
Sex Chromosome Mosaicism and Hybrid Speciation among Tiger Swallowtail Butterflies
Hybrid speciation, or the formation of a daughter species due to interbreeding between two parental species, is a potentially important means of diversification, because it generates new forms from existing variation. However, factors responsible for the origin and maintenance of hybrid species are largely unknown. Here we show that the North American butterfly Papilio appalachiensis is a hybrid species, with genomic admixture from Papilio glaucus and Papilio canadensis. Papilio appalachiensis has a mosaic phenotype, which is hypothesized to be the result of combining sex-linked traits from P. glaucus and P. canadensis. We show that P. appalachiensis' Z-linked genes associated with a cooler thermal habitat were inherited from P. canadensis, whereas its W-linked mimicry and mitochondrial DNA were inherited from P. glaucus. Furthermore, genome-wide AFLP markers showed nearly equal contributions from each parental species in the origin of P. appalachiensis, indicating that it formed from a burst of hybridization between the parental species, with little subsequent backcrossing. However, analyses of genetic differentiation, clustering, and polymorphism based on molecular data also showed that P. appalachiensis is genetically distinct from both parental species. Population genetic simulations revealed P. appalachiensis to be much younger than the parental species, with unidirectional gene flow from P. glaucus and P. canadensis into P. appalachiensis. Finally, phylogenetic analyses, combined with ancestral state reconstruction, showed that the two traits that define P. appalachiensis' mosaic phenotype, obligatory pupal diapause and mimicry, evolved uniquely in P. canadensis and P. glaucus, respectively, and were then recombined through hybridization to form P. appalachiensis. These results suggest that natural selection and sex-linked traits may have played an important role in the origin and maintenance of P. appalachiensis as a hybrid species. In particular, ecological barriers associated with a steep thermal cline appear to maintain the distinct, mosaic genome of P. appalachiensis despite contact and occasional hybridization with both parental species.
Published in the journal:
. PLoS Genet 7(9): e32767. doi:10.1371/journal.pgen.1002274
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002274
Summary
Hybrid speciation, or the formation of a daughter species due to interbreeding between two parental species, is a potentially important means of diversification, because it generates new forms from existing variation. However, factors responsible for the origin and maintenance of hybrid species are largely unknown. Here we show that the North American butterfly Papilio appalachiensis is a hybrid species, with genomic admixture from Papilio glaucus and Papilio canadensis. Papilio appalachiensis has a mosaic phenotype, which is hypothesized to be the result of combining sex-linked traits from P. glaucus and P. canadensis. We show that P. appalachiensis' Z-linked genes associated with a cooler thermal habitat were inherited from P. canadensis, whereas its W-linked mimicry and mitochondrial DNA were inherited from P. glaucus. Furthermore, genome-wide AFLP markers showed nearly equal contributions from each parental species in the origin of P. appalachiensis, indicating that it formed from a burst of hybridization between the parental species, with little subsequent backcrossing. However, analyses of genetic differentiation, clustering, and polymorphism based on molecular data also showed that P. appalachiensis is genetically distinct from both parental species. Population genetic simulations revealed P. appalachiensis to be much younger than the parental species, with unidirectional gene flow from P. glaucus and P. canadensis into P. appalachiensis. Finally, phylogenetic analyses, combined with ancestral state reconstruction, showed that the two traits that define P. appalachiensis' mosaic phenotype, obligatory pupal diapause and mimicry, evolved uniquely in P. canadensis and P. glaucus, respectively, and were then recombined through hybridization to form P. appalachiensis. These results suggest that natural selection and sex-linked traits may have played an important role in the origin and maintenance of P. appalachiensis as a hybrid species. In particular, ecological barriers associated with a steep thermal cline appear to maintain the distinct, mosaic genome of P. appalachiensis despite contact and occasional hybridization with both parental species.
Introduction
Inter-specific hybridization is widespread in nature and may have important consequences in evolution, from the transfer of adaptive alleles between species to the formation of hybrid species [1]–[8]. Although prevalent in plants, hybrid speciation is apparently uncommon among animals, which may be explained by two factors: (a) hybrid populations may be weakly reproductively isolated from the parental species, so instead of maintaining their genomic identity, they may fuse with one of the parental species by backcrossing, and (b) hybrids may be unable to compete for ecological niches already occupied by parental species [3]–[5]. Hence, hybrid populations are likely to evolve as a species only when hybrids mate assortatively with other hybrids [7], and/or when they adapt to a new environment [6]. Both processes involve selection on specific phenotypic and ecological traits that favor the origin and maintenance of hybrid species. Selective introgression of genes responsible for divergent ecological traits from both parental species into a hybrid species would provide strong evidence for natural selection in the origin of a hybrid species [9]. However, selective regimes that may favor the formation of hybrid species are poorly understood and genetic evidence for selective introgression of ecologically important traits in a hybrid species is limited [9], [10]. For instance, several putative examples of animal hybrid species are known, ranging from fish to crustaceans and insects [11], but the potential role of natural selection in their origin and maintenance is largely unknown. Also, there are no examples of hybrid species that maintain their genomic identity while in contact with both parental species, which is important because sympatry may suggest that hybrid species are maintained by natural selection. Indeed, other known or suspected hybrid species are allopatric relative to either one or both parental species [6], [7], [11]. Here we show that the tiger swallowtail butterfly Papilio appalachiensis exhibits many hallmarks of a hybrid species, including a mosaic genome derived from P. glaucus and P. canadensis. At the same time, its genome is significantly differentiated while being in contact with both its parental species. We further show that its hybrid phenotype, which appears to be under selection, was produced by combining sex chromosomes of its parental species. Thus, Papilio appalachiensis may be an informative case for understanding the role of natural selection in both the origin and maintenance of hybrid species.
Tiger swallowtail butterflies of North America (Figure 1A) are a monophyletic group consisting of eight closely-related species. Sister species glaucus and canadensis show clear and strong evidence for interspecific divergence and multiple forms of reproductive isolation: one-way assortative mate preference [12], [13], reduced hatching success of hybrid eggs [13], [14], Haldane's Rule and hybrid incompatibility in glaucus/canadensis crosses [14], [15], divergent thermal habitat preference [16]–[18], differential host-plant preference/usage [18]–[22] and larval development times [23], and differential survival on preferred host plants [14], [19], [24] (also see Materials and Methods). This broad range of isolating mechanisms shows that glaucus and canadensis are good biological species [13], [25]. They have diverged ecologically along a steep thermal gradient in spite of hybridizing across a long and narrow hybrid zone [25]–[27]. This hybrid zone is characteristically bimodal, where hybrids are uncommon relative to the parental species [28] (also see Materials and Methods). The hybrid zone is maintained by thermal ecology: glaucus is adapted to a warmer thermal habitat compared to canadensis. This has shaped key ecological differences between the two species, including voltinism and pupal diapause (Figure 1B). These traits are Z-linked (Lepidoptera have ZZ males and ZW females) [15], [21], [29]. The thermal landscape also contributes indirectly to the evolution of Batesian mimicry, which is under frequency-dependent selection and produces sexual dimorphism and female dimorphism in glaucus [30], [31]. Papilio glaucus is palatable to vertebrate predators, and has two female forms: a yellow, male-like, non-mimetic form and a melanic form that mimics the chemically defended Battus philenor (Figure 1A) [32]. The distribution of B. philenor is limited in the north by its larval host plant, producing a latitudinal gradient in the frequency of the glaucus mimetic female form [30], [33], [34]. The Mendelian locus controlling female color in glaucus is W-linked, with a Z-linked mimicry enabler allele nearly fixed in glaucus and a mimicry suppressor allele fixed in canadensis [26], [29].

Recently described P. appalachiensis appears to be a phenotypic mixture of glaucus and canadensis, with which it is sympatric and parapatric, respectively (Figure 1A), but is apparently reproductively isolated from both (see Materials and Methods, and below). It shows affinity with canadensis for traits related to its cooler thermal habitat but with glaucus for traits related to mimicry and sexual/female dimorphism (Figure 1B) [35]–[38]. This mosaic phenotype, which is critical to its existence in cooler, high-elevation habitats along the Appalachian Mountains but within the range of B. philenor, has led to the suggestion that appalachiensis may be a hybrid species [35], [37]. In support of this hypothesis, preliminary allozyme data have shown that appalachiensis has a canadensis-like allele at the Z-linked Lactate Dehydrogenase (Ldh) gene and a glaucus-like allele at the Z-linked Phosphogluconate Dehydrogenase (Pgd) gene [38].
Here we test the hypothesis that appalachiensis is a hybrid species with genomic admixture from glaucus and canadensis. If it is a hybrid species, its specific mixture of traits combined with the known sex-linkage of these traits make a clear prediction regarding the ancestry of appalachiensis' sex chromosomes: appalachiensis has inherited most of its Z chromosome from canadensis and its W chromosome from glaucus. We also test whether it has genome-wide admixture from the parental species, as expected of hybrid species that are formed with little subsequent backcrossing with parental species. In addition, we test alternative hypotheses regarding the apparently mixed genotype and phenotype of appalachiensis. Finally, we estimate divergence times between glaucus, canadensis and appalachiensis and study the evolutionary history of the traits that make up appalachiensis' hybrid phenotype in order to shed light on the evolutionary dynamics of the origin and maintenance of this hybrid species.
Results/Discussion
We tested the prediction that appalachiensis has mosaic sex chromosomes by sequencing six genes along the length of the Z chromosome and the mitochondrial cytochrome oxidase I (COI) gene (Materials and Methods, Table S2). While there are currently no known W-linked molecular markers in butterflies, we can infer the history of the W chromosome by sequencing mitochondrial DNA because both are maternally inherited and hence linked in Lepidoptera (see Materials and Methods). As predicted, our sequence data showed clear genetic discordance with appalachiensis having a mitochondrial genome similar to that of glaucus, and a Z chromosome similar to canadensis (Figure 2, Tables S3 and S4).

While the observed sex chromosome mosaicism suggests that appalachiensis evolved by combining key, sex-linked ecological traits from glaucus and canadensis, it does not tell us whether appalachiensis has a hybrid ancestry across the rest of its genome. Such widespread genomic admixture would be expected for a hybrid species formed with little subsequent backcrossing with the parental species. Alternatively, appalachiensis may be largely similar to one parental species across its genome, with a selectively introgressed W (and mtDNA) or Z chromosome from the other parental species [36]. To test these alternatives, we screened 2,035 nuclear amplified fragment length polymorphism (AFLP) markers and identified 249 that distinguished glaucus from canadensis. We then genotyped these ancestry informative markers in glaucus, canadensis, appalachiensis and laboratory-generated glaucus x canadensis F1 and F2 hybrids. We used the program STRUCTURE to genetically cluster the samples under the assumption that they represented two, three or four admixed populations. This analysis revealed that appalachiensis has genome-wide admixture indicative of historical hybridization between glaucus and canadensis (Figure 3A, Figure S2). Under the assumption of two populations (K = 2), appalachiensis and laboratory-generated hybrids showed similar and nearly equal admixture from glaucus and canadensis. At K = 3, appalachiensis formed its own cluster, and at K = 4 it remained a distinct cluster while subdivision within glaucus became apparent. In contrast, at K = 3 and K = 4, the laboratory-generated hybrids continued to show similar admixture from both parental species. A locus-by-locus analysis of molecular variance (AMOVA) of the AFLP dataset further confirmed widespread genomic admixture in appalachiensis: allele frequencies at 67 AFLP markers were similar to glaucus and significantly different from those of canadensis, 74 AFLP markers were similar to canadensis and significantly different from glaucus, and 92 markers had frequencies intermediate between glaucus and canadensis but significantly different from neither (Figure 3B). This analysis also revealed 16 AFLPs for which the frequency in appalachiensis was significantly different from both parental species (Figure 3B). Importantly, co-occurring appalachiensis and glaucus individuals, as identified morphologically at the time of collection from Spruce Knob in May and June 2006 (see Materials and Methods), showed species-specific sequences (Figure 2A) and AFLP signatures (Figure 3A). This suggests that these two co-occurring species are genetically distinct and that the clusters seen in Figure 2 and 3 are not merely geographically isolated subpopulations of the same species.

Examining the evolutionary context of the hybrid phenotype of appalachiensis requires a well-resolved phylogeny of tiger swallowtails. However, resolving this phylogeny from sequence data is challenging owing to recent divergence, incomplete lineage sorting and ongoing hybridization [39], [40] (also see Materials and Methods). Therefore, we used 1,607 polymorphic AFLP markers to generate a bootstrap-supported neighbor-joining tree of the entire clade. All tiger swallowtail nodes were strongly supported, except the glaucus, canadensis and appalachiensis clades (Figure 4A; also see Table S5). Within the glaucus-canadensis-appalachiensis groups, individuals formed species-specific clusters with only 12 out of the 184 samples clustering outside their own species (Figure 4A). We resolved the ancestral nodes between appalachiensis, glaucus and canadensis based on Figure 2 and Figure 3, and mapped relevant ecological traits (Table S6) on the resulting phylogeny (Figure 4B). We found that the mimetic female form suppressor is basal and widespread among tiger swallowtails. Mimicry and the enabler either evolved independently in garcia and glaucus, or were lost in alexiares and canadensis. The ancestral yellow female form was uniquely lost in garcia, whereas female dimorphism was uniquely gained in glaucus and then introgressed into appalachiensis. Similarly, obligatory pupal diapause and univoltinism evolved uniquely in canadensis and introgressed into appalachiensis. Thus, we not only identified the specific traits that recombined to form the hybrid species, we also traced the origin and spread of those traits through evolutionary history, down to the time they were brought together through hybridization to generate a new species.

The evolutionary history and direction of introgression in appalachiensis contrasts with the phenomenon of canadensis “late flight”, which has emerged recently near the northern limit of the glaucus-canadensis hybrid zone in Vermont [38], [41]. The late flight occurs in July and is allochronic relative to true (“early flight”) canadensis, which flies in May and June [38], [41]. The late flight has been hypothesized to be a result of hybridization between glaucus and canadensis, potentially representing an early stage in the evolution of an appalachiensis-like entity [41]. We used our DNA sequence and AFLP data to test whether late flight canadensis is of hybrid origin, and whether it is similar to appalachiensis. We found that unlike appalachiensis, the late flight is entirely canadensis-like at both its mitochondrial and Z-linked genes, except for three late flight individuals that were heterozygous for glaucus - and canadensis-like haplotypes at Kettin (Figure 2A). However, AFLP data show a strong signature of genome-wide admixture in late flight individuals (Figure 3C). Furthermore, the AFLP signature of the late flight is indistinguishable from that of laboratory-generated glaucus x canadensis F1 and F2 hybrids but distinct from appalachiensis (Figure 3C, Figure S2B), as expected if the late flight is a result of recent hybridization. The recent history of hybridization in the late flight is also supported by coalescent simulations (Figure S4, also see below and in Materials and Methods). Together, these data show that its history of introgression is dissimilar to appalachiensis. The late flight has the potential to speciate through allochronic flight period and larval host plant specialization [20], [41]; however, unlike appalachiensis, it may be a transient phase in the northward movement of the glaucus-canadensis hybrid zone in a changing thermal landscape. Moreover, although selection may maintain some characteristics of a mosaic sex chromosome in the late flight [20], [37], our data suggest that the late flight may not be experiencing natural selection for the same sex chromosome mosaicism that seems to have been essential in generating and maintaining the hybrid phenotype of appalachiensis.
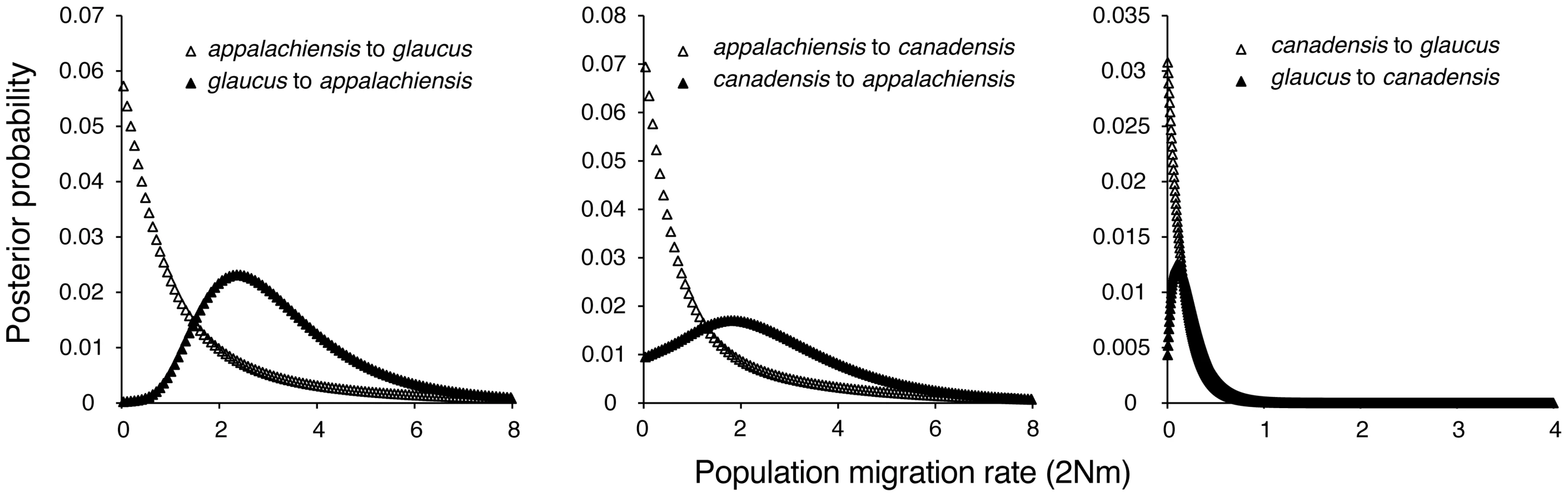
We tested two alternative hypotheses for the apparent hybrid ancestry of appalachiensis: (1) appalachiensis, glaucus and canadensis diverged from one another in a standard bifurcating manner and ancestral variation sorted out in such a way that appalachiensis now appears to have a hybrid genotype and phenotype, and (2) appalachiensis is not a reproductively isolated species but a very recent hybrid population that is constantly supplied by ongoing hybridization between glaucus and canadensis. We addressed the first hypothesis with our DNA sequence data by estimating divergence times and rates of historical introgression among the three species with the coalescent approach implemented in IMa2. Instead of supporting a bifurcating model of speciation, this analysis showed that glaucus and canadensis diverged approximately 580,000 years ago, whereas appalachiensis diverged from both the parental species approximately 100,000 years ago during the Pleistocene (Figure 5). The IMa2 analysis also showed essentially unidirectional introgression from glaucus and canadensis into appalachiensis (Figure 6), consistent with the hypothesized hybrid origin of appalachiensis.

We tested the second hypothesis by comparing among the three focal species: (a) the extent of linkage-disequilibrium, and (b) the proportion of species-specific genetic polymorphisms and haplotypes. Recent hybridization is expected to produce elevated linkage disequilibrium as a result of bringing together two distinct chromosomal copies from each parent species, the linkage disequilibrium decaying through successive generations as a result of recombination [42]. Indeed, linkage disequilibrium in both our DNA sequence and AFLP data was elevated in the laboratory-generated hybrids and late flight canadensis, both of which are products of very recent hybridization (Table 1). In contrast, linkage disequilibrium was significantly lower in appalachiensis and within the range of that seen in glaucus and canadensis (Table 1). Together, this evidence supports appalachiensis' historical origin and highlights the differences in the hybrid history between appalachiensis, late flight canadensis and laboratory-generated hybrids. If appalachiensis is indeed an old and subsequently isolated hybrid species, then it should possess unique mutations that it would have accumulated since it split from its parental species. These unique mutations should be detectable even if their number may be relatively small and they may not be fixed given: (a) the short estimated divergence time of ca 100,000 years, and (b) possibly low level of ongoing hybridization with the parental species. We calculated the proportion of species-specific single nucleotide polymorphisms and haplotypes among the three focal species based on our sequence data from seven genes (Table 2). The number and proportion of species-specific polymorphisms among glaucus, canadensis and appalachiensis varied greatly among genes and species, but all three species had a substantial number of species-specific polymorphisms (Table 2). The proportion of species-specific polymorphisms averaged across the seven genes was 0.58 (±0.221) for canadensis, 0.59 (±0.158) for glaucus, and 0.41 (±0.19) for appalachiensis. Thus, appalachiensis had lower average species-specific polymorphisms compared to its parental species, as expected from its younger age, but the difference was not significant (ANOVA, F(2,18) = 1.90, p = 0.179). We conclude that appalachiensis appears to have been isolated from the parental species long enough to accumulate species-specific mutations whose numbers are comparable with those of the parental species. This pattern is robust and holds true if we consider the number of species-specific haplotypes instead of polymorphisms: the proportion of species-specific haplotypes, averaged across all seven genes, was 0.71(±0.270) for canadensis, 0.9(±0.06) for glaucus, and 0.68(±0.098) for appalachiensis (ANOVA: F(2,18) = 3.47, p = 0.0533) (Table 2). These polymorphism and haplotype data support the hypothesis that appalachiensis is a good species rather than a recent or constantly supplied hybrid population. Moreover, additional lines of evidence are in conflict with the second hypothesis: (a) appalachiensis has a unique morphological and behavioral phenotype (Figure 1B and Figure S1), (b) there is significant genetic differentiation among appalachiensis, glaucus and canadensis, and appalachiensis forms a very well-supported and distinct genomic cluster that is apparent in both the DNA sequence (Figure 2) and AFLP data (Figure 3, Table S5), (c) IMa2 simulations showed appalachiensis to have diverged from the parental species long ago (Figure 5), and (d) Papilio appalachiensis is parapatric with canadensis and occurs southward to Georgia in the southern Appalachian Mountains, very far outside canadensis' range (Figure 1A; Materials and Methods). Therefore, appalachiensis could not be a product of ongoing hybridization between glaucus and canadensis. It is also important to note that its phenotype is unlike that of the laboratory-generated glaucus x canadensis hybrids.


Conclusions
Our data show that Papilio appalachiensis displays multiple hallmarks of a hybrid species. Furthermore, it has two unique features that inform us on the dynamics of hybrid speciation. First, it may potentially be the sole example of a hybrid species that spatially overlaps with both parental species. Second, its sex-linked ecologically important traits, and therefore sex chromosome mosaicism, appear to be under selection. In this case the sharp ecological clines along the latitudinal and altitudinal thermal gradients seem to maintain three, rather than two, spatially overlapping tiger swallowtail species. The evolution and persistence of appalachiensis in contact with its parental species suggests that hybridization among animals may result in selectively favored hybrid species that contribute to biodiversity. Moreover, sex chromosomes play an important role in speciation [43], and this has specifically been demonstrated in the Lepidoptera [44], [45], but this is the first example in which sex-linked traits seem to have contributed to hybrid speciation.
As a whole, our data suggest a scenario for the origin and maintenance of appalachiensis. During one of the late Pleistocene glacial retreats, canadensis populations retreated from their southern range while glaucus populations advanced northward and upward into the mountains. The changing thermal landscape likely brought the advancing glaucus populations into contact with a relict canadensis population in the Appalachian Mountains. The ensuing hybridization seems to have been largely unidirectional, with canadensis males preferentially mating with glaucus females, as they do today [12]. Hence, appalachiensis is now fixed for the glaucus mitochondrial genome, along with its W-linked female-limited mimicry and dimorphism. Both these traits are under frequency-dependent selection in the present appalachiensis habitat because with glacial retreats, selection for mimicry has also moved northward along with the range of B. philenor. However, the cold Appalachian thermal habitat ensured that the Z-linked genes associated with this lifestyle persisted from its canadensis ancestry in the proto-appalachiensis populations [16], [37]. This selection also persists today [38]. This unique combination of traits under varied selection may have been critical in helping appalachiensis evolve as a distinct species. The maintenance of genome-wide admixture in appalachiensis (Figure 3) also suggests that it was formed via a relatively brief burst of hybridization between the parental species, with little backcrossing with glaucus despite their continued sympatry throughout appalachiensis' range. A few other hybrid species have likely emerged in a similarly short span of time with potentially little backcrossing [8], [46], [47]. Interestingly, our estimated divergence times of approximately 100,000 and 90,000 years between glaucus/canadensis and appalachiensis (see Figure 5) fall precisely in the last interglacial period in North America, known as the Sangamonian Stage (125,000 to 75,000 years ago), which is congruent with the scenario just outlined. It may be possible to explore this scenario with additional population genetic data for these species.
Papilio appalachiensis may provide a rare genetic example of the creative role of hybridization in evolution [8]. Hybrids are often considered maladapted and viewed as lying in fitness valleys between the adaptive peaks that the parental species occupy in the adaptive landscape [3]. In case of tiger swallowtails, however, the northward and upward movement of the toxic B. philenor in the Appalachian Mountains may have created a new, unoccupied adaptive peak with selection for a combination of cold thermal habitat and mimicry. Neither of the parental species appears to be able to occupy this peak. Our results suggest an intriguing scenario in which the hybrid species (appalachiensis), with its precise combination of phenotypic traits, may have landed directly on this novel, unoccupied adaptive peak.
The primary goals of this work were to test whether appalachiensis exhibited genetic evidence of hybrid ancestry, mosaic sex chromosomes, and genome-wide admixture. Our results reveal several intriguing patterns that appear to support appalachiensis' status as a hybrid species and suggest an evolutionary scenario for its origin and maintenance. Our work also highlights specific areas where additional data will enrich our understanding of this system in particular and the dynamics of hybrid speciation in general. One such area is identifying the specific mechanisms that generate reproductive isolation between appalachiensis and both of its parental species. Our genetic data, and the low frequency of glaucus/appalachiensis hybrids from areas where they co-occur in West Virginia, suggest that these two species exhibit some reproductive isolation. However, the ecological, behavioral and genetic factors that contribute to this isolation remain unknown, as does the exact degree of isolation. Specifically, we do not currently understand the mechanisms of isolation or the precise geographic ranges and areas of overlap between appalachiensis and canadensis. In addition, more widespread sampling of appalachiensis from across its range would allow us to test its genomic integrity throughout the range, and also to estimate the extent to which it hybridizes with each of the parental species. Given the latitudinal range of appalachiensis relative to glaucus and canadensis, it is possible that appalachiensis experiences variable amounts of recent introgression with each of the two parental species. Our current sampling was limited so we were unable to test this hypothesis. However, genetic data for our appalachiensis sample from its southern range in North Carolina and co-occurring samples of appalachiensis and glaucus from West Virginia suggest that appalachiensis may be fairly homogeneous throughout its range, but more widespread sampling is warranted. Scriber [48] has recently pointed out that ongoing climate warming may diminish the cool thermal mountain refuges in the southern Appalachian Mountains of northern Georgia, western North Carolina and eastern Tennessee in appalachiensis' southernmost range. The changing thermal landscape may induce increased introgression of glaucus genes into appalachiensis, which may diminish the genomic contribution from canadensis in these appalachiensis populations. Clearly, the rich biological detail of this system holds promise to test various aspects of hybrid speciation and persistence in animals.
Materials and Methods
Study System
The parental species, P. glaucus and P. Canadensis
Tiger swallowtails (Papilio glaucus species group) form a small, monophyletic species group of the American subgenus Pterourus of Papilio [40]. Many tiger swallowtail species look similar in general appearance, although they have consistent differences in ecological traits as well as larval, pupal and adult morphology [49], [50]. Nonetheless, there had been much confusion about subspecies and species status of three parapatric taxa: rutulus, canadensis and glaucus, since they occasionally hybridize when in contact at their range margins [50] (also see Figure 1A). Due to the lack of complete sympatry and occasional hybridization, many earlier authors had treated rutulus and canadensis as subspecies of glaucus [25], [50], [51]. The three taxa have now been solidly established as distinct species through the past three decades of work by JMS and his collaborators. This work has revealed multiple mechanisms of reproductive isolation separating glaucus and canadensis that indicate that they are good biological species [12]–[25] (also see the Introduction).
Interspecific hybridization is common in nature and there are many well-studied interspecific hybrid zones [3], [4], [52], [53]. Several features have been used to distinguish interspecific hybrid zones from contact zones between subspecies, most importantly: (a) interspecific hybrid zones are marked by a reduction in hybrids relative to the parental forms (“bimodal hybrid zone”) [54], and (b) the maintenance of “genotypic clusters” in spite of hybridization [55]. The glaucus/canadensis hybrid zone meets these two hallmarks of interspecific hybrid zones: (a) hybrids make up less than 20% of the community in the very middle of the glaucus/canadensis hybrid zone [28], and (b) our present genetic data as well as previous allozyme work [29], [56], [57] show that glaucus and canadensis form distinct genotypic clusters despite hybridization. The rate of hybridization reported for glaucus and canadensis in the middle of the hybrid zone is within the range of rate of hybridization known for other well-established species pairs [52].
Lastly, more recent phylogenetic work has shown rutulus and canadensis to be well-separated from glaucus [40]. Our present work (Figure 2, Figure 3, Figure 4, Figure 5, Table S5) further reinforces the view that rutulus, canadensis and glaucus are genetically strongly diverged parapatric species, not subspecies, despite occasional hybridization.
The putative hybrid species, P. appalachiensis
Papilio appalachiensis has a distinct and readily recognizable phenotype spanning morphology, ecology and behavior, and this phenotype is maintained in sympatry/parapatry with the parental species, glaucus and canadensis [35], [36]. There are several factors that suggest that appalachiensis is a distinct species rather than a recent hybrid population. For example, its melanic female form is unlike any other tiger swallowtail species, and both the male and female wing patterns are unlike any laboratory-generated F1 and F2 glaucus x canadensis hybrids. Similar to the glaucus/canadensis hybrid zone (see above), appalachiensis shows two hallmarks of a species: (a) it has a bimodal hybrid zone with glaucus, and (b) it forms a unique genetic cluster distinct from its parental species (see Figure 2 and Figure 3, and Results/Discussion). Papilio appalachiensis shows highly reduced rate of hybridization with glaucus, with which it completely overlaps in distribution (Figure 1A). Although appalachiensis is found at mid - and high elevations in the Appalachian Mountains and glaucus is usually found at lower elevations, our work shows that they co-occur widely both along the elevational and latitudinal gradients at least for several weeks during late May and June. For example, KK found fresh individuals of both species feeding side by side on the same flowering honeysuckle bushes below Spruce Knob, West Virginia, in May and June 2006. Thus, there is ample opportunity for hybridization between the two taxa. Nonetheless, we collected only 16 suspected glaucus/appalachiensis hybrids (phenotypically intermediate between glaucus and appalachiensis), along with 114 phenotypically pure appalachiensis and 172 phenotypically pure glaucus within the overlap zone of appalachiensis and glaucus from Tennessee to northeastern West Virginia. Thus, the proportion of suspected hybrids between the two species was very small, approximately 5.6%, making this a clearly bimodal hybrid zone. These numbers are well below the estimates of hybridization for many other hybridizing species [4] and point to reduced hybridization as one would expect for distinct species. As a clarification, individuals were scored as putative hybrids between appalachiensis and glaucus based on the diagnostic wing color patterns given in the original species description [35], [36], mainly: (1) square versus crescent-shaped submarginal yellow spots on the hindwing, (2) contiguous yellow band versus row of spots on forewing underside, and (3) extent of blue on the upper hindwing of females. They were first identified as probable hybrids by KK and then all specimens were blindly scored by H. Pavulaan, who independently arrived at the same conclusion (Pavulaan originally described appalachiensis). If our species assignments had been wrong, underestimating the true rate of hybridization based on phenotypic characters, the genetic clustering shown in Figure 2 and Figure 3 would have been very different.
The contact zone and nature of isolation between appalachiensis and canadensis are currently not well understood. These two species are believed to be sympatric in the Appalachian Mountains from West Virginia northward [35], [58]. Almost all of our appalachiensis samples were collected near Spruce Knob, West Virginia (Table S1, Figure S5), very near localities where canadensis reportedly occurs. Based on our published [37], [38] as well as unpublished work, we suspect that canadensis has recently moved northward along the Appalachian Mountains, perhaps completely out of West Virginia, due to ongoing climate warming. It is likely that appalachiensis and canadensis maintain a hybrid zone similar to the glaucus-canadensis hybrid zone, and this hybrid zone may also have moved northward due to the ongoing warming in this area. Current work by MLA and JMS aims to delineate the present northern boundary of appalachiensis distribution, the southern boundary of canadensis in the Appalachian Mountains, and the boundaries of the contact zone between the two species. However, based on current understanding [35], [58], appalachiensis and canadensis are narrowly sympatric or parapatric.
Specimen Collection and Lab Methods
Table S1 lists details of the 244 wild-caught individuals and 23 laboratory-generated hybrids used for genetic analysis, and Figure S5 shows localities where these specimens were collected. We preserved bodies in ethanol and stored wings in glassine envelopes. We measured wingspan from the preserved wings using vernier calipers, from the base of the forewing to its tip (Figure S1). We extracted genomic DNA from legs and thoracic muscle tissue using DNeasy Blood & Tissue Kit (Qiagen, Germantown, MD).
We sequenced one mitochondrial (Cytochrome oxidase I, COI) and six Z-linked genes (Kettin (Ket), Tyrosine hydroxylase (TH), Triosephosphate isomerase (Tpi), Period (Per), Lactate dehydrogenase (Ldh) and Phenylalanine hydroxylase (PAH)) to study potential admixture in appalachiensis (NCBI accession numbers JF764373-JF764558, JF951433-JF951722). Primer sequences and lengths of aligned sequences are given in Table S2. PCR protocols used for previously published primers are given in the literature [27], [59]. We designed primers for TH and PAH by aligning sequences from Bombyx mori and Papilio xuthus (Table S2). Previous allozyme work had shown that appalachiensis has a glaucus-like Pgd allele [38], so we also developed several primers for that gene. Unfortunately, none of these primers worked. Future comparisons of Pgd sequences among the focal species will be useful for inferring the extent of mosaicism along the appalachiensis Z chromosome. We amplified all Z-linked genes with Invitrogen Platinum hot-start taq (Invitrogen Corp, Carlsbad, CA) and a touchdown protocol with annealing temperatures from 65–50°C.
We generated amplified fragment length polymorphism (AFLP) data using a Plant Mapping Kit (Applied Biosystems, Foster City, CA) and 10 primer pairs: EcoRI-ACA/MseI-CAC, EcoRI-ACA/MseI-CAG, EcoRI-ACA/MseI-CAT, EcoRI-ACA/MseI-CTC, EcoRI-ACA/MseI-CTG, EcoRI-ACT/MseI-CAC, EcoRI-ACT/MseI-CAG, EcoRI-ACT/MseI-CAT, EcoRI-ACT/MseI-CTC, and EcoRI-ACT/MseI-CTG.
Analyses of DNA Sequence Data
We cleaned DNA sequences using Sequencher (Gene Codes Corp., Ann Arbor, MI). When available, we included reference sequences from GenBank and aligned the sequence datasets online using Multiple Sequence Comparison by Log-Expectation program (MUSCLE; http://www.ebi.ac.uk/Tools/muscle/index.html). We corrected the sequence output alignments by eye, paying special attention to indels in introns of Z-linked genes. For population genetic analyses, indels longer than one base pair were scored as a single polymorphism.
We used this dataset to analyze sequence similarity between glaucus, canadensis and appalachiensis by performing species pair-wise locus-by-locus analyses of molecular variance (AMOVAs) using Arlequin [60] to calculate fixation index (FST) values (Figure 2B, Table S4). Given their young ages and occasional hybridization, there is considerable polymorphism but few fixed differences among the three species. Hence, we tested the genic ancestry of appalachiensis in relation to the parental species for polymorphisms that were strongly differentiated (p≤0.001) between glaucus and canadensis (Figure 2A). However, we used entire sequences to calculate the FST values in Figure 2B. Specimens that were sequenced for only one gene are included in Table S3 (missing data are marked grey) and were used in calculating FST values in Figure 2B, but excluded from Figure 2A. Genotypes here refer to nucleotide bases at specific SNP or indel polymorphisms in the genes, which can be diploid (Z-linked polymorphisms scored in males) or haploid (mtDNA, and Z-linked polymorphisms scored in females). The Z-linked genes in Figure 2A and Tables S2, S3, S4 are listed in the same order in which they occur on the Bombyx mori Z chromosome.
The Use of Mitochondrial DNA to Infer the History of the W Chromosome
We wanted to determine whether appalachiensis inherited its W chromosome, where the gene controlling female-limited mimicry is situated, from canadensis or glaucus. Unfortunately, no W-linked molecular markers exist for these butterflies. Therefore, we used a mitochondrial gene because the W chromosome and the mitochondrial genome are both maternally inherited and thus linked in Lepidoptera. Andolfatto et al. [61] had attempted to use mtDNA to examine the evolutionary history of the W-linked female-limited mimetic phenotype in glaucus. Surprisingly, they found no association between mtDNA genealogies and mimicry, and attributed this finding to paternal leakage of mtDNA and recombination among maternal and paternal mtDNA. In fact, there are three other, potentially more likely explanations for the polyphyly of mimicry with respect to the mitochondrial gene tree: (1) multiple origins of mimicry resulting in more than one W-linked mimicry allele, (2) loss of mimicry in some individuals resulting in more than one non-mimicry allele, and (3) the presence of a low-frequency Z-linked mimicry suppressor allele in glaucus [26], [61]. Thus, the observed disassociation between mtDNA and mimicry does not necessarily mean disassociation between mtDNA and the W chromosome. Unfortunately, the above alternatives cannot be rigorously tested at present because the molecular identities of the mimicry and enabler genes are yet unknown. However, it is important to remember that there is overwhelming evidence from throughout the animal kingdom showing that mtDNA is almost exclusively maternally inherited [62]–[64]. Finally, it is critical to note that rare paternal inheritance of mtDNA could only influence our interpretation regarding the history of the W chromosome if it occurred during the specific time period during which glaucus and canadensis hybridized to form appalachiensis. If this had occurred, we would predict a mixture of glaucus and canadensis mtDNA haplotypes in appalachiensis, which we do not see (Figure 2A). Instead, our extensive dataset reveals that glaucus and canadensis mtDNA haplotypes are exclusively species-specific, and that of appalachiensis is like glaucus (Figure 2A). This pattern shows that paternal leakage of mtDNA has not occurred between glaucus and canadensis during the origin of appalachiensis. Thus, mtDNA is an informative marker for inferring the species-level ancestry of appalachiensis' W chromosome.
Analyses of AFLP Data
We analyzed AFLP data using Genemapper Software Version 4.0 (Applied Biosystems, Foster City, CA). Samples were analyzed with the default bin width of 1.0 bp, and the fragment analysis range of 50–250 bps. We analyzed the AFLP data in two ways: we identified population clusters and genomic admixture among the focal species using the program STRUCTURE [65], and we constructed a phylogeny of tiger swallowtails using PAUP* [66]. We performed population clustering in STRUCTURE using three datasets: (a) focal species alone (glaucus (39 samples), canadensis (41 samples) and appalachiensis (38 samples); total 118 samples) (Figure S2A), (b) focal species along with laboratory-generated glaucus/canadensis hybrids (23 hybrid samples; total 141 samples) (Figure 3A), and (c) focal species along with laboratory-generated glaucus/canadensis hybrids and wild caught late flight canadensis individuals (30 late flight canadensis individuals; total 171 samples) (Figure 3C, Figure S2B).
We performed three separate STRUCTURE analyses because each was aimed at testing a different hypothesis and thus required that a different subset of samples be included. For instance, our first analysis focused only on the three focal species to determine whether appalachiensis exhibited evidence of admixture between canadensis and glaucus yet was genetically distinct from both of them (Figure S2A). To do this, we used a well-established approach that has been used previously to both support and refute the hypothesis of hybrid speciation [6], [67]. When admixture is present, as in the case of hybrid speciation, STRUCTURE can estimate the proportion of each individual's genome that is derived from distinct gene pools (admixture proportions). Gompert et al. [6] showed that a hybrid species with genomic mosaicism displayed admixture proportions split between the parental species when analyzed with the parental species at K = 2, but then formed a distinct cluster at K = 3 (just as we have shown here for appalachiensis) [6]. In comparison, Kronforst et al. [67] showed that if one does the same analysis with a species that does not appear to be a hybrid species, at K = 2 it clusters entirely with its sister species but then forms its own cluster at K = 3 [67]. Thus, we used STRUCTURE to estimate admixture proportions instead of assignment probabilities of the sampled individuals. We did this because we were asking what proportion of genome of each appalachiensis individual was glaucus - versus canadensis-like (which is shown by admixture proportions), not whether a particular appalachiensis individual was assigned to one or the other parental species with a certain probability (which is shown by assignment probabilities). Using this method, we discovered a pattern in appalachiensis (Figure 3) consistent with it being a hybrid species [6]. We used a similar approach for our second analysis focused on comparing the genetic signature of appalachiensis to lab generated glaucus/canadensis hybrids (Figure 3A), while our third analysis focused on the ancestry of the late-flight canadensis population (Figure 3C).
Our STRUCTURE analyses were based on 249 AFLP markers that were significantly differentiated (p≤0.05) between glaucus and canadensis as judged by FST values from a locus-by-locus AMOVA. We analyzed each dataset in STRUCTURE with these ancestry-informative AFLP markers with 200,000 burn-in generations and 1,000,000 generations of data collection under the assumption of two, three and four admixed populations (K = 2, 3 or 4). We used STRUCTURE to estimate admixture proportions rather than assignment probabilities, and visualized the STRUCTURE outputs with the program Distruct [68] (Figure 3). Analyses based on the full AFLP dataset (polymorphisms with a minor allele frequency ≥5%) produced similar results but with less resolution (Figure S3).
For Figure 3B, we used the same 249 AFLP markers that were significantly differentiated between glaucus and canadensis (see last paragraph), did pair-wise AMOVAs among the three species, and categorized AFLP loci as: 1) “glaucus-like”: appalachiensis allele frequency is significantly different from canadensis but not from glaucus, 2) “canadensis-like”: appalachiensis allele frequency is significantly different from glaucus but not from canadensis, 3) “intermediate”: appalachiensis allele frequency is intermediate but not significantly different from either parental species, and 4) “different”: appalachiensis allele frequency is different from both parental species. The pie chart (Figure 3B) shows the number of appalachiensis AFLP loci that fall in the four categories, highlighting the intermediacy of appalachiensis with respect to glaucus and canadensis.
Phylogenetic Methods
In order to study the larger evolutionary context of appalachiensis' unique hybrid phenotype, we used the AFLP dataset to resolve phylogenetic relationships between tiger swallowtails. The AFLP phylogenetic dataset included 184 individuals with P. garamas as the outgroup, and 1,607 polymorphic AFLP markers. Laboratory-generated glaucus/canadensis hybrids and suspected wild-caught glaucus/appalachiensis hybrids were excluded from the phylogenetic analysis. We generated a neighbor-joining tree of tiger swallowtails in PAUP* [66] (Figure 4A), with bootstrap support based on 2,000 pseudo-replicates. The distance-based neighbor-joining method was used because there is no model of evolution for AFLP data [69]–[73]. AFLPs are reliable for phylogenetic reconstruction and, in fact, provide answers consistent with other sources of data in addition to being able to resolve relationships that DNA sequence data cannot [74]–[76]. Zakharov et al. [40] only included glaucus, canadensis, rutulus and multicaudata, so only half of the eight tiger swallowtail species were covered in their phylogeny. Note that our relationships among those species are exactly the same as in Zakharov et al., which shows that AFLPs resolve phylogenies consistent with other sources of molecular data. The critical species for inferring the evolution of mimicry among tiger swallowtails are alexiares and garcia, which we included in our phylogeny for the first time. Several nuclear and mitochondrial genes tested were not informative in resolving relationships between some tiger swallowtail species (analysis not presented here), so fast-evolving molecular markers such as AFLPs were needed to resolve these relationships.
We mapped on this phylogeny the ecological and morphological traits (Table S6) relevant to the hybrid phenotype of appalachiensis using the software MacClade 4 [77]. Since phylogenetic programs do not model speciation and trait evolution on reticulated trees, for the initial mapping of traits we excluded appalachiensis from the tree and evolved traits from the outgroup (P. garamas) to glaucus and canadensis. Based on the signatures of hybrid speciation in Figure 2 and Figure 3, appalachiensis and its mosaic inheritance of traits from glaucus and canadensis were subsequently manually added to the tree, post-hoc.
Estimating Divergence Times and Gene Flow
We used our DNA sequence data and the program IMa2 [78]–[81] to estimate divergence times (Figure 5) and historical gene flow (Figure 6) among appalachiensis, glaucus and canadensis. This coalescent-based method uses comparative DNA sequence data and an “Isolation with Migration” model of population divergence to estimate model parameters such as effective population sizes, divergence times, and bi-directional migration rates. While IMa2 can analyze more than two populations at a time, it requires populations to be related by a specified bifurcating tree. Since we were testing the predicted hybrid origin of appalachiensis, which does not follow a bifurcating mode of speciation, we analyzed our data as three pair-wise comparisons among the three focal species. For each analysis, we ran IMa2 with 100 Metropolis-coupled chains with a 150,000-step burn-in followed by 20 million steps of data collection. An important requirement of the IMa2 method is that there is free recombination among sequenced loci: loci do not necessarily have to be on separate chromosomes but they should not be too close to one another. Since six of our seven sequenced loci were on the Z chromosome, and they spanned the length of that chromosome, we satisfied this requirement by using data from only three genes for these analyses: Kettin and PAH, which are on opposite ends of the Z chromosome, and the mitochondrial gene COI. Another requirement of IMa2 is that there is no recombination within loci. We satisfied this requirement by trimming the sequence dataset to a region with no evidence of recombination using the program IMgc Online [82]. We used this trimmed dataset with three genes for estimating both divergence times and gene flow. Converting parameter estimates to units such as time in years since divergence or population migration rates (2Nm) requires mutation rates for each gene. We estimated these by comparing sequences to the within tiger swallowtail outgroup, P. multicaudata. Average mtDNA divergence between multicaudata and the three focal species was 3.17%. Using previous COI divergence estimate of 1.15% per lineage per million years [83], we estimated that P. multicaudata split from the common ancestor of our focal species approximately 1.4 million years ago. This date differs from that estimated by Zakharov et al. [40] for all of Papilio, but our date may be more accurate since it was estimated specifically for tiger swallowtails. However, the actual dates are less important for our purpose since the relevant findings from the IMa2 analysis were that: (a) appalachiensis originated much more recently compared to the time when glaucus and canadensis split from one another (Figure 5), and (b) appalachiensis split from both glaucus and canadensis around the same time (Figure 5).
A similar analysis was done to estimate divergence times between glaucus, canadensis and late-flight canadensis (Figure S4).
Comparisons of Linkage Disequilibrium
We estimated the extent of linkage disequilibrium across both our AFLP and Z-linked DNA sequence data and compared this among populations. For each dataset (AFLPs or concatenated Z-linked gene sequences) we calculated pair-wise associations among all polymorphisms using the exact test for linkage disequilibrium implemented in Arlequin. We summarized these results by calculating the average percentage of polymorphisms that were significantly (p≤0.01) associated with each other (Table 1) and then compared them among species using an ANOVA.
Comparisons of Species-Specific Polymorphisms
We used our DNA sequence dataset to calculate the number of species-specific polymorphisms for each of our focal species (Table 2). We calculated species-specific polymorphisms as the proportion of genetic polymorphisms unique to one species, out of the total number of polymorphisms (including shared and species-specific) seen among all the sequenced samples of that species. We used the software DnaSP [84] to count the total number of polymorphisms and the subset that were species-specific for each species at each gene. We then compared the average proportion of species-specific polymorphisms among species with an ANOVA. We also performed a similar analysis comparing the proportion of species-specific haplotypes rather than polymorphisms, and found similar results (Table 2).
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. RiesebergLHRaymondORosenthalDMLaiZLivingstoneK 2003 Major ecological transitions in wild sunflowers facilitated by hybridization. Science 301 1211 1216
2. LewontinRCBirchLC 1966 Hybridization as a source of variation for adaptation to new environments. Evolution 20 315 336
3. MalletJ 2007 Hybrid speciation. Nature 446 279 283
4. MalletJ 2005 Hybridization as an invasion of the genome. Trends Ecol Evol 20 229 237
5. CoyneJAOrrHA 2004 Speciation. Sunderland, Massachusetts Sinauer Associates
6. GompertZFordyceJAForisterMLShapiroAMNiceCC 2006 Homoploid hybrid speciation in an extreme habitat. Science 314 1923 1925
7. MavarezJSalazarCABerminghamESalcedoCJigginsCD 2006 Speciation by hybridization in Heliconius butterflies. Nature 441 868 871
8. NolteAWTautzD 2010 Understanding the onset of hybrid speciation. Trends Gen 26 54 58
9. GrossBLRiesebergLH 2005 The ecological genetics of homoploid hybrid speciation. J Heredity 96 241 252
10. SalazarCBaxterSWPardo-DiazCWuGSurridgeA 2010 Genetic evidence for hybrid trait speciation in Heliconius butterflies. PLoS Genet 6 e1000930 doi:10.1371/journal.pgen.1000930
11. MavarezJLinaresM 2008 Homoploid hybrid speciation in animals. Mol Ecol 17 4181 4185
12. DeeringMDScriberJM 2002 Field bioassays show heterospecific mating preference asymmetry between hybridizing North American Papilio butterfly species (Lepidoptera: Papilionidae). J Ethol 20 25 33
13. ScriberJMDeeringMStumpA 2003 Hybrid zone ecology and tiger swallowtail trait clines in North America. BoggsCLWattWBEhrlichPR Butterflies: Ecology and Evolution Taking Flight Chicago University of Chicago Press 367 391
14. ScriberJMLederhouseRCDowellRV 1995 Hybridization studies with North American swallowtails. ScriberJMTsubakiYLederhouseRC Swallowtail Butterflies: Their Ecology and Evolutionary Biology Gainesville, Florida Scientific Publishers, Inc 269 281
15. HagenRHScriberJM 1995 Sex chromosomes and speciation in tiger swallowtails. ScriberJMTsubakiYLederhouseRC Swallowtail Butterflies: Their Ecology and Evolutionary Biology Gainesville, , Florida Scientific Publishers, Inc. 211 227
16. MercaderRJScriberJM 2008 Asymmetrical thermal constraints on the parapatric species boundaries of two widespread generalist butterflies. Ecol Entomol 33 537 545
17. LederhouseRCAyresMPScriberJM 1995 Physiological and behavioral adaptations to variable thermal environments in North American swalowtail butterflies. ScriberJMTsubakiYLederhouseRC Swallowtail Butterflies: Their Ecology and Evolutionary Biology Gainesville, , Florida Scientific Publishers, Inc 71 81
18. ScriberJMLederhouseRC 1992 The thermal environment as a resource dictating geographic patterns of feeding specialization of insect herbivores. HunterMROhgushiTPricePW Effects of Resource Distribution on Animal-Plant Interactions San Diego, California Academic Press 429 466
19. LindrothRLScriberJMHsiaMTS 1986 Differential responses of tiger swallowtail subspecies to secondary metabolites from tulip tree and quaking aspen. Oecologia 70 13 19
20. MercaderRJAardemaMLScriberJM 2009 Hybridization leads to host-use divergence in a polyphagous butterfly sibiling species pair. Oecologia 158 651 662
21. MercaderRJScriberJM 2007 Diversification of host use in two polyphagous butterflies: differences in oviposition specificity or host rank hierarchy. Ent Exp Appl 125 89 101
22. BossartJLScriberJM 1995 Genetic variation in oviposition preference in tiger swallowtail butterflies: interspecific, interpopulation and interindividual comparisons. ScriberJMTsubakiYLederhouseRC Swallowtail Butterflies: Their Ecology and Evolutionary Biology Gainesville, Florida Scientific Publishers, Inc 183 193
23. RitlandDBScriberJM 1985 Larval developmental rates of three subspecies of tiger swallowtail butterflies, Papilio glaucus, and their hybrids in relation to temperature. Oecologia 65 185 193
24. ScriberJMLindrothRLNitaoJK 1989 Differential toxicity of a phenolic glycoside from quaking aspen to Papilio glaucus butterfly subspecies, hybrids and backcrosses. Oecologia 81 186 191
25. HagenRHLederhouseRCBossartJLScriberJM 1991 Papilio canadensis and P. glaucus (Papilionidae) are distinct species. J Lep Soc 45 245 258
26. ScriberJMHagenRHLederhouseRC 1996 Genetics of mimicry in the tiger swallowtail butterflies, Papilio glaucus and P. canadensis (Lepidoptera: Papilionidae). Evolution 50 222 236
27. PutnamASScriberJMAndolfattoP 2007 Discordant divergence times among Z-chromosome regions between two ecologically distinct swallowtail butterfly species. Evolution 61 912 927
28. LuebkeHJScriberJMYandellBS 1988 Use of Multivariate Discriminant Analysis of male wing morphometrics to delineate a hybrid zone for Papilio glaucus glaucus and P. g. canadensis in Wisconsin. Amer Midland Natur 119 366 379
29. HagenRHScriberJM 1989 Sex-linked diapause, color, and allozyme loci in Papilio glaucus: linkage analysis and significance in a hybrid zone. J Heredity 80 179 185
30. RiesLMullenSP 2008 A rare model limits the distribution of its more common mimic: A twist on frequency-dependent Batesian mimicry. Evolution 62 1798 1803
31. KunteK 2009 Female-limited mimetic polymorphism: A review of theories and a critique of sexual selection as balancing selection. Anim Behav 78 1029 1036
32. BrowerJVZ 1958 Experimental studies of mimicry in some North American butterflies: Part II. Battus philenor and Papilio troilus, P. polyxenes and P. glaucus. Evolution 12 123 136
33. BrowerLPBrowerJVZ 1962 The relative abundance of model and mimic butterflies in natural populations of the Battus philenor mimicry complex. Ecology 43 154 158
34. BurnsJM 1966 Preferential mating versus mimicry: Disruptive selection and sex-limited dimorphism in Papilio glaucus. Science 153 551 553
35. PavulaanHWrightDM 2002 Pterourus appalachiensis (Papilionidae: Papilioninae), a new swallowtail butterfly from the Appalachian Region of the United States. Tax Rep Internl Lep Sur 3 1 20
36. PavulaanHWrightDM 2004 Discovery of a black female form of Pterourus appalachiensis (Papilionidae: Papilioninae) and additional observations of the species in West Virginia. Tax Rep Internl Lep Sur 6 1 10
37. ScriberJMOrdingGJ 2005 Ecological speciation without host plant specialization; possible origins of a recently described cryptic Papilio species. Ent Exp Appl 115 247 263
38. ScriberJMOrdingGJMercaderRJ 2008 Introgression and parapatric speciation in a hybrid zone. TilmonKJ Specialization, Speciation, and Radiation: The Evolutionary Biology of Herbivorous Insects Berkeley, , California University of California Press 69 87
39. SperlingFAH 2003 Butterfly molecular systematics: From species definitions to higher-level phylogenies. BoggsCLWattWBEhrlichPR Butterflies: Ecology and Evolution Taking Flight Chicago University of Chicago Press 431 458
40. ZakharovEVCaterinoMSSperlingFAH 2004 Molecular phylogeny, historical biogeography, and divergence time estimates for Swallowtail Butterflies of the genus Papilio (Lepidoptera: Papilionidae). Syst Biol 53 193 215
41. OrdingGJMercaderRJAardemaMLScriberJM 2010 Allochronic isolation and incipient hybrid speciation in tiger swallowtail butterflies. Oecologia 162 523 531
42. BartonNHGaleKS 1993 Genetic analysis of hybrid zones. HarrisonRG Hybrid Zones and the Evolutionary Process USA Oxford University Press 13 45
43. QvarnströmABaileyRI 2009 Speciation through evolution of sex-linked genes. Heredity 102 4 15
44. SperlingFAH 1994 Sex-linked genes and species differences in Lepidoptera. Can Ent 126 807 818
45. ProwellDP 1998 Sex linkage and speciation in Lepidoptera. HowardDJBerlocherSH Endless forms: Species and speciation Oxford Oxford University Press 309 319
46. SchwarzDMattaBMShakir-BotteriNLMcPheronBA 2005 Host shift to an invasive plant triggers rapid animal hybrid speciation. Nature 436 546 549
47. NolteAWFreyhofJStemshornKCTautzD 2005 An invasive lineage of sculpins, Cottus sp. (Pisces, Teleostei) in the Rhine with new habitat adaptations has originated from hybridization between old phylogeographic groups. Proc R Soc B 272 2379 2387
48. ScriberJM 2011 Impacts of climate warming on hybrid zone movement: Geographically diffuse and biologically porous “species borders”. Insect Sci 18 121 159
49. BrowerLP 1959 Speciation in butterflies of the Papilio glaucus group. I. Morphological relationships and hybridization. Evolution 13 40 63
50. ScottJA 1986 The Butterflies of North America: A Natural History and Field Guide. Stanford, USA Stanford University Press
51. RothschildWJordanK 1906 A revision of the American Papilios. Novitates Zoologicae 13 411 744
52. MalletJBeltranMNeukirchenWLinaresM 2007 Natural hybridization in heliconiine butterflies: the species boundary as a continuum. BMC Evol Biol 7 28
53. SperlingFAH 1990 Interspecific hybrids in Papilio butterflies: Poor taxonomy or interesting evolutionary problem? Can J Zool 68 1790 1799
54. JigginsCDMalletJ 2000 Bimodal hybrid zones and speciation. Trends Ecol Evol 15 250 255
55. MalletJ 1995 A species definition for the modern synthesis. Trends Ecol Evol 10 294 299
56. HagenRHScriberJM 1991 Systematics of the Papilio glaucus and P. troilus species groups (Lepidoptera: Papilionidae): inferences from allozymes. Annals Ent Soc Amer 84 380 395
57. HagenRH 1990 Population structure and host use in hybridizing subspecies of Papilio glaucus (Lepidoptera: Papilionidae). Evolution 44 1914 1930
58. AllenTJ 1997 The butterflies of West Virginia and their caterpillars University of Pittsburgh Press
59. NazariVZakharovEVSperlingFAH 2007 Phylogeny, historical biogeography, and taxonomic ranking of Parnassiinae (Lepidoptera, Papilionidae) based on morphology and seven genes. Mol Phylogen Evol 42 131 156
60. ExcoffierLLischerH 2009 Arlequin: An Integrated Software Package for Population Genetics Data Analysis. Ver 3.11
61. AndolfattoPScriberJMCharlesworthB 2003 No association between mitochondrial DNA haplotypes and a female-limited mimicry phenotype in Papilio glaucus. Evolution 57 305 316
62. BirkyCWJr 1995 Uniparental inheritance of mitochondrial and chloroplast genes: mechanisms and evolution. Proc Natl Acad Sci, USA 92 11331 11338
63. MoritzCDowlingTEBrownWM 1987 Evolution of animal mitochondrial DNA: relevance for population biology and systematics. Annual Review of Ecology and Systematics 18 269 292
64. AviseJCArnoldJBallRMBerminghamELambT 1987 Intraspecific phylogeography: the mitochondrial DNA bridge between population genetics and systematics. Ann Rev Ecol Syst 18 489 522
65. PritchardJKStephensMDonnellyP 2000 Inference of population structure using multilocus genotype data. Genetics 155 945 959
66. SwoffordDL 2002 PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4 Sunderland, , MA Sinauer Associates
67. KronforstMRSalazarCLinaresMGilbertLE 2007 No genomic mosaicism in a putative hybrid butterfly species. Proc R Soc B 274 1255 1264
68. RosenbergNA 2007 Distruct: A Program for the Graphical Display of Population Structure. Ver 1.1
69. AlbertsonRCMarkertJADanleyPDKocherTD 1999 Phylogeny of a rapidly evolving clade: The cichlid fishes of Lake Malawi, East Africa. Proc Natl Acad Sci, USA 96 5107 5110
70. BeardsleyPMYenAOlmsteadRG 2003 AFLP phylogeny of Mimulus section Erythranthe and the evolution of hummingbird pollination. Evolution 57 1397 1410
71. HollingsworthPMEnnosRA 2004 Neighbour joining trees, dominant markers and population genetic structure. Heredity 92 490 498
72. MendelsonTCShawKL 2005 Rapid speciation in an arthropod. Nature 433 375 376
73. RudhARogellBHoglundJ 2007 Non-gradual variation in colour morphs of the strawberry poison frog Dendrobates pumilio: genetic and geographical isolation suggest a role for selection in maintaining polymorphism. Mol Ecol 16 4284 4294
74. ChamberlainNLHillRIKapanDDGilbertLEKronforstMR 2009 Polymorphic butterfly reveals the missing link in ecological speciation. Science 326 847 850
75. QuekS-PCountermanBAMouraPAdCardosoMZMarshallCR 2010 Dissecting comimetic radiations in Heliconius reveals divergent histories of convergent butterflies. Proc Natl Acad Sci, USA 107 7365 7370
76. SavageWKMullenSP 2009 A single origin of Batesian mimicry among hybridizing populations of admiral butterflies (Limenitis arthemis) rejects an evolutionary reversion to the ancestral phenotype. Proc R Soc B 276 2557 2565
77. MaddisonDRMaddisonWP 2003 MacClade 4. Version 4.06 Sunderland, , MA Sinauer Associates
78. NielsenRWakeleyJ 2001 Distinguishing migration from isolation. A Markov chain Monte Carlo approach. Genetics 158 885 96
79. HeyJNielsenR 2004 Multilocus methods for estimating population sizes, migration rates and divergence time, with applications to the divergence of Drosophila pseudoobscura and D. persimilis. Genetics 167 747 760
80. WonYJHeyJ 2005 Divergence population genetics of chimpanzees. Mol Biol Evol 22 297 307
81. HeyJNielsenR 2007 Integration within the Felsenstein equation for improved Markov Chain Monte Carlo methods in population genetics. Proc Natl Acad Sci, USA 104 2785 2790
82. WoernerAECoxMPHammerMF 2007 Recombination-filtered genomic datasets by information naximization. Bioinformatics 23 1851 1853
83. BrowerAVZ 1994 Rapid morphological radiation and convergence among races of the butterfly Heliconius erato inferred from patterns of mitochondrial DNA evolution. Proc Natl Acad Sci, USA 91 6491 6495
84. LibradoPRozasJ 2009 DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25 1451 1452
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2011 Číslo 9
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Retrotransposon-Induced Heterochromatin Spreading in the Mouse Revealed by Insertional Polymorphisms
- The Evolutionarily Conserved Longevity Determinants HCF-1 and SIR-2.1/SIRT1 Collaborate to Regulate DAF-16/FOXO
- Genome-Wide Analysis of Heteroduplex DNA in Mismatch Repair–Deficient Yeast Cells Reveals Novel Properties of Meiotic Recombination Pathways
- Association of eGFR-Related Loci Identified by GWAS with Incident CKD and ESRD
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy