
Ribosomal DNA Deletions Modulate Genome-Wide Gene Expression: “–Sensitive” Genes and Natural Variation
The ribosomal rDNA gene array is an epigenetically-regulated repeated gene locus. While rDNA copy number varies widely between and within species, the functional consequences of subtle copy number polymorphisms have been largely unknown. Deletions in the Drosophila Y-linked rDNA modifies heterochromatin-induced position effect variegation (PEV), but it has been unknown if the euchromatic component of the genome is affected by rDNA copy number. Polymorphisms of naturally occurring Y chromosomes affect both euchromatin and heterochromatin, although the elements responsible for these effects are unknown. Here we show that copy number of the Y-linked rDNA array is a source of genome-wide variation in gene expression. Induced deletions in the rDNA affect the expression of hundreds to thousands of euchromatic genes throughout the genome of males and females. Although the affected genes are not physically clustered, we observed functional enrichments for genes whose protein products are located in the mitochondria and are involved in electron transport. The affected genes significantly overlap with genes affected by natural polymorphisms on Y chromosomes, suggesting that polymorphic rDNA copy number is an important determinant of gene expression diversity in natural populations. Altogether, our results indicate that subtle changes to rDNA copy number between individuals may contribute to biologically relevant phenotypic variation.
Published in the journal:
. PLoS Genet 7(4): e32767. doi:10.1371/journal.pgen.1001376
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1001376
Summary
The ribosomal rDNA gene array is an epigenetically-regulated repeated gene locus. While rDNA copy number varies widely between and within species, the functional consequences of subtle copy number polymorphisms have been largely unknown. Deletions in the Drosophila Y-linked rDNA modifies heterochromatin-induced position effect variegation (PEV), but it has been unknown if the euchromatic component of the genome is affected by rDNA copy number. Polymorphisms of naturally occurring Y chromosomes affect both euchromatin and heterochromatin, although the elements responsible for these effects are unknown. Here we show that copy number of the Y-linked rDNA array is a source of genome-wide variation in gene expression. Induced deletions in the rDNA affect the expression of hundreds to thousands of euchromatic genes throughout the genome of males and females. Although the affected genes are not physically clustered, we observed functional enrichments for genes whose protein products are located in the mitochondria and are involved in electron transport. The affected genes significantly overlap with genes affected by natural polymorphisms on Y chromosomes, suggesting that polymorphic rDNA copy number is an important determinant of gene expression diversity in natural populations. Altogether, our results indicate that subtle changes to rDNA copy number between individuals may contribute to biologically relevant phenotypic variation.
Introduction
The ribosomal rDNA gene array is an epigenetically-regulated repeated locus responsible for producing over 90% of cellular RNAs. The large (35S) rDNA array gives origin to the Nucleolus Organizer Region (NOR) and the 35S rRNA genes are present from fewer than 50 to more than 25,000 copies among eukaryotes [1]. However, even though copy number varies widely between and within species [2], [3], the functional consequences of copy number polymorphisms have been largely unknown, and often assumed to have negligible functional consequence. This is in part because redundant rDNA arrays are found in eukaryotic genomes. In humans, for example, rDNA arrays containing the 35S rRNA genes are found on chromosomes 13, 14, 15, 21, and 22. Similarly, redundant rDNA genes within each array can compensate for copy number polymorphisms to maintain a sufficient supply of rRNAs [4]–[6]; indeed, only a fraction of the rDNA units in a particular array are transcribed at any given time [7], and flexibility in the transcriptional elongation rate allows for control of rRNA output in conditions of rapid cell division and growth. Hence, even though rDNA copy number and rRNA production is regulated by the cell, the relevance of supernumerary or inactive rDNA repeat units has remained elusive. Recent work has investigated whether inactive rDNA copies are necessary for genome stability [8].
In Drosophila melanogaster, rDNA arrays exist as a pair of functionally-redundant loci on the X and Y chromosomes [5], [6], which have further suggested that variable copy number in the Y-linked rDNA array might have little, if any, functional significance. Indeed, even though X0 males are sterile due to loss of Y-linked fertility genes, they appear morphologically normal despite having no Y-linked rDNA [9]. Nevertheless, Paredes and Maggert [10] have recently shown that induced variation in the Y-linked rDNA copy number modifies heterochromatin-induced position effect variegation (PEV), and natural changes in rDNA copy number through development correlate with PEV. Taken together, these findings suggested that polymorphisms in rDNA copy number might be relevant to the maintenance of genome-wide chromatin structure.
Polymorphic naturally occurring Y chromosomes induce Y-linked Regulatory Variation (YRV), which affects the expression of autosomal and X-linked genes [11]. In Drosophila melanogaster, YRV is observed in males differing only in the origin of their Y chromosomes and it is manifested as the differential expression of hundreds of non-Y-linked genes [11]. The source of YRV cannot be simply ascribed to polymorphisms in protein-coding genes [12], [13], nor is it easily mapped to sub-regions of the Y chromosome because of the lack of recombination along the Y-chromosome and the difficulty in manipulating large segments of heterochromatin. The similar contributions of induced deletions in the Y-linked rDNA array and naturally occurring Y chromosome variation to PEV [10]–[13], suggested that naturally occurring rDNA copy number polymorphism might be a significant source of regulatory variation. The hypothesized association between Y-linked regulatory variation and rDNA copy number might provide a molecular framework for the contribution of the Drosophila melanogaster Y chromosome to adaptive phenotypic variation and fitness.
Here we tested the hypothesis that induced deletions in the Y-linked rDNA in an otherwise isogenic background might modulate variation in genome-wide gene expression. Our results indentified hundreds to thousands of genes whose expression is sensitive to deletions in the rDNA. Importantly, many of the induced changes in expression could be replicated in both male and female genotypes. Further, we show significant associations between genome-wide gene expression variation arising from Y chromosomes with targeted deletions in the rDNA loci and variation arising from naturally occurring polymorphic Y chromosomes, which harbor polymorphisms in rDNA copy number. This suggests that variation in rDNA copy number may be an underappreciated source of natural variation with important consequences to the modulation of gene expression across the genome.
Results
To address the relevance of rDNA copy number to genome-wide gene expression we used Y chromosomes containing targeted reductions of rDNA copy number. These chromosomes are otherwise expected to remain structurally unchanged. Although it is possible that these chromosome differ in ways that we cannot detect, four lines of evidence suggest that alterations of the rDNA copy number are the sole difference between these chromosomes: (1) I-CreI-mediated chromosome alterations were genetically and cytologically limited to the rDNA [14], (2) no other identifiable I-CreI consensus site has been cloned from Drosophila, (3) the position effect variegation phenotype arising form these alterations were reverted concomitant with rDNA magnification [10], and (4) Drosophila rDNA is known to be comprised solely of rDNA and two retroelements [15], [16]. Although we cannot formally rule out the possibility that heretofore uncharacterized non-rDNA sequences are in the rDNA loci, for ease of discussion we consider that deletions in the rDNA are the major source of variation between these chromosomes.
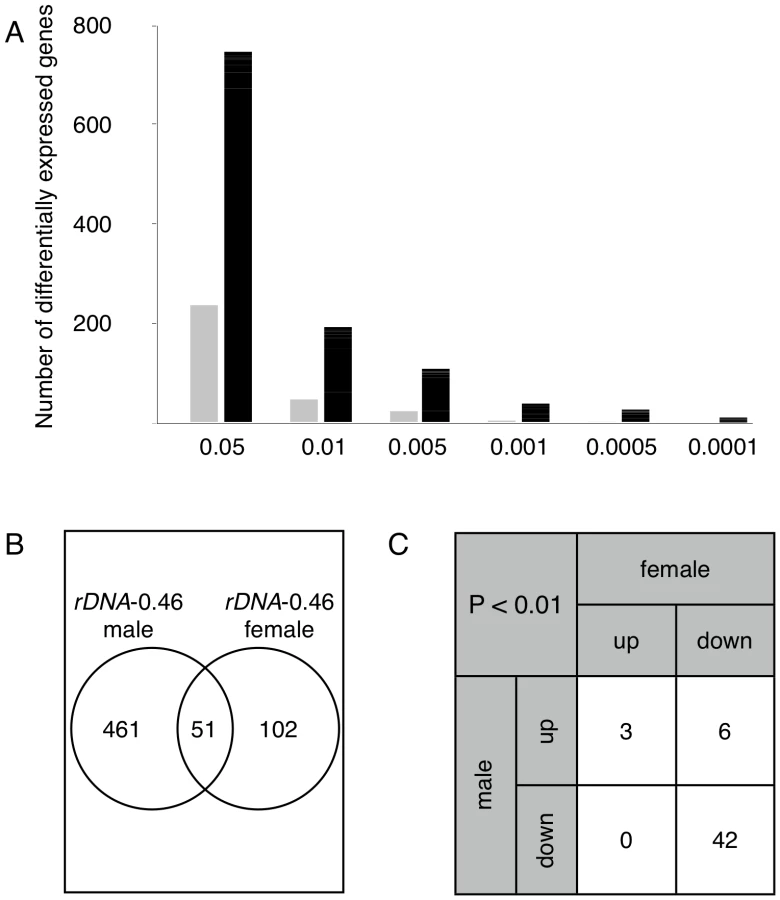
Four Y chromosomes that differ in the copy number of rDNA repeats were introgressed into an isogenic background of autosomes and X chromosome (Figure S1); genome-wide expression was interrogated with microarrays (Figure S2). We compared adult male flies bearing a wild-type Y chromosome to three strains carrying mutant derivative Y chromosomes with reduced rDNA repeats – two mildly (YrDNA-mild, with rDNA copy numbers approximately 87% and 85% of wild-type) and one grossly (YrDNA-gross, 46% of wild type) reduced array. We observed substantial gene expression variation among these strains as compared to the random expectation across a range of P-values (Figure 1A). We expected that laboratory induced deletions in the rDNA array would result in gene expression modulation that would not only be replicated in each independently generated mutant, but would also be more pronounced in mutants lacking a larger proportion of the original locus. Our data are in agreement with both these expectations, and further suggest that rDNA copy number may be the major source of variation between these chromosomes. Accordingly, the numbers of differentially expressed genes were positively correlated with the rDNA deletion size. We observed that the Y chromosome with the lowest rDNA copy number (YrDNA-gross) induced the highest number of expression changes, whereas the two Y chromosomes with mildly-deleted arrays resulted in smaller numbers of differentially expressed genes. This finding is consistent with evidence that chromosomes with fewer rDNA copies have a stronger effect on position effect variegation [10]. Furthermore, the genes identified as differentially expressed in the mild deletions were a subset of those induced in the strain with grossly deleted rDNA (Figure 1B). Accordingly, 42–71% (P<0.001, FDR <0.05) of the genes identified with chromosomes containing mild rDNA deletions were also identified by the Y chromosome with the smallest rDNA copy number. In support of the reproducibility of the gene expression modulation that was observed in PEV across the lines [10], 24% of differentially expressed genes were shared by at least two chromosomes with reduced rDNA arrays, whereas fewer than 0.2% were expected to be shared by chance in randomly permuted datasets (P<0.001). In addition, the direction and magnitudes of changes in expression were significantly correlated (ρ = 0.78–0.84, P<10E−16) between Y chromosomes harboring rDNA deletions (Figure 1C), suggesting a similar responsiveness of the affected genes regardless the size of the rDNA deletion. These data support our contention that a common lesion – namely rDNA copy number variation – underlies this phenotype, and points to the relevance of rDNA copy number variation to modulation of genome-wide gene expression.

Natural polymorphisms and induced deletions of the rDNA show no overt dominant phenotype [3], [17]. Even X0 males, which other than being sterile due to loss of Y-linked fertility genes, appear morphologically normal despite having no Y-linked rDNA [9]. Therefore, we expected that induced rDNA deletion would have impacts on individual gene expression that were generally small. Indeed, we found that 85% of genes whose expression differed significantly from wild-type had changes in expression level of no more than 50% (Figure S3A). It was unexpected and unlikely that the number of genes showing only a 10% change in relative expression would be less than those showing a 20% change. Instead this result is likely an underestimation of genes exhibiting small changes caused by limitation of statistical power to detect them. We projected a linear regression of the number of genes whose differential expression was altered by 10–20%, 20–30%, 30–40%, and 40–50% in an attempt to correct the first decile (Figure S3B–S3D). From that extrapolation, we estimate that as many as 1200–1700 differentially expressed genes at P<0.05 and 360–435 at P<0.01 might escape statistical detection despite our high level of replication (Figure S2). Since we analyzed 8073 unique gene IDs from our microarrays, these estimates suggest that as much as 40% of the genome might be subtly affected by partial rDNA deletion.
To determine the generality of rDNA induced expression changes, we investigated the differential expression between the rDNA-deleted YrDNA-gross and its ancestral wild-type Y chromosome in females with XXY karyotype. We observed hundreds of gene expression differences between XXYrDNA-gross and isogenic XXY females bearing the wild-type Y chromosome. The number of differentially expressed genes was fewer in XXY females than in males (Figure 2A), possibly due to the smaller fractional rDNA size difference in the female karyotype which contains two wild-type X-linked arrays. Nevertheless, we observed a significant enrichment of differentially expressed genes commonly shared between males and females harboring the same Y chromosome; at P<0.01, 51 genes were shared between the sexes whereas only a single gene was expected to be shared by chance alone. As before, expected numbers were calculated from randomly permuted datasets (Figure 2B, cf. Figure S4A). While the number of affected genes differed between the sexes, we observed a significant association between down-regulated genes (Figure 2C, Figure S5), and a significant correlation in fold-changes between sexes (ρ = 0.45, P<10E–16). A similar trend of up - and down-regulation and commonly shared genes remained across a range of P-values used for ascertaining differential expression (Figure S4B, S4C). Together, our data show that rDNA copy number variation commonly affects male and female transcription and identifies a similar set of “rDNA-sensitive” genes. These data suggest that the response of a gene to rDNA deletion is an attribute of the gene structure and/or its regulation, rather than a sex-dependent effect.
One mechanism for the wide-ranging effects of rDNA copy number on gene expression might arise if deletions of the rDNA compromised the spread of heterochromatin structure to euchromatin from centric heterochromatic regions [18], [19]. Indeed, this simple model is suggested by the observation that three heterochromatin-induced variegating alleles were affected by rDNA deletion [10]. However, studies have established that the spreading of heterochromatin-associated factors is neither long-ranged, processive, or simple [20], [21]. Nonetheless, to address the issue we tested two strong predictions of a simple heterochromatin spreading model: that (i) genes residing in the proximity of heterochromatin will be more strongly affected by changes in rDNA copy number, and that (ii) the majority of gene expression changes will be seen as increase in expression as repressive heterochromatin is reduced. We therefore tested for an enrichment of differentially expressed genes according to their cytological location by plotting the number of differentially expressed genes from all three chromosomes (Figure 3A black bars, Figure 3B black line). We compared the number of differentially expressed genes to the number of analyzed genes from the microarray (Figure 3B gray bars and line), which corrected for uneven gene density across the genome and for any bias in the representation of the genome on our microarrays.

We found that while the number of differentially expressed genes appears to drop near the cytological bands juxtaposed to centric heterochromatic blocks (bands 20, 40/41, and 80/81), this is indistinguishable from our expectation based on lower representation of genes from these regions. Moreover, we could discern no general trend for genes near the telomeres (bands 1, 21, and 60/61). Furthermore, while X-linked loci were less affected than were chromosome 2 - and 3-linked genes (43–47% the frequency), these trends were not statistically different from that expected given the distribution of represented genes (Figure S6A). Next, we analyzed the genomic region at the transition between the X-linked 20 megabases of centric heterochromatin and adjacent euchromatin. None of the first 20 genes were differentially expressed. The same held true for the telomere of the long arm of the X chromosome. Finally, we assessed the density of genes responding to rDNA copy number along all the chromosome arms in a sliding 2 megabase window at 1 megabase increments. Significance was determined empirically by analyzing 1000 sets of randomly sampled genes taken from the entire genome. Each set of random data had a sample size equal to the number of differentially expressed genes. No significant overrepresentation of differentially expressed genes was detected in any increment. Thus, our analysis indicated that the genes affected by rDNA deletion are randomly distributed in the genome, and are neither influenced by proximity to heterochromatin nor chromosome linkage. There was also no preponderant increase or decrease of differential expression. Corroborating our conclusion that rDNA deletion affects the same set of genes in males and females, we saw that the distribution of affected genes in males and females was coincident (Figure 3C, Figure S6B). Taken together these results argue against a simple heterochromatin spreading model but instead suggest that loci with sensitivity to rDNA copy number variation are scattered through the genome.
An intriguing possibility is that polymorphisms of the rDNA copy number in naturally occurring Y chromosomes could account in part for the differential genome-wide modulation of gene expression exerted by these chromosomes. One indication that such rDNA-driven Y-linked variation is relevant came from categorizing the differential expressed genes by Gene Ontogeny (GO) category. These analyses pointed to five categories that overlapped with those discovered by comparing differential gene expression due to natural Y chromosome polymorphisms (Table 1) [11]. Importantly, energy metabolism and rDNA activity are closely associated [22], [23]. Our data suggest that rDNA copy number itself might play a role regulating energy metabolism through modulation of gene expression of genes responsible for energy management or membrane metabolism. Accordingly, we observed that in both males and females “rDNA sensitive” genes are highly enriched for proteins localized to the mitochondrial membrane (46 genes in males, P<2.1E–10; 38 genes in females P<7.2E–12) and involved in electron transport (44 genes in males, P<2.3E–05; 38 genes in females, P<1.4E–09). These enrichments are significant in both males and females, but collectively account for 16% and 9% respectively of the differentially expressed genes (at P<0.05). Although energy and lipid metabolism may be influenced by rDNA copy number, they are not the sole targets.

To address the relevance of rDNA copy number in natural populations, we integrated data from gene expression variation due to natural Y-linked polymorphisms and gene expression variation due to deletions in the rDNA array. We found an unexpected level of overlap in the identity of differentially expressed genes when compared to the level expected by chance alone (Figure 4A, Figure S7). We found 124 genes with evidence for differential expression (P<0.005) in comparisons between chromosomes with induced changes in rDNA copy number and in comparisons between naturally-occurring chromosomes. We assessed statistical significance of this overlap in two ways. First, the same procedure for identifying differential expression was applied to datasets in which the expression data were randomly permuted. These analyses showed that only five genes were identified as differentially expressed in both permuted datasets. Second, we generated random samples of size 568 (the observed number of differentially expressed genes shared between the induced rDNA chromosomes) and 683 (the observed number of differentially expressed genes shared between the natural chromosomes), and counted the number of genes that appeared in the overlap of both sets in each of 10,000 replicated of this experiment. The analysis indicates that the observed overlap of 124 genes is significantly enriched compared to random datasets (P<0.0001, Figure 4B). This enrichment shows that a gene affected by YRV is predictive of being affected by rDNA copy number.

Finally, we analyzed the set of genes whose expression is affected by rDNA deletion, and found that absolute fold changes due to these deletions is significantly correlated with expression variation arising due to naturally-occurring Y chromosome polymorphisms (ρ = 0.25–0.55, P<1.0E–12, Figure 5). These correlation coefficients indicate that approximately 5–30% of gene expression variation detected on natural Y chromosomes might be due to polymorphisms in the rDNA loci. Using real-time PCR we confirmed that the natural Y chromosomes possessed polymorphisms in rDNA copy number, the range of which included the two mild rDNA deletions used in this study (Figure 6). Moreover, the effect of naturally occurring Y chromosomes from YOhio and YZimbabwe on heterochromatin-induced position effect variegation were consistent with effects seen for induced deletions: lower rDNA copy number correlated with increased suppression of variegation [10], [12].


Discussion
Our work identified the consequences of induced rDNA deletions to the expression of hundreds to thousands of genes scattered through the genome of both males and females. The data are reproducible between independently-generated chromosomes derived from a common ancestor, with lines containing smaller deletions showing smaller effects on gene expression. Although we cannot exclude the possibility that some undetected Y-linked differences other than rDNA copy number might confound some of the effects we have seen, we interpret the changes in gene expression to be, at least in part, a consequence of induced deletions in the rDNA locus. Previous studies lend credence to this interpretation [10], [14], [17]. Furthermore, the results herein reported are well in line with the expectation that the measured rDNA deletions partly underlie the expression phenotypes we report. Accordingly, we observed that the Y chromosome with the lowest rDNA copy number (YrDNA-gross) induced the highest number of expression changes, whereas the two Y chromosomes with mildly-deleted arrays resulted in smaller numbers of differentially expressed genes. This finding is consistent with evidence that chromosomes with fewer rDNA copies have a stronger effect on position effect variegation [10]. Furthermore, the genes identified as differentially expressed in the mild deletions were a subset of those induced in the strain with grossly deleted rDNA. Furthermore, we showed that such rDNA-responsive genes are not physically clustered or near heterochromatin; instead, affected genes are distributed across the genome and show functional enrichments for genes encoding proteins localized to the mitochondria and involved in electron transfer. This finding is reminiscent of previous observations for a close link between rDNA activity and energy metabolism. Furthermore, our analyses suggested that naturally occurring copy number polymorphism in the rDNA array might be an important and underappreciated source of regulatory variation in natural populations.
Epigenetic regulation of chromatin structure is necessary for the stability of the rDNA array and the nucleolus. Indeed, destabilization of rDNA copy number and nucleolar structure are markers of aging and cancer progression [24], [25] and our findings raise the possibility that rDNA alterations might underlie some of these pleiotropic phenotypes through its consequences on genome-wide gene expression. Mechanistically, the transcriptional activity of the rDNA locus is regulated through histone modifications and allows for nucleolus formation from even a single rDNA repeat unit [8], [26]–[28]. Furthermore, many chromatin components known to regulate rDNA activity and stability are dosage sensitive. For instance, the Sir2-family proteins are required for the silencing of genes inserted into the rDNA repeated array, rDNA stability, and telomeric silencing [29], [30]. In Drosophila, Sir2 has an essential role in both euchromatic gene repression and heterochromatic silencing [29]. Hence, it is conceivable that rDNA expression and rDNA copy number are related, and perturbations to either might yield heritable alteration in genome structure with consequences on gene expression of unlinked genes. Accordingly, we propose that short and long rDNA arrays may result in the differential recruitment of nuclear proteins for rRNA expression/processing and nucleolar structure. In such a model, variation of protein dosages due to either the release of proteins required for silencing of non-expressed rDNA repeats (which might be more abundant in the larger array) or due to the recruitment of proteins required for the higher expression of a smaller number of expressed rDNA repeats (as might be the case in the shorter rDNA arrays) might be expected. Indeed, Michel and colleagues showed that spontaneous rDNA deletions in yeast caused decreased cellular abundance of Sir2 mRNA and protein [31]. Hence, the widespread effects of rDNA copy number on genome-wide expression might arise from imbalances in the nuclear protein pool, which includes chromatin and transcription factors, that might result from the requirements for silencing and expression of variable numbers of rDNA repeats or structural elements within nucleoli. Alternatively, Paredes and Maggert showed alterations in nucleolus stability as a consequence of reduced rDNA copy number [10]. The dynamic composition of the nucleolus [32] may differ in nucleoli coalesced around long or short rDNA arrays.
Paredes and Maggert showed that rDNA copy number altered the balance of euchromatin and heterochromatin in Drosophila models of position effect variegation [10]. Those observations suggested a global effect of rDNA copy number on chromatin structure. Significantly, multiple studies show a complex regulation of heterochromatin spreading at natural euchromatin-heterochromatin transition zones and chromosome rearrangement breakpoint junctions. Yasuhara and Wakimoto showed that increased levels of HP1 association at variegating euchromatic-heterochromatic junctions is not homogenous across the chromosome [20], and both Talbert and Henikoff, and Ahmad and Henikoff showed that heterochromatin spreading does not translate into uniform responses for closely-linked genes [33], [34]. Most recently, Vogel and colleagues showed that while a variegating white+ gene was responsive to changes in chromatin environment, the majority of linked genes remained unresponsive to the spreading of HP1 from nearby heterochromatin [21]. Hence, it was not surprising that genes responding to rDNA copy number were distributed across the genome. More important, this observation does not contradict the view that rDNA copy number impacts global chromatin structure. To the contrary, recent studies suggested that heterochromtain-induced PEV [12], [13] and euchromatic gene expression might be related [11], and rDNA deletions affect heterochromatin-induced PEV [10]: the data we present here supports the unity of these observations by showing that deletions within the rDNA affect euchromatic gene expression, uncover a significantly overlapping subset of genes, and account for a fraction of naturally occurring YRV.
Although rDNA copy number polymorphism may only account for a small fraction of YRV, our data indicate a relevant contribution with significant enrichments in the number of differentially expressed genes that are shared by both rDNA deletions lines and naturally occurring Y chromosomes. This is unexpected because rDNA copy number is typically thought to be without genetic consequence. The remainder of YRV may map to other heterochromatin of the Y chromosome, either simple repeats or transposable elements. Similarly, the remainder of rDNA-linked variation may be ameliorated or epistatically masked by compensatory polymorphisms elsewhere in the heterochromatin. Finally, our results raise the issue of how widespread these effects may be in other chromosomes and organisms containing highly variable rDNA loci. Indeed, the large and varied number of rDNA copies is a common characteristic of eukaryotic genomes [2]. The human diploid karyotype has ten 35S and two 5S arrays, each highly variable with regard to size and activity [35]. In plants such as Arabidopsis, the 5S rDNA arrays contain heritable regulatory chromatin structure [36], and in flax, heritable induced phenotypic variation is mapped to rDNA copy number [37], [38]. Although rDNA copy number in Drosophila may vary over an order of magnitude, the upper limits remain consistent across many studies [1]. What limits the expansion of rDNA is yet to be established, but our results suggest that superfluous rDNA repeats – inactive or active – have genetic consequence to gene expression, which might impose a heretofore underappreciated pressure against excessive expansion or contraction. We envision this genetic consequence to be heritable as a result of copy number polymorphism, suggesting that rDNA copy number is an important consideration in understanding the evolution of a genome. Indeed, the rDNA array has a uniquely dynamic biology of copy number loss and gain that occurs meiotically and somatically through development [5], [39]–[41]. Hence, the ability of the rDNA to change in copy number and activity might provide a novel mechanism for adaptation to environmental changes by maintaining a euchromatin/heterochromatin balance that is most conducive for fitness.
Materials and Methods
Drosophila stocks and crosses
Drosophila melanogaster Y-chromosomes containing targeted deletions in the rDNA locus were generated as previously described [17]. These chromosomes were derived from a common single male ancestor in the laboratory that was subjected to targeted deletions within the rDNA locus, and thus created an allelic series of Y chromosomes virtually identical with the exception of variation in rDNA copy number. These Y chromosomes were introgressed into an isogenic (X chromosome, autosomes, and mitochondrial genome) laboratory stock [11] by crossing XY males to females homozygous for the markers yellow, brown, ebony, cubitus interruptus, and eyeless (y/y; bw/bw; e/e; ci ey/ci ey) for two generations (Figure S1). This isogenic stock is expected to contain very little genetic variation, and upon receipt was subjected to no fewer than eight additional generations of brother-sister mating to reinforce homozygosity of the genetic background. Four Y chromosomes were analyzed: The original Y chromosome that contains a wild type rDNA array (100%), two derived chromosomes with mild deletions 87% (YrDNA-0.87) and 85% (YrDNA-0.85) of wild-type, and one grossly reduced derived chromosome that contains 46% (YrDNA-0.46) of wild-type. Flies were grown under 24 h light at constant temperature (25°C) and humidity (80%). XXY female flies were obtained by crossing males from the isogenic Y chromosome substitution lines described above to females from a laboratory stock containing a compound (attached) X chromosome, C(1)M4, y.
rDNA copy number analyses
rDNA copy number in Y-chromosome substitution strains containing the rDNA deletions were checked with a random subset of flies used for gene expression analysis. rDNA copy number was measured as described in Paredes and Maggert [17]. For each stock whose Y-linked rDNA array was measured males were crossed with C(1)DX, y1 f1 bb0, which lacks all X-linked rDNA. The resulting C(1)DX/Y females contain only Y-linked rDNA, whose copy number was determined with quantitative Real-time PCR using conserved rDNA primers that amplified 18S subunit rRNA gene sequence. For Real-time PCR analyses, five biological replicates consisting of three experimental replicates from individual flies were sampled for each genotype. Real-time PCR analyses were carried out with the Power Sybr Green Master Mix (Applied Biosystems, Foster City, California), and amplification profiles were obtained with an ABI Step-One Real-time PCR (Applied Biosystems, Foster City, California). Reactions were quality-confirmed for the absence of dimers or nonspecific amplification by analyzing melting curve kinetics and electrophoretic analysis of products.
Gene expression analyses
Microarrays were approximately 18,000-feature cDNA arrays spotted with Drosophila melanogaster cDNA PCR products. For RNA extraction, newly emerged male flies were collected and aged for three days at 25°C, after which they were flash frozen in liquid nitrogen and stored at −80°C. When females were analyzed, they were collected within 7 hours of eclosion to assure they were unmated prior to aging under the same conditions as were males. Total RNA was extracted from whole flies using TRIZOL (Gibco-BRL, Life Technologies, Gaithersburg, Maryland). cDNA synthesis, labeling with fluorescent dyes (Cy3 and Cy5) and hybridization reactions were carried out using 3DNA protocols and reagents (Genisphere Inc., Hatfield, Pennsylvania). Slides were scanned using AXON 4000B scanner (Axon Instruments, Foster City, California) and the GenePix Pro 6.0 software. Stringent quality-control criteria were used to ensure reliability of foreground intensity reads for both Cy3 and Cy5 channels. Foreground fluorescence of dye intensities was normalized by the Loess method in the library Limma [42], [43] of the software R. Significance of variation in gene expression due to Y chromosome origin was assessed with linear models and empirical Bayes moderated F statistics in Limma [42], [43]. P values were adjusted for multiple testing by using the method of Benjamini and Hochberg to control the false discovery rate [44]. Test results were considered to be significant if the adjusted P values were less than 0.05, nominally controlling the expected false discovery rate to no more than 5%. Differential expression was also assessed using the Bayesian Analysis of Gene Expression Levels (BAGEL) model [45]. Results were robust to the choice of linear models in Limma or BAGEL. False discovery rates were estimated based on the variation observed when randomized versions of the original dataset were analyzed. Similarly, expected values for the overlap between independent datasets were estimated by applying the same statistical procedure (Limma or BAGEL) to permuted versions of the datasets. Significance of the overlap was further assessed by generating samples of sizes that matched the number of differentially expressed genes in separate datasets (induced rDNA deletions and natural Y chromosomes) and determining the number of genes found in both random samples; the mean number was 38.54, and the maximum was 62 genes in 10,000 trials. Enrichment in gene ontology categories was assessed using a modified Bonferroni correction with GeneMerge [46]. Microarray gene expression data can be obtained at the GEO database (GSE27695).
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. LongEO
DawidIB
1980 Repeated genes in eukaryotes. Annu Rev Biochem 49 727 764
2. ProkopowichCD
GregoryTR
CreaseTJ
2003 The correlation between rDNA copy number and genome size in eukaryotes. Genome 46 48 50
3. LyckegaardEM
ClarkAG
1989 Ribosomal DNA and Stellate gene copy number variation on the Y chromosome of Drosophila melanogaster. Proc Natl Acad Sci U S A 86 1944 1948
4. KarpenGH
SchaeferJE
LairdCD
1988 A Drosophila rRNA gene located in euchromatin is active in transcription and nucleolus formation. Genes Dev 2 1745 1763
5. TartofKD
1973 Regulation of ribosomal RNA gene multiplicity in Drosophila melanogaster. Genetics 73 57 71
6. TerracolR
Prud'hommeN
1981 26S and 18S rRNA synthesis in bobbed mutants of Drosophila melanogaster. Biochimie 63 451 455
7. FrenchSL
OsheimYN
CiociF
NomuraM
BeyerAL
2003 In exponentially growing Saccharomyces cerevisiae cells, rRNA synthesis is determined by the summed RNA polymerase I loading rate rather than by the number of active genes. Mol Cell Biol 23 1558 1568
8. IdeS
MiyazakiT
MakiH
KobayashiT
2010 Abundance of ribosomal RNA gene copies maintains genome integrity. Science 327 693 696
9. BridgesCB
1916 Non-Disjunction as Proof of the Chromosome Theory of Heredity (Concluded). Genetics 1 107 163
10. ParedesS
MaggertKA
2009 Ribosomal DNA contributes to global chromatin regulation. Proc Natl Acad Sci U S A 106 17829 17834
11. LemosB
AraripeLO
HartlDL
2008 Polymorphic Y chromosomes harbor cryptic variation with manifold functional consequences. Science 319 91 93
12. LemosB
BrancoAT
HartlLD
2010 Epigenetic modulation of gene expression by polymorphic Y chromosomes: Sexual conflict, chromatin components, and immune response. Proc Natl Acad Sci U S A in press
13. JiangPP
HartlDL
LemosB
2010 Y Not a Dead End: Epistatic Interactions Between Y-linked Regulatory Polymorphisms and Genetic Background Affect Global Gene Expression in Drosophila melanogaster. Genetics
14. MaggertKA
GolicKG
2005 Highly efficient sex chromosome interchanges produced by I-CreI expression in Drosophila. Genetics 171 1103 1114
15. PellegriniM
ManningJ
DavidsonN
1977 Sequence arrangement of the rDNA of Drosophila melanogaster. Cell 10 213 214
16. WellauerPK
DawidIB
1977 The structural organization of ribosomal DNA in Drosophila melanogaster. Cell 10 193 212
17. ParedesS
MaggertKA
2009 Expression of I-CreI endonuclease generates deletions within the rDNA of Drosophila. Genetics 181 1661 1671
18. RudolphT
YonezawaM
LeinS
HeidrichK
KubicekS
2007 Heterochromatin formation in Drosophila is initiated through active removal of H3K4 methylation by the LSD1 homolog SU(VAR)3-3. Mol Cell 26 103 115
19. TalbertPB
HenikoffS
2006 Spreading of silent chromatin: inaction at a distance. Nat Rev Genet 7 793 803
20. YasuharaJC
WakimotoBT
2008 Molecular landscape of modified histones in Drosophila heterochromatic genes and euchromatin-heterochromatin transition zones. PLoS Genet 4 e16 doi:10.1371/journal.pgen.0040016
21. VogelMJ
PagieL
TalhoutW
NieuwlandM
KerkhovenRM
2009 High-resolution mapping of heterochromatin redistribution in a Drosophila position-effect variegation model. Epigenetics Chromatin 2 1
22. GuarenteL
2000 Sir2 links chromatin silencing, metabolism, and aging. Genes Dev 14 1021 1026
23. MurayamaA
OhmoriK
FujimuraA
MinamiH
Yasuzawa-TanakaK
2008 Epigenetic control of rDNA loci in response to intracellular energy status. Cell 133 627 639
24. JohnsonFB
SinclairDA
GuarenteL
1999 Molecular biology of aging. Cell 96 291 302
25. DryginD
RiceWG
GrummtI
2010 The RNA polymerase I transcription machinery: an emerging target for the treatment of cancer. Annu Rev Pharmacol Toxicol 50 131 156
26. PengJC
KarpenGH
2007 H3K9 methylation and RNA interference regulate nucleolar organization and repeated DNA stability. Nat Cell Biol 9 25 35
27. PengJC
KarpenGH
2009 Heterochromatic genome stability requires regulators of histone H3 K9 methylation. PLoS Genet 5 e1000435 doi:10.1371/journal.pgen.1000435
28. KobayashiT
2008 A new role of the rDNA and nucleolus in the nucleus--rDNA instability maintains genome integrity. Bioessays 30 267 272
29. RosenbergMI
ParkhurstSM
2002 Drosophila Sir2 is required for heterochromatic silencing and by euchromatic Hairy/E(Spl) bHLH repressors in segmentation and sex determination. Cell 109 447 458
30. FuruyamaT
BanerjeeR
BreenTR
HartePJ
2004 SIR2 is required for polycomb silencing and is associated with an E(Z) histone methyltransferase complex. Curr Biol 14 1812 1821
31. MichelAH
KornmannB
DubranaK
ShoreD
2005 Spontaneous rDNA copy number variation modulates Sir2 levels and epigenetic gene silencing. Genes Dev 19 1199 1210
32. BoisvertFM
van KoningsbruggenS
NavascuesJ
LamondAI
2007 The multifunctional nucleolus. Nat Rev Mol Cell Biol 8 574 585
33. TalbertPB
HenikoffS
2000 A reexamination of spreading of position-effect variegation in the white-roughest region of Drosophila melanogaster. Genetics 154 259 272
34. AhmadK
HenikoffS
2001 Modulation of a transcription factor counteracts heterochromatic gene silencing in Drosophila. Cell 104 839 847
35. StultsDM
KillenMW
PierceHH
PierceAJ
2008 Genomic architecture and inheritance of human ribosomal RNA gene clusters. Genome Res 18 13 18
36. RiddleNC
RichardsEJ
2002 The control of natural variation in cytosine methylation in Arabidopsis. Genetics 162 355 363
37. CullisCA
2005 Mechanisms and control of rapid genomic changes in flax. Ann Bot 95 201 206
38. SchneebergerRG
CullisCA
1991 Specific DNA alterations associated with the environmental induction of heritable changes in flax. Genetics 128 619 630
39. HawleyRS
TartofKD
1985 A two-stage model for the control of rDNA magnification. Genetics 109 691 700
40. TartofKD
1974 Unequal mitotic sister chromatin exchange as the mechanism of ribosomal RNA gene magnification. Proc Natl Acad Sci U S A 71 1272 1276
41. CohenS
YacobiK
SegalD
2003 Extrachromosomal circular DNA of tandemly repeated genomic sequences in Drosophila. Genome Res 13 1133 1145
42. SmythGK
2004 Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat Appl Genet Mol Biol 3 Article3
43. GentlemanR
2005 Bioinformatics and Computational Biology Solutions Using R and Bioconductor. New York Springer
44. BenjaminiY
HochbergY
1995 Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society 57 289 300
45. TownsendJP
HartlDL
2002 Bayesian analysis of gene expression levels: statistical quantification of relative mRNA level across multiple strains or treatments. Genome Biol 3 RESEARCH0071
46. Castillo-DavisCI
HartlDL
2003 GeneMerge--post-genomic analysis, data mining, and hypothesis testing. Bioinformatics 19 891 892
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2011 Číslo 4
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Survival Motor Neuron Protein Regulates Stem Cell Division, Proliferation, and Differentiation in
- PTG Depletion Removes Lafora Bodies and Rescues the Fatal Epilepsy of Lafora Disease
- Evolution of Vertebrate Transient Receptor Potential Vanilloid 3 Channels: Opposite Temperature Sensitivity between Mammals and Western Clawed Frogs
- An Evolutionary Genomic Approach to Identify Genes Involved in Human Birth Timing
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy