
A Zinc Finger Motif-Containing Protein Is Essential for Chloroplast RNA Editing
Transcripts encoding chloroplast and mitochondrial proteins of flowering plants are profoundly affected by RNA editing. In Arabidopsis, over 600 genomically-encoded Cs are modified to Us in organelle transcripts, altering the encoded amino acids and creating stop and start codons. Pentatricopeptide proteins are known to bind to cis-elements near C targets of editing and chloroplast RNA editing also requires members of two additional protein families. Nevertheless, not all protein components of the editosome have been identified. We now report the discovery of a member of fourth gene family essential for chloroplast RNA editing: OZ1, member of a family of Arabidopsis RanBP2-type zinc finger proteins. Identifying all of the proteins in the RNA editosome is critical for understanding the mechanism behind the remarkable specificity of C-to-U editing.
Published in the journal:
. PLoS Genet 11(3): e32767. doi:10.1371/journal.pgen.1005028
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1005028
Summary
Transcripts encoding chloroplast and mitochondrial proteins of flowering plants are profoundly affected by RNA editing. In Arabidopsis, over 600 genomically-encoded Cs are modified to Us in organelle transcripts, altering the encoded amino acids and creating stop and start codons. Pentatricopeptide proteins are known to bind to cis-elements near C targets of editing and chloroplast RNA editing also requires members of two additional protein families. Nevertheless, not all protein components of the editosome have been identified. We now report the discovery of a member of fourth gene family essential for chloroplast RNA editing: OZ1, member of a family of Arabidopsis RanBP2-type zinc finger proteins. Identifying all of the proteins in the RNA editosome is critical for understanding the mechanism behind the remarkable specificity of C-to-U editing.
Introduction
In vascular plants, specific cytidines are converted to uridines by RNA editing in the chloroplast transcripts [1–3]. A typical land plant modifies 30 to 40 C targets in chloroplasts, usually changing the encoded amino acid, and also acts upon hundreds of Cs in plant mitochondria [4,5]. The process is believed to be a correction mechanism to restore functional mRNAs in chloroplasts and mitochondria, whose genomes have undergone otherwise detrimental T-to-C changes [6].
The composition of the molecular machines that carry out plant organellar RNA editing, the editosomes, is not yet fully understood. Editosomes are found between the 200 and 400 kD markers on size exclusion columns [7]. Specificity of editing is achieved through the recognition of a cis-element 5’ adjacent to the editable cytidine by a pentatricopeptide repeat (PPR) motif-containing protein [8–10]. Recognition codes that match particular PPRs with nucleotides within the bound RNA region have been proposed [11,12]. Multiple PPRs in editing factors are followed by a so-called E domain and many PPR protein editing factors also contain a C-terminal DYW domain [13]. The DYW domain exhibits sequence motifs characteristic of cytidine deaminases [14] and is required for editing activity of some PPR-DYW proteins but is dispensable in others [15–20]. Attempts to demonstrate deaminase activity of purified DYW domains have failed so far [19,21]. Nevertheless, it remains possible that a DYW domain providing an enzymatic activity needed to deaminate cytidine to uridine could be present in all editosomes even if not on all PPR recognition factors. A protein named DYW1, which lacks any PPRs, was found to be required for editing of a chloroplast C target that is recognized by a PPR-E factor that lacks a DYW domain [22]. Mutating conserved residues characteristic of deaminases in DYW1 or in the DYW domains of QED1 and RARE1 results in impaired editing [23,24].
In addition to the large PPR protein family that provides site-specific recognition, members of two other plant protein families have been identified as components of editosomes, the RIP/MORF family and the ORRM family [7,25,26]. As each of these additional proteins are needed for efficient editing of some C targets but not others, editosomes that act upon particular C targets differ not only in the site-specific PPR protein recognition factor they contain, but also in which members of these additional families comprise the protein complex. The chloroplast editing factor ORRM1 contains both a RIP domain and an RRM (RNA Recognition Motif) domain [25]. The ORRM1 protein belongs to a distinct clade of RRM-containing proteins, and the RRM domain by itself is able to provide RNA editing activity to orrm1 mutants [25].
In order to identify components of chloroplast editosomes that contain ORRM1, Arabidopsis thaliana orrm1 mutants were complemented with an epitope-tagged ORRM1 protein. A candidate ORRM1-interacting protein, encoded by At5g17790, was identified in immunoprecipitates. Through analysis of mutant and silenced tissue, we demonstrated that the candidate protein is a novel chloroplast editing factor. The protein, which we have named OZ1 (Organelle Zinc finger 1), belongs to the RanBP2 type zinc finger protein family, and is required for editing of 14 sites in chloroplasts and affects editing efficiency of another 16 chloroplast C targets. OZ1 is a member of an Arabidopsis protein family that encodes three additional proteins predicted to be targeted to chloroplasts or mitochondria. Identification of OZ1 as a chloroplast editing factor implicates a previously unsuspected class of zinc finger-containing proteins as potentially involved in RNA editing or other aspects of plant organelle RNA metabolism.
Results
N-terminal tagging of ORRM1 preserves editing activity
Preliminary experiments demonstrated that an epitope tag placed at the C-terminus of ORRM1 disrupted its function (S1 Fig A and B). We therefore produced an ORRM1 expression vector with a RecA transit sequence followed by three tandem FLAG tags fused with a strepII tag (3FS tag) (S1 Fig A and C). This construct, designated, RecA-3FS-mORRM1, resulted in significant increase of editing of matK C640, from 11% to 20%, following transfections of orrm1 protoplasts (S1C Fig). The low complementation level can be largely attributed to the size of the vector used in this assay. While the plasmid harboring RecA-RRM is around 6kb (S1 Fig), the N-terminal tagged ORRM1 is integrated into a binary vector around 14kb, and plasmids over 10 kb are known to exhibit lower transfection efficiency [27].
We investigated whether the epitope-tagged protein (Fig. 1A) could restore editing in transgenic plants obtained by root transformation of orrm1 mutant plants. Transgenic plants of normal phenotype were obtained and RNA was extracted for use in editing assays. Editing extent of matK C640, ndhB C872 and ndhG C50, which exhibit decreased editing in orrm1, was examined by bulk sequencing (Fig. 1B). Editing of all three sites was restored to wild-type level in the RecA-3FS-mORRM1 transgenic plants.

Co-immunoprecipitation of ORRM1 followed by mass spectrometry identifies a candidate interacting protein
Total leaf proteins were used to perform ORRM1 immunoprecipitation (IP). Wild-type Arabidopsis, Columbia ecotype, was included as a negative control for comparison in order to eliminate non-specific binding proteins. We observed that the affinity of the strepII tag on ORRM1 to streptactin resin was poor, which was probably due to its internal position caused by the N-terminal fusion of the FLAG tag (Fig. 1A). Therefore we used only anti-FLAG antibodies for immunoprecipitation.
As is shown in Fig. 2, the anti-FLAG antibody recognizes one band from the transgenic plant samples, but none in the wild-type sample. The unique band’s electrophoretic mobility is slightly slower than that expected for the predicted 42 kD size of the tagged ORRM1, possibly due to post-translational modifications. Anti-FLAG resins retained almost all tagged ORRM1 protein from the extract (Fig. 2A). The elutions from both ORRM1 and negative control were separated by a SDS-PAGE gel and silver stained. The bait, 3FS-mORRM1, is clearly seen in the transgenic plant IP but missing in the Col negative control (Fig. 2B). The immunoprecipitates were subjected to MS/MS mass spectrometry in order to identify ORRM1-binding proteins. The protein encoded by At5g17790 was selected for further investigation because after the ORRM1 peptides, it had the largest number of matches in MS/MS spectra and was not detected in the negative controls. S1 Table describes the peptides detected that resulted in the identification of the At5g17790 as a candidate ORRM1-interacting protein.

Characterization of mutants in At5g17790
At5g17790 contains two tandem C2X10C2 zinc finger domains [28] called RanBP2 type zinc fingers (X2GDWICX2CX3NFARRX2CXRCX2-PRPEX2; pFAM00641), which were characterized in the Ran Binding Protein 2 (RanBP2). Ran is a small GTPase and RanBP2 is a nucleoporin that binds Ran via the zinc finger motifs. This gene previously was identified as mutated in a variegated Ds insertional mutant of Arabidopsis thaliana Landsberg erecta, but the cause of the chloroplast developmental aberration was not determined [28]. We obtained one T-DNA insertional line in A. thaliana ecotype Columbia from ABRC, SAIL_358_H03 (Fig. 3A). In contrast to the mutant in the Landsberg ecotype, the homozygous Columbia mutant showed a uniform yellow phenotype as a young seedling, as shown in Fig. 3B. Subsequent growth on sucrose media result in the appearance of light green, non-variegated leaves (Fig. 3C). These older mutant seedlings could be transferred to soil, where the pale green leaves were able to support autotrophic growth (Fig. 3D). The protein encoded by At5g17790 was given the name OZ1 (Organelle Zinc finger 1).

OZ1 mutation leads to altered editing at most chloroplast sites
RNA from 4-week-old oz1–1 homozygous mutants and the siblings was extracted and the editing extent was examined by bulk sequencing as shown in Fig. 4A. The oz1 mutation causes altered editing of various chloroplast C targets. For example, editing of rpoA C200 and ndhB C872 is completely lost in oz1–1 while editing of rpoB C338 is partially disrupted (Fig. 4A). No obvious effect was observed on psbE C214 editing in the oz1–1 mutant. On the contrary, at rpoC1 C488, the mutant editing level is up-regulated compared to the wild-type (Fig. 4A).

Because poisoned primer extension (PPE) is a more sensitive method to measure editing extent than bulk sequencing [4,7], all chloroplast editing sites were assayed both in oz1 and its siblings using PPE (Fig. 4B and Table 1). The oz1–1 allele is clearly recessive because no significant editing difference is seen between heterozygotes and wild-type plants. Editing of rpoA C200 is 0% in oz1–1 by PPE, which confirmed the result from bulk sequencing. ndhB C746 editing dropped from 97% in wild-type to 68% in the mutant. Editing of rpoC1 C488 increases from 25% in wild type to 58% in the mutant, in agreement with the bulk sequencing data.

The assay data for the complete set of chloroplast sites is shown in Table 1. 14 sites have major loss of editing (>90% decrease in editing) in oz1–1 and 15 other sites have significantly decreased editing (>5%, P<0.05). Although editing defects are massive, editing events on the same transcript are not all affected in the same pattern by oz1, hence the editing defects are unlikely to be a secondary effect caused by some change in a transcript itself. For example, ndhD C2 and ndhD C878 lost over 90% of the wild-type editing extent in oz1–1, but ndhD C383 is not affected at all. On the contrary, the sites recognized by the same PPR protein are largely affected in the same way by oz1–1 mutation. ndhD C878 and ndhB C467 share the same PPR recognition factor CRR28 and both of them lose over 90% editing in oz1–1 (Table 1). Likewise, editing of both ndhB C836 and ndhG C50 (controlled by OTP82) and editing of both rpoA C200 and clpP C559 (recognized by CLB19) are similarly affected. ndhF C290 ndhB C1481 and psbZ C50 are all recognized by OTP84, and in oz1–1, all sites exhibit mild defects in editing (5%-20%) (Table 1).
Given that OZ1 is immunoprecipitated by ORRM1, we also compared the OZ1-dependent sites and ORRM1-dependent sites to examine if these two factors participate in the same editing events. Indeed, editing efficiencies of the 14 OZ1-dependent sites are all severely affected in the orrm1 mutant. Many other sites mildly affected by the oz1–1 mutation are also orrm1-dependent (Table 1). 8 sites are controlled only by OZ1 but not by ORRM1. Taken together, OZ1 is a genuine editing factor for the majority of C targets in chloroplasts.
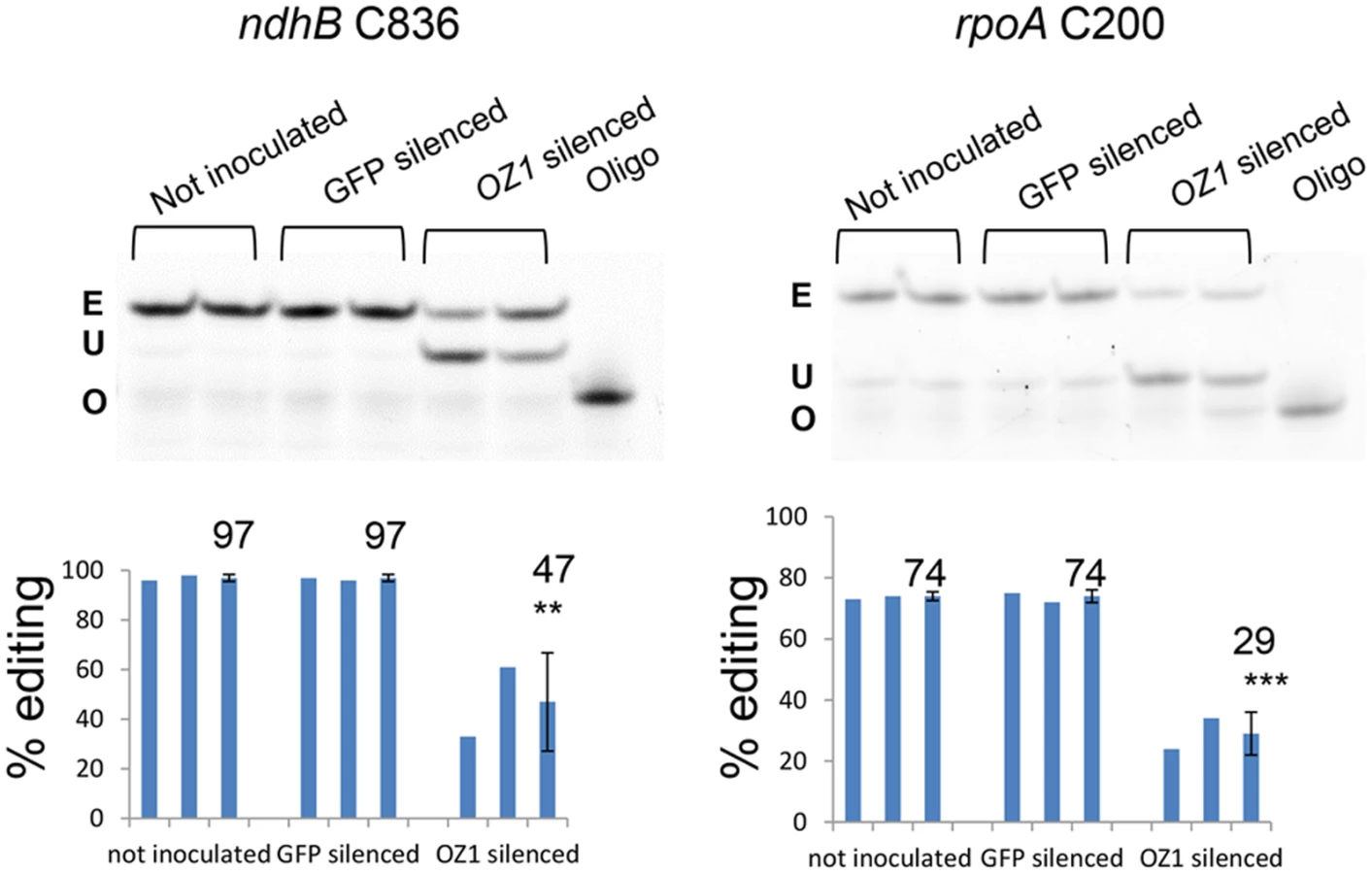
Transient silencing of OZ1 leads to chloroplast editing defects
Since a second T-DNA mutant in the coding region of OZ1 was not available, we performed Virus Induced Gene Silencing (VIGS) to transiently silence OZ1 expression in young Arabidopsis seedlings. To monitor the silencing efficiency, a GFP co-silencing marker harbored in the VIGS construct was used [29]. Agrobacteria carrying either the OZ1/GFP co-silencing construct or the GFP silencing construct alone were inoculated into 2 week-old 35S::GFP expressing Arabidopsis seedlings. After growth in long days for 5 more weeks, the editing extents in RNA from GFP-silenced leaves and from uninoculated plants were analyzed by PPE. There were no differences between leaves of GFP-silenced plants and untreated plants (Fig. 5). ndhB C836 editing extent decreased from 97% in the untreated control to 47% in OZ1 silenced plants (P<0.01). rpoA C200 editing extent dropped from 74% in untreated control to 29% in OZ1 silenced plants (P<0.001). These results agree with the data from oz1–1 mutants, in which editing is abolished at both sites. The residual editing in the silenced plants is probably caused by incomplete depletion of OZ1 protein.
Transient expression of OZ1 in oz1–1 protoplasts complements the editing defects
Although the young oz1–1 mutant has a severely defective phenotype, the plants gradually recover some chlorophyll. In a 6-week - old oz1–1 plant, the old leaves remain pale yellow while the new leaves are light green (Fig. 3C). To investigate whether editing defects are rescued in the light green leaves, editing of RNA extracted from both the pale yellow leaves and the light green leaves were analyzed by bulk sequencing.
Although pigmentation has been partially recovered in light green leaves, plastid editing is still defective in those leaves compared to wild type Col (Fig. 6). No obvious difference in editing between yellow and green leaves was observed. This finding indicates that the defects in editing are not due to a pleiotropic effect caused by some other chloroplast developmental problem in yellow chlorophyll-deficient leaves. Green leaves were therefore used to prepare protoplasts. OZ1 was cloned into a pSAT4a vector to create 35S::OZ1, a plant transient expression vector driven by a 35S promoter for transfections of oz1–1 protoplasts. A chloroplast targeted YFP construct (35S::cpYFP) was included as a negative control. Monitoring of transfection efficiency of the YFP constructs by microscopy indicated expression of YFP in over 50% of the protoplasts.

RNA was extracted from protoplasts two days after the transfection and analyzed by PPE to examine the editing efficiency (Fig. 7). No significant difference in editing was seen between the untransfected control and the 35S::cpYFP-transfected control. Introduction of 35S::OZ1 significantly increases the editing level for all the sites we tested. rpoA C200 increased from 3% to 21%, ndhB C836 from 19% to 31% and rps12-(i1) C58 from 3% to 15%. This confirms that the editing defects in the oz-1–1 mutant can be reduced by introduction of OZ1.

Stable expression of OZ1 in oz1–1 mutant plants also complements the editing defects
Because of the poor growth of the homozygous oz1 mutant plant (Fig. 3D), we decided to transform the heterozygous plant by floral dipping with a construct expressing OZ1 under the control of a 35S promoter. Genotyping the transgenic plants growing on a selectable plate allowed us to recover several independent plants homozygous mutant for the endogenous oz1 alleles but expressing the OZ1 transgene. The introduction of a functional OZ1 complements the editing defect in all the transgenic plants assayed (Fig. 8A). Positional effects on the transgene are known to affect expression and likely resulted in the range of responses in the transformed plants. For example, in different plants, rpoA C200 editing extent ranged from 13%-65% (Fig. 8A). The restoration of editing extent in some transgenic mutant plants is much more pronounced than with the transient expression in the oz1 mutant protoplasts, e.g. 89% vs. 31% for ndhB C836, and reaches almost the level observed in the wild-type plant. In addition to reverting the editing defects, the introduction of OZ1 in planta also suppress the yellow phenotype observed in the mutant plant (Fig. 8B). The reversion of both editing and phenotypic defects by expression of a functional OZ1 demonstrates the role of this protein in both phenomena.

OZ1 interacts with ORRM1
A yeast two-hybrid (Y2H) assay was employed to examine the interaction between OZ1 and ORRM1. Both OZ1 and ORRM1 are plastid-targeted proteins, so the predicted transit peptide sequences were removed from each before cloning them into AD/BD fusion constructs. As shown in Fig. 9A, OZ1 interacts with ORRM1 in yeast. The interaction is not affected by the position of the fusion protein since both AD-OZ1/BD-ORRM1 and its reciprocal pair AD-ORRM1/BD-OZ1 showed interaction, implicating a genuine interaction between these two proteins. ORRM1 was further divided into nORRM1 and cORRM1, encompassing the RIP-RIP and the RRM domain respectively. nORRM1 but not cORRM1 interacts with OZ1, indicating the RIP-RIP domain actually mediates the interaction with OZ1 (Fig. 9B).
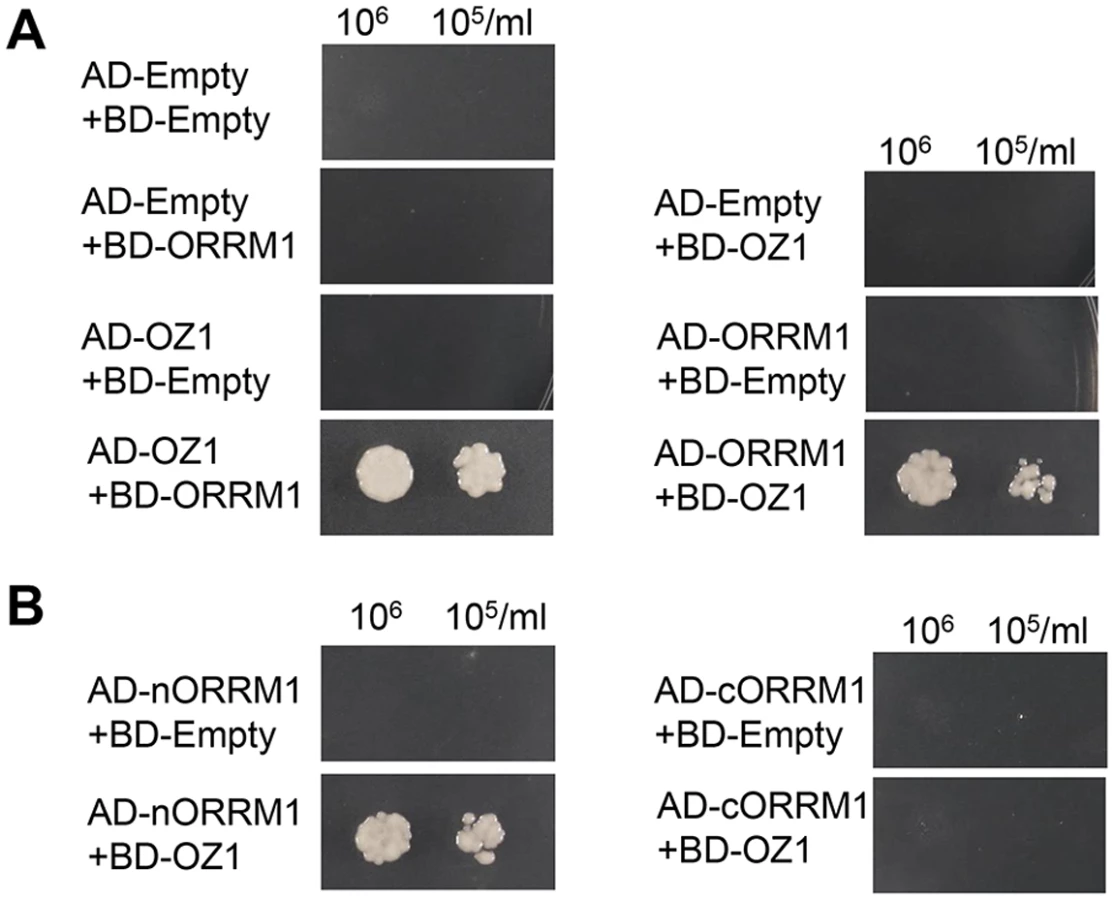
OZ1 interacts with other components of chloroplast editosomes
We suspected that OZ1 might also interact with other components of chloroplast editosomes in addition to ORRM1, such as additional PPR site recognition factors and members of the RIP/MORF protein family. In order to determine whether OZ1 can dimerize and/or interact with other components of the editing complex, we performed a series of Y2H assays. OZ1 fused to either AD or BD does not show any auto-activation for HIS and ADE reporters, while yeast with AD-OZ1/BD-OZ1 is able to grow on histidine and adenine deficient media, indicating self-interaction (Fig. 10A). OZ1 also interacts with OTP82 and CRR28, as shown in Fig. 10B, a result expected from the effect of the oz1–1 mutation on C targets controlled by OTP82 and CRR28 (Table 1). OZ1 exhibits a weaker interaction with RIP1; fewer colonies are seen in the RIP1/OZ1 combination (Fig. 10B). However, no interaction was observed between OZ1 and RIP2 or RIP9 (Fig. 10B), even though RIP2 and RIP9 are essential for editing of a large number of chloroplast C targets. We considered the possibility that OZ1 associates with RIP2 and RIP9 via ORRM1. To test this hypothesis, we performed a Y2H assay for ORRM1 and RIP proteins (Fig. 10C). Both RIP1 and RIP2 can interact with ORRM1. RIP9 fused with the GAL4 binding domain strongly autoactivates HIS and ADE reporters, so RIP9 was not tested in this experiment. ORRM1 with the GAL4 activation domain shows no autoactivation (Fig. 9A). Our data is consistent with ORRM1 as a mediator of interaction between OZ1 and RIP2 within the chloroplast editosome.

OZ1 belongs to a small family in Arabidopsis
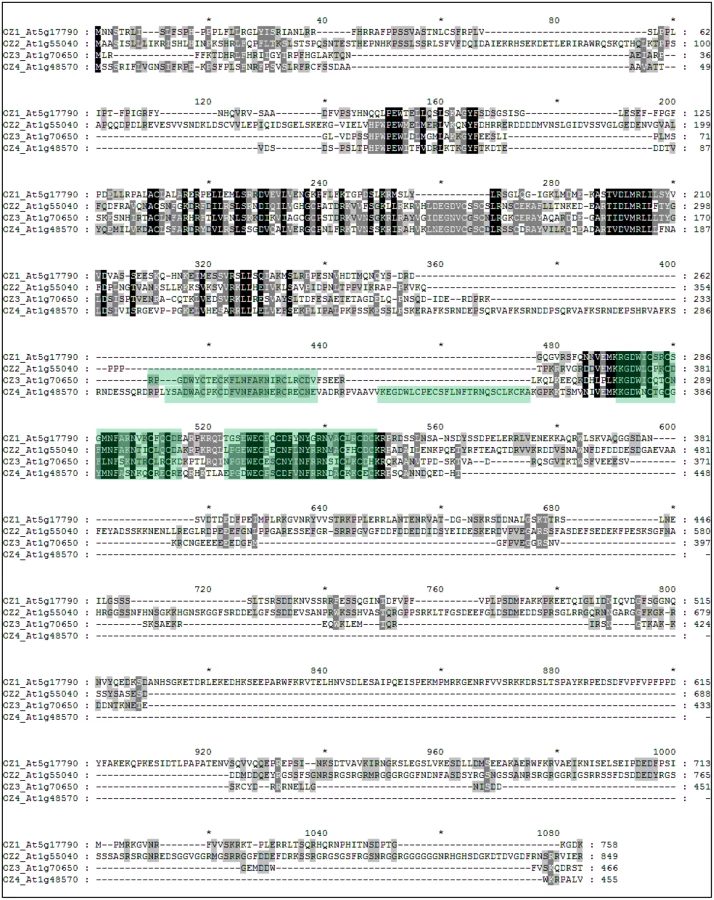
Three highly similar RanBP2 zinc finger proteins were found in Arabidopsis protein database in a BLAST search with OZ1. In 2004, these proteins were reported to comprise a four-member gene family of unknown function [28]; however, changes in gene models result in a new alignment (Fig. 11). The protein sequence alignment by T-coffee shows presence of multiple highly conserved regions in the N-terminal portion of the protein, past the predicted transit sequences, with various numbers of zinc finger motifs and more variable C-terminal regions (Fig. 11).
In order to investigate the subcellular location of OZ1, we fused the N-terminal sequences encoding 100 amino acids to yellow fluorescent protein (YFP) and transfected protoplasts. Confocal microscopic imaging revealed that OZ1-YFP is located in the chloroplasts at punctate loci (Fig. 12). Previously, the entire coding region of OZ1 (VAR3) was fused to GFP and observed to be located within chloroplasts at punctate loci [28]. OZ1 was also detected in chloroplast nucleoid preparations by mass spectrometry (ppdb.tc.cornell.edu). In addition, Target P predicts all of the other OZ1 family members to be organelle targeted [30], one in mitochondria and two in plastids (S2 Table). Proteomics studies have also found OZ4 (At1g48570) in both chloroplast nucleoids and stroma (ppdb.ts.cornell.edu).

Except for the zinc finger motif, no other annotated domain or motif was found in the OZ1 family. In order to find hidden uncharacterized motifs, motif scanning was performed using MEME against all four members to look for motifs between 15aa to 70aa. Five motifs were returned (Fig. 13). The zinc finger domain has 4 characteristic cysteine residues. As shown in Fig. 13, the zinc finger motif is shared by all four members, but the number of repeats varies. OZ1 and OZ2 (At1g55040) contain two zinc finger motifs, while OZ3 (At1g70650) has three and OZ4 (At1g48570) has four. The regions preceding the zinc finger motifs are relatively highly conserved, briefly spanning 3 distinct domains. The region downstream of the zinc finger domains is quite variable. OZ1 has three repeats of motif 5, which is either missing or poorly conserved in the other members (S2 Fig). Portions of motif 5 were previously identified as three “long repeats” in At5g17790 [28].

We performed homology searches to determine whether orthologs of the four Arabidopsis OZ family members could be detected in other well-characterized plant genomes. We were able to identify putative orthologs for all 4 genes in poplar, grape, rice, and maize. Moss and Selaginella, which exhibit chloroplast RNA editing, also encode OZ-like proteins but with a lower similarity (Fig. 14). We could not detect proteins similar to the OZ family in Chlamydomonas or Volvox, where editing does not occur.

Discussion
Because a second coding region mutant in OZ1 was not available, we performed transient silencing, transient complementation, and stable complementation to verify the function of OZ1 in editing. Given that VIGS can only knock down gene expression, the editing level of the OZ1-dependent sites were reduced but not totally abolished. Introduction of a 35S::OZ1 construct into mutant protoplasts or into transgenic plants greatly increased the editing extents of the editing defective sites, demonstrating that the editing defects seen in the oz1 mutant can be attributed to loss of OZ1. The absence of OZ1 results in reduced editing efficiency at most of the affected C targets, but editing of rpoC1 C488 is increased. Possibly, the loss of OZ1 results in reduction of sequestration of an editing factor needed for rpoC1 C488 that is present in limiting amounts when OZ1 is present. If OZ1 is not needed for editing of rpoC1 C488, its loss could make available more of an unknown editing factor needed for efficient rpoC1 C488 editing.
Although 14 chloroplast C targets have major loss of editing with nine sites showing no detectable editing by PPE, and editing efficiencies of 16 other editing sites are significantly altered, the oz1 mutant can survive on sucrose as a yellow seedling and then undergo sufficient chloroplast development to support autotrophic growth. Even in the green, fully photosynthetic leaves, the editing defects are still observed. Most of the sites at which editing is abolished in the oz1–1 mutant are in non-coding regions or in NADH dehydrogenase genes that are not needed in low light growth chamber conditions. The virescent phenotype could be largely due to the complete loss of editing of rpoA C200. Absence of editing at this site also occurs when the PPR editing factor gene CLB19 is mutated [15], and the phenotype of oz1–1 is similar to the clb19 mutant. rpoA encodes a subunit of the plastid-encoded RNA polymerase (PEP); loss of editing at this particular site results in defective PEP and yellow seedling phenotype in early developmental stages. However, mutants survive defects in PEP and later partially recover because of the presence of the nuclear-encoded plastid polymerase that can remedy some of the impaired gene expression. The phenotype of oz1–1 differs from the previously characterized Ds insertion mutant var3 in At5g17790, which was found to exhibit variegated leaves [28].
The action of OZ1 is clearly site-specific, because C targets that reside on the same transcript are differently affected. For example, ndhD C878 has a major loss of editing while ndhD C383 is barely affected. Which editing sites are affected is likely determined by the editing factors with which OZ1 interacts. C targets that share PPR recognition factors are similarly affected in the oz1 mutant. In Y2H assays, OZ1 binds to CRR28 and OTP82, PPR proteins that are required for editing of sites that also require OZ1. Furthermore, OZ1 interacts with ORRM1, and all 14 severely affected chloroplast sites are also affected in the orrm1 mutant. OZ1 interacts with RIP1, though the interaction is not as strong as that with ORRM1. We did not observe direct interaction of OZ1 with RIP2 or RIP9. However, ORRM1 can bind to RIP1 and RIP2. Previously we reported interaction between the RIP-RIP domain of ORRM1 and CRR28 and OTP82 [25]. Another group determined that RIP2 and RIP9 interact with CRR28 [31]. RIP2 and RIP9 have been reported to interact with PPO1, protoporphyrinogen IX oxidase 1, which is required for efficient editing of a number of chloroplast sites [31]. However, PPO1 does not interact with CRR28 or other PPR editing factors, and it is presently unknown whether PPO1 also interacts with either ORRM1 or OZ1 [31]. All of the interaction data, taken together, is consistent with the presence of multi-component editing complexes that contain ORRM1, OZ1, and at least one RIP protein and a PPR protein, at unknown stoichiometry. An example of the model for the editosome acting upon ndhD C878, drawn according to the yeast two-hybrid data, is shown in Fig. 15. Some complexes are also likely to contain PPO1, but we cannot place this protein into our diagram until its interaction with ORRM1 and OZ1 is investigated in the future.

Three proteins that share high similarity with OZ1 are also predicted to be targeted to organelles according to Target P (S2 Table). OZ1 contains three long repeats at its C terminus; these domains of unknown function are less conserved in other family members. The only well-documented and most significant domain found in this family is the Ran binding protein 2 type zinc finger motif, which is a conserved 30-amino-acid consensus (X2GDWICX2CX3NFARRX2CXRCX2-PRPEX2; pFAM00641) characterized in RAN binding protein 2 (RanBP2) and other nucleoporins. OZ1 contains two tandem RanBP2 zinc-finger domains while other members have various number of this domain. It is highly possible that other members of the OZ family have similar role in chloroplast or mitochondrial RNA editing. Such redundancy could explain the residual editing for some sites in the oz1 mutant Chloroplast RNA editing in vitro has been shown to be Zn2+ dependent [32]. Zinc binding is characteristic of cytidine deaminases, and the DYW domain, which contains cytidine deaminase motifs and is present on a subset of PPR protein editing factors, has been shown to bind zinc ions [16,23]. The requirement for zinc in plant organelle RNA editing has been thought to be due to the need for cytidine deaminase activity. The discovery of OZ1 implicates the OZ family as another possible source of the zinc requirement for editing to occur. Further experiments will be needed to determine whether the zinc fingers present in the OZ family actually bind zinc and whether they are important in RNA and/or protein binding in the RNA editosome.
Material and Methods
Mutant lines and phenotyping
The T-DNA insertional A. thaliana Columbia ecotype mutant SAIL_358_H03 in the OZ1 gene was obtained from the Arabidopsis Biological Resource Center (https://abrc.osu.edu/). After 3 days of stratification, the seeds were placed onto petri dishes containing Murashige-Skoog medium in a 25o room with 14 hour day length. Mutant plants and the wild-type siblings were then transferred into soil after 7 weeks on tissue culture medium. Leaves from 4 week - old and 8 week - old plants were collected for further analysis.
Plasmid constructs and oligonucleotides
The ORRM1 coding sequence was cloned using primer pair ORRM1_1F and ORRM1_R_WO from previous constructs. The sequence was first cloned into PCR8/GW/TOPO (Life Technologies, Carlsbad, CA), and then shuttled into a modified PBI121 vector with a 3XFLAG-strepII C-terminal tag. Alternatively, coding sequence of the ORRM1 mature form (without the predicted 54aa transit peptide) was fused to a N terminal 3XFLAG-strepII tag sequence and an artificial transit peptide sequence from RecA in an overlapping PCR using primer pairs RecA_F, RecA_R, 3FS_F, 3FS_R, ORRM1_163F and ORRM1_R. This chimeric gene was cloned into PCR8 vector first and then the PBI121 vector using LR ClonaseII (Life Technologies, Carlsbad, CA). The C-terminal tag on the vector was eliminated by the endogenous stop codon of ORRM1 in the sequence. A RecA-RRM construct was described previously. The OZ1 coding sequence was cloned using primer pair OZ1_F and OZ1_R from A. thaliana cDNA. The PCR product was first ligated to PCR8/GW/TOPO and then transferred into the destination vectors pSAT4a and pAUL13 [33] to create a transient expression construct and a stable complementation construct, respectively.
All oligonucleotides used in this study are shown in S3 Table.
Protoplast complementation
Protoplasts from orrm1 or oz1 mutants were prepared following the protocol from Jen Sheen’s lab [27]. Light green leaves from the oz1 mutant were used for protoplast preparation. 10 μg of plasmid DNA was used to transfect 2x104 cells. The transfected protoplasts were incubated in the dark at room temperature for 3 days (orrm1 complementation) or 1 day (oz1 complementation) before harvest.
Virus-induced gene silencing
An OZ1 gene-specific region was selected from the CATMA database [34] and amplified using primer pair OZ1_VIGS_F and OZ1_VIGS_R. The fragment was first integrated into PCR8/GW/TOPO and then into the silencing vector PTRV2/GW/GFP by an LR reaction. Agrobacteria harboring the silencing construct were used to infiltrate 2 week-old Arabidopsis seedlings that expressed GFP driven by 35S promoter as previously described [7]. 5 weeks after infiltration, silencing efficiency was monitored by the expression of the co-silenced GFP in each individual. Silenced plants, which exhibited a dark red color from stem to leaf under UV light, were collected for further analysis.
Generation of transgenic plants
The 3XFLAG-strepII-ORRM1ΔCTP and OZ1 constructs in PBI121 and PAUL13, respectively, were used to transform the Agrobacterium GV3101 strain. A standard root transformation protocol was followed to transform mutant orrm1 roots. The roots were first induced on Callus Inducing Medium (CIM) for 2 days and then infected with Agrobacteria in liquid media [35]. Roots were incubated on CIM for another 2 days until they were overgrown by Agrobacteria and then bacteria were removed by several washing steps with liquid CIM containing timentin and carbenicillin. Roots were then cut into 0.5mm pieces and put onto Shoot Inducing Medium (SIM) containing 100mg/L Basta for selection. After the shoots grew out, they were removed from the calli and transferred onto a Root Inducing Medium (RIM) [35]. Fully grown transgenic plants with healthy roots were then transferred into soil. Because the root transformation was not successful with the oz1 mutant, a standard floral dip protocol was used to transform a heterozygous plant.
Co-immunoprecipitation
10 g of leaves of each line were ground in liquid nitrogen into fine powder. Total leaf protein was extracted using grinding buffer (150mM NaCl, 50mM Tris-HCl pH7.4, 1mM EDTA, 0.2%NP-40 and 1x cocktail protease inhibitor (Sigma-Aldrich, St. Louis, MO). The extract was cleared by 13,000 rpm centrifugation, 0.45 μm filtration and then 30 minutes incubation with unconjugated agarose beads (Vector Laboratories, Burlingame, CA) to minimize non-specific binding. 200 μl anti-FLAG agarose resins (Sigma-Aldrich, St. Louis, MO) were first blocked with 4%BSA before 2 hours incubation with around 30 μg pre-cleared protein extract. A washing step was done using washing buffer (150mM NaCl, 50mM Tris-HCl pH7.4, 1mM EDTA, 0.2% NP-40). The IP was eluted with elution buffer (2M MgCl2, 50mM Tris-HCl ph8.0, 150mM NaCl, 0.5%CHAPS). The final sample was prepared using SDS-PAGE sample prep kit (Thermo Scientific, Waltham, MA).
Immunoblotting, silver staining and mass spectrometry
1% of the IP samples were loaded onto an Any kD MINI-PROTEAN TGX precast gel (Bio-Rad, Hercules, CA) followed by standard procedures of immunoblotting. α-FLAG-M2-HRP (Sigma-Aldrich, St. Louis, MO) was used to detect FLAG-tagged proteins. 10% of the IP samples were separated on a 10% polyacrylamide gel before being subjected to silver staining compatible to mass spectrometry. Co-purifying proteins with FLAG-tagged ORMM1 were identified by tandem mass spectrometry using a nanoLC-LTQ-Orbitrap instrument, followed by database searching with MASCOT against TAIR10 [36].
RNA editing extent measurement
DNA contaminants were removed from RNA samples by TURBO DNase (Life Technologies, Carlsbad, CA). The cDNA was reverse transcribed from the RNAs using the pooled reverse primers as previously described [7]. PCR products harboring the editing sites were either bulk Sanger-sequenced or subjected to PPE assay [29,37].
Yeast two-hybrid assay
Mature OZ1 coding sequence (without the N-terminal predicted 33 aa transit peptide) was amplified using primer pair OZ1_100F and OZ1_R from cDNA and cloned into PCR8/GW/TOPO (Life Technologies, Carlsbad, CA). Mature RIP2 and RIP9 coding sequences were amplified using primer pairs RIP2_133F and RIP2_R, RIP9_175F and RIP9_R from A. thaliana cDNA, respectively. PCR products were first cloned into PCR8/GW/TOPO and then pGADT7GW and pGBKT7GW destination vectors through homologous recombination by LR clonaseII (Life Technologies, Carlsbad, CA). RIP1, RARE1, CRR28, OTP82, ORRM1 constructs produced for Y2H assays were previously described [7]. Empty vectors were used as negative controls. Two mating types of the PJ69–4 yeast strain, a and α, were used. Single transformants were obtained by transformation while double transformants were produced through mating. Yeast harboring testing pairs were grown in leucine and tryptophan deficient media overnight before they were diluted with water to OD600 0.5, 0.05, or 0.005. 10 μl of each dilution was spotted onto leucine-, tryptophan-, histidine-, adenine-deficient media plates. Growth results were collected after 3 days incubation at 30°C.
Subcellular localization in protoplasts
The first 100 codons of OZ1 was amplified from cDNA with primers listed in S3 Table, and cloned into PCR8/GW/TOPO (Life Technologies, Carlsbad, CA). The fragment was subsequently transferred to the pEXSG-YFP Gateway destination vector [38] by recombination using LR Clonase II (Life Technologies, Carlsbad, CA), creating a gene encoding a YFP fusion protein driven by a tandem CaMV 35S promoter. Protoplasts were prepared from leaves of 3-week old Arabidopsis accession Ler and transfected as described above. Images were acquired 3 days after transfection using a Zeiss 710 confocal microscope at the Cornell Biotechnology Resource Center (BRC).
Supporting Information
{kind=link}
{kind=link}
Zdroje
1. Covello PS, Gray MW (1989) RNA editing in plant mitochondria. Nature 341 : 662–666. 2552326
2. Gualberto JM, Lamattina L, Bonnard G, Weil J-H, Grienenberger J-M (1989) RNA editing in wheat mitochondria results in the conservation of protein sequences. Nature 341 : 660–662. 2552325
3. Hiesel R, Wissinger B, Schuster W, Brennicke A (1989) RNA editing in plant mitochondria. Science 246 : 1632–1634. 2480644
4. Bentolila S, Oh J, Hanson MR, Bukowski R (2013) Comprehensive high-resolution analysis of the role of an Arabidopsis gene family in RNA editing. PLoS Genet 9: e1003584. doi: 10.1371/journal.pgen.1003584 23818871
5. Chateigner-Boutin A-L, Small I (2007) A rapid high-throughput method for the detection and quantification of RNA editing based on high-resolution melting of amplicons. Nucleic Acids Res 35: e114. 17726051
6. Gray MW, Covello PS (1993) RNA editing in plant mitochondria and chloroplasts. FASEB J 7 : 64–71. 8422976
7. Bentolila S, Heller WP, Sun T, Babina AM, Friso G, et al. (2012) RIP1, a member of an Arabidopsis protein family, interacts with the protein RARE1 and broadly affects RNA editing. Proc Natl Acad Sci USA 109: E1453–E1461. doi: 10.1073/pnas.1121465109 22566615
8. Chaudhuri S, Maliga P (1996) Sequences directing C to U editing of the plastid psbL mRNA are located within a 22 nucleotide segment spanning the editing site. EMBO J 15 : 5958–5964. 8918473
9. Hayes ML, Hanson MR (2007) Identification of a sequence motif critical for editing of a tobacco chloroplast transcript. RNA 13 : 281–288. 17158709
10. Okuda K, Nakamura T, Sugita M, Shimizu T, Shikanai T (2006) A pentatricopeptide repeat protein is a site recognition factor in chloroplast RNA editing. J Biol Chem 281 : 37661–37667. 17015439
11. Barkan A, Rojas M, Fujii S, Yap A, Chong YS, et al. (2012) A combinatorial amino acid code for RNA recognition by pentatricopeptide repeat proteins. PLoS Genet 8: e1002910. doi: 10.1371/journal.pgen.1002910 22916040
12. Yin P, Li Q, Yan C, Liu Y, Liu J, et al. (2013) Structural basis for the modular recognition of single-stranded RNA by PPR proteins. Nature 504 : 168–171. doi: 10.1038/nature12651 24162847
13. Lurin C, Andrés C, Aubourg S, Bellaoui M, Bitton F, et al. (2004) Genome-wide analysis of Arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell 16 : 2089–2103. 15269332
14. Salone V, Rüdinger M, Polsakiewicz M, Hoffmann B, Groth-Malonek M, et al. (2007) A hypothesis on the identification of the editing enzyme in plant organelles. FEBS Lett 581 : 4132–4138. 17707818
15. Chateigner-Boutin A-L, Ramos-Vega M, Guevara-García A, Andrés C, De La Luz Gutiérrez-Nava M, et al. (2008) CLB19, a pentatricopeptide repeat protein required for editing of rpoA and clpP chloroplast transcripts. Plant J 56 : 590–602. doi: 10.1111/j.1365-313X.2008.03634.x 18657233
16. Hayes ML, Giang K, Berhane B, Mulligan RM (2013) Identification of two pentatricopeptide repeat genes required for RNA editing and zinc binding by C-terminal cytidine deaminase-like domains. J Biol Chem 288 : 36519–36529. doi: 10.1074/jbc.M113.485755 24194514
17. Kotera E, Tasaka M, Shikanai T (2005) A pentatricopeptide repeat protein is essential for RNA editing in chloroplasts. Nature 433 : 326–330. 15662426
18. Okuda K, Myouga F, Motohashi R, Shinozaki K, Shikanai T (2007) Conserved domain structure of pentatricopeptide repeat proteins involved in chloroplast RNA editing. Proc Natl Acad Sci USA 104 : 8178–8183. 17483454
19. Okuda K, Chateigner-Boutin A-L, Nakamura T, Delannoy E, Sugita M, et al. (2009) Pentatricopeptide repeat proteins with the DYW motif have distinct molecular functions in RNA editing and RNA cleavage in Arabidopsis chloroplasts. Plant Cell 21 : 146–156. doi: 10.1105/tpc.108.064667 19182104
20. Okuda K, Hammani K, Tanz SK, Peng L, Fukao Y, et al. (2010) The pentatricopeptide repeat protein OTP82 is required for RNA editing of plastid ndhB and ndhG transcripts. Plant J Cell Mol Biol 61 : 339–349.
21. Nakamura T, Sugita M (2008) A conserved DYW domain of the pentatricopeptide repeat protein possesses a novel endoribonuclease activity. FEBS Lett 582 : 4163–4168. doi: 10.1016/j.febslet.2008.11.017 19041647
22. Boussardon C, Salone V, Avon A, Berthomé R, Hammani K, et al. (2012) Two interacting proteins are necessary for the editing of the ndhD-1 site in Arabidopsis plastids. Plant Cell 24 : 3684–3694. doi: 10.1105/tpc.112.099507 23001034
23. Boussardon C, Avon A, Kindgren P, Bond CS, Challenor M, et al. (2014) The cytidine deaminase signature HxE(x)n CxxC of DYW1 binds zinc and is necessary for RNA editing of ndhD-1. New Phytol 203 : 1090–1095. doi: 10.1111/nph.12928 25041347
24. Wagoner JA, Sun T, Lin L, Hanson MR. (2014) Cytidine deaminase motifs within the DYW domain of two pentatricopeptide repeat-containing proteins are required for site-specific chloroplast RNA editing. J Biol Chem.
25. Sun T, Germain A, Giloteaux L, Hammani K, Barkan A, et al. (2013) An RNA recognition motif-containing protein is required for plastid RNA editing in Arabidopsis and maize. Proc Natl Acad Sci USA 110: E1169–E1178. doi: 10.1073/pnas.1220162110 23487777
26. Takenaka M, Zehrmann A, Verbitskiy D, Kugelmann M, Härtel B, et al. (2012) Multiple organellar RNA editing factor (MORF) family proteins are required for RNA editing in mitochondria and plastids of plants. Proc Natl Acad Sci USA 109 : 5104–5109. doi: 10.1073/pnas.1202452109 22411807
27. Yoo S-D, Cho Y-H, Sheen J (2007) Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat Protoc 2 : 1565–1572. 17585298
28. Næsted H, Holm A, Jenkins T, Nielsen HB, Harris CA, et al. (2004) Arabidopsis VARIEGATED 3 encodes a chloroplast-targeted, zinc-finger protein required for chloroplast and palisade cell development. J Cell Sci 117 : 4807–4818. 15340011
29. Robbins JC, Heller WP, Hanson MR (2009) A comparative genomics approach identifies a PPR-DYW protein that is essential for C-to-U editing of the Arabidopsis chloroplast accD transcript. RNA 15 : 1142–1153. doi: 10.1261/rna.1533909 19395655
30. Emanuelsson O, Nielsen H, Brunak S, von Heijne G (2000) Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J Mol Biol 300 : 1005–1016. 10891285
31. Zhang F, Tang W, Hedtke B, Zhong L, Liu L, et al. (2014) Tetrapyrrole biosynthetic enzyme protoporphyrinogen IX oxidase 1 is required for plastid RNA editing. Proc Natl Acad Sci USA 111 : 2023–2028. doi: 10.1073/pnas.1316183111 24497494
32. Hegeman CE, Hayes ML, Hanson MR (2005) Substrate and cofactor requirements for RNA editing of chloroplast transcripts in Arabidopsis in vitro. Plant J 42 : 124–132. 15773858
33. Lyska D, Engelmann K, Meierhoff K, Westhoff P (2013) pAUL: a gateway-based vector system for adaptive expression and flexible tagging of proteins in Arabidopsis. PloS One 8: e53787. doi: 10.1371/journal.pone.0053787 23326506
34. Crowe ML, Serizet C, Thareau V, Aubourg S, Rouzé P, et al. (2003) CATMA: a complete Arabidopsis GST database. Nucleic Acids Res 31 : 156–158. 12519971
35. Weigel D, Glazebrook J (2006) Root transformation of Arabidopsis. Cold Spring Harb Protoc 2006: pdb.prot4671.
36. Friso G, Olinares PDB, van Wijk KJ (2011) The workflow for quantitative proteome analysis of chloroplast development and differentiation, chloroplast mutants, and protein interactions by spectral counting. Methods Mol Biol 775 : 265–282. doi: 10.1007/978-1-61779-237-3_14 21863448
37. Peeters NM, Hanson MR (2002) Transcript abundance supercedes editing efficiency as a factor in developmental variation of chloroplast gene expression. RNA 8 : 497–511. 11991643
38. Jakoby MJ, Weinl C, Pusch S, Kuijt SJH, Merkle T, et al. (2006) Analysis of the subcellular localization, function, and proteolytic control of the Arabidopsis cyclin-dependent kinase inhibitor ICK1/KRP1. Plant Physiol 141 : 1293–1305. 16766674
39. Notredame C, Higgins DG, Heringa J (2000) T-Coffee: A novel method for fast and accurate multiple sequence alignment. J Mol Biol 302 : 205–217. 10964570
40. Yagi Y, Tachikawa M, Noguchi H, Satoh S, Obokata J, Nakamura T (2013) Pentatricopeptide repeat proteins involved in plant organellar RNA editing. RNA Biol. 10 : 1419–1425. doi: 10.4161/rna.24908 23669716
41. Yap A, Kindgren P, Colas des Francs-Small C, Kazama T, Tanz SK, et al. (2015) AEF1/MPR25 is implicated in RNA editing of plastid atpF and mitochondrial nad5 and also promotes atpF splicing in Arabidopsis and rice. Plant J.
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2015 Číslo 3
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Clonality and Evolutionary History of Rhabdomyosarcoma
- Morphological Mutations: Lessons from the Cockscomb
- Maternal Filaggrin Mutations Increase the Risk of Atopic Dermatitis in Children: An Effect Independent of Mutation Inheritance
- Transcriptomic Profiling of Reveals Reprogramming of the Crp Regulon by Temperature and Uncovers Crp as a Master Regulator of Small RNAs
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy