
Blood Fluke Exploitation of Non-Cognate CD4 T Cell Help to Facilitate Parasite Development
Schistosoma blood flukes, which infect over 200 million people globally, co-opt CD4+ T cell-dependent mechanisms to facilitate parasite development and egg excretion. The latter requires Th2 responses, while the mechanism underpinning the former has remained obscure. Using mice that are either defective in T cell receptor (TCR) signaling or that lack TCRs that can respond to schistosomes, we show that naïve CD4+ T cells facilitate schistosome development in the absence of T cell receptor signaling. Concurrently, the presence of naïve CD4+ T cells correlates with both steady-state changes in the expression of genes that are critical for the development of monocytes and macrophages and with significant changes in the composition of peripheral mononuclear phagocyte populations. Finally, we show that direct stimulation of the mononuclear phagocyte system restores blood fluke development in the absence of CD4+ T cells. Thus we conclude that schistosomes co-opt innate immune signals to facilitate their development and that the role of CD4+ T cells in this process may be limited to the provision of non-cognate help for mononuclear phagocyte function. Our findings have significance for understanding interactions between schistosomiasis and other co-infections, such as bacterial infections and human immunodeficiency virus infection, which potently stimulate innate responses or interfere with T cell help, respectively. An understanding of immunological factors that either promote or inhibit schistosome development may be valuable in guiding the development of efficacious new therapies and vaccines for schistosomiasis.
Published in the journal:
. PLoS Pathog 6(4): e32767. doi:10.1371/journal.ppat.1000892
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1000892
Summary
Schistosoma blood flukes, which infect over 200 million people globally, co-opt CD4+ T cell-dependent mechanisms to facilitate parasite development and egg excretion. The latter requires Th2 responses, while the mechanism underpinning the former has remained obscure. Using mice that are either defective in T cell receptor (TCR) signaling or that lack TCRs that can respond to schistosomes, we show that naïve CD4+ T cells facilitate schistosome development in the absence of T cell receptor signaling. Concurrently, the presence of naïve CD4+ T cells correlates with both steady-state changes in the expression of genes that are critical for the development of monocytes and macrophages and with significant changes in the composition of peripheral mononuclear phagocyte populations. Finally, we show that direct stimulation of the mononuclear phagocyte system restores blood fluke development in the absence of CD4+ T cells. Thus we conclude that schistosomes co-opt innate immune signals to facilitate their development and that the role of CD4+ T cells in this process may be limited to the provision of non-cognate help for mononuclear phagocyte function. Our findings have significance for understanding interactions between schistosomiasis and other co-infections, such as bacterial infections and human immunodeficiency virus infection, which potently stimulate innate responses or interfere with T cell help, respectively. An understanding of immunological factors that either promote or inhibit schistosome development may be valuable in guiding the development of efficacious new therapies and vaccines for schistosomiasis.
Introduction
Extensive co-evolution of parasitic organisms and their hosts has given rise to complex host-parasite relationships in which exploitation of host responses to infection by parasites is a recurring theme. Nowhere is this complexity in host-parasite relationships better exemplified than in the parasitic helminths, which infect a third of the world's human population [1] by establishing chronic infections that persist for years, often in the face of vigorous immune responses [2]. Schistosoma blood flukes account for a significant proportion of these helminth infections, causing considerable morbidity and mortality. After initiating infection by direct skin penetration, schistosomes migrate in the bloodstream to hepatic pre-sinusoidal venules, where rapid growth and development ensues, culminating in mating of adult worms and the production of eggs.
It was previously shown that Schistosoma mansoni co-opts CD4+ T cell-dependent mechanisms to facilitate both parasite development during pre-patent infection and the excretion of parasite eggs after the onset of oviposition [3], [4], [5]. The latter requires formation of Th2-dependent granulomas in the bowel wall to allow passage of eggs from the portal vasculature into the intestinal lumen [3], [4], [6]. However, the mechanism by which CD4+ T cells facilitate development of schistosome worms has not been elucidated. While homeostatic maintenance of peripheral CD4+ T cells through the action of γc cytokines is required to provide a permissive environment for schistosome development [7], previous studies did not identify a role for effector Th1/Th2 responses or any single effector cytokine in parasite development [5], [8], [9].
In this study we tested whether activation of CD4+ T cells through the T cell receptor (TCR) by schistosome antigens is required for schistosome development to proceed. Unexpectedly, our data indicate that CD4+ T cells that lack specificity for schistosome antigens can facilitate schistosome development in the absence of antigen-mediated T cell activation. Interestingly, the presence of naïve, resting CD4+ T cells also correlates both with steady-state transcriptional changes in the expression of genes that are critical for development of monocytes and macrophages and with changes in the composition of peripheral monocyte populations. Further, direct stimulation of the mononuclear phagocyte system bypasses the requirement for CD4+ T cells in schistosome development, suggesting that innate cells of the mononuclear phagocyte system facilitate schistosome development and that CD4+ T cells influence the parasites indirectly by modulating monocyte/macrophage function. Together, our results demonstrate that blood flukes exploit innate immune signals to facilitate their development and suggest that the role of CD4+ T cells in this process may be limited to the provision of non-cognate help for innate mononuclear cell function.
Results
TCR signaling blockade does not impair parasite development
To evaluate the role of T cell activation in schistosome development, we first examined schistosome development in Bcl10 (Unigene accession no. Mm.239141) -deficient and protein kinase C θ (PKCθ; Mm.329993) –deficient mice, where impairment of NF-κB activation in response to TCR ligation renders T cells unresponsive to antigen [10], [11]. Unexpectedly, S. mansoni recovered from Bcl10-/- and wild type mice were indistinguishable in size (Figure 1A) and deposited comparable numbers of eggs (Figure 1B), unlike schistosomes from recombination activating gene 1 (RAG-1; Mm.828) -deficient mice that exhibit a severely stunted phenotype and greatly reduced rates of egg production ([5] and data not shown; see also Figure 1D and E). As expected, CD4+ T cell responses to S. mansoni were impaired by Bcl10 deletion, as measured by acquisition of an activated phenotype (Figures S1A and B) and cytokine production (Figure S1C), suggesting that activation of CD4+ T cells through the TCR and subsequent CD4+ T cell responses are dispensable for normal schistosome development. Identical results were obtained with PKCθ-/- mice (data not shown). To directly test whether Bcl10-/- CD4+ T cells can facilitate schistosome development, parasite development was examined in RAG-1-/- mice that were reconstituted with Bcl10-/- CD4+ T cells. Adoptive transfer of Bcl10-/- CD4+ T cells prior to infection partially restored worm growth (Figures 1C and D), resulting in a phenotype that was intermediate between RAG-1-/- recipients of wild type CD4+ T cells and non-reconstituted RAG-1-/- mice. Likewise, adoptive transfer of Bcl10-/- CD4+ T cells produced an intermediate egg production phenotype that was not significantly different from RAG-1-/- recipients of wild type CD4+ T cells or non-reconstituted RAG-1-/- mice, even though egg production in these latter two groups was significantly different from each other (Figure 1E). Identical results were obtained when adoptive transfers were performed with PKCθ-/- CD4+ T cells (data not shown). Because TCR signals are required for T cell homeostasis [12], Bcl10-/- CD4+ T cells exhibited variable levels of engraftment that were consistently lower than those of wild type cells (Figure 1F). However, there was a significant positive correlation between the number of Bcl10-/- CD4+ T cells and the mean length of the worms recovered from each recipient (Figure 1G). These data suggest that CD4+ T cells do not require intact antigen receptor signaling to facilitate parasite development. Further, our data suggest there is a simple requirement for sufficient numbers of CD4+ T cells to allow parasite development to proceed and predict a minimum of approximately 2×106 CD4+ T cells as the number required to restore parasite growth to wild type levels (Figure 1G).

Non-responsive CD4+ T cells facilitate S. mansoni development
To test whether TCR specificity for schistosome antigens is necessary for CD4+ T cells to facilitate schistosome development, we examined S. mansoni development in TCR-transgenic RAG-/- mice that transgenically express previously rearranged MHC class II-restricted TCRs specific for irrelevant antigens. In RAG-/- mice possessing only chicken ovalbumin (OVA)-specific (OT-II/RAG-1-/- C57BL/6, DO11.10/RAG-2-/- BALB/c) or pigeon cytochrome C (PCC)-specific (Cyt5-CC7/RAG-2-/- B10.A) CD4+ T cells, schistosome development was enhanced relative to RAG-/- controls that lack CD4+ T cells, as determined by assessment of parasite size (Figures 2A and B) and egg production (Figure 2C). The extent to which parasite development was rescued varied between the different strains, with DO11.10/RAG-2-/- mice supporting parasite development that was comparable to wild type mice and parasites from OT-II/RAG-1-/- and Cyt5-CC7/RAG-2-/- exhibiting intermediate levels of development. As expected, OVA - and PCC-specific CD4+ T cells from S. mansoni-infected RAG-/- mice were completely unresponsive to schistosome antigens (Figures S2A and S2B), though these cells retained the ability to respond to the appropriate antigen (Figure S2B). These data demonstrate that CD4+ T cells do not require specificity for schistosome antigens and do not need antigen stimulation in order to facilitate schistosome development. Furthermore, the MHCII molecule by which the TCR is restricted is also likely irrelevant, as the three transgenic TCRs used recognize different murine MHCII molecules. Taken together, the data in Figures 1 and 2 suggest that CD4+ T cells influence the outcome of schistosome infection by mechanisms that are independent of antigen receptor specificity and signaling.
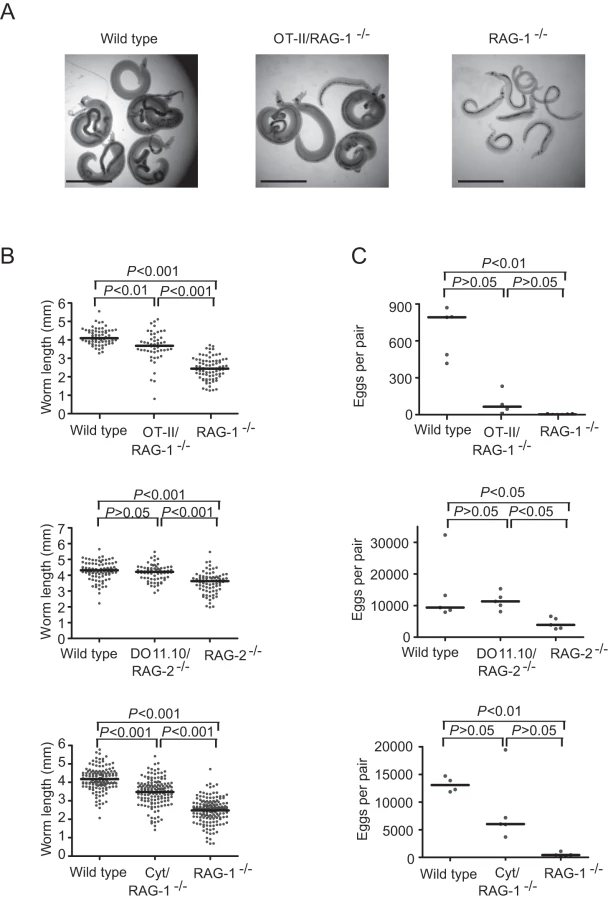
The presence of naïve CD4+ cells correlates with steady-state changes in mononuclear phagocyte development
Regardless of antigen specificity, naïve CD4+ T cells are maintained by active homeostatic processes [12], requiring γc cytokines and interactions with professional APCs that express MHCII [13], [14], [15], [16], [17], [18]. Because the effect of CD4+ T cells on schistosome development is independent of antigen receptor specificity and antigen-mediated activation (Figures 1 and 2), we hypothesized that steady-state homeostatic interactions between naïve CD4+ T cells and MHCII+ APCs in host tissues may play a role in promoting schistosome development. To identify transcriptional changes induced by the steady-state homeostatic interactions of naïve CD4+ T cells, we used a whole genome microarray to compare transcript levels in liver tissue from non-infected RAG-1-/- and OT-II/RAG-1-/- mice (NCBI GEO series accession number GSE8340.) Using this approach, we identified 165 genes that were differentially expressed only in the presence of naïve CD4+ T cells (Figure 3A and Table S1). None of these genes are specifically expressed by CD4+ T cells, suggesting that OT-II CD4+ T cells contribute relatively little to the total hepatic RNA, being relatively few in number and transcriptionally quiescent. Supervised average-linkage hierarchical clustering of the expression data revealed that the transcriptional profile of hepatic tissue from OT-II/RAG-1-/- mice was more similar to that of wild type mice than RAG-1-/- mice, with the single largest grouping of genes comprising those that were expressed at higher levels in wild type and OT-II/RAG-1-/- mice when compared to RAG-1-/- mice (Figure 3A).

To identify biological functions associated with the differentially expressed genes, we employed a structured, knowledge-based approach to identify networks of differentially expressed genes with related functions [19]. Ingenuity Pathways Analysis software (Ingenuity Systems, www.ingenuity.com) was used to overlay differentially expressed genes onto a global molecular network developed from information contained in the Ingenuity Pathways Knowledge Base. Networks of up to 35 differentially expressed genes were then algorithmically generated based on the functional connectivity between the genes. Numerical network scores were calculated to rank networks according to their degree of relevance to the molecules in the dataset. Using this approach, five high-scoring networks (score>20) of differentially expressed genes were identified, a representative of which (score = 45) is illustrated in Figure 3B. Biological functions significantly associated with the networks, together with the genes associated with each function, are displayed in Table S2. Biological functions with the most significant associations included Cell Morphology (P = 1.37×10−4−4.80×10−2), Cellular Compromise (P = 1.37×10−4−4.31×10−2), Cell Cycle (P = 2.28×10−4−4.58×10−2), Cell Death (P = 6.30×10−4−4.31×10−2) and Cell-to-Cell Signaling and Interaction (P = 7.51×10−4−4.31×10−2), suggesting significant changes in cellular development within the liver in the presence of naïve CD4+ T cells. Interestingly, genes significantly up-regulated in the presence of naïve CD4+ T cells included Csf1/macrophage colony-stimulating factor (M-CSF) (UniGene accession no. Mm.795), which is an essential growth factor for monocytes and monocyte-derived macrophages [20], and other genes involved in myeloid cell development and function (Myd116 (Mm.4048) [21], Cd2 (Mm.22842) [22], Gli1 (Mm.391450) [23] and Plld (Mm.29933) [24]; highlighted in Figure 3A), suggesting specific alterations in mononuclear phagocyte development. Other genes that were up-regulated in the presence of naïve CD4+ T cells are associated with the cellular response to stress (highlighted in Figure 3A), including genes involved in the response to mis-folded proteins [25], e.g. several chaperones (Hsp90aa1 (UniGene Accession No. Mm.341186) [26], Hsp90ab1 (Mm.2180) [26], Hsp110 (Mm.270681) [27], p23 (Mm.305816) [28], Dnajb9 (Mm.27432) [29]), enzymes involved in protein folding (Sep15 (Mm.29812) [30], a disulfide isomerase (Mm.33692) [31], a peptidyl-prolyl cis-trans isomerase (Mm.32842)), enzymes involved in protein degradation (Ubxd2 (Mm.29812) [32], the Syvn1 ubiquitin ligase (Mm.149870) [33], Usp43 (Mm.158885)), and DNA-binding proteins involved in the transcriptional response to stress (an XBP-1-like transcription factor (Mm.187453) [34], the ER transmembrane transcription factor Creb3l2 (Mm.391651) [35], H2afx (Mm.245931) [36], and Junb (Mm.1167) [37]). Junb in particular is a central regulator of the cellular response to stress and is targeted for phosphorylation by stress-activated protein kinases [38]. Presumably because of the role of these cells in phagocytosis and in antigen processing and presentation, stress responses are constitutively active in immature APCs and are required for APC development and survival [39], Indeed, the stress response gene Junb is also a critical regulator of myeloid cell differentiation and is classified as a myeloid differentiation primary response (MyD) gene [40]. Also up-regulated in the presence of naïve CD4+ T cells were genes involved in another critical aspect of phagocyte and APC function, namely vesicle formation and transport, e.g. Rabgap1l (Mm.25833) [41], Arhgap21 (Mm.28507) [42], Arfip2 (Mm.41637) [43], Kif13b (Mm.23611) [44], and Diap1 (Mm.195916), which is also implicated in receptor-mediated phagocytosis in myeloid cells [45]. Thus, transcriptional changes consistent with alterations in mononuclear phagocyte and/or APC development and function correlate with the presence of naïve CD4+ T cells in OT-II/RAG-1-/- mice.
To explore the possibility that mononuclear phagocyte development was altered by the presence of CD4+ T cells, leukocytes from the spleen and liver of non-infected RAG-1-/- and OT-II/RAG-1-/- mice were compared by flow cytometry. After gating to exclude granulocytes and OT-II T cells, mononuclear phagocytes were identified by expression of the myeloid marker CD11b (Mm.262106) [46]. In both mouse strains, CD11b+ cells segregated into two populations based on relative CD11b expression levels and forward scattering (FSC) properties – CD11bhi FSChi and CD11blo FSClo (Figure 3C). However, the relative proportions of the two populations differed markedly in the two strains, with RAG-1-/- mice possessing relatively more CD11blo cells than OT-II/RAG-1-/- mice (Figure 3C). Further analysis revealed that the two CD11b+ populations also differed significantly in expression of CD115 (Mm.22574, the receptor for M-CSF), with CD11hi cells exhibiting higher CD115 expression than CD11lo cells (Figure 3D; P = 0.0016). Both CD11b+ populations also expressed Ly6C (Mm.1583, a monocyte marker), although again, CD11bhi cells exhibited higher levels than CD11blo cells (Figure 3D). Together, the greater forward scatter and elevated expression of CD115 by CD11bhi cells strongly suggest these are functionally more mature mononuclear phagocytes than CD11blo cells, as increased size and expression of CD11b and CD115 are all implicated in monocyte activation and maturation [47], [48], [49]. Finally, systematic analysis of CD11bhi and CD11blo populations in age-matched groups of non-infected OT-II/RAG-1-/- and RAG-1-/- mice revealed that cells of the functionally more mature CD11bhi phenotype were significantly more abundant in OT-II/RAG-1-/- mice than RAG-1-/- mice (Figure 3E). Thus, our data suggest that, compared to OT-II/RAG-1-/- mice, RAG-1-/- mice exhibit a defect in mononuclear phagocyte development, with accumulation of cells that possess an immature CD11blo phenotype. Furthermore, this alteration in myeloid cell development correlates with reduced expression of genes that are required for mononuclear phagocyte development and function (Figure 3A, Figure 3B, Tables S1 and S2). As the only difference between RAG-1-/- and OT-II/RAG-1-/- mice is that, in the latter, transgenic expression of a previously rearranged MHC II-restricted TCR allows for development of a monospecific population of CD4+ T cells with specificity for chicken ovalbumin, our data suggest that steady-state MHCII-TCR-mediated interactions between mononuclear cells and naïve CD4+ T cells enhances developmental progression of mononuclear phagocytes to a functionally more mature state.
Direct activation of mononuclear phagocytes rescues schistosome development in the absence of CD4+ T cells
Since enhanced parasite development and egg production in OT-II/RAG-1-/- mice correlated with enhanced steady-state maturation of monocytes in these animals, we hypothesized that non-responsive OT-II CD4+ T cells facilitate schistosome development indirectly, through steady-state interactions with mononuclear cells that promote monocyte maturation. To test this hypothesis, we tested whether direct stimulation of mononuclear phagocyte maturation, in the absence of any CD4+ T cells, could substitute for CD4+ T cells in restoring parasite development in RAG-/- mice. As the Toll-like receptor (TLR) -4 (Mm.38049) ligand lipopolysaccharide (LPS) has previously been shown to stimulate the in vivo maturation of monocytes [50], [51], we administered ultrapure LPS to schistosome-infected RAG-/- mice during pre-patent infection and assessed the effect on parasite growth and egg production at 6 weeks post infection. Consistent with other reports [50], we found that LPS administration caused a significant decrease in the relative numbers of CD11b+ monocytes (Figure 4A), indicative of the terminal maturation and subsequent apoptosis of these cells in response to TLR-4 ligation. Treatment of both BALB/c RAG-2-/- (Figures 4B-D) and C57BL/6 RAG-1-/- (Figures 4E and 4F) mice with LPS led to significant increases in both parasite length (Figures 4B, 4C and 4E) and egg production (Figure 4D and 4F). Indeed, parasite growth (Figure 4E) and egg production (Figure 4F) were responsive to increasing doses of LPS. Together these data indicate that innate immune signals alone are sufficient to support schistosome development and suggest that blood flukes exploit the role of CD4+ T cells in providing help for mononuclear phagocyte maturation and function.

Discussion
Parasites of the genus Schistosoma likely arose over 70 million years ago [52] and have undergone complex co-evolution with their definitive hosts, resulting in parasite adaptations that both evade and exploit host immune functions. Previous studies showed that schistosomes require host CD4+ T cells for normal development [5], [53], [54] and to mediate egress of eggs from the body across the bowel wall [3], [55]. While egg excretion requires induction of an effector Th2 response to egg antigens [6], the CD4+ T cell effector functions that facilitate blood fluke development have not been elucidated. Because previous studies failed to identify a specific role for either Th1 or Th2 responses in schistosome development [5], [8], the question of whether any CD4+ T effector functions are required to promote schistosome development remained unresolved. Therefore, the purpose of this study was to test whether antigen receptor signaling and subsequent activation of CD4+ T cells are necessary for normal parasite development to proceed. Our results show that, in contrast to the requirement for effector T cell responses to facilitate egg excretion, neither recognition of schistosome antigens nor TCR-mediated activation of CD4+ T cells are required for normal parasite development. Our findings suggest that none of the effector functions typically associated with CD4+ T cell responses are directly implicated in facilitating schistosome development, and may explain why previous attempts to identify a single T cell factor that modulates schistosome development, using knockout mice deficient in individual cytokines or their receptors [8], have been unsuccessful.
The schistosome requirement for CD4+ T cells, but the lack of necessity for traditional T cell effector functions, suggests that the steady-state homeostatic activities of naïve CD4+ T cells make the host environment more conducive to schistosome development. While the role of homeostatic interactions with MHCII+ cells in maintaining the peripheral CD4+ T cell pool is well established, the necessity of these same interactions for efficient APC maturation is being increasingly recognized [18], [56], [57]. In T cell-deficient mice, APC development and function are compromised but can be restored by reconstitution of peripheral T cell populations [18]. Furthermore, it was recently demonstrated that T cell conditioning of APCs for efficient maturation occurs under steady-state conditions before the initiation of T cell responses, in the absence of cognate antigen, and is mediated, at least in part, by the co-stimulatory molecule B7-H1 expressed by naïve T cells [57]. We propose that similar steady-state interactions between naïve OT-II T cells and MHCII+ cells in OT-II/RAG-1-/- mice account for the baseline increases in APC-related gene expression and the alterations in mononuclear cell maturation we observed in these mice.
Given the ability of naïve T cells to prime for efficient APC maturation, we hypothesized that exploitation of APC function by schistosomes, rather than of CD4+ T cells directly, accounts for the enhanced parasite development we observed in TCR-transgenic RAG-/- mice. While we cannot exclude the possibility that unknown factors elaborated by resting, naïve T cells directly influence schistosome development, our hypothesis provides a parsimonious explanation for why naïve CD4+ T cells that do not respond to schistosome infection can still influence parasite development. That schistosome development can be restored in the complete absence of CD4+ T cells, through direct maturation of APCs, supports this hypothesis by demonstrating that CD4+ T cells are not directly required for parasite development.
As the sentinels of the immune system, circulating monocytes and tissue macrophages and dendritic cells, also known as the mononuclear phagocyte system, specifically express pattern recognition receptors (PRRs), including TLRs, which allow for the detection of invading pathogens [58]. Indeed, monocytes express high levels of TLRs and are the predominant producers of proinflammatory cytokines during endotoxic shock [59]. Thus while other cells in RAG-/- mice express PRRs, the monocytes, macrophages and dendritic cells are the predominant responders to pathogen-associated molecular patterns (PAMPs) such as LPS and are the likely mediators of the effect of LPS on schistosome development. Monocytes, macrophages and dendritic cells are ontologically related, as monocytes are the macrophage and dendritic cell precursors that migrate into the tissues, both during steady-state conditions and during infection. In response to infection, PRR ligation stimulates increased monocyte flux to meet the elevated demand for antigen-presenting and effector cells to combat infection [60]. Our demonstration that LPS administration restores schistosome development is therefore further evidence in support of the hypothesis we propose above, that schistosome development is influenced by innate mononuclear cell function. Indeed, our data point to mononuclear cell function as the common mechanism underpinning the enhancement of schistosome development by both naïve CD4+ T cells and LPS. This hypothesis is under further investigation.
Several hypotheses can be proposed to explain how mononuclear phagocytes might influence shistosome development. Recruited to sites of infection and tissue damage, mononuclear cells produce cytokines and chemokines in response to activation that could act as cues for developing schistosomes [51]. Alternatively, the localized release of proinflammatory cytokines by mononuclear cells would be predicted to increase blood flow and vascular permeability, perhaps increasing the supply of host factors required for normal parasite development. At sites of inflammation, monocyte-derived cells also contribute to blood vessel remodeling and angiogenesis, by secreting angiogenic factors and trans-differentiating into endothelial cells [61]. As schistosomes are intravascular parasites and early parasite development occurs within portal venules, vessel remodeling may be required to allow for parasite growth. Yet another possibility is that mononuclear phagocytes may directly damage schistosomes, therefore requiring their local depletion for schistosome development to proceed normally. Activated macrophages and dendritic cells can produce nitric oxide [62], a molecule that is toxic to developing schistosomes [63] and mediates immunity to schistosome infection in animals vaccinated with irradiated cercariae [64]. Priming of APC maturation by CD4+ T cells or TLR ligands might allow for more rapid progression of these cells to apoptosis in response to activation. In RAG-/- mice, developmental impairment of APCs might allow for their persistence, constituting a persistent source of nitric oxide and/or other molecules that impair schistosome development. If this is the case, analysis of innate responses to developing schistosomes in RAG-/- mice may identify innate effector mechanisms that can be harnessed to enhance immunity to schistosome infection. These hypotheses are currently being tested.
Our data demonstrate that blood flukes do not respond to CD4+ T cells directly, but rather respond to signals that originate from the innate immune system. These findings corroborate previous studies where tumor necrosis factor (TNF; Mm.1293), an innate proinflammatory cytokine, was shown to stimulate parasite egg laying in CD4+ T cell-deficient Prkdcscid/scid mice [65]. While TNF does not appear to stimulate parasite development directly [5], [8], the overlap between the TNF receptor and TLR signaling pathways could account for the ability of both ligands to enhance schistosome development [66], [67]. Our findings also have implications for understanding epidemiological associations between schistosomiasis and other infections [68], such as salmonellosis. While schistosome infection has been implicated in persistence of Salmonella infection [69], our data also suggest that proinflammatory stimuli produced in response to bacterial LPS could play a role in exacerbating schistosome infection by supporting parasite development in co-infected individuals. These putative immunological and epidemiological associations are currently under investigation. The possible requirement of schistosomes for proinflammatory signals to support normal development is intriguing, as schistosomes, and helminths in general, are largely unable to stimulate such responses themselves due to a lack of potent TLR ligands [70]. It is therefore tempting to speculate that, under evolutionary pressure to avoid immune detection, schistosomes have lost the ability to stimulate the inflammatory feedback required for their successful development and now rely on other mechanisms to generate these essential inflammatory signals.
In summary, our investigation of the mechanism by which CD4+ T cells facilitate schistosome development has revealed that blood flukes require neither CD4+ T cell responses nor associated effector functions. Furthermore, our data show that schistosomes do not respond directly to CD4+ T cells, as their requirement for these cells can be bypassed completely by direct stimulation of innate immune responses. Indeed, our data suggest that the role of CD4+ T cells in facilitating schistosome development may be limited to the provision of non-cognate T cell help for the maturation of MHCII+ APCs. We provide two lines of evidence in support of this hypothesis. First, non-responsive, naïve CD4+ T cells, which also condition immature APCs to undergo maturation, support improved parasite development. Second, direct stimulation of APC maturation in the absence of CD4+ T cells restores schistosome development. These data, together with previous findings that macrophages mediate vaccine-induced immunity to schistosome infection [64], implicate mononuclear cells as central host determinants of the outcome of schistosome infection in the definitive host. A detailed understanding of the interactions between blood flukes and the mononuclear phagocyte system could therefore identify opportunities to modulate mononuclear cell function in ways that impair or prevent the establishment of schistosome infections.
Materials and Methods
Ethics statement
All animal studies were performed in accordance with protocols approved by the USUHS Institutional Animal Care and Use Committee.
Experimental mice
RAG-1-/- mice were purchased from Jackson Laboratory (Bar Harbor, ME) and bred in-house to generate sufficient numbers for experiments. Bcl10-/- mice [11] were kindly provided by Dr. Tak Mak. OT-II mice [71] were the kind gift of Dr. Francis Carbone. Bcl10-/- and OT-II mice, originally with a mixed 129/C57BL/6 background, were backcrossed to the C57BL/6 background. OT-II mice were then bred with RAG-1-/- in house, to generate OTII/RAG-1-/- mice. C57BL/6 (National Cancer Institute, Frederick, MD) and 129× C57BL/6 F1 hybrid wild type mice (Taconic) were used as positive controls, although no differences in parasitological parameters were found in parasites recovered from either wild type strain (data not shown). BALB/cTac-TgN(DO11.10)-Rag2tm1 (DO11.10/RAG-2-/-) mice [72] and B10.A/AiTac-[Tg]TCRCyt5CC7-I-[KO]-Rag2tm1 (Cyt/RAG-2-/-) mice [73] were kindly provided by Dr. Dragana Jankovic. RAG-2-/- and wild type mice on the BALB/c and B10.A2 backgrounds (Taconic) were used as controls, respectively.
Mice were infected percutaneously via the tail skin with 150 S. mansoni cercariae (Puerto Rican strain) shed from infected Biomphalaria glabrata snails [74]. All animal studies were performed in accordance with protocols approved by the USUHS Institutional Animal Care and Use Committee.
Parasite recovery and measurement of parasitological parameters
Parasites were recovered from the portal system by perfusion [74] 42 days post-infection, immediately fixed in 4% neutral-buffered formaldehyde and photographed using a Nikon Coolpix 4500 4.0 megapixel digital camera connected to a Vistavision trinocular dissecting microscope at 20× magnification. Length of male parasites was determined from digital images using ImageJ software (http://rsb.info.nih.gov/ij). Quantitative analysis of parasite length was performed on male worms as male schistosomes always outnumber females in experimental infections and female growth is significantly influenced by pairing with males [75]. Liver tissue was digested in 0.7% trypsin (50 ml) in phosphate-buffered saline (PBS) for 2–3 hours at 37°C, and eggs were counted under a dissecting microscope. Egg production per schistosome pair was determined by dividing the total number of eggs calculated for each mouse liver by the number of parasite pairs recovered.
Cell isolation and adoptive transfer
Lymph nodes and spleens from wild type C57BL/6 mice or Bcl10-/- mice were dispersed through a 70-µm nylon strainer. Single cell suspensions were washed and red blood cells were lysed using ACK lysing buffer (Quality Biological, Inc.) Cells were incubated with anti-CD4 (Mm.2209) coated microbeads (Miltenyi Biosciences) and separated using Midi-Macs magnetic columns (Miltenyi Biosciences). Flow cytometric analysis of isolated CD4+ T cells routinely demonstrated a purity of ∼99%. 3×106 cells suspended in PBS were transferred into RAG-1-/- mice by intravenous injection into a lateral tail vein. Control recipients received PBS alone. All tissue culture reagents used were free of endotoxin as determined by routine testing. Recipient animals were then infected with cercariae 24 hours post transfer, as described above. To verify the efficacy of adoptive transfers at necropsy, splenocytes from reconstituted RAG-1-/- mice were surface labeled with APC-Cy7-conjugated antibodies to CD4, FITC-conjugated antibodies to CD8 (Mm.1858), APC-conjugated antibodies to TCRβ (Mm.333026), PE-conjugated antibodies to NK1.1 (Mm.6180) and PerCp-Cy5.5-conjugated antibodies to CD19 (Mm.4360) (BD Biosciences) and analyzed using a LSR II Optical Bench flow cytometer with FACSDiva and Winlist software, version 5.0 (Verity Software House). The total number of T cells in each recipient after adoptive transfer was estimated by multiplying the percent of CD4+TCRβ+ cells by the total number of splenocytes for each mouse.
Analysis of cytokine production
CD11c+ cells were isolated from wild type spleens through incubation of cells suspensions with anti-CD11c (Mm.22378) coated microbeads (Miltenyi Biosciences and separated using Midi-Macs magnetic columns (Miltenyi Biosciences). CD4+ cells were isolated from spleens and livers of wild type, Bcl10-/- and OTII/RAG-1-/- mice as described above. CD4+ T cells and CD11c+ cells, pulsed with 50 µg/ml schistosome worm antigen preparation (SWAP)[76], [77], 5 µg/ml OVA or 1 µg/µl anti-CD3 (Mm.210361) (BD Bioscience), were co-cultured for 72 hours in 96 well plates at a ratio of 5×105 CD4+ cells to 5×104 CD11c+ cells. Culture supernatants were assessed for IFNγ (Mm.240327) and IL-10 (Mm.874) by ELISA using BD Opt EIA Mouse IFNγ and IL-10 antibody pairs and ELISA reagents (BD Bioscience) and analyzed using a Spectramax M2 Plate reader (Molecular Devices).
Analysis of cell surface molecule expression
For analysis of T cell activation in cells recovered from spleen and liver, expression of CD44 (Mm.423621), CD62L (Mm.1461), CD69 (Mm.74745), and CD25 (Mm.915) was examined by flow cytometry, after gating on CD4+TCRβ+NK1.1- cells. Cells isolated from spleens and livers were surface labeled with FITC-conjugated antibodies to CD44 or CD69, PE-conjugated antibodies to TCRβ or NK1.1, PerCp-Cy5.5-conjugated antibodies to NK1.1 or CD4, APC-conjugated antibodies to CD62L, and APC-Cy7-conjugated antibodies to CD4 or CD25, (BD Biosciences). For analysis of mononuclear populations, cells were stained with PE-conjugated anti-CD11b (Mm.262106), PerCP-Cy5.5-conjugated anti-Ly6C (Mm.1583) and APC-conjugated anti-CD115 (Mm.22574). Aqua dead cell stain (Invitrogen) was used to discriminate live and dead cells and granulocytes were excluded based on their high side scatter. All samples were analyzed using a LSR II Optical Bench flow cytometer with FACSDiva (BD Biosciences) and Winlist software, version 5.0 (Verity Software House).
Microarray
Livers from naïve female wild type C57BL/6, RAG-1-/-, and OT-II/RAG-1-/- (N = 9 for each genotype) mice were pooled into groups of three and prepared for cDNA microarray analysis. Briefly, RNA was isolated via RNAzol (Tel-Test, Friendswood, TX) and the RNeasy protocol (Qiagen) and analyzed for purity and concentration on a NanoDrop® ND-1000 Spectrophotometer (Wilmington, DE). cDNA was prepared from two 30 µg aliquots of each pooled sample and labeled with either Cy3 or Cy5 fluorescent probes. One 30 µg aliquot from each pool was used to create a background control pool, while the second aliquot was used as the comparative sample. For further isolation and labeling protocol details please refer to http://www.niaid.nih.gov/dir/services/rtb/microarray/protocols.asp. Samples were hybridized as described by Schaupp [78] to Mmbe custom arrays manufactured by the NIAID Microarray Facility. Further information about these arrays can be found at http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GPL1057. Hybridizations were performed in triplicate. Images were scanned by GenePix4000B Scanner (Axon Instruments/Molecular Devices, Sunnyvale, CA) and analyzed using the mAdb program (http://madb.niaid.nih.gov/). Signals were calculated as mean intensity – median background. Data were analyzed using Significance Analysis of Microarrays (SAM), version 2.11 (Stanford University) and student's T tests (EXCEL, with P values <0.05 considered significant) to identify only those genes that exhibited differential expression in RAG-1-/- liver tissue compared to both OT-II/RAG-1-/- and wild type liver tissue (i.e. genes differentially expressed in RAG-1-/- liver tissue compared to either OT-II/RAG-1-/- or wild type alone were not considered specific to the presence of naïve CD4+ T cells). We applied supervised average-linkage hierarchical clustering on differentially regulated genes, as implemented in the program Cluster version 2.11 (M. Eisen; http://www.microarrays.org/software), separately to both the genes and arrays. The results were analyzed, and figures generated, using TreeView version 1.60 (http://www.microarrays.org/software). Initial gene annotation was performed through GoMiner (http://discover.nci.nih.gov/gominer/). Functional analyses and networks of differentially expressed genes were then generated through the use of Ingenuity Pathways Analysis (Ingenuity® Systems, www.ingenuity.com). To identify networks of differentially expressed genes that were functionally related, Ingenuity Pathways Analysis software was used to overlay differentially expressed genes onto a global molecular network developed from information contained in the Ingenuity Pathways Knowledge Base. To facilitate analysis, networks were limited to a size of up to 35 differentially expressed genes. A numerical network score was then calculated for each network, as described under Statistical Analysis below, to determine the probability of obtaining the same network by random chance.
LPS treatment
In RAG-/- mice, APC maturation was induced by biweekly intraperitoneal injection of low doses of ultrapure LPS, E. coli 0111:B4 (InvivoGen, San Diego), at doses of 2 µg or 20 µg LPS/mouse. Monocyte numbers were monitored by flow cytometry, as described above.
Statistical analysis
Because unequal variances were observed among some of the groups analyzed in this study, stringent non-parametric tests were used throughout to test the significance of differences between experimental groups. For two groups, significance of differences between experimental groups was tested using Mann-Whitney tests, and for three groups the significance of differences was tested using Kruskal-Wallis tests followed by Dunns' multiple comparison tests. Statistical analyses were performed with GraphPad Prism Version 4.0 software (GraphPad Software, Inc., San Diego, CA). P values of less than 0.05 were considered significant. Experiments were repeated at least twice, with 4–5 animals per group. For microarray data, Functional Analysis of differentially expressed genes was performed using Ingenuity Pathways Analysis (Ingenuity® Systems, www.ingenuity.com) to identify networks of differentially expressed genes that were functionally related. For putative networks, a numerical score was calculated from the hypergeometric distribution of the network using the right-tailed Fisher's Exact Test. The network score is the negative log of the Fisher's Exact Test P value. The probability that each biological function assigned to the network is due to chance alone was also tested using Fischer's Exact Test. P values of less than 0.05 were considered significant.
Accession numbers
The gene expression data discussed in this manuscript have been deposited in the NCBI's GEO database and are available under GEO series accession number GSE8340.
Supporting Information
Zdroje
1. ColleyDG
LoVerdePT
SavioliL
2001 INFECTIOUS DISEASE: Medical Helminthology in the 21st Century. Science 293 1437 1438
2. MaizelsRM
BundyDA
SelkirkME
SmithDF
AndersonRM
1993 Immunological modulation and evasion by helminth parasites in human populations. Nature 365 797 805
3. DoenhoffM
MusallamR
BainJ
McGregorA
1978 Studies on the host-parasite relationship in Schistosoma mansoni - infected mice: the immunological dependence of parasite egg excretion. Immunology 35 771 778
4. DoenhoffMJ
1997 A role for granulomatous inflammation in the transmission of infectious disease: schistosomiasis and tuberculosis. Parasitology 115 S113 125
5. DaviesSJ
GroganJL
BlankRB
LimKC
LocksleyRM
2001 Modulation of Blood Fluke Development in the Liver by Hepatic CD4+ Lymphocytes. Science 294 1358 1361
6. HerbertDR
HolscherC
MohrsM
ArendseB
SchwegmannA
2004 Alternative macrophage activation is essential for survival during schistosomiasis and downmodulates T helper 1 responses and immunopathology. Immunity 20 623 635
7. BlankRB
LambEW
TochevaAS
CrowET
LimKC
2006 The Common gamma Chain Cytokines Interleukin (IL)-2 and IL-7 Indirectly Modulate Blood Fluke Development via Effects on CD4+ T Cells. J Infect Dis 194 1609 1616
8. DaviesSJ
LimKC
BlankRB
KimJH
LucasKD
2004 Involvement of TNF in limiting liver pathology and promoting parasite survival during schistosome infection. Int J Parasitol 34 27 36
9. DaviesSJ
McKerrowJH
2003 Developmental plasticity in schistosomes and other helminths. Int J Parasitol 33 1277 1284
10. SunZ
ArendtCW
EllmeierW
SchaefferEM
SunshineMJ
2000 PKC-theta is required for TCR-induced NF-kappaB activation in mature but not immature T lymphocytes. Nature 404 402 407
11. RulandJ
DuncanGS
EliaA
del Barco BarrantesI
NguyenL
2001 Bcl10 is a positive regulator of antigen receptor-induced activation of NF-kappaB and neural tube closure. Cell 104 33 42
12. ErnstB
LeeDS
ChangJM
SprentJ
SurhCD
1999 The peptide ligands mediating positive selection in the thymus control T cell survival and homeostatic proliferation in the periphery. Immunity 11 173 181
13. FontenotJD
RasmussenJP
GavinMA
RudenskyAY
2005 A function for interleukin 2 in Foxp3-expressing regulatory T cells. Nat Immunol 6 1142 1151
14. Van ParijsL
RefaeliY
LordJD
NelsonBH
AbbasAK
1999 Uncoupling IL-2 signals that regulate T cell proliferation, survival, and Fas-mediated activation-induced cell death. Immunity 11 281 288
15. ElleryJM
NichollsPJ
2002 Possible mechanism for the alpha subunit of the interleukin-2 receptor (CD25) to influence interleukin-2 receptor signal transduction. Immunol Cell Biol 80 351 357
16. PeschonJJ
MorrisseyPJ
GrabsteinKH
RamsdellFJ
MaraskovskyE
1994 Early lymphocyte expansion is severely impaired in interleukin 7 receptor-deficient mice. J Exp Med 180 1955 1960
17. von Freeden-JeffryU
VieiraP
LucianLA
McNeilT
BurdachSE
1995 Lymphopenia in interleukin (IL)-7 gene-deleted mice identifies IL-7 as a nonredundant cytokine. J Exp Med 181 1519 1526
18. ShreedharV
MoodycliffeAM
UllrichSE
BucanaC
KripkeML
1999 Dendritic cells require T cells for functional maturation in vivo. Immunity 11 625 636
19. CalvanoSE
XiaoW
RichardsDR
FelcianoRM
BakerHV
2005 A network-based analysis of systemic inflammation in humans. Nature 437 1032 1037
20. AuffrayC
SiewekeMH
GeissmannF
2009 Blood monocytes: development, heterogeneity, and relationship with dendritic cells. Annu Rev Immunol 27 669 692
21. LiebermannDA
HoffmanB
2002 Myeloid differentiation (MyD) primary response genes in hematopoiesis. Oncogene 21 3391 3402
22. CrawfordK
StarkA
KitchensB
SternheimK
PantazopoulosV
2003 CD2 engagement induces dendritic cell activation: implications for immune surveillance and T-cell activation. Blood 102 1745 1752
23. ZhaoC
ChenA
JamiesonCH
FereshtehM
AbrahamssonA
2009 Hedgehog signalling is essential for maintenance of cancer stem cells in myeloid leukaemia. Nature 458 776 779
24. LiuXS
LiXH
WangY
ShuRZ
WangL
2007 Disruption of palladin leads to defects in definitive erythropoiesis by interfering with erythroblastic island formation in mouse fetal liver. Blood 110 870 876
25. ToddDJ
LeeAH
GlimcherLH
2008 The endoplasmic reticulum stress response in immunity and autoimmunity. Nat Rev Immunol 8 663 674
26. DavenportEL
MorganGJ
DaviesFE
2008 Untangling the unfolded protein response. Cell Cycle 7 865 869
27. EastonDP
KanekoY
SubjeckJR
2000 The hsp110 and Grp1 70 stress proteins: newly recognized relatives of the Hsp70s. Cell Stress Chaperones 5 276 290
28. RaoRV
NiaziK
MollahanP
MaoX
CrippenD
2006 Coupling endoplasmic reticulum stress to the cell-death program: a novel HSP90-independent role for the small chaperone protein p23. Cell Death Differ 13 415 425
29. LeeAH
IwakoshiNN
GlimcherLH
2003 XBP-1 regulates a subset of endoplasmic reticulum resident chaperone genes in the unfolded protein response. Mol Cell Biol 23 7448 7459
30. LabunskyyVM
YooMH
HatfieldDL
GladyshevVN
2009 Sep15, a thioredoxin-like selenoprotein, is involved in the unfolded protein response and differentially regulated by adaptive and acute ER stresses. Biochemistry 48 8458 8465
31. AlanenHI
WilliamsonRA
HowardMJ
HatahetFS
SaloKE
2006 ERp27, a new non-catalytic endoplasmic reticulum-located human protein disulfide isomerase family member, interacts with ERp57. J Biol Chem 281 33727 33738
32. LiangJ
YinC
DoongH
FangS
PeterhoffC
2006 Characterization of erasin (UBXD2): a new ER protein that promotes ER-associated protein degradation. J Cell Sci 119 4011 4024
33. YamasakiS
YagishitaN
NishiokaK
NakajimaT
2007 The roles of synoviolin in crosstalk between endoplasmic reticulum stress-induced apoptosis and p53 pathway. Cell Cycle 6 1319 1323
34. KoongAC
ChauhanV
Romero-RamirezL
2006 Targeting XBP-1 as a novel anti-cancer strategy. Cancer Biol Ther 5 756 759
35. KondoS
SaitoA
HinoS
MurakamiT
OgataM
2007 BBF2H7, a novel transmembrane bZIP transcription factor, is a new type of endoplasmic reticulum stress transducer. Mol Cell Biol 27 1716 1729
36. KanekoH
IgarashiK
KataokaK
MiuraM
2005 Heat shock induces phosphorylation of histone H2AX in mammalian cells. Biochem Biophys Res Commun 328 1101 1106
37. CunhaDA
LadriereL
OrtisF
Igoillo-EsteveM
GurzovEN
2009 GLP-1 agonists protect pancreatic {beta}-cells from lipotoxic endoplasmic reticulum stress through upregulation of BiP and JunB. Diabetes
38. RinconM
DavisRJ
2009 Regulation of the immune response by stress-activated protein kinases. Immunol Rev 228 212 224
39. IwakoshiNN
PypaertM
GlimcherLH
2007 The transcription factor XBP-1 is essential for the development and survival of dendritic cells. J Exp Med 204 2267 2275
40. SjinRM
KrishnarajuK
HoffmanB
LiebermannDA
2002 Transcriptional regulation of myeloid differentiation primary response (MyD) genes during myeloid differentiation is mediated by nuclear factor Y. Blood 100 80 88
41. ItohT
SatohM
KannoE
FukudaM
2006 Screening for target Rabs of TBC (Tre-2/Bub2/Cdc16) domain-containing proteins based on their Rab-binding activity. Genes Cells 11 1023 1037
42. MenetreyJ
PerderisetM
CicolariJ
DuboisT
ElkhatibN
2007 Structural basis for ARF1-mediated recruitment of ARHGAP21 to Golgi membranes. Embo J 26 1953 1962
43. HabermannB
2004 The BAR-domain family of proteins: a case of bending and binding? EMBO Rep 5 250 255
44. BolisA
CovielloS
VisigalliI
TaveggiaC
BachiA
2009 Dlg1, Sec8, and Mtmr2 regulate membrane homeostasis in Schwann cell myelination. J Neurosci 29 8858 8870
45. Colucci-GuyonE
NiedergangF
WallarBJ
PengJ
AlbertsAS
2005 A role for mammalian diaphanous-related formins in complement receptor (CR3)-mediated phagocytosis in macrophages. Curr Biol 15 2007 2012
46. SunderkotterC
NikolicT
DillonMJ
Van RooijenN
StehlingM
2004 Subpopulations of mouse blood monocytes differ in maturation stage and inflammatory response. J Immunol 172 4410 4417
47. ChituV
StanleyER
2006 Colony-stimulating factor-1 in immunity and inflammation. Curr Opin Immunol 18 39 48
48. ShiC
SimonDI
2006 Integrin signals, transcription factors, and monocyte differentiation. Trends Cardiovasc Med 16 146 152
49. TieuBC
LeeC
SunH
LejeuneW
RecinosA3rd
2009 An adventitial IL-6/MCP1 amplification loop accelerates macrophage-mediated vascular inflammation leading to aortic dissection in mice. J Clin Invest 119 3637 3651
50. ZanoniI
OstuniR
CapuanoG
ColliniM
CacciaM
2009 CD14 regulates the dendritic cell life cycle after LPS exposure through NFAT activation. Nature 460 264 268
51. GuhaM
MackmanN
2001 LPS induction of gene expression in human monocytes. Cell Signal 13 85 94
52. LockyerAE
OlsonPD
OstergaardP
RollinsonD
JohnstonDA
2003 The phylogeny of the Schistosomatidae based on three genes with emphasis on the interrelationships of Schistosoma Weinland, 1858. Parasitology 126 203 224
53. HarrisonRA
DoenhoffMJ
1983 Retarded development of Schistosoma mansoni in immunosuppressed mice. Parasitology 86 429 438
54. LambEW
CrowET
LimKC
LiangYS
LewisFA
2007 Conservation of CD4+ T cell-dependent developmental mechanisms in the blood fluke pathogens of humans. Int J Parasitol 37 405 415
55. DoenhoffMJ
SabahAA
FletcherC
WebbeG
BainJ
1987 Evidence for an immune-dependent action of praziquantel on Schistosoma mansoni in mice. Trans R Soc Trop Med Hyg 81 947 951
56. MurailleE
De TrezC
PajakB
BraitM
UrbainJ
2002 T cell-dependent maturation of dendritic cells in response to bacterial superantigens. J Immunol 168 4352 4360
57. TalayO
ShenCH
ChenL
ChenJ
2009 B7-H1 (PD-L1) on T cells is required for T-cell-mediated conditioning of dendritic cell maturation. Proc Natl Acad Sci U S A 106 2741 2746
58. AkiraS
UematsuS
TakeuchiO
2006 Pathogen recognition and innate immunity. Cell 124 783 801
59. TsujimotoH
OnoS
EfronPA
ScumpiaPO
MoldawerLL
2008 Role of Toll-like receptors in the development of sepsis. Shock 29 315 321
60. SerbinaNV
JiaT
HohlTM
PamerEG
2008 Monocyte-mediated defense against microbial pathogens. Annu Rev Immunol 26 421 452
61. David DongZM
AplinAC
NicosiaRF
2009 Regulation of angiogenesis by macrophages, dendritic cells, and circulating myelomonocytic cells. Curr Pharm Des 15 365 379
62. SerbinaNV
Salazar-MatherTP
BironCA
KuzielWA
PamerEG
2003 TNF/iNOS-producing dendritic cells mediate innate immune defense against bacterial infection. Immunity 19 59 70
63. AhmedSF
OswaldIP
CasparP
HienyS
KeeferL
1997 Developmental differences determine larval susceptibility to nitric oxide-mediated killing in a murine model of vaccination against Schistosoma mansoni. Infect Immun 65 219 226
64. WynnTA
OswaldIP
EltoumIA
CasparP
LowensteinCJ
1994 Elevated expression of Th1 cytokines and nitric oxide synthase in the lungs of vaccinated mice after challenge infection with Schistosoma mansoni. J Immunol 153 5200 5209
65. AmiriP
LocksleyRM
ParslowTG
SadickM
RectorE
1992 Tumour necrosis factor alpha restores granulomas and induces parasite egg-laying in schistosome-infected SCID mice [see comments]. Nature 356 604 607
66. PapaS
BubiciC
ZazzeroniF
FranzosoG
2009 Mechanisms of liver disease: cross-talk between the NF-kappaB and JNK pathways. Biol Chem 390 965 976
67. SekiE
BrennerDA
2008 Toll-like receptors and adaptor molecules in liver disease: update. Hepatology 48 322 335
68. LambertucciJR
RayesAA
SerufoJC
Gerspacher-LaraR
Brasileiro FilhoG
1998 Schistosomiasis and associated infections. Mem Inst Oswaldo Cruz 93 Suppl 1 135 139
69. RochaH
KirkJW
HeareyCDJr
1971 Prolonged Salmonella bacteremia in patients with Schistosoma mansoni infection. Arch Intern Med 128 254 257
70. van RietE
HartgersFC
YazdanbakhshM
2007 Chronic helminth infections induce immunomodulation: consequences and mechanisms. Immunobiology 212 475 490
71. BarndenMJ
AllisonJ
HeathWR
CarboneFR
1998 Defective TCR expression in transgenic mice constructed using cDNA-based alpha - and beta-chain genes under the control of heterologous regulatory elements. Immunol Cell Biol 76 34 40
72. MurphyKM
HeimbergerAB
LohDY
1990 Induction by antigen of intrathymic apoptosis of CD4+CD8+TCRlo thymocytes in vivo. Science 250 1720 1723
73. SederRA
PaulWE
DavisMM
Fazekas de St GrothB
1992 The presence of interleukin 4 during in vitro priming determines the lymphokine-producing potential of CD4+ T cells from T cell receptor transgenic mice. J Exp Med 176 1091 1098
74. SmithersSR
TerryRJ
1965 The infection of laboratory hosts with cercariae of Schistosoma mansoni and the recovery of the adult worms. Parasitology 55 695 700
75. HernandezDC
LimKC
McKerrowJH
DaviesSJ
2004 Schistosoma mansoni: sex-specific modulation of parasite growth by host immune signals. Exp Parasitol 106 59 61
76. ColleyDG
CookJA
FreemanGLJr
BartholomewRK
JordanP
1977 Immune responses during human schistosomiasis mansoni. I. In vitro lymphocyte blastogenic responses to heterogeneous antigenic preparations from schistosome eggs, worms and cercariae. Int Arch Allergy Appl Immunol 53 420 433
77. GazzinelliG
KatzN
RochaRS
ColleyDG
1983 Immune responses during human schistosomiasis mansoni. X. Production and standardization of an antigen-induced mitogenic activity by peripheral blood mononuclear cells from treated, but not active cases of schistosomiasis. J Immunol 130 2891 2895
78. SchauppCJ
JiangG
MyersTG
WilsonMA
2005 Active mixing during hybridization improves the accuracy and reproducibility of microarray results. Biotechniques 38 117 119
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2010 Číslo 4
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- The Effect of Vaccination on the Evolution and Population Dynamics of Avian Paramyxovirus-1
- Reconstitution of SARS-Coronavirus mRNA Cap Methylation
- Increased Monocyte Turnover from Bone Marrow Correlates with Severity of SIV Encephalitis and CD163 Levels in Plasma
- Deficiencies in Jasmonate-Mediated Plant Defense Reveal Quantitative Variation in Pathogenesis
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy