcription, rather than a reorganization of the nucleolus as a result of drug exposure. We confirmed that drug treatment reduced active rRNA expression by culturing eviscerated whole wild-type larvae in tissue culture medium for 24 hours in the presence of rapamycin or actinomycin D alone. Real-time PCR quantification of cDNAs of pre-processed rRNA junctions (as in Fig 1A) were reduced to 80% (+19.8%/-15.9%) and 69% (+13.1%/-11%) (N = 10 larvae for each condition), respectively, compared to control larvae cultured without any drug. That nucleolar instability is enhanced by insulin and mitigated by rapamycin and actinomycin D suggests that the consequential effect on rDNA stability occurs downstream of the convergence of the activities of these pharmacological agents.

Loss of rDNA manifests on meiotically-inherited chromosomes
Wild-caught Drosophila strains exhibit a wide variance in rDNA copy number [34, 35]; the source of this variance, however, is unknown. For this variability to be explained by environmentally induced instability during the life history of these chromosomes, germline rDNA would have to be susceptible to environmental influence. To look for possible germline effects of diet, we used a genetic strategy to specifically measure copy number of Y-linked rDNA. We chose to focus our attention on the Y-linked array because it is preferentially active in males [43, 71] and because of the ease with which the Y chromosome is manipulated genetically [55]. We genetically-isolated Y-linked rDNA arrays by crossing adult males to females bearing an rDNA-deficient compound X chromosome (C(1)DX) (Fig 5A). Female progeny of this cross were viable and carried the patroclinous Y-linked rDNA as their sole source of rRNA genes; any differences between rDNA in daughters were due to permanent germline changes to the chromosomes occurring in the fathers.

When male flies were raised as larvae on either “Standard” cornmeal medium (“Standard” or “Std”), or SY10 or SY30 (Fig 5A, symbol (1)) then moved to Standard food and outcrossed to C(1)DX virgin females, the progeny ((2)) had no detectable difference in the rDNA copy number (Fig 5B). Thus while the soma was undergoing diet-induced rDNA loss at this stage (Fig 1D), the germline was not susceptible to diet induced loss of rDNA; this was not unexpected because the germline cells are relatively quiescent in larvae.
Conversely, the adult germline was found to be sensitive to dietary conditions. Adult males (Fig 5C, (3)), raised from eggs on Standard medium, were collected 1–4 days after eclosion and allowed to mate with C(1)DX virgin females on Standard food for one day; the female progeny ((4)) of this cross served as “Control” (i.e., time-zero) datum for subsequent comparisons. The next day, the males were transferred to experimental conditions (Std, SY10, or SY30), with or without rapamycin, and were allowed to feed. Males ((5)) were crossed with fresh C(1)DX virgins after 20 days. In this way, we were able to sample the germline of the same group of males before and after treatment (Fig 5C).
Female progeny (“Treated” in Fig 5C, (6)) had no fewer Y-linked rDNA than their half-sisters if their fathers had spent 20 days eating Standard food (Fig 5D, compare “Control” and “Std”). In stark contrast, daughters of fathers who ate SY10 or SY30 for 20 days exhibited a reduced Y-rDNA copy number, which was different from daughters both of young fathers and fathers who had fed on Standard food for 20 days. rDNA copy number reduction was greater from SY30-fed fathers than from SY10-fed fathers. Thus the father’s germline underwent loss of rDNA in pre-meiotic mitotic (stem-cell) divisions during the 20 day period that they were exposed to SY10 or SY30, and the amount of loss was proportional to dietary largess.
Loss was mitigated when 10 µM rapamycin was included in the SY10 and SY30 food. It has previously been confirmed that rapamycin concentrations up to 200 μM have no effect on Drosophila feeding rates [72] suggesting that the effects of rapamycin on germline rDNA loss are pharmacological in origin as opposed to behavioral. Spermatogenesis and germline stem cell proliferation in adults are regulated by both diet and nutrient sensing pathways [73, 74]; it is likely that germline rDNA instability in response to increased dietary yeast concentration is a result of the modulation of these pathways, along with any subsequent changes in rDNA regulation.
Altered rDNA copy number can be stable for many years
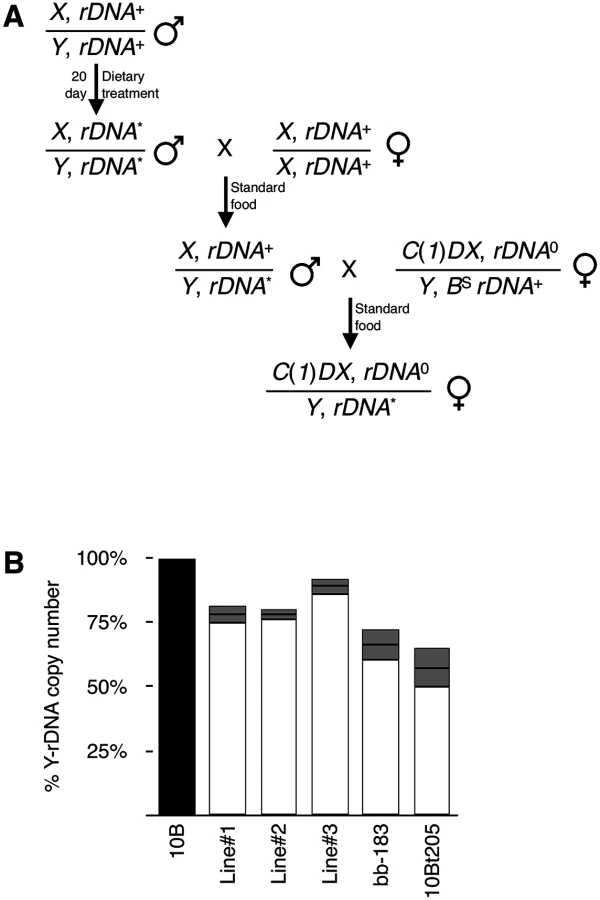
In order for a population to maintain a steady-state rDNA size, natural loss must be balanced by expansion. In Drosophila, rDNA magnification may serve this purpose, although magnification is not wide-spread and is only observed on some chromosomes under certain conditions [30–32, 75]; the determinative characteristics of chromosomes that exhibit magnification remain unknown. To test for this sort of reversion of diet-induced rDNA loss, we established independent lines from SY30-fed males and kept them on Standard food as with any Drosophila strain (Fig 6A). This allowed us to monitor transgenerational rDNA copy number for reversion or continued instability. We tested pooled males from three such independent lines that had been kept for two generations on Standard food after being raised for one generation and 20 days as adult on SY10 or SY30 and found that lost rDNA remained lost. Data shown in Fig 6B (Lines 1, 2, and 3) show standard error of the mean (S.E.M.) to report the average rDNA copy number of individuals line founded by single males. This observation is consistent with published findings (as well as our anecdotal observations) that while some engineered rDNA deletions lines exhibited moderate (~5%) expansion shortly after production [55], they have been otherwise stable, without selection, over many subsequent generations. Indeed, we tested one such line and found that in relation to the progenitor stock, changes—magnification or continued loss—had not occurred after six years on Standard food, corresponding to no fewer than sixty generations (Fig 6B, bb-183). Loss of rDNA was also achieved by passaging the Y,10B in a mutant heterozygous for Su(var)205, which encodes the Heterochromatin Protein 1 gene product. This mutation has been shown to cause rDNA instability [23] and rDNA loss [57], but when the Y,10B “tempered” by the mutation (10Bt205) was returned to a genetic background wild-type for Su(var)205, and passaged for a year (approximately 25 generations) the rDNA loss was not reverted (Fig 6C). These observations suggest that, just like any other polymorphism, rDNA deletions (naturally-occurring or otherwise) persist over multiple generations and that magnification, in contrast, is relatively rare.
Discussion
In this work we have established that ribosomal DNA (rDNA) copy number polymorphisms can be created by manipulating the diet of wild-type flies. By directly altering insulin-like signaling and phenocopying nucleolar instability in culture using recombinant insulin, we showed that normal IIS signaling can be a significant source of rDNA copy number variation in the soma. Diet-induced rDNA copy number changes occur in both the soma and germline. As a result, they are both permanent within an organism and are capable of being transmitted to subsequent generations, hence may act as a codex for dietary history of an individual or for a population.
Dietary modulation can account for loss of rDNA, but some unknown factor must be responsible for establishing some limit to the loss. We do not know the mechanism for this maintenance, although it could be an as-yet unobserved intentional regulated processes that assures minimal rDNA copy number, or it could be by normal selective pressures exerted by the Minute or bobbed phenotypes that result from very low ribosome number. Alternatively, loss may be balanced by gain of rDNA through unequal sister chromatid exchange, gene conversion, re-replication, or cycles of excision, rolling-circle replication, and re-integration. Meiotic magnification and somatic pseudo-magnification at the rDNA have long been known in Drosophila, although the identification of a mechanism has eluded researchers for over 40 years [32, 76]. Part of the asymptotic limit to loss may be the natural ecology of Drosophila, wherein older males (with greater loss, Fig 5) may be less likely to mate, produce fewer offspring, or produce an altered sex ratio; ecological experiments would be needed to address these possible contributions.
The rDNA is the major site of nuclear energy utilization—transcription, processing, packaging, and export—and was known to be responsive to the energy status of the cell. This response by rDNA to diet, and its fortuitous cleavage by I-CreI, allowed us to identify rDNA copy number as a factor which stabilizes the genome. This observation is now confirmed in Drosophila [26, 43] and similar hypotheses have been proposed for yeast rDNA [40, 77]. However it seems unlikely that the rDNA is alone in this ability. Half of the genome of Drosophila is composed of interspersed or tandem repeats—the transposable elements, highly-repetitive DNAs, expressed repeat gene clusters—and these sequences may account for some of the remaining regulatory variation that as yet has been unmapped [63]. It will require the ability to alter and measure copy numbers of the other repeated DNAs of the genome to ascertain if complex or quantitative traits map to these large blocks of “junk.”
Our observation of diet-induced rDNA loss integrates with previous results which indicate that rDNA copy number polymorphisms account for a large fraction of Y-linked gene regulatory variation (termed “YRV”) [43, 56, 63, 78], including the ability of heterochromatin to induce gene silencing (position effect variegation). Ecological phenotypic variation implied by gene expression differences may be quite significant in competitive, food-rich or food-scarce natural environments. Our observations may directly explain why food and culture conditions alter the extent of position effect variegation, and may further explain why chromosomes from different strains—natural isolates or mutant stocks—differ in their ability to suppress position effect variegation [43].
We believe that the diet - and IIS-induced rDNA instability we observe is a general, or at least common, feature of Y-linked rDNA because it has been measurable in males of many strains used in our work. For instance, we specifically tested two other Y chromosomes: a wild-type male from a laboratory Canton-S stock and a freshly wild-caught (“Texas-B”) male by backcrossing males from these strains to a y1; bw1; e1; eyR strain to genetically isolate the Y chromosome [63]. rDNA copy number of flies raised on SY10 or SY30 was compared and otherwise-isogenic males bearing the Canton-S exhibited a 38% decrease in rDNA copy number, while the Texas-B chromosome exhibited an 8% decrease. Thus, while diet-induced loss appears to be a common feature of Y-linked rDNA genes, there are likely other genetic factors that influence the rate or bounds of loss. Additionally, the two presumably-unrelated transgenic lines (the Y from the UAS-InR strain presented in Fig 3 and the Y from the Fibrillarin-RFP strain presented in Fig 4) both showed nucleolar instability under conditions with increased IIS signaling. The same phenomenon of rDNA loss was less clear in females, who appeared to exhibit small amounts of loss that was not statistically robust. Because the biology of X-linked rDNA arrays differs from that of the Y-linked arrays [71, 79, 80], and the consequence of X-X exchange at the rDNA is very different from that of X-Y exchange, we had no reason to believe that the phenomenon was related and pursued it no further.
rDNA instability is observed in a number of eukaryotes and is associated with a variety of complex phenotypes including position effect variegation in Drosophila [24, 43], replicative lifespan in yeast [65], plant size in flax [81], cancer progression in humans [82–84], and the aforementioned “hidden variation” of Y-linked Regulatory Variation. Our findings provide a mechanism for the influence of diet on all of these processes. Our findings are likely generally relevant to many organisms due to the conserved structure of ribosomal DNA arrays, the common copy number polymorphisms at that locus [34], and the common modes of rDNA regulation [85]. While we focused on diet, other processes that influence rRNA transcription (e.g. cell proliferation, DNA damage, determination and differentiation, stress, aging, temperature, etc. [86]) would presumably also affect rDNA stability via similar mechanisms, and thus, the rDNA may be a common mediator of induced and heritable effects. We do not expect that induced changes to the genome are limited to the rDNA, in fact satellite sequences show copy number polymorphisms that are only now being investigated [57, 87]. In terms of epigenetic inheritance, it is unclear whether diet-induced rDNA copy number polymorphisms may act as an inducible and heritable modifying mutation that subsequently destabilizes epigenetic silencing.
Materials and Methods
Fly strains and husbandry
y1 w67c23/Dp(1;Y) y+, P{w = RS5}10B was described previously [55, 88], and was used for all feeding experiments. Constitutively active insulin receptor expression was performed using the following stocks: w*; P{Ubi-GAL4}2/CyO, w1118; P{Sgs3-GAL4.PD}TP1, y1 w1118; P{UAS-InR.R418P}2, and y1 w1118; P{w[+mC] = UAS-InR.K1409A}2 [89], all of which were obtained from the Bloomington Drosophila Stock Center. w1118; P{w[+mC] = UAS-mRFP-Fib} was used for in vitro salivary gland culturing experiments to visualize nucleolar fibrillarin [70], and was a gift from Dr. Patrick DiMario. Expression was under control of the core hsp70 promoter on the pUAST backbone, which provided ample expression to visualize without a GAL4 driver. C(1)DX, y1 f 1 bb0 was used to genetically isolate Y chromosomes for real-time PCR analysis. The rDNA deletion strain used in Fig 2 and 6 for comparison was y1 w67c23/Dp(1;Y) y+, P{w = RS5}10B, Df(rDNA)bb-183 [55], and the 10Bt205 strain in Fig 6 is described in Aldrich and Maggert [57]. All crosses were performed at 25°C and 80% humidity. Unless otherwise noted in the experiment all stocks were maintained on standard cornmeal molasses medium.
Feeding experiments
We used two experimental media for our feeding experiments. The first was based on SYA media used in dietary restriction studies [52] and contained 5% sucrose, 10% hydrolyzed yeast, 5% cornmeal (w/v), and 1% agar (w/v). These ingredients were boiled in deionized water and mixed until fully dissolved. Media was allowed to cool to 55°C before the addition of propionic acid and tegosept to 0.3% each. We refer to this diet as SY10. SY30 was identical except it contained 30% yeast (w/v). One-third of the required yeast was added in increments during heating to allow easy dissolution. Standard (cornmeal) medium is 5% cornmeal (w/v), 3% yeast (w/v), 1% agar (w/v), 7% molasses (v/v), and supplemented with proprionic acid and tegosept as above.
For experiments testing the effects of larval diet, we collected embryos overnight on apple juice agar plates covered with yeast paste. Embryos were washed off plates using 1X PBS and transferred to experimental media at a uniform density [90]. To measure adult germline effects, males raised on Standard medium were collected 1–4 days post eclosion and crossed in groups of five to C(1)DX/Y virgins on Standard medium. After 24 hours, the males were removed and placed on either Standard medium, or Experimental media with or without 10 μM Rapamycin (LKT Laboratories), while females were maintained on Standard medium and allowed to lay eggs. Progeny from these females served as the baseline for subsequent comparisons. Males were transferred to fresh experimental media every 3 days for 20 days, after which they were again crossed to virgin C(1)DX females on Standard medium. Additional adult males fed in the above manner were crossed to y1 w67c23 females to establish stocks for subsequent analysis.
RNA analyses
Expression of the rRNA faces three problems. First, it is very stable (by RNA standards), with a half-life of at least two days [91], so steady-state rRNA levels are insufficient to detect changes in rRNA transcription. Second, it is very abundant, accounting for approximately 50% of transcription and 80–90% of steady-state RNA [20], so small differences in loading result in large variance in apparent rRNA concentration. Additionally, selection of any mRNA as a comparison (“denominator” in relative-abundance calculations) introduces even more variance as the quantification of differences in rRNA is more sensitive than differences in mRNAs with lower abundance (i.e., a 10% difference of 1000 is easier to detect than a 10% difference of 10). Third, the a priori assumption that any mRNA may not change in conditions in which rRNA expression changes may not be valid. For these reasons, we measured active pre-rRNA by detecting cDNAs derived from the ETS-18S junction, using reverse transcription primer GGAGGACGAGAAAATTGACAGACGCTTTGAGACAAGCATATAA. This primer was designed to be complementary to the 18S at the junction of ETS and 18S, and have a novel sequence at the 5’ end for use in subsequent real time PCR.
RNA and DNA were co-purified to satisfy the need for a stable (non-regulable) denominator for rRNA transcription levels. Total nucleic acids were purified from pools of either one hundred second instar larvae (for Fig 1A) or ten dissected and everted third instar larvae (for measuring effects of rapamycin and actinomycinD). Tissue was homogenized in a solution containing 50 mM EDTA, 100 mM Tris pH 7.0, and 1% SDS. Homogenate was extracted twice in equal volumes phenol:chloroform:isoamyl alcohol (25 : 24 : 1, buffered with 1 M Tris pH 7.0). Under these conditions, all nucleic acids partition to the aqueous phase [92], which was further extracted with chloroform followed by diethyl ether. Nucleic acids were precipitated with 0.8 volumes propanol, washed with RNAse-free 70% ethanol, and resuspended in DNAse - and RNAse-free water. Reverse transcription was performed in 20 µL reactions with 2 µg nucleic acid, 2 units M-MuLV Reverse Transcriptase (New England Biolabs), 1X Reverse Transcriptase Buffer, 2 µM of each dNTP, and 4 µM primer for 1 hour at 42°C followed by 10 minutes at 90°C. Samples were diluted 1 : 250 for subsequent real time PCR reactions, as described below and before [55, 57]. Melt-curve analysis was used to assure single melt peaks, and reaction efficiencies were determined using LinRegPCR [93] (average efficiency for the tRNA gene was 92% and for the ETS-18S was 82%). Efficiency correction and fold changes were calculated as before [94].
For detection of retrotransposon R1 and R2 expression, RNA was extracted as previously described [95] from three pools of 20 male wandering third instar larvae. Reverse-Transcriptase reactions for R1, R2, and rho1 cDNAs were individually primed using CCAGCATACGTATGCTCGCTG (for R1), GGGAGTGATTGGAGTTGTTTCCG (for R2), or CTAGCGAATCGGGTGAATCCACTG (for rho1). Real-time PCR reactions were then performed as below using those same primers and additionally GGGACAGCTTAGTGCACTCTAC (for R1), CCCCGGAGTTGCTAATCTAACC (for R2), and GTGGAGCTGGCCTTGTGGG (for rho1). Negative controls for R1 and R2 used the rho1 cDNA template reaction and R1 or R2 real time primer pairs.
Immunofluorescence and microscopy
Whole mount salivary gland immunofluorescence was performed as in (11). Glands were dissected from 3rd instar larvae in 1X PBS and were then transferred to PBT (PBS containing 0.1% Triton X-100) and fixed in PBT containing 3.7% formaldehyde and blocked for 1 hour in PBT supplemented with 1% BSA. Glands were washed and incubated overnight at 4°C with a mouse anti-Fibrillarin primary antibody (Abcam) diluted 1 : 1000 in PNBT (PBT containing 1% BSA and 500 mM NaCl). Goat anti-mouse conjugated to rhodamine was used as a secondary antibody (1 : 1000). DNA was counterstained with 1 ng/mL DAPI (MP Biomedicals). All images were obtained using a Zeiss Axioskop 2 epifluorescence microscope running AxioVision (v. 4.6.3.0) with a 20X objective (numerical aperature = 0.5). Sequential excitation was performed at 543 nm (for Rhodamine and RFP) and 405 nm (for DAPI).
In vitro salivary gland culture
Larvae containing the homozygous P{w[+mC] = UAS-mRFP-Fib} were reared in bottles and aged to the early wandering stage of the third instar. Males were dissected in 1X PBS and salivary glands were moved to Schneider media supplemented with 50 mg/mL streptomycin, 50 mg/mL penicillin, and 10% heat-inactivated fetal bovine serum (Gibco). Five to ten glands per condition were co-cultured and treated with 5 µM human insulin (Sigma) for 22–24 hours, then stained with 1 ng/mL DAPI for one hour prior to visualization. For “pre-treatment,” salivary glands were treated with 10 µM rapamycin or 0.6 µM actinomycin D (Acros) for two hours prior to insulin addition. For “post-treatment,” glands were treated with rapamycin or actinomycin D two hours prior to DAPI addition. Images were taken of all stained salivary gland lobes near the anterior end at 20X, post-processed for bright/contrast, and scored for nucleolar structure. A nucleus was determined to contain “multiple” nucleoli if more than one separate focus of fibrillarin fluorescence was discernible. Glands that had no or poor DAPI staining were assumed to be damaged by treatment and excluded from analysis. The entire dissection and culture experiment was performed eight times over the course of two weeks and all data pooled to calculate frequencies presented in Fig 4.
Quantitative polymerase chain reaction (real-time PCR)
Real-time PCR analysis was performed as described in [55]. DNA was extracted from newly eclosed adult male flies in pools of three or more and quantified using a NanoDrop ND-1000. Real-time PCR was performed with a StepOne Real-time PCR system and Power SYBR Green reagents (Applied Biosciences) using 4 ng DNA per reaction. 18S rDNA was amplified using primers AGCCTGAGAAACGGCTACCA and AGCTGGGAGTGGGTAATTTAC, while the endogenous loading control, tRNAK-CTT, was amplified using CTAGCTCAGTCGGTAGAGCATGA and CCAACGTGGGGCTCGAAC. Relative differences were calculated using the “ΔΔCT” method. Each data point presented consists of at least three independent biological samples quantified in triplicated or quadruplicated technical replicates. Error bars generally represent the standard deviation of biological replicates, as justified in the text [96]; error bars are generally asymmetric around the mean (“+” values are higher than “-”values”) because we calculate error on CT values prior to exponential transformation (2CT) for presentation. P-values were calculated from ΔΔCT values prior to exponential transformation (to get “fold” values) using StatPlus:mac v. 5.8 (AnalystSoft) or Apple Numbers.
Zdroje
1. Anway M.D., et al., Epigenetic transgenerational actions of endocrine disruptors and male fertility. Science, 2005. 308(5727): p. 1466–9. 15933200
2. Jirtle R.L. and Skinner M.K., Environmental epigenomics and disease susceptibility. Nat Rev Genet, 2007. 8(4): p. 253–62. 17363974
3. Arico J.K., et al., Epigenetic Patterns Maintained in Early Caenorhabditis elegans Embryos Can Be Established by Gene Activity in the Parental Germ Cells. PLoS Genet, 2011. 7(6): p. e1001391. doi: 10.1371/journal.pgen.1001391 21695223
4. Feil R. and Fraga M.F., Epigenetics and the environment: emerging patterns and implications. Nat Rev Genet, 2011. 13(2): p. 97–109. doi: 10.1038/nrg3142 22215131
5. Greer E.L., et al., Transgenerational epigenetic inheritance of longevity in Caenorhabditis elegans. Nature, 2011. 479(7373): p. 365–71. doi: 10.1038/nature10572 22012258
6. Seong K.H., et al., Inheritance of stress-induced, ATF-2-dependent epigenetic change. Cell, 2011. 145(7): p. 1049–61. doi: 10.1016/j.cell.2011.05.029 21703449
7. Crews D., et al., Epigenetic transgenerational inheritance of altered stress responses. Proc Natl Acad Sci U S A, 2012. 109(23): p. 9143–8. doi: 10.1073/pnas.1118514109 22615374
8. Padmanabhan N., et al., Mutation in folate metabolism causes epigenetic instability and transgenerational effects on development. Cell, 2013. 155(1): p. 81–93. doi: 10.1016/j.cell.2013.09.002 24074862
9. Ahmad K. and Henikoff S., Epigenetic consequences of nucleosome dynamics. Cell, 2002. 111(3): p. 281–4. 12419239
10. Deal R.B., Henikoff J.G., and Henikoff S., Genome-wide kinetics of nucleosome turnover determined by metabolic labeling of histones. Science, 2010. 328(5982): p. 1161–4. doi: 10.1126/science.1186777 20508129
11. Hathaway N.A., et al., Dynamics and memory of heterochromatin in living cells. Cell, 2012. 149(7): p. 1447–60. doi: 10.1016/j.cell.2012.03.052 22704655
12. Teves S.S., Deal R.B., and Henikoff S., Measuring genome-wide nucleosome turnover using CATCH-IT. Methods Enzymol, 2012. 513: p. 169–84. doi: 10.1016/B978-0-12-391938-0.00007-0 22929769
13. Ptashne M., Epigenetics: core misconcept. Proc Natl Acad Sci U S A, 2013. 110(18): p. 7101–3. doi: 10.1073/pnas.1305399110 23584020
14. Struhl K., Is DNA methylation of tumour suppressor genes epigenetic? Elife, 2014. 3: p. e02475. doi: 10.7554/eLife.02475 24623307
15. Ahmad K. and Golic K.G., Somatic reversion of chromosomal position effects in Drosophila melanogaster. Genetics, 1996. 144(2): p. 657–70. 8889528
16. Conconi A., et al., Two different chromatin structures coexist in ribosomal RNA genes throughout the cell cycle. Cell, 1989. 57(5): p. 753–61. 2720786
17. Riddle N.C. and Richards E.J., Genetic variation in epigenetic inheritance of ribosomal RNA gene methylation in Arabidopsis. Plant J, 2005. 41(4): p. 524–32. 15686517
18. Eickbush D.G., et al., Epigenetic regulation of retrotransposons within the nucleolus of Drosophila. Mol Cell Biol, 2008. 28(20): p. 6452–61. doi: 10.1128/MCB.01015-08 18678644
19. Grummt I. and Langst G., Epigenetic control of RNA polymerase I transcription in mammalian cells. Biochim Biophys Acta, 2013. 1829(3–4): p. 393–404.
20. Warner J.R., The economics of ribosome biosynthesis in yeast. Trends Biochem Sci, 1999. 24(11): p. 437–40. 10542411
21. Pellegrini M., Manning J., and Davidson N., Sequence arrangement of the rDNA of Drosophila melanogaster. Cell, 1977. 10(2): p. 213–4. 402223
22. French S.L., et al., In exponentially growing Saccharomyces cerevisiae cells, rRNA synthesis is determined by the summed RNA polymerase I loading rate rather than by the number of active genes. Mol Cell Biol, 2003. 23(5): p. 1558–68. 12588976
23. Peng J.C. and Karpen G.H., H3K9 methylation and RNA interference regulate nucleolar organization and repeated DNA stability. Nat Cell Biol, 2007. 9(1): p. 25–35. 17159999
24. Paredes S. and Maggert K.A., Ribosomal DNA contributes to global chromatin regulation. Proc Natl Acad Sci U S A, 2009. 106(42): p. 17829–34. doi: 10.1073/pnas.0906811106 19822756
25. Guerrero P.A. and Maggert K.A., The CCCTC-binding factor (CTCF) of Drosophila contributes to the regulation of the ribosomal DNA and nucleolar stability. PLoS One, 2011. 6(1): p. e16401. doi: 10.1371/journal.pone.0016401 21283722
26. Larson K., et al., Heterochromatin formation promotes longevity and represses ribosomal RNA synthesis. PLoS Genet, 2012. 8(1): p. e1002473. doi: 10.1371/journal.pgen.1002473 22291607
27. Saka K., et al., Cellular senescence in yeast is regulated by rDNA noncoding transcription. Curr Biol, 2013. 23(18): p. 1794–8. doi: 10.1016/j.cub.2013.07.048 23993840
28. Cohen S., Yacobi K., and Segal D., Extrachromosomal circular DNA of tandemly repeated genomic sequences in Drosophila. Genome Res, 2003. 13(6A): p. 1133–45. 12799349
29. Takeuchi Y., Horiuchi T., and Kobayashi T., Transcription-dependent recombination and the role of fork collision in yeast rDNA. Genes Dev, 2003. 17(12): p. 1497–506. 12783853
30. Tartof K.D., Unequal mitotic sister chromatid exchange and disproportionate replication as mechanisms regulating ribosomal RNA gene redundancy. Cold Spring Harb Symp Quant Biol, 1974. 38: p. 491–500. 4208791
31. Hawley R.S. and Tartof K.D., A two-stage model for the control of rDNA magnification. Genetics, 1985. 109(4): p. 691–700. 3921426
32. Bianciardi A., et al., Ribosomal DNA organization before and after magnification in Drosophila melanogaster. Genetics, 2012. 191(3): p. 703–23. doi: 10.1534/genetics.112.140335 22505623
33. Dawid I.B., Wellauer P.K., and Long E.O., Ribosomal DNA in Drosophila melanogaster. I. Isolation and characterization of cloned fragments. J Mol Biol, 1978. 126(4): p. 749–68. 106128
34. Long E.O. and Dawid I.B., Repeated genes in eukaryotes. Annu Rev Biochem, 1980. 49: p. 727–64. 6996571
35. Lyckegaard E.M. and Clark A.G., Ribosomal DNA and Stellate gene copy number variation on the Y chromosome of Drosophila melanogaster. Proc Natl Acad Sci U S A, 1989. 86(6): p. 1944–8. 2494656
36. Stage D.E. and Eickbush T.H., Sequence variation within the rRNA gene loci of 12 Drosophila species. Genome Res, 2007. 17(12): p. 1888–97. 17989256
37. Boulon S., et al., The nucleolus under stress. Mol Cell, 2010. 40(2): p. 216–27. doi: 10.1016/j.molcel.2010.09.024 20965417
38. Ide S., et al., Abundance of ribosomal RNA gene copies maintains genome integrity. Science, 2010. 327(5966): p. 693–6. doi: 10.1126/science.1179044 20133573
39. Boyd M.T., Vlatkovic N., and Rubbi C.P., The nucleolus directly regulates p53 export and degradation. J Cell Biol, 2011. 194(5): p. 689–703. doi: 10.1083/jcb.201105143 21893597
40. Kobayashi T., Regulation of ribosomal RNA gene copy number and its role in modulating genome integrity and evolutionary adaptability in yeast. Cell Mol Life Sci, 2011. 68(8): p. 1395–403. doi: 10.1007/s00018-010-0613-2 21207101
41. Paredes S., The Ribosomal DNA Genes Influence Genome-Wide Gene Expression in Drosophila melanogaster., in Biology. 2011, Texas A&M University. p. 175. doi: 10.1016/j.jphotobiol.2011.08.009 21955546
42. Tsurumi A. and Li W.X., Global heterochromatin loss: a unifying theory of aging? Epigenetics, 2012. 7(7): p. 680–8. doi: 10.4161/epi.20540 22647267
43. Zhou J S.T., Martinsen L, Lemos B, Eickbush TH, Hartl DL, Y chromosome mediates ribosomal DNA silencing and modulates the chromatin state in Drosophila. Proc Natl Acad Sci U S A, 2012. doi: 10.1073/pnas.1207367109(Online).
44. Chen S., et al., Repression of RNA polymerase I upon stress is caused by inhibition of RNA-dependent deacetylation of PAF53 by SIRT7. Mol Cell, 2013. 52(3): p. 303–13. doi: 10.1016/j.molcel.2013.10.010 24207024
45. Grummt I., The nucleolus-guardian of cellular homeostasis and genome integrity. Chromosoma, 2013. 122(6): p. 487–97. 24022641
46. Gibbons J.G., et al., Ribosomal DNA copy number is coupled with gene expression variation and mitochondrial abundance in humans. Nat Commun, 2014. 5: p. 4850. doi: 10.1038/ncomms5850 25209200
47. Maggert K.A., Genetics: polymorphisms, epigenetics, and something in between. Genet Res Int, 2012. 2012: p. 867951. doi: 10.1155/2012/867951 22567405
48. Grummt I. and Ladurner A.G., A metabolic throttle regulates the epigenetic state of rDNA. Cell, 2008. 133(4): p. 577–80. doi: 10.1016/j.cell.2008.04.026 18485866
49. Murayama A., et al., Epigenetic control of rDNA loci in response to intracellular energy status. Cell, 2008. 133(4): p. 627–39. doi: 10.1016/j.cell.2008.03.030 18485871
50. Smith D.L. Jr., et al., Calorie restriction effects on silencing and recombination at the yeast rDNA. Aging Cell, 2009. 8(6): p. 633–42. doi: 10.1111/j.1474-9726.2009.00516.x 19732044
51. Fontana L., Partridge L., and Longo V.D., Extending healthy life span—from yeast to humans. Science, 2010. 328(5976): p. 321–6. doi: 10.1126/science.1172539 20395504
52. Grandison R.C., et al., Effect of a standardised dietary restriction protocol on multiple laboratory strains of Drosophila melanogaster. PLoS One, 2009. 4(1): p. e4067. doi: 10.1371/journal.pone.0004067 19119322
53. Zhou J., et al., Y chromosome mediates ribosomal DNA silencing and modulates the chromatin state in Drosophila. Proc Natl Acad Sci U S A, 2012. 109(25): p. 9941–6. doi: 10.1073/pnas.1207367109 22665801
54. Zhang Q., Shalaby N.A., and Buszczak M., Changes in rRNA transcription influence proliferation and cell fate within a stem cell lineage. Science, 2014. 343(6168): p. 298–301. doi: 10.1126/science.1246384 24436420
55. Paredes S. and Maggert K.A., Expression of I-CreI Endonuclease Generates Deletions Within the rDNA of Drosophila. Genetics, 2009. 181(4): p. 1661–71. doi: 10.1534/genetics.108.099093 19171942
56. Paredes S., et al., Ribosomal DNA deletions modulate genome-wide gene expression: "rDNA-sensitive" genes and natural variation. PLoS Genet, 2011. 7(4): p. e1001376. doi: 10.1371/journal.pgen.1001376 21533076
57. Aldrich J.C. and Maggert K.A., Quantitative PCR reveals naturally occurring and mutationally-induced repetitive sequence variation on the Drosophila Y chromosome. PLoS ONE, 2014. 9: e109906. doi: 10.1371/journal.pone.0109906 25285439
58. Maggert, K.A., Reduced rDNA Copy Number Does Not Affect "Competitive" Chromosome Pairing in XYY Males of Drosophila melanogaster. G3 (Bethesda), 2014.
59. Karpen G.H., Schaefer J.E., and Laird C.D., A Drosophila rRNA gene located in euchromatin is active in transcription and nucleolus formation. Genes Dev, 1988. 2(12B): p. 1745–63. 3149250
60. Peng J.C. and Karpen G.H., Epigenetic regulation of heterochromatic DNA stability. Curr Opin Genet Dev, 2008. 18(2): p. 204–11. doi: 10.1016/j.gde.2008.01.021 18372168
61. Spofford J.B., Position-effect variegation in Drosophila, in The Genetics and Biology of Drosophila, Ashburner M. and Novitski E., Editors. 1976, Academic Press. p. 955–1019.
62. Ashburner M., Golic K.G., and Hawley R.S., Drosophila: a laboratory handbook. 2nd ed. 2005, Cold Spring Harbor, N.Y.: Cold Spring Harbor Laboratory Press. xxviii, 1409 p.
63. Lemos B., Araripe L.O., and Hartl D.L., Polymorphic Y chromosomes harbor cryptic variation with manifold functional consequences. Science, 2008. 319(5859): p. 91–3. doi: 10.1126/science.1148861 18174442
64. Jiang P.P., Hartl D.L., and Lemos B., Y Not a Dead End: Epistatic Interactions Between Y-linked Regulatory Polymorphisms and Genetic Background Affect Global Gene Expression in Drosophila melanogaster. Genetics, 2010.
65. Kwan E.X., et al., A Natural Polymorphism in rDNA Replication Origins Links Origin Activation with Calorie Restriction and Lifespan. PLoS Genet, 2013. 9(3): p. e1003329. doi: 10.1371/journal.pgen.1003329 23505383
66. Jakubczak J.L., et al., Turnover of R1 (type I) and R2 (type II) retrotransposable elements in the ribosomal DNA of Drosophila melanogaster. Genetics, 1992. 131(1): p. 129–42. 1317313
67. Eickbush T.H., et al., Evolution of R1 and R2 in the rDNA units of the genus Drosophila. Genetica, 1997. 100(1–3): p. 49–61. 9440280
68. Grewal S.S., Evans J.R., and Edgar B.A., Drosophila TIF-IA is required for ribosome synthesis and cell growth and is regulated by the TOR pathway. J Cell Biol, 2007. 179(6): p. 1105–13. 18086911
69. Grewal S.S., Insulin/TOR signaling in growth and homeostasis: a view from the fly world. Int J Biochem Cell Biol, 2009. 41(5): p. 1006–10. doi: 10.1016/j.biocel.2008.10.010 18992839
70. Rosby R., et al., Knockdown of the Drosophila GTPase nucleostemin 1 impairs large ribosomal subunit biogenesis, cell growth, and midgut precursor cell maintenance. Mol Biol Cell, 2009. 20(20): p. 4424–34. doi: 10.1091/mbc.E08-06-0592 19710426
71. Greil F. and Ahmad K., Nucleolar Dominance of the Y Chromosome in Drosophila melanogaster. Genetics, 2012.
72. Bjedov I., et al., Mechanisms of life span extension by rapamycin in the fruit fly Drosophila melanogaster. Cell Metab, 2010. 11(1): p. 35–46. doi: 10.1016/j.cmet.2009.11.010 20074526
73. Ueishi S., Shimizu H., and H.I Y, Male germline stem cell division and spermatocyte growth require insulin signaling in Drosophila. Cell Struct Funct, 2009. 34(1): p. 61–9. 19384053
74. Wang L., McLeod C.J., and Jones D.L., Regulation of adult stem cell behavior by nutrient signaling. Cell Cycle, 2011. 10(16): p. 2628–34. 21814033
75. Endow S.A., Molecular characterization of ribosomal genes on the Ybb - chromosome of Drosophila melanogaster. Genetics, 1982. 102(1): p. 91–9. 6813192
76. Ritossa F.M., Unstable redundancy of genes for ribosomal RNA. Proc Natl Acad Sci U S A, 1968. 60(2): p. 509–16. 5248808
77. Kobayashi T., A new role of the rDNA and nucleolus in the nucleus—rDNA instability maintains genome integrity. Bioessays, 2008. 30(3): p. 267–72. doi: 10.1002/bies.20723 18293366
78. Sackton T.B., et al., Interspecific Y chromosome introgressions disrupt testis-specific gene expression and male reproductive phenotypes in Drosophila. Proc Natl Acad Sci U S A, 2011. 108(41): p. 17046–51. doi: 10.1073/pnas.1114690108 21969588
79. Tartof K.D., Regulation of ribosomal RNA gene multiplicity in Drosophila melanogaster. Genetics, 1973. 73(1): p. 57–71. 4631601
80. Endow S.A. and Glover D.M., Differential replication of ribosomal gene repeats in polytene nuclei of Drosophila. Cell, 1979. 17(3): p. 597–605. 113105
81. Schneeberger R.G. and Cullis C.A., Specific DNA alterations associated with the environmental induction of heritable changes in flax. Genetics, 1991. 128(3): p. 619–30. 1678726
82. Montanaro L., Trere D., and Derenzini M., Nucleolus, ribosomes, and cancer. Am J Pathol, 2008. 173(2): p. 301–10. doi: 10.2353/ajpath.2008.070752 18583314
83. Derenzini M., Montanaro L., and Trere D., What the nucleolus says to a tumour pathologist. Histopathology, 2009. 54(6): p. 753–62. doi: 10.1111/j.1365-2559.2008.03168.x 19178588
84. Drygin D., Rice W.G., and Grummt I., The RNA polymerase I transcription machinery: an emerging target for the treatment of cancer. Annu Rev Pharmacol Toxicol, 2010. 50: p. 131–56. doi: 10.1146/annurev.pharmtox.010909.105844 20055700
85. McStay B. and Grummt I., The epigenetics of rRNA genes: from molecular to chromosome biology. Annu Rev Cell Dev Biol, 2008. 24: p. 131–57. doi: 10.1146/annurev.cellbio.24.110707.175259 18616426
86. Grummt I., The nucleolus-guardian of cellular homeostasis and genome integrity. Chromosoma, 2013.
87. Wei K.H., et al., Correlated variation and population differentiation in satellite DNA abundance among lines of Drosophila melanogaster. Proc Natl Acad Sci U S A, 2014. 111(52): p. 18793–8. doi: 10.1073/pnas.1421951112 25512552
88. Maggert K.A. and Golic K.G., Highly efficient sex chromosome interchanges produced by I-CreI expression in Drosophila. Genetics, 2005. 171(3): p. 1103–14. 16020774
89. Wu Q., et al., Regulation of hunger-driven behaviors by neural ribosomal S6 kinase in Drosophila. Proc Natl Acad Sci U S A, 2005. 102(37): p. 13289–94. 16150727
90. Clancy D.J. and Kennington W.J., A simple method to achieve consistent larval density in bottle cultures. Drosophila Information Service, 2001. 84: p. 168–169.
91. Winkles J.A., Phillips W.H., and Grainger R.M., Drosophila ribosomal RNA stability increases during slow growth conditions. J Biol Chem, 1985. 260(12): p. 7716–20. 3922989
92. Shimada H., et al., Normalization using ploidy and genomic DNA copy number allows absolute quantification of transcripts, proteins and metabolites in cells. Plant Methods, 2010. 6: p. 29. doi: 10.1186/1746-4811-6-29 21190547
93. Ramakers C., et al., Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci Lett, 2003. 339(1): p. 62–6. 12618301
94. Pfaffl M.W., A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res, 2001. 29(9): p. e45. 11328886
95. Bogart K. and Andrews J., Extraction of Total RNA from Drosophila, in CGB Technical Report 2006–10. 2006, The Center for Genomics and Bioinformatics, Indiana University, Bloomington, Indiana.
96. Krzywinski M. and Altman N., Points of significance: error bars. Nat Methods, 2013. 10(10): p. 921–2. doi: 10.1038/nmeth.2659 24161969
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2015 Číslo 4
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Lack of GDAP1 Induces Neuronal Calcium and Mitochondrial Defects in a Knockout Mouse Model of Charcot-Marie-Tooth Neuropathy
- Proteolysis of Virulence Regulator ToxR Is Associated with Entry of into a Dormant State
- Frameshift Variant Associated with Novel Hoof Specific Phenotype in Connemara Ponies
- Ataxin-2 Regulates Translation in a New BAC-SCA2 Transgenic Mouse Model
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy