
The CYCLIN-A CYCA1;2/TAM Is Required for the Meiosis I to Meiosis II Transition and Cooperates with OSD1 for the Prophase to First Meiotic Division Transition
Meiosis halves the chromosome number because its two divisions follow a single round of DNA replication. This process involves two cell transitions, the transition from prophase to the first meiotic division (meiosis I) and the unique meiosis I to meiosis II transition. We show here that the A-type cyclin CYCA1;2/TAM plays a major role in both transitions in Arabidopsis. A series of tam mutants failed to enter meiosis II and thus produced diploid spores and functional diploid gametes. These diploid gametes had a recombined genotype produced through the single meiosis I division. In addition, by combining the tam-2 mutation with AtSpo11-1 and Atrec8, we obtained plants producing diploid gametes through a mitotic-like division that were genetically identical to their parents. Thus tam alleles displayed phenotypes very similar to that of the previously described osd1 mutant. Combining tam and osd1 mutations leads to a failure in the prophase to meiosis I transition during male meiosis and to the production of tetraploid spores and gametes. This suggests that TAM and OSD1 are involved in the control of both meiotic transitions.
Published in the journal:
. PLoS Genet 6(6): e32767. doi:10.1371/journal.pgen.1000989
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1000989
Summary
Meiosis halves the chromosome number because its two divisions follow a single round of DNA replication. This process involves two cell transitions, the transition from prophase to the first meiotic division (meiosis I) and the unique meiosis I to meiosis II transition. We show here that the A-type cyclin CYCA1;2/TAM plays a major role in both transitions in Arabidopsis. A series of tam mutants failed to enter meiosis II and thus produced diploid spores and functional diploid gametes. These diploid gametes had a recombined genotype produced through the single meiosis I division. In addition, by combining the tam-2 mutation with AtSpo11-1 and Atrec8, we obtained plants producing diploid gametes through a mitotic-like division that were genetically identical to their parents. Thus tam alleles displayed phenotypes very similar to that of the previously described osd1 mutant. Combining tam and osd1 mutations leads to a failure in the prophase to meiosis I transition during male meiosis and to the production of tetraploid spores and gametes. This suggests that TAM and OSD1 are involved in the control of both meiotic transitions.
Introduction
Meiosis is a central feature in the reproductive program of all sexually reproducing eukaryotes. The process of meiosis involves two rounds of chromosome segregation that follow a single round of chromosome duplication leading to the production of haploid gametes. Meiosis differs from mitosis, somatic cell division, in a number of ways. In meiosis:
-
Homologous chromosomes pair and closely associate along a proteinaceous structure called the synaptonemal complex (SC). This process culminates at a substage of prophase called pachytene.
-
Crossovers occur between homologs during prophase.
-
Homologous chromosomes separate at anaphase of the first division. To ensure accurate chromosome segregation at anaphase I each homolog must remain connected to the other through metaphase I. Since the SC disappears before the end of prophase, it cannot ensure linkage of homologs at metaphase I. This connection is maintained until anaphase I by chiasmata, the cytological manifestation of crossovers.
-
Meiosis has a second division with no intervening DNA synthesis. Sister chromatids, resulting from the meiotic S phase, remain associated until metaphase II and are separated from each other at anaphase II, leading to the production of four haploid spores [1], [2].
To generate haploid spores the meiocyte must enter meiosis I, pass through the meiosis I to meiosis II transition and exit meiosis II. Errors in these transitions, are not uncommon and may lead to parthenogenesis or teratoma formation, or to the production of gametes with the somatic number of chromosomes (2n gametes) [3], [4]. The formation of 2n gametes is thought to be an important mechanism for generating polyploids. Polyploidy has played a key role in the evolution of many fungal, plant, invertebrate and vertebrate lineages, and is particularly frequent in plants [5], [6]. 2n gametes are also an important tool for plant breeding [7].
Cyclins and cyclin-dependent kinases (Cdk) form complexes that are essential for progression through both the mitotic and meiotic cell cycles. The transition from meiosis I to meiosis II requires a fine balance in Cyclin–Cdk activity: it must be sufficiently low to exit meiosis I but must nonetheless be maintained at a level sufficiently high to suppress DNA replication and promote entry into meiosis II [8], [9]. Precisely how the mitotic machinery is modified for the purpose of meiosis is not fully understood. Most of the knowledge currently available originates from studies carried out in unicellular fungi, Xenopus laevis or mouse oocyte systems.
In oocytes, entry into both meiosis I and meiosis II is driven by Cdc2/Cyclin B complexes (the molecular components of the maturation promoting factor, MPF) [10]. The rate of Cyclin B synthesis and degradation by the APC (anaphase promoting complex) determines the timing of the transitions occurring during meiosis [10], [11]. At the end of meiosis I, Cyclin B is only partially degraded [12] and the residual, low level of Cdc2/CyclinB activity is essential for entry into meiosis II [13]. This partial Cyclin B degradation is fine-tuned by the Erp1/Emi2 APC inhibitor [14]–[16]. In Schizosaccharomyces pombe, Mes1 is a key player in the meiosis I to meiosis II transition. Like Erp1/Emi2, the Mes1 protein partially inhibits cyclin degradation by the anaphase promoting complex (APC), thereby allowing entry into meiosis II [17], [18]. In Saccharomyces cerevisiae, the simultaneous deletion of two of the six B-type cyclins results in a single reductional division during meiosis, with the production of two-spored asci (dyads), suggesting that specialist cyclins may be responsible for mediating the meiosis I to meiosis II transition [19]–[21].
Very little is known about control of the meiotic cell cycle in plants. In maize, the elongate mutant produces diploid female gametes because it is unable to undergo female meiosis II, but the corresponding gene has not been identified [22]. The only gene involved in the meiosis I to meiosis II transition isolated to date is the Arabidopsis OSD1 (OMISSION OF SECOND DIVISION) gene, the molecular function of which remains unknown. As osd1 mutants fail to enter the second division in both male and female meiosis, functional 2n gametes and tetraploid progeny are produced [23]. To look for other regulators of meiotic progression, we focused our attention on cyclins. The Arabidopsis thaliana genome contains 10 A-type and 11 B-type cyclins but functions in the cell cycle have been identified for only a few of these molecules, probably because redundancy attenuates the effects of mutations in single cyclin genes [24], [25]. A noticeable exception to this rule is CYCA1;2, also known as TAM (TARDY ASYNCHRONOUS MEIOSIS). In the tam-1 mutant, a single substitution of an amino acid (Thr283-Ile) slows cell cycle progression during male meiosis [26], [27]. Here, we revisit the function of CYCA1;2/TAM by isolating a series of alleles, including null alleles, and show that CYCA1;2/TAM is crucial for the meiosis I to meiosis II transition. The tam mutants fail to enter meiosis II, leading to the production of dyads of spores and diploid gametes that have experienced genetic exchange. This demonstrates that the meiosis I to meiosis II transition in plants involves a non redundant Cyclin-cdk activity. Furthermore, combining tam and osd1 mutations leads to a failure of the transition from prophase to the first meiotic division (meiosis I) during male meiosis and to the production of single-spore meiotic products and tetraploid pollen grains. This shows that CYCA1;2/TAM and OSD1 are also involved in the prophase to meiosis I transition. The implications of these results for the control of meiotic transitions are discussed.
In addition, taking advantage of a genetic strategy we used previously to investigate OSD1 function [23], we combined tam mutants with mutations that affect homologous recombination (Atspo11-1) and chromosome segregation (Atrec8), essentially converting meiosis into a mitotic-like division. The tam/Atspo11-1/Atrec8 line, called MiMe-2, produces diploid gametes that are genetically identical to their parents, mimicking apomeiosis, a key element of apomixis or asexual reproduction through seeds. However, MiMe-2 also resulted in the production of diploid and aneuploid gametes, probably due to the lower penetrance of tam mutations compared to osd1 mutations.
Results
The tam Mutants Produce Diploid Gametes
The Arabidopsis TAM (Tardy Asynchronous Meiosis) gene has been implicated in the control of meiotic progression [26], [27]. The tam-1 mutant displays slower cell cycle progression during male meiosis, although it does eventually, like wild type, produce tetrads. However, previous to this study, only one mutant allele with a point mutation leading to a single amino-acid substitution in the protein had been studied. As this point mutation is temperature-sensitive and probably corresponds to a hypomorphic allele, we revisited the role of the TAM gene by isolating and characterizing three independent insertion mutants from public mutant collections and three additional point mutations (Figure 1A and Figure S1). The tam-2 (sail_505_C06 [28], [29]) T-DNA insertion is in the fourth exon (ATG+1130 pb) and is accompanied by a large deletion (corresponding to half of the TAM coding sequence, the entire TAM promoter region and the first 52 bp of the next gene, At1 g77400), the tam-3 (SALK_080686 [30]) T-DNA insertion is in the seventh intron (ATG+1690 bp) and the tam-4 (CSHL_ET12273 [31]) Ds insertion is in the first exon (ATG+62 bp). The tam-2 and tam-3 mutations are in the Columbia (Col-0) background, whereas the tam-4 mutation is in the Landsberg erecta (Ler) background (Figure 1A). Additionally, three point mutations were isolated in an ethyl methanesulfonate (EMS) chemical mutagenesis screen. The tam-5 mutation is a G to A transition at nucleotide 378 in the coding sequence that creates a stop codon (TGG → TAG; Trp77 → stop). The tam-6 mutation is also a G to A transition at the -1 position of the 3′ splice site between intron 2 and exon 3. The tam-5 and tam-6 alleles are both recessive and do not complement one another. The tam-7 mutation is a G to A transition at position -4 of the 3′ splice site between intron 1 and exon 2. Similar splice site proximal mutations have been observed in other Arabidopsis mutants including det1-1 and det1-3 [32]. The tam-7 allele is dominant and thus could not be used for genetic complementation of the other tam alleles. All three EMS tam alleles are in the Columbia (Col-0) background. The T-DNA alleles were used preferentially for detailed analysis.

The tam mutants displayed no defects during somatic development. In Arabidopsis, male meiosis produces a group of four spores, organized in a tetrahedron, called a tetrad (Figure 1B). Each spore gives rise to a pollen grain. In these six independent tam mutants, the products of male meiosis were mostly dyads of spores instead of tetrads (Figure 1B and Table 1). In addition to balanced dyads the stronger mutants also produced triads and unbalanced products together with a very small number of tetrads. Complementation tests with tam-2, tam-3 and tam-4 mutants confirmed that these mutations were allelic. Unlike the previously described temperature sensitive tam-1 mutant, these six mutants expressed the dyad phenotype at normal growing temperatures (20°C). Furthermore, the dyad stage in tam-1 does not appear to be terminal, as meiosis always progresses to tetrad production [27], whereas the tam-2, tam-3, tam-4, tam-5, tam-6 and tam-7 systematically produced mostly dyads. This suggests that the tam-1 mutant presents only a delay in the progression of meiosis, whereas the other mutants do not progress beyond the dyad stage (as confirmed by pollen analysis, see below). This phenotype is reminiscent of the phenotype of the osd1 mutant, which produces spores directly after meiosis I. Its gametes are thus diploid and its offspring polyploid. Thus, we determined ploidy levels among the offspring of diploid tam-2 and tam-4 mutants. In the progeny of selfed homozygous mutants, we observed tetraploids, triploids, and occasionally diploid plants (Table 2). If pollen from tam-2 or tam-4 mutant plants was used to fertilize a wild-type plant, almost all the resulting progeny were triploid, with only a few diploid plants identified (Table 2). If tam-2 or tam-4 mutant ovules were fertilized with wild-type pollen grains we isolated diploid and triploid plants (Table 2). Thus, the frequency of diploid spores, resulting in functional gametes, was high in the tam-2 and tam-4 mutants, for both the male (∼90%) and female (∼30%) lineages. Tam mutants produced only slightly fewer seeds than the corresponding wild-type lines (36±4 seeds/silique in tam-2; 42±4 in wild type Col-0; 43±4 in tam-4; 52±3 in wild type Ler), but many (>50%) of the seeds produced by tam-2 mutants and a few (<10%) of those produced by tam-4 mutants were shriveled. This finding was not unexpected, because tam mutants produce triploid seeds with an excess of the paternal genome that is associated with the production of shriveled seed, particularly in the Col-0 background [33]. The germination rate of seeds produced by tam-2 was 63% (n = 264), suggesting that the proportion of triploid seeds may be underestimated in tam-2 progeny.


The tam Mutants Skip the Second Division of Meiosis
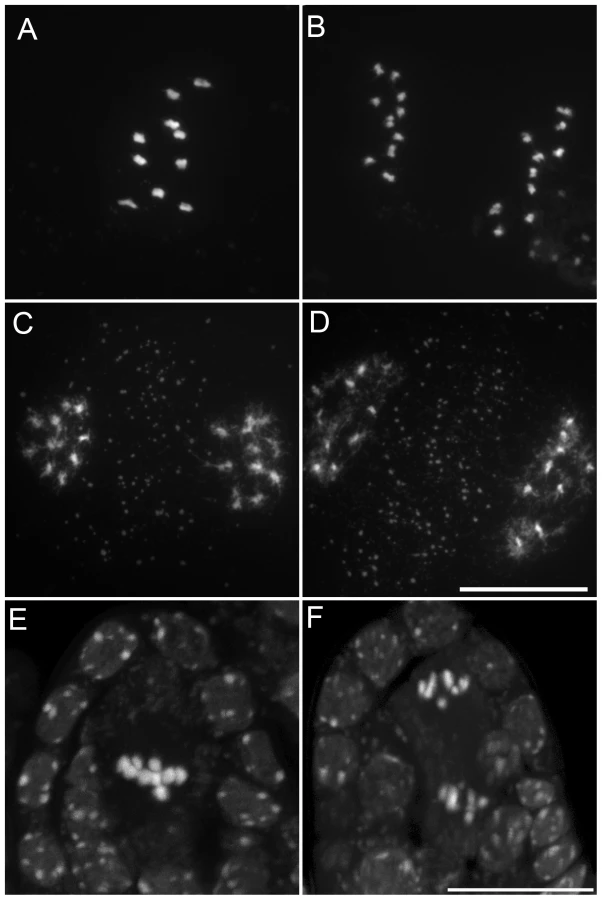
We characterized the mechanisms underlying dyad production in tam mutants, by investigating chromosome behavior during male meiosis using a meiocyte spreading technique (Figure 2 and Figure 3). In wild type, the ten chromosomes appeared as threads at leptotene, underwent synapsis at zygotene and were fully synapsed, along the SC at pachytene (Figure 2A). After the disappearance of the SC at diplotene the resulting five bivalents condensed, revealing the presence of chiasmata (Figure 2B). The bivalents organized on the metaphase I plate (Figure 2C) and homologs segregated to opposite poles at anaphase I (Figure 2D and 2E). The two sets of five homologs aligned on the two metaphase II plates (Figure 2F). The second round of segregation at anaphase II (Figure 2G) led to the formation of four sets of five chromosomes, that decondensed to form the spore nuclei (Figure 2H). Meiosis I in tam mutants was indistinguishable from that in the wild type. All stages of prophase were observed, including full synapsis at pachytene (Figure 3A) and chiasmata at diakinesis (Figure 3B). The bivalents observed at diakinesis, condensed and aligned on the metaphase plate (Figure 3C). We quantified the chiasma frequency by studying the shape of metaphase I bivalents, as described by [34], [35], and found no difference between tam-2 (9±0.8 chiasmata per cell, n = 70) and the wild type (9.1±1 chiasmata per cell, n = 53). The bivalents segregated at anaphase I and decondensed at telophase I (Figure 3D–3F). However, we found no meiosis II figures (among >1000 male meiocytes for each tam-2, tam-3 and tam-4, from prophase to spore formation), consistent with the dyad production in tam mutants resulting from the absence of a second meiotic division. Both typical meiosis I and meiosis II figures were observed during female meiosis (Compare Figure 4 and Figure 5), consistent with the observed production of ∼70% haploid female gametes in tam mutants and the notion that female diploid megaspores are also produced by skipping the second meiotic division.




If diploid gametes are indeed generated by skipping the second meiotic division, any parental heterozygosity at centromeres should be lost in the tam diploid gametes, because sister centromeres cosegregate at division I. Conversely, we would also expect heterozygosity to increase towards telomeres, due to recombination between the locus concerned and the centromere. We tested this hypothesis in two ways, using the fluorescent tagged line (FTL) Arabidopsis tetrads system providing direct information about the genetic content of pollen grains (Figure 6, Figure 7, Figure 8) and by genotyping male and female tam-2 triploid offspring for polymorphic molecular markers (Figure 8) [23], [36], [37].


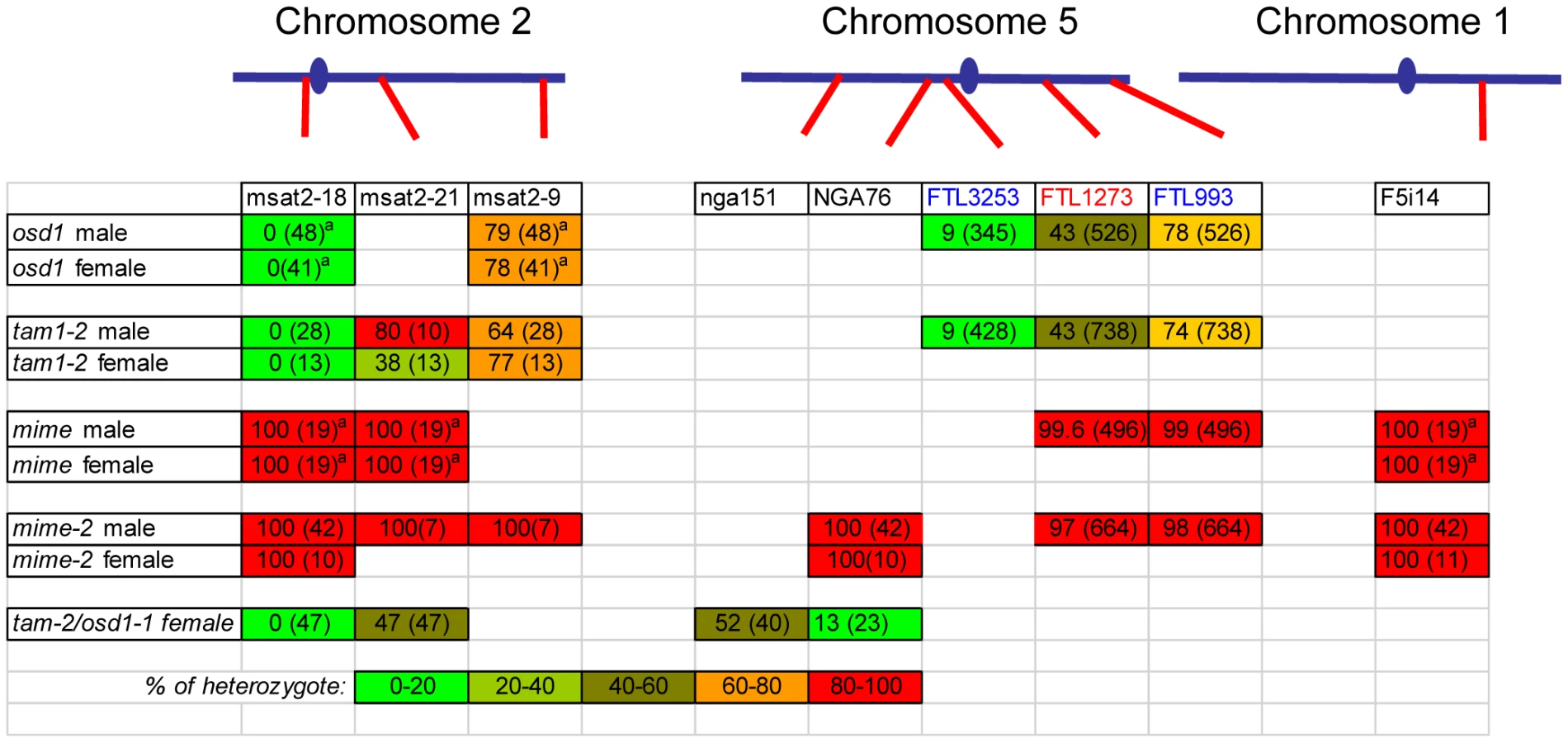
The FTL system is a visual assay based on the use of reporter constructs encoding fluorescent proteins produced in the pollen of the quartet mutant (qrt1-2) [38]. In this mutant, the pollen grains from each meiosis remain physically attached. We carried out crosses to generate plants with both the qrt1-2 and tam-2 mutations, heterozygous for one or two reporter transgenes conferring pollen fluorescence. As controls, we used both wild type and osd1 mutant plants. We first used an FTL transgene located close to the centromere on chromosome 5 (FTL3253 encoding AmCyan) [39]. In wild-type (qrt1-2 background) control plants, all tetrads consisted of two fluorescent and two non-fluorescent pollen grains (Figure 6A), reflecting the segregation of the four chromatids, two of which carried the transgene (n = 120). In the osd1-1 and tam-2 mutants, we observed mostly dyads of pollen (Figure 6B–6E). Furthermore, in both mutants, the vast majority of dyads (91%, n = 526 and n = 738, in osd1-1 and tam-2 respectively) consisted of one fluorescent pollen grain and one non-fluorescent pollen grain (Figure 6B and 6C). Thus, in these dyads, one of the pollen grains contained the two sister chromatids carrying the transgene, whereas the other pollen grain inherited the other two sister chromatids. This segregation pattern is fully consistent with the absence of a second meiotic division. In a small proportion of dyads (9% in both osd1-1 and tam-2 mutants) both pollen grains were fluorescent, indicating recombination between the transgene and the centromere (Figure 6D and 6E). We then carried out the same experiment with two linked FTL transgenes located at some distance from the centromere on chromosome 5 (FTL1273 DsRed2, FTL993 ECFP) [39]. In the wild type (qrt1-2 background), we observed tetrads without (Figure 7A) and with (Figure 7B) recombination of the markers as described in a previous study [36]. The frequency of recombination between the two markers, measured as the percentage of non-parental chromatids deduced from the fluorescence distribution, was 32%. In both the osd1-1 and tam-2 mutants, we observed segregation patterns in the dyads reflecting crossing over between the two markers (29% (n = 262) and 22% (n = 367), respectively) and between the markers and the centromere (22% and 22%, respectively) (Figure 7C–7J). The frequency of heterozygosity at the FTL marker loci in diploid pollen grains was determined (Figure 8). The frequency of heterozygosity was low next to the centromere and increased with distance from the centromere. All these findings are consistent with the tam and osd1 diploid pollen grains resulting from a single first division of meiosis that includes recombination, with no second division [23].
For confirmation of this genetic analysis, we genotyped triploid offspring generated from male and female gametes from the tam-2 mutant. We first introduced genetic polymorphisms into tam-2 (Col-0 background) by crossing this mutant to the No-0 accession. In the F2 generation, we used PCR to select plants homozygous for the tam-2 mutation but heterozygous for a series of microsatellite markers. These plants were crossed, as male or female parents, with plants from a third genetic background (Landsberg erecta, Ler). Genotyping of the resultant triploid progeny for the trimorphic molecular markers revealed the genetic make-up of the 2n gametes produced by the mutant. The Ler allele was present in all the plants (brought by the wild type Ler haploid gamete), while the presence/absence of the Col-0/No-0 allele in the triploids corresponds to the genotype of the 2N mutant gametes. All the diploid gametes tested had the predicted genetic characteristics, similar to those of the diploid gametes produced by the osd1 mutant (Figure 8). The markers were homozygous at the centromere, but displayed segregation at other loci, due to recombination. These results confirmed that the absence of a second meiotic division was indeed responsible for the production of both male and female 2n gametes in tam-2.
Turning Meiosis into Mitosis
Our tam alleles displayed phenotypes very similar to that of the osd1 mutant, with no second meiotic division, leading to the production of viable diploid male and female gametes. Thus, tam diploid gametes differ from apomeiotic (mitosis-like) gametes in that they are genetically different from the mother plant [40], [41]. We have previously shown that, by combining the osd1 mutation with the Atspo11-1 mutant, which eliminates recombination, and the Atrec8 mutation, which modifies chromatid segregation, it is possible to convert meiosis into a mitosis-like division (apomeiosis). We called this triple mutant MiMe for “mitosis instead of meiosis” [23]. We investigated whether tam mutation could replace the osd1 mutation to convert meiosis into mitosis, by constructing the tam-2/Atspo11-1/Atrec8 triple mutant, which we named MiMe-2. During male meiosis, MiMe-2 plants mostly generated dyads (91.2% 446/489), with only a small number of unbalanced products. Observations of chromosome behavior during male and female meiosis showed that the division process resembled mitosis: 10 univalents aligned on the metaphase plate, with the separation of sister chromatids at anaphase (Figure 9). The selfed progeny of MiMe-2 plants was mostly tetraploid, with some aneuploids (Table 2). Backcrossing MiMe-2 plants, as the female parent, onto wild-type plants resulted mostly in triploids with a few aneuploid plants (Table 2), whereas backcrossing MiMe plants, as the male parent, onto the wild type, generated a mixture of mostly triploid plants, diploid and aneuploid plants (Table 2). Thus, the mitosis-like division observed in MiMe-2 plants gives rise to functional diploid gametes, together with small numbers of haploid and aneuploid gametes, in both the male and female lineages. MiMe-2 plants also displayed lower levels of fertility than either the wild-type or the tam-2 mutant (wild type: 42±4 seeds/fruit; tam-2: 37±4, MiMe2: 20±3). This finding was not unexpected, as the tam mutation does not display full penetrance. In meiocytes lacking Atrec8 and Atspo11 that undergo a second division, this division is unbalanced, probably leading to the production of aneuploid gametes in MiMe-2 plants [42]. We analyzed the genetic content of the MiMe-2 diploid gametes, using both the FTL lines and molecular markers (Figure 7K and 7L and Figure 8). We introduced the same FTL transgenes as described above (FTL1273 DsRed2, FTL993 ECFP) into MiMe and MiMe-2 plants. Almost all the pollen grains of both genotypes displayed both types of fluorescence (Figure 7K and 7L; 98% and 95%, respectively), indicating that they had inherited both transgenes, and confirming the occurrence of a mitosis-like division, rather than meiosis, in both lines. The few cases in which the two pollen grains were not expressing both fluorescent proteins may be explained by chromosome missegregation or occasional extinction of the transgenes. In addition, all the diploid MiMe-2 gametes (male and female) systematically retained heterozygosity for each genetic marker tested (Figure 8). They were thus genetically identical to the mother plant. These results confirm that MiMe-2 plants undergo a mitosis-like division instead of a normal meiotic division, generating gametes genetically identical to the parent plant, but at a lower regularity than MiMe plants [23].
tam/osd1 Double Mutant Skip Division II at Female Meiosis and Skip Both Divisions at Male Meiosis
Our tam alleles displayed phenotypes very similar to that of the osd1 mutant, with no second meiotic division, leading to the production of viable diploid male and female gametes. We then combined tam-2 and osd1-1 mutations. The double mutant was almost sterile, producing very few seeds by selfing. Reciprocal crosses with wild type revealed that tam-2/osd1-1 double mutant was female fertile but male sterile. If tam-2/osd1-1 mutant ovules were fertilized with wild-type pollen grains we isolated almost exclusively triploid plants (Table 2). Observation of female meiosis in the double mutant revealed normal meiosis I but an absence of meiosis II (Figure 10). The genetic analysis of the female gametes, performed as above, showed that all the diploid ovules tested had the predicted genetic characteristics for an absence of second meiotic division (Figure 8). These results show that tam-2/osd1-1 double mutants display the same female meiosis phenotype as single mutants, with female meiocytes failing to enter meiosis II, leading to the production of 2n ovules.

We investigated the origin of the male sterility phenotype observed in tam-2/osd1-1 double mutant plants, by assessing pollen viability [43]. Figure 11A shows a wild-type anther treated with Alexander stain which produces a red pigment in viable pollen. Anthers of tam-2 and osd1-1 single mutants contained viable pollen grains, although less numerous and slightly bigger than wild type (Figure 11B and 11C). In contrast, tam-2/osd1-1 anther contained very few pollen grains (9±11) with variable size (Figure 11D–11F). In wild type, meiosis produces four spores (Figure 12A), whereas both tam and osd1 mutants produce dyads of spores (Figure 12B and 12C). In contrast, observation of male meiotic products in tam-2/osd1-1 revealed only “monads”, with a single-spore product (Figure 12D, n = 498). We then investigated the behavior of male meiotic chromosomes in tam-2/osd1-1 mutants (Figure 13). Prophase was indistinguishable from wild type: the ten chromosomes appeared as threads at leptotene, underwent synapsis at zygotene and were fully synapsed at pachytene. After the disappearance of the SC at diplotene, the resulting five bivalents condensed, revealing the presence of chiasmata. However, we observed spores with a single nucleus (Figure 13E and 13F), consistent with the observation of monads. Furthermore, only two figures typical of metaphase/anaphase I and no figures of telophase I or meiosis II were found among >1600 meiocytes. This suggests that most male meiocytes skip both meiosis I and II and produce spores directly after replication and prophase, without chromosome segregation. An expected consequence of such a defect is the production of 4n pollen grains. We did not succeed in crossing tam-2/osd1-1 mutants as male but we determined ploidy levels among the seeds produced by selfing and found a large proportion of 6n plants in the progeny of 2n plants (Table 2). As crosses with wild type showed that tam-2/osd1-1 produce 2n ovules, the occurrence of 6n plants strongly suggests that 4n pollen grains are produced, in accordance with the skipping of both rounds of segregation at male meiosis in tam-2/osd1-1. A large proportion of 4n plants is also found in the selfed progeny showing that tam-2/osd1-1 also produces 2n pollen grains. These 2n pollen grains likely originate from the few meiocytes that enter meiosis I and produces 2n spores, that later outcompete 4n pollen.



Discussion
A Cyclin A Mediates the Meiosis I to Meiosis II Transition
We show here that one of the ten known type A cyclins in Arabidopsis, CYCA1;2/TAM, is required for the transition between meiosis I and meiosis II. No phenotype has been reported for mutants lacking any of the other CYCA, probably due to the high level of redundancy [24]. By contrast, none of the other cyclins were able to compensate for CYCA1;2 in the meiosis I to meiosis II transition. CYCA1;2 may have a specialist function or pattern of expression, required for this transition, that cannot be supplied by any of the other cyclins from Arabidopsis. Alternatively, the lack of CYCA1;2 may decrease generic cyclin/CDK complex activity, causing the meiosis I-meiosis II transition to fail.
CYCA1;2 and OSD1 Are Involved in the Prophase to Meiosis I Transition at Male Meiosis
Both cyca1;2/tam and the previously described osd1 mutants fail to enter meiosis II and produce spores after meiosis I. Remarkably, male meiocytes lacking both OSD1 and CYCA1;2/TAM genes fail to enter meiotic division I, producing spores directly after prophase. This shows that in addition to their crucial function in driving meiosis I to meiosis II transition, these two genes are involved in the prophase to meiosis I transition. This suggests that they both contribute to an activity promoting entry into meiosis I and entry into meiosis II, most likely a CDK activity. The molecular function of OSD1 is currently unknown, however, it has been proposed by analogy to Erp1/Emi2 and Mes1 that it may inhibit APC activity, thus promoting CDK activity [23]. TAM/CYCA1;2 being a cyclin, may directly promote CDK activity. We believe that the meiosis I to meiosis II transition is easily disturbed, because fine regulation of the levels of cyclin/CDK activity is required to ensure both exit from meiosis I and entry into meiosis II [8]. Thus a moderate decrease of CDK activity in osd1 and cyca1;2/tam single mutants may cause failure to enter meiosis II without impairing the prophase to meiosis I transition. In contrast the coincident depletion of OSD1 and CYCA1;2/TAM, may further decrease CDK activity, impairing entry into meiosis I. Unfortunately the direct measurement of CDK activity during meiosis in Arabidopsis is currently not possible. The combination of osd1 and/or cyca1;2/tam mutants with other mutants affecting CDK activity may help to test this model.
In S. cerevisiae, the simultaneous deletion of two (out of six possible) B-type cyclins (Clb1 and Clb3 or Clb1 and Clb4) leads to a failure to enter meiosis II [20], [21]. However, although Clb3 activity is specific to meiosis II, this is not the case for Clb1 and Clb4 [19], and the Clb1 Clb3 Clb4 triple mutant barely sporulates, suggesting that all three proteins have functions in meiosis progression other than the meiosis I-meiosis II transition, similar to OSD1 and CYCA1;2/TAM.
Different Control of Male and Female Meiosis
In the osd1/tam double mutant, male meiocytes fail to enter meiosis I after prophase, whereas female meiocytes proceed to meiosis I and fail to enter meiosis II revealing a striking difference in the control of male and female meiosis progression. We suggest that other cyclins may partly compensate for the absence of CYCA1;2, during female meiosis, and that the meiosis I to meiosis II transition may be driven by different mixtures of cyclins in male and female meiosis. This possibility is supported by the cyca1;2/tam single mutants being weakly affected in the female lineage, compared to the male lineage and to osd1 male and female lineage. An analysis of the effects on meiosis of the depletion of other cyclins, either alone or together with TAM/CYCA1;2, is required to test this hypothesis.
Cyclin A proteins specific to male meiosis have already been described in mammals. The mouse and human genomes each contain two different A-type cyclins. One of these cyclins, Cyclin A1, is restricted to the germ line whereas Cyclin A2 is ubiquitously expressed [44]–[47]. The loss of Cyclin A1 function has no effect on viability and results in male sterility, with male meiosis progressing to the late prophase and then leading to apoptosis; it has no effect on female fertility [48]. Control of male and female meiosis by a different set of cyclins may thus be a general phenomenon.
Plant Cyclins and Meiosis
The Arabidopsis genome encodes 50 cyclins or putative cyclins [25]. Two of these cyclins, SDS and CYCA1;2/TAM, have been directly implicated in meiosis. CYCA1;2/TAM is a type A cyclin involved in cell cycle progression [27] [25 and this study], whereas SDS forms an outgroup, and plays a specific role in recombination with no evidence for any role in cell cycle progression [49], [50]. SDS is required to bias crossovers between homologous chromosome at meiosis rather than between sister chromatid, as in mitosis [50]. A viable hypomorphic mutant of CDKA displays meiotic defects potentially corresponding to a combination of the sds and tam defects [51] (e.g. recombination and progression defects), consistent with both SDS and TAM/CYCA1;2 being involved in CDKA activation. In wheat, Cdc2/CDKA genes are essential components of the Ph1 locus, which is responsible for preventing recombination between homeologous chromosomes [52]. This suggests that cyclin/CDK activity may finely regulate various meiotic events.
Relevance for Apomixis
Apomixis, or asexual clonal reproduction through seeds, has great potential for agricultural applications [40], [41]. It can be separated into three developmental components: an absence or alteration of meiosis (apomeiosis), the fertilization-independent development of the embryo from the egg cell (parthenogenesis), and the initiation of endosperm development with or without fertilization [40], [41], [53]. We recently showed that fully penetrant apomeiosis can be induced in Arabidopsis, when meiosis is replaced by a mitosis-like division in the MiMe genotype, which combines mutations in three genes, AtSPO11-1, to eliminate recombination, AtREC8, to ensure the separation of sister chromatids rather than homologues and OSD1 to abolish the second division [23]. We have now identified a second gene, CYCA1;2/TAM, the mutation of which abolishes the second division of meiosis. The MiMe-2 genotype, which combines the Atspo11-1, AtRec8 and a newly described tam mutation, gives rise to the same phenotype as the MiMe genotype, with the conversion of meiosis into a mitosis-like division. Thus, apomeiosis can be engineered by combining various mutations. However, as tam mutations have a lower penetrance than osd1 mutations, MiMe-2 plants produce apomeiotic gametes less frequently than MiMe plants. Thus, osd1 mutations may be more suitable than tam mutations for agricultural applications, if the results obtained in Arabidopsis can be extrapolated to crop plants. In addition to apomeiosis in MiMe or MiMe-2 plants, apomixis will required the introduction of parthenogenesis and autonomous endosperm formation.
Materials and Methods
Growth Conditions and Genotyping
Arabidopsis plants were cultivated as previously described [54]. For cytometry experiments, Arabidopsis plants were cultivated on Arabidopsis medium [55], at 21°C, under a 16-h to 18-h photoperiod and 70% relative humidity. The tam-2 (Sail_505-C06 Columbia accession), tam-3 (Salk_080686 Columbia accession), tam-4 (CSHL_Et12273 Landsberg erecta accession), tam-5, tam-6 and tam-7 mutants were genotyped by PCR. For tam-2, tam-3 and tam-4 two primer pairs were used. The first pair is specific to the wild-type allele and the second pair is specific to the left border of the inserted sequence. tam-2: N874380U (5′ GACTTGATGGATCCACAGC 3′) & N874380L (5′ CAGAAATCCTCCACTTGCG 3′); N874380L & LB3Sail (5′ TAGCATCTGAATTTCATAACCAATCTCGATACAC 3′). tam-3: N580686U (5′ GAAGAGTATAGGCTTTCGCCC 3′) & N580686L (5′ TGCAACCACATCAGATGAGAG 3′); N580686L & LBSalk2 (5′ GCTTTCTTCCCTTCCTTTCTC 3′). tam-4: ET12273R (5′ TAATGGGACCCACTGATGATC 3′) & ET12273L (5′ ACCTCAGATACACGCAAATGC 3′); ET12273L & Ds5-1 (5′ GAAACGGTCGGGAAACTAGCTCTAC 3′). tam-5 F2P24.10_3 (5′ CATCGCTTTGGAGCAATTCGGTGT) & F2P24.10_555 (ACCAAAACCTGCTTTATCTCGCAATT. tam-6 F2P24.10_cf (CACCATGTCTTCTTCGTCGAGAAATCTATCTCA) & F2P24.10_572 (TGGCCGATCTTATTTGAACAATTCACCTC). tam-7 F2P24.10_4 (AAGACACCGAATTGCTCCAAAGCG) & F2P24.10_yjd (ATGTAATCTAGAGCCGGTCTTTTGTTCAA). The PCR products for tam-5, tam-6 and tam-7 were designed as derived amplified polymorphic sequences (dCAPS) [56] using the Sainsbury atPRIMER webtool (http://www.atprimer.tsl.ac.uk/cgi-bin/form1.cgi). The dCAPS for tam-5, tam-6 and tam-7 are cleaved with Mfe I (125 bp versus 103 bp+22 bp), Xba I (316 bp versus 136 bp+180 bp) and Mfe I (396 bp versus 365 bp+31 bp) respectively. The cleaved amplified polymorphic sequences (CAPS) used to genotype qrt1-2 were described by Francis et al [39]. The primers used to genotype osd1-1, Atspo11-1-3 and Atrec8-3 were described in a previous study [23]. The tam-2 and tam-4 T-DNA right border junction was analyzed by sequencing PCR products. For tam-2, the specific primers used were 77400F (5′TTGGCGAATCGTGGCGAGAA 3′) and Rb3Sail (5′TAACAATTTCACACAGGAAACAGCTATGAC3′. For tam-4, the specific primers used were ET12273R and Ds3-4 (5′ CCGTCCCGCAAGTTAAATATG3′).
Fluorescent Tagged Lines
The seeds for FTL analysis were obtained from G.P. Copenhaver [36], [57]. We obtained tam-2/qrt1-2 and FTL3253 +/ − plants by selfing double heterozygous tam-2/qrt1-2, FTL3253 +/ − plants. We obtained osd1-1/qrt1-2 and FTL3253 +/ − plants by selfing a double heterozygote osd1-1/qrt1-2 FTL3253 +/ − plants. We obtained tam-2/spo11-1-3/rec8-3/qrt1-2 and FTL993 +/ − FTL1273 +/ − plants by selfing a qrt1-2 mutant, triple heterozygous tam-2/spo11-1-3/rec8-3 and FTL993 +/ − FTL1273 +/ − plant. We obtained osd1-1/spo11-1-3/rec8-3/qrt and FTL993 +/ − FTL1273 +/ − plants by crossing a qrt1-2 mutant, triple heterozygote osd1-1/spo11-1-3 /rec8-3 FTL993 −/ − FTL1273 +/+ plant with a qrt1-2 mutant, triple heterozygous osd1-1/spo11-1-3/rec8-3 FTL993 +/+ FTL1273 −/ − plant.
Plants of interest were selected by PCR genotyping. For each line, the first pair of primers is specific to the wild-type allele and the second pair is specific to the T-DNA insertion:
FTL3253 (AmCyan, nucleotide position 9304032 on chromosome 5): FTL3253U (5′ AACTTAGATGCCGAAGAAATG 3′) & FTL3253L (5′ GAGATTCTATACAGATTGATCC 3′);
FTL3253U & LB-TDNA_FTL (5′ GGCATGCAAGCTGATAATTC3′)
FTL993 (CFP, nucleotide position 25731311 on chromosome 5): FTL993U (5′ AGTGACAAGAATCCTAGTCG 3′) & FTL993L (5′GTCTCTACTAAGAGCTCCTC 3′); FTL993L & LB-TDNA_FTL
FTL1273 (DsRed2, nucleotide position 18164269 on chromosome 5): FTL1273U (5′ TACTTAGTCTAGGGTATACAC3′)& FTL1273L (5′ TATAATCGTTCGTCAACGAG 3′); FTL1273L & LB-TDNA_FTL. The I3 line (qrt1-2 with two insertions on chromosome 3, FTL1500 CFP at nucleotide position 498916 and FTL1371 DsRed2 at nucleotide position 4319513) as described in Francis et al., [39] was used for EMS mutagenesis as described below.
Genetic Analysis
Genetic complementation is typically tested by crossing homozygous mutants, but with the tam mutants this would introduce a complicating variable since the progeny would be tetraploid. Instead we assessed complementation by crossing heterozygous tam plants, selecting double heterozygous progeny with PCR and scoring their phenotype.
A tam-2 mutant with Col/No-0 polymorphisms was obtained by crossing a heterozygous tam-2 mutant with a No-0 plant and selfing the F1 generation. We then selected tam mutants heterozygous for several Col/No-0 polymorphisms in the F2 generation and crossed them to wild-type Ler plants. The triploid plants obtained were genotyped to infer the genotype of the tam 2n gametes.
We obtained tam-2/spo11-1-3/rec8-3 mutants with Col-0/Ler polymorphisms by crossing a triple heterozygous tam-2/spo11-1-3/rec8-3 mutant (Col-0) with a wild-type Ler and selfing the F1 progeny. Similarly, plants of interest in the F2 generation were crossed with wild-type No-0 and the triploid progeny were genotyped. The trimorphic (Col-0/Ler/No-0) microsatellite markers used to genotype the tam-2 (Col-0/No-0) x Ler population and the tam-2/spo11-1-3/rec8-3 (Col-0/Ler) triple mutant x No-0 F1 population have been described previously [23]. Microsatellite The NGA76 microsatellite was amplified (Tm: 57°C) with the 5′GGAGAAAATGTCACTCTCCACC 3′ and 5′AGGCATGGGAGACATTTACG 3′ primers (35 cycles of 30 sec at 94°C, 30 sec at Tm and 45 sec at 72°C).
Cytology and Flow Cytometry
Final meiotic products were observed, chromosome spreads generated, and genome sizes were determined as described previously [23]. Pollen fluorescence was analysed as previously described [36]. Images were acquired with a LEICA DM RXA2 epifluorescence microscope using eCFP and DSRed2 filters (Chroma Technologies) and processed with Photoshop 8 (Adobe Systems Inc.).
Mutagenesis
EMS alleles of the TAM locus were generated following the protocol of Weigel and Glazebrook [58]. 0.5 grams of seed were imbibed in 30 ml of sterile water for 4 hrs. and then mutagenized with 0.2% ethyl methane sulfonate for 16 hrs. at room temperature with gentle agitation. Mutagenized seeds were rinsed with 30 ml of sterile water eight times and then dried before planting. Seeds were harvested from individually collected M1 plants. M2 plants were screened for a pollen dyad phenotype (background was qrt1-2 which produces pollen tetrads).
Supporting Information
Zdroje
1. GertonJL
HawleyRS
2005 Homologous chromosome interactions in meiosis: diversity amidst conservation. Nat Rev Genet 6 477 487
2. ZicklerD
KlecknerN
1999 Meiotic chromosomes: integrating structure and function. Annu Rev Genet 33 603 754
3. HashimotoN
WatanabeN
FurutaY
TamemotoH
SagataN
1994 Parthenogenetic activation of oocytes in c-mos-deficient mice. Nature 370 68 71
4. BretagnolleF
ThompsonJ
1995 Gametes with the somatic chromosome number: mechanisms of their formation and role in the evolution of autopolyploid plants. New Phytologist 129 1 22
5. OttoSP
2007 The evolutionary consequences of polyploidy. Cell 131 452 462
6. OttoSP
WhittonJ
2000 Polyploid incidence and evolution. Annu Rev Genet 34 401 437
7. RamannaMS
JacobsonE
2003 Relevance of sexual polyploidization for crop improvement. Euphytica 133 3 18
8. MarstonAL
AmonA
2004 Meiosis: cell-cycle controls shuffle and deal. Nat Rev Mol Cell Biol 5 983 997
9. PesinJA
Orr-WeaverTL
2008 Regulation of APC/C Activators in Mitosis and Meiosis. Annu Rev Cell Dev Biol 24 475 499
10. JonesKT
2004 Turning it on and off: M-phase promoting factor during meiotic maturation and fertilization. Mol Hum Reprod 10 1 5
11. LedanE
PolanskiZ
TerretME
MaroB
2001 Meiotic maturation of the mouse oocyte requires an equilibrium between cyclin B synthesis and degradation. Dev Biol 232 400 413
12. KobayashiH
MinshullJ
FordC
GolsteynR
PoonR
1991 On the synthesis and destruction of A - and B-type cyclins during oogenesis and meiotic maturation in Xenopus laevis. J Cell Biol 114 755 765
13. IwabuchiM
OhsumiK
YamamotoTM
SawadaW
KishimotoT
2000 Residual Cdc2 activity remaining at meiosis I exit is essential for meiotic M-M transition in Xenopus oocyte extracts. EMBO J 19 4513 4523
14. MadgwickS
HansenDV
LevasseurM
JacksonPK
JonesKT
2006 Mouse Emi2 is required to enter meiosis II by reestablishing cyclin B1 during interkinesis. J Cell Biol 174 791 801
15. OheM
InoueD
KanemoriY
SagataN
2007 Erp1/Emi2 is essential for the meiosis I to meiosis II transition in Xenopus oocytes. Dev Biol 303 157 164
16. TangW
WuJQ
GuoY
HansenDV
PerryJA
2008 Cdc2 and Mos regulate Emi2 stability to promote the meiosis I-meiosis II transition. Mol Biol Cell 19 3536 3543
17. IzawaD
GotoM
YamashitaA
YamanoH
YamamotoM
2005 Fission yeast Mes1p ensures the onset of meiosis II by blocking degradation of cyclin Cdc13p. Nature 434 529 533
18. KimataY
TrickeyM
IzawaD
GannonJ
YamamotoM
2008 A mutual inhibition between APC/C and its substrate Mes1 required for meiotic progression in fission yeast. Dev Cell 14 446 454
19. CarlileTM
AmonA
2008 Meiosis I is established through division-specific translational control of a cyclin. Cell 133 280 291
20. DahmannC
FutcherB
1995 Specialization of B-type cyclins for mitosis or meiosis in S. cerevisiae. Genetics 140 957 963
21. KiburzBM
AmonA
MarstonAL
2008 Shugoshin promotes sister kinetochore biorientation in Saccharomyces cerevisiae. Mol Biol Cell 19 1199 1209
22. BarrellPJ
GrossniklausU
2005 Confocal microscopy of whole ovules for analysis of reproductive development: the elongate1 mutant affects meiosis II. Plant J 43 309 320
23. d'ErfurthI
JolivetS
FrogerN
CatriceO
NovatchkovaM
2009 Turning meiosis into mitosis. PLoS Biol 7 e1000124 doi:10.1371/journal.pbio.1000124
24. InzeD
De VeylderL
2006 Cell cycle regulation in plant development. Annu Rev Genet 40 77 105
25. WangG
KongH
SunY
ZhangX
ZhangW
2004 Genome-wide analysis of the cyclin family in Arabidopsis and comparative phylogenetic analysis of plant cyclin-like proteins. Plant Physiol 135 1084 1099
26. MagnardJL
YangM
ChenYC
LearyM
McCormickS
2001 The Arabidopsis ene Tardy Asynchronous Meiosis is required for the normal pace and synchrony of cell division during male meiosis. Plant Physiol 127 1157 1166
27. WangY
MagnardJL
McCormickS
YangM
2004 Progression through meiosis I and meiosis II in Arabidopsis anthers is regulated by an A-type cyclin predominately expressed in prophase I. Plant Physiol 136 4127 4135
28. SchollRL
MayST
WareDH
2000 Seed and molecular resources for Arabidopsis. Plant Physiol 124 1477 1480
29. SessionsA
BurkeE
PrestingG
AuxG
McElverJ
2002 A high-throughput Arabidopsis reverse genetics system. Plant Cell 14 2985 2994
30. AlonsoJM
StepanovaAN
LeisseTJ
KimCJ
ChenH
2003 Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653 657
31. SundaresanV
SpringerP
VolpeT
HawardS
JonesJD
1995 Patterns of gene action in plant development revealed by enhancer trap and gene trap transposable elements. Genes Dev 9 1797 1810
32. BrownJW
1996 Arabidopsis intron mutations and pre-mRNA splicing. Plant J 10 771 780
33. DilkesBP
SpielmanM
WeizbauerR
WatsonB
Burkart-WacoD
2008 The maternally expressed WRKY transcription factor TTG2 controls lethality in interploidy crosses of Arabidopsis. PLoS Biol 6 e308 doi:10.1371/journal.pbio.0060308
34. Sanchez MoranE
ArmstrongSJ
SantosJL
FranklinFC
JonesGH
2001 Chiasma formation in Arabidopsis thaliana accession Wassileskija and in two meiotic mutants. Chromosome Res 9 121 128
35. MercierR
JolivetS
VezonD
HuppeE
ChelyshevaL
2005 Two meiotic crossover classes cohabit in Arabidopsis: one is dependent on MER3,whereas the other one is not. Curr Biol 15 692 701
36. BerchowitzLE
CopenhaverGP
2008 Fluorescent Arabidopsis tetrads: a visual assay for quickly developing large crossover and crossover interference data sets. Nat Protoc 3 41 50
37. d'ErfurthI
JolivetS
FrogerN
CatriceO
NovatchkovaM
2008 Mutations in AtPS1 (Arabidopsis thaliana Parallel Spindle 1) lead to the production of diploid pollen grains. PLoS Genet 4 e1000274 doi:10.1371/journal.pgen.1000274
38. PreussD
RheeSY
DavisRW
1994 Tetrad analysis possible in Arabidopsis with mutation of the QUARTET (QRT) genes. Science 264 1458 1460
39. FrancisKE
LamSY
HarrisonBD
BeyAL
BerchowitzLE
2007 Pollen tetrad-based visual assay for meiotic recombination in Arabidopsis. Proc Natl Acad Sci U S A 104 3913 3918
40. BicknellRA
KoltunowAM
2004 Understanding apomixis: recent advances and remaining conundrums. Plant Cell 16 Suppl S228 245
41. SpillaneC
CurtisMD
GrossniklausU
2004 Apomixis technology development-virgin births in farmers' fields? Nat Biotechnol 22 687 691
42. ChelyshevaL
DialloS
VezonD
GendrotG
VrielynckN
2005 AtREC8 and AtSCC3 are essential to the monopolar orientation of the kinetochores during meiosis. J Cell Sci 118 4621 4632
43. AlexanderMP
1969 Differential staining of aborted and nonaborted pollen. Stain Technol 44 117 122
44. RavnikSE
WolgemuthDJ
1996 The developmentally restricted pattern of expression in the male germ line of a murine cyclin A, cyclin A2, suggests roles in both mitotic and meiotic cell cycles. Dev Biol 173 69 78
45. SweeneyC
MurphyM
KubelkaM
RavnikSE
HawkinsCF
1996 A distinct cyclin A is expressed in germ cells in the mouse. Development 122 53 64
46. YangR
MorosettiR
KoefflerHP
1997 Characterization of a second human cyclin A that is highly expressed in testis and in several leukemic cell lines. Cancer Res 57 913 920
47. RavnikSE
WolgemuthDJ
1999 Regulation of meiosis during mammalian spermatogenesis: the A-type cyclins and their associated cyclin-dependent kinases are differentially expressed in the germ-cell lineage. Dev Biol 207 408 418
48. LiuD
MatzukMM
SungWK
GuoQ
WangP
1998 Cyclin A1 is required for meiosis in the male mouse. Nat Genet 20 377 380
49. AzumiY
LiuD
ZhaoD
LiW
WangG
2002 Homolog interaction during meiotic prophase I in Arabidopsis requires the SOLO DANCERS gene encoding a novel cyclin-like protein. EMBO J 21 3081 3095
50. De MuytA
PereiraL
VezonD
ChelyshevaL
GendrotG
2009 A high throughput genetic screen identifies new early meiotic recombination functions in Arabidopsis thaliana. PLoS Genet 5 e1000654 doi:10.1371/journal.pgen.1000654
51. DissmeyerN
NowackMK
PuschS
StalsH
InzeD
2007 T-loop phosphorylation of Arabidopsis CDKA;1 is required for its function and can be partially substituted by an aspartate residue. Plant Cell 19 972 985
52. GriffithsS
SharpR
FooteTN
BertinI
WanousM
2006 Molecular characterization of Ph1 as a major chromosome pairing locus in polyploid wheat. Nature 439 749 752
53. KoltunowAM
GrossniklausU
2003 Apomixis: a developmental perspective. Annu Rev Plant Biol 54 547 574
54. VignardJ
SiwiecT
ChelyshevaL
VrielynckN
GonordF
2007 The interplay of RecA-related proteins and the MND1-HOP2 complex during meiosis in Arabidopsis thaliana. PLoS Genet 3 e176 doi:10.1371/journal.pgen.0030176
55. EstelleMA
SomervilleC
1987 Auxin-resistant mutants of Arabidopsis thaliana with an altered morphology. Mol Gen Genet 206 200 206
56. NeffMM
NeffJD
ChoryJ
PepperAE
1998 dCAPS, a simple technique for the genetic analysis of single nucleotide polymorphisms: experimental applications in Arabidopsis thaliana genetics. Plant J 14 387 392
57. BerchowitzLE
FrancisKE
BeyAL
CopenhaverGP
2007 The Role of AtMUS81 in Interference-Insensitive Crossovers in A. thaliana. PLoS Genet 3 e132 doi:10.1371/journal.pgen.0030132
58. WeigelD
GlazebrookJ
2002 Arabidopsis: A Laboratory Manual
59. Wang Y, Jha AK, Chen R, Doonan JH, Yang M Polyploidy-associated genomic instability in Arabidopsis thaliana. Genesis 48 254 263
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2010 Číslo 6
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Web-Based, Participant-Driven Studies Yield Novel Genetic Associations for Common Traits
- The IG-DMR and the -DMR at Human Chromosome 14q32.2: Hierarchical Interaction and Distinct Functional Properties as Imprinting Control Centers
- Cushing's Syndrome and Fetal Features Resurgence in Adrenal Cortex–Specific Knockout Mice
- Amplification of a Cytochrome P450 Gene Is Associated with Resistance to Neonicotinoid Insecticides in the Aphid
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy