
Phosphorylation-Dependent Differential Regulation of Plant Growth,
Cell Death, and Innate Immunity by the Regulatory Receptor-Like Kinase
BAK1
Plants rely heavily on receptor-like kinases (RLKs) for perception and
integration of external and internal stimuli. The Arabidopsis regulatory
leucine-rich repeat RLK (LRR-RLK) BAK1 is involved in steroid hormone responses,
innate immunity, and cell death control. Here, we describe the differential
regulation of three different BAK1-dependent signaling pathways by a novel
allele of BAK1, bak1-5. Innate immune signaling mediated by the
BAK1-dependent RKs FLS2 and EFR is severely compromised in
bak1-5 mutant plants. However, bak1-5
mutants are not impaired in BR signaling or cell death control. We also show
that, in contrast to the RD kinase BRI1, the non-RD kinases FLS2 and EFR have
very low kinase activity, and we show that neither was able to
trans-phosphorylate BAK1 in vitro. Furthermore, kinase activity
for all partners is completely dispensable for the ligand-induced
heteromerization of FLS2 or EFR with BAK1 in planta, revealing
another pathway specific mechanistic difference. The specific suppression of
FLS2 - and EFR-dependent signaling in bak1-5 is not due to a
differential interaction of BAK1-5 with the respective ligand-binding RK but
requires BAK1-5 kinase activity. Overall our results demonstrate a
phosphorylation-dependent differential control of plant growth, innate immunity,
and cell death by the regulatory RLK BAK1, which may reveal key differences in
the molecular mechanisms underlying the regulation of ligand-binding RD and
non-RD RKs.
Published in the journal:
. PLoS Genet 7(4): e32767. doi:10.1371/journal.pgen.1002046
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002046
Summary
Plants rely heavily on receptor-like kinases (RLKs) for perception and
integration of external and internal stimuli. The Arabidopsis regulatory
leucine-rich repeat RLK (LRR-RLK) BAK1 is involved in steroid hormone responses,
innate immunity, and cell death control. Here, we describe the differential
regulation of three different BAK1-dependent signaling pathways by a novel
allele of BAK1, bak1-5. Innate immune signaling mediated by the
BAK1-dependent RKs FLS2 and EFR is severely compromised in
bak1-5 mutant plants. However, bak1-5
mutants are not impaired in BR signaling or cell death control. We also show
that, in contrast to the RD kinase BRI1, the non-RD kinases FLS2 and EFR have
very low kinase activity, and we show that neither was able to
trans-phosphorylate BAK1 in vitro. Furthermore, kinase activity
for all partners is completely dispensable for the ligand-induced
heteromerization of FLS2 or EFR with BAK1 in planta, revealing
another pathway specific mechanistic difference. The specific suppression of
FLS2 - and EFR-dependent signaling in bak1-5 is not due to a
differential interaction of BAK1-5 with the respective ligand-binding RK but
requires BAK1-5 kinase activity. Overall our results demonstrate a
phosphorylation-dependent differential control of plant growth, innate immunity,
and cell death by the regulatory RLK BAK1, which may reveal key differences in
the molecular mechanisms underlying the regulation of ligand-binding RD and
non-RD RKs.
Introduction
Plants are under constant pressure to respond rapidly and accurately to changing environmental and developmental conditions. Hence they need to translate extracellular signals into appropriate intracellular responses. Cell surface receptor-like-kinases (RLKs) are one of the major components in this extracellular sensing. The model plant species Arabidopsis and rice show a huge expansion of the RLK family compared to other eukaryotes with >600 and >1100 members, respectively [1]. However, only a very limited number of plant RLKs have an assigned function ranging from development to responses to biotic and abiotic stresses [2]–[4].
Plant RLKs share a common domain organization with the well-studied mammalian receptor tyrosine kinases (RTKs) [5], [6]. The activation of RTKs is initiated by ligand binding to the extra-cellular domain leading to conformational changes that are transmitted by a single trans-membrane domain and induce receptor homo - and/or hetero-oligomerization [7]. This leads to activation by trans - and auto-phoshorylation of the activation loop, correct positioning of the cytoplasmic asymmetric kinase dimer, and release of the inhibition by the C-terminal and/or juxta-membrane regions [8]–[10]. Downstream signaling is initiated by sequential auto - or trans-phosphorylation of specific residues in the cytoplasmic domain serving as docking sites for downstream signaling partners, and/or direct phosphorylation of signaling substrates [11].
Kinases, including RLKs, can be subdivided into RD and non-RD kinases depending on the conservation of the amino-acid residue preceding the core catalytic aspartate (Asp) residue in subdomain VIb of the kinase domain [12], [13]. Most RD kinases require auto-phosphorylation of the activation loop for full kinase activity. In contrast, non-RD kinases do not require activation loop phosphorylation and are activated by different mechanisms [13].
Notably, several plant RD - and non-RD ligand-binding receptor kinases (RKs) share the common RD-type regulatory RLK BAK1 as signaling partner [14], [15]. The leucine-rich repeat (LRR)-RLK BAK1 (At4g33430) is a member of the somatic embryogenesis-related kinase (SERK) family and is also named SERK3 [16], [17]. BAK1 was initially identified as a positive regulator of brassinosteroid (BR) responses, forming a ligand-dependent complex in vivo with the LRR-RK BRI1 (At4g39400), the main BR receptor [18]–[21]. Over-expression of BAK1 suppresses weak bri1 alleles, and bak1 knock-out mutants are hypo-sensitive to BR and resemble weak bri1 alleles [18], [19], [21].
BAK1 is also involved in cell death control as bak1 knock-out mutants have a spreading lesion phenotype upon pathogen infection and premature senescence [22], [23]. This loss of cell death control is aggravated in double-mutant combinations with its closest paralog BKK1/SERK4 (At2g13790), and strong bak1 bkk1 allele combinations are seedling lethal even in sterile conditions [22], [24]. Additionally, BAK1 interacts with BIR1 (At5g48380), another LRR-RLK, mutants of which also show constitutive uncontrolled cell death [25].
BAK1 was also identified as an important regulator of pathogen-associated-molecular-pattern (PAMP)-triggered immunity (PTI) [26], [27]. Bak1 null mutants are compromised in their responsiveness to several PAMPs including flg22 (derived from bacterial flagellin), elf18 (derived from bacterial EF-Tu), HrpZ, lipopolysaccharides, peptidoglycans, and damage-associated molecular patterns (DAMPs), such as AtPep1 [26]–[29]. Furthermore, BAK1-silenced Nicotiana benthamiana (N. benthamiana) plants are less sensitive to the PAMPs INF1 and csp22 (derived from bacterial cold shock protein) [26]. BAK1 rapidly forms ligand-dependent heteromers with the flg22 and elf18 pattern-recognition receptors (PRRs), the ligand-binding LRR-RKs FLS2 (At5g46330) and EFR (At5g20480), respectively [26], [27], [30](Roux et al., submitted). BAK1 also interacts in a ligand-independent manner with the AtPep1 PRRs, the ligand-binding LRR-RKs AtPEPR1/2 (At1g73080/At1g17750) in yeast two-hybrid assays [15]. The importance of the heteromerization with BAK1 in plant innate immunity is substantiated by the targeting of the ligand-induced BAK1-FLS2 interaction by the bacterial virulence effector AvrPto to block PTI signaling [29], [31], [32] Importantly, the function of BAK1 in cell death control and innate immunity seems to be independent of its function in BR signaling [14].
Clearly, BAK1 is an important regulator implicated in multiple independent signaling pathways leading to growth, cell death control and innate immunity. Although BAK1 forms ligand-dependent heteromers with several ligand-binding LRR-RKs [20], [21], [26], [27], it is not required for ligand binding [27], [33]. In that respect, BAK1 should be considered as a regulatory RLK rather than a co-receptor. It is, however, not fully understood how BAK1 regulates these different pathways.
A previous study suggests that BAK1 functions as a signal enhancer for the RD-kinase BRI1 [21]. This conclusion is based on biochemical studies into auto - and trans-phosphorylation events revolving around BRI1-BAK1 followed by phenotypic analysis of BAK1 phospho-mimetic and phospho-dead mutants. Interestingly none of the BAK1 mutant alleles had a strong differential effect on PTI and BR signaling [21]. The activation of BAK1 by BRI1 is further supported by a recent report showing that a tyrosine auto-phosphorylation site in the C-terminus of BAK1 is required for trans-activation of BRI1 [34]. Interestingly, this auto-phoshorylation site of BAK1 is not required for flg22-induced seedling growth inhibition (SGI) [34]. Given this differential requirement of phosphosites and the different mode of regulation of non-RD kinases versus RD kinases [12], [13], it is unclear whether the BRI1-BAK1 model can be generalized to non-RD kinases. Since non-RD kinases are mostly associated with functions in innate immunity across kingdoms [35], it is of great interest to elucidate potential regulatory mechanisms of non-RD kinases and to reveal potential differences to RD kinases.
Here, we demonstrate the phosphorylation-dependent differential regulation of the RD-kinase BRI1 and the non-RD kinases FLS2 and EFR by BAK1. We identified a novel mutant allele of BAK1, bak1-5, that is strongly impaired in PTI signaling but displays a wild-type-like BR signaling capacity. Furthermore, bak1-5 is not impaired in cell death control. This unexpected phenotype is not due to a differential complex formation between BAK1-5 and the RD and non-RD kinases, but requires the kinase activity of BAK1-5 suggesting a phosphorylation-dependent differential regulation. Moreover, our work reveals dramatic differences in the trans-phosphorylation events between BAK1 and BRI1 or EFR in vitro, and the requirement of kinase activity for complex formation in planta.
Results
Identification of the novel BAK1 allele bak1-5
To identify novel regulators of EFR function/signaling in Arabidopsis thaliana, we previously performed a forward-genetic screen for elf18-insensitive (elfin) mutants based on loss SGI triggered by elf18 [36]. Out of 103 non-efr elfin mutants recovered, only one, elfin27-6, showed a clear defect in the SGI induced by both elf18 and flg22, even at high peptide concentrations (1 mM) (Figure 1A, Figure S1). This suggested that this mutant was affected in an important component shared by both EFR - and FLS2-dependent signaling pathways.

Using a map-based cloning approach we identified the corresponding mutation as a single mis-sense substitution in the 10th exon of BAK1 (Figure 1B). We therefore tentatively renamed elfin27-6 as bak1-5. This mutation leads to a C408Y change in the subdomain VIa of the cytoplasmic kinase preceding the catalytic loop (Figure 1C). This Cys residue is conserved in ∼17% of all RLKs in Arabidopsis thaliana (data not shown).
Next, we tested whether the bak1-5 mutation affects the accumulation of the BAK1 protein. To this end, we performed immunoblot analysis on protein extracts of Col-0, bak1-5 and bak1-4 (SALK_116202) mutant plants using anti-BAK1 antibodies. As shown in Figure 1D, full-length mutant BAK1-5 protein accumulated to similar levels as the wild-type protein, whereas the corresponding band was completely missing in bak1-4 null mutants.
To confirm that the C408Y mutation causes the observed elfin phenotype, we first transformed the null mutant bak1-4 with BAK1 or BAK1-5 genomic sequences under the control of their own regulatory sequences. As expected, the wild-type transgene was able to complement the compromised flg22 - and elf18-induced reactive oxygen species (ROS) burst of bak1-4 (Figure 1E and Figure S2A–S2B). Consistently, transgenic plants expressing BAK1-5 were strongly impaired in flg22 - and elf18-induced ROS burst and thus phenocopied the bak1-5 mutant (Figure 1E and Figure S2A–S2B).
To further prove that the bak1-5 mutation causes the elfin phenotype, and to ascertain whether bak1-5 is a recessive or dominant mutation, we took advantage of the fact that bak1-4, in contrast to bak1-5 (Figure 1A and Figure S1), is not impaired in the SGI triggered by elf18 (Figure 1F and Figure S1) [27]. We tested the contribution of BAK1-5 to the impaired elf18-induced SGI in an allelism test between bak1-5 and bak1-4. Only homozygous bak1-5 and bak1-5×bak1-4 heterozygous F1 seedlings showed a strong impairment in elf18-induced SGI (Figure 1F). Interestingly bak1-5×Col-0 heterozygous F1 plants showed an intermediate phenotype between wild-type Col-0 and bak1-5 seedlings (Figure 1F). This indicates that bak1-5 is a semi-dominant allele and suggests that BAK1-5 has as a dose-sensitive dominant-negative effect on the endogenous wild-type BAK1. This semi-dominant-negative effect was not restricted to SGI, but was also observed when elf18-induced ROS burst was measured in leaves of bak1-5×Col-0 heterozygous F1 plants (Figure S2C).
Therefore, bak1-5 is a novel semi-dominant allele of BAK1 with a specific phenotype related to PAMP responsiveness.
bak1-5 is strongly impaired in EFR - and FLS2-dependent PTI signaling
Previous results showed that the null bak1-4 mutant plants were strongly impaired in early and late responses to flg22, but were not impaired in late elf18 responses [27]. In particular, elf18-induced SGI in bak1-4 was indistinguishable from wild-type (Figure 1 E and Figure S1) [27]. Since the novel allele bak1-5 was impaired in both flg22 - and elf18-triggered SGI, we investigated the impact of the bak1-5 mutation on early and late responses triggered by flg22 and elf18.
We found that the ROS burst induced by flg22 and elf18 treatment was strongly reduced in bak1-5 leaves (Figure 2A), whereas leaves of the null mutant bak1-4 showed only a delayed and slightly reduced ROS burst (Figure 2A), as previously reported [26], [27].

Next, we analysed the impact of bak1-5 on the activation of MAP kinases (MPKs) by flg22 and elf18. Consistent with previous observations, the activation of MPK3, 4 and 6 after flg22 and elf18 treatment was delayed and reduced in bak1-4 seedlings (Figure 2B). Surprisingly, the activation of these MPKs by flg22 and elf18 was differentially regulated in bak1-5 seedlings. The activation of MPK3 and 6 by flg22 and elf18 was also delayed, but the level of activation ultimately reached levels similar to that observed in wild-type seedlings at 15 mins. Notably, MPK4 was not activated at all during the time-course of the experiment (Figure 2B).
Since MPK activation is linked to PAMP-induced transcriptional reprogramming [37], [38] we then assessed whether PAMP-induced gene expression was also affected in bak1-5 seedlings using three different PTI marker genes [39] over a 3-hour time-course experiment. The induction of the three genes by flg22 and elf18 was partially impaired in bak1-4 over the time-course although this effect was minor at certain time-points (Figure 2C). In contrast, after flg22 or elf18 treatment the transcript levels of all three PTI-marker genes were drastically reduced in bak1-5 over the time-course (Figure 2C). Interestingly, the steady-state expression of the marker genes was already significantly lower in bak1-5 when compared to wild-type (Figure S3).
Our results clearly demonstrate that bak1-5 plants were strongly affected in all flg22 and elf18 responses measured. Strikingly, the new allele bak1-5 was more strongly impaired in PTI signaling than the null allele bak1-4 suggesting a mis-regulation of PTI signaling. This effect was particularly apparent with EFR-dependent responses, as bak1-4 null mutants were not affected in elf18-triggered late responses, whereas bak1-5 mutants were.
Finally, we tested if the strong impairment of bak1-5 in EFR - and FLS2-dependent PTI signaling compromised resistance to bacterial pathogen. For this purpose we spray-infected four week-old plants with the weakly virulent strain Pto DC3000 COR− that has been previously shown to be compromised in fully suppressing PTI signaling [40]. Consistently, bacteria grew to slightly higher titters in leaves of PRR single mutants fls2 or efr, and to even higher levels in the double mutant fls2 efr when compared to wild-type (Figure 2D). As reported previously [23], bak1-4 mutants were as susceptible as wild-type to bacterial spray-infection (Figure 2D); most likely due to the only slight impairment in PTI signaling and the compromised cell death control [23]. In contrast, bak1-5 plants were hyper-susceptible and supported bacterial multiplication to similar levels as in efr fls2 leaves (Figure 2D). The impairment of bak1-5 in bacterial resistance was further supported by the increased disease symptoms observed after spray-infection with Pto DC3000 COR− (Figure S4). In addition, Col-0, bak1-4 and bak1-4 plants expressing BAK1 displayed no significant disease symptoms after spray-infection with Pto DC3000 COR-, whereas bak1-5 or bak1-4 plants expressing BAK1-5 clearly develop chlorotic lesions associated with disease (Figure S4).
Therefore, the compromised PTI signaling capacity of bak1-5 leads to a reduced ability to launch effective defence responses culminating in hyper-susceptibility to bacteria.
bak1-5 is not impaired in brassinosteroid signaling
Next, we tested if bak1-5 was also impaired in BR signaling, as all previously reported bak1 loss-of-function alleles are hyposensitive to BR [18], [19]. Classically, the reported bak1 loss-of-function alleles display a semi-dwarf cabbage-like rosette when grown under short-day conditions similar to weak bri1 mutant plants [18], [19]. Surprisingly, bak1-5 plants did not show any growth impairment under these conditions and looked comparable to wild-type plants (Figure 3A). Consistently, the expression of both BAK1 and BAK1-5 was able to rescue the semi-dwarf cabbage-like rosette phenotype of bak1-4 (Figure S5).

As plant morphology does not always correlate with defects in other BR responses [17], we compared the effect of exogenous treatments with brassinolide (BL), the most bioactive BR [41], or the BR biosynthesis inhibitor brassinazole (BRZ) [42] on bak1-4 and bak1-5 plants. First, we quantitatively investigated the BR-responsiveness of etiolated seedlings grown under different BR regimes [43]. As expected, bak1-4 hypocotyls were much smaller than wild-type, were hypo-sensitive to the growth inhibition effect of BL, and hyper-sensitive to BRZ (Figure 3B). In contrast, although bak1-5 hypocotyls were slightly smaller than wild-type, they displayed a wild-type-like responsiveness to BRZ and BL (Figure 3B).
To test for subtle changes in BR sensitivity in the bak1-4 and bak1-5 seedlings, we performed BL marker gene analysis by quantitative real-time RT-PCR. For this purpose, we investigated the expression pattern of two well-characterised BL marker genes, SAUR-AC1 (At4g38850) as an auxin co-regulated gene, and EXP8 (At2g40610) as a BL-specific gene [44]. In order to fully capture the signaling capability of either bak1 allele, we included a pre-treatment with BRZ to reduce any hormone level adaptation within genotypes that may have altered BR signaling capacity as previously reported for bzr1-1D [45]. BL treatment on its own did not reveal any significant differences between the genotypes for EXP8 expression (Figure 3C, left). However, the induction of SAUR-AC1 by BL was clearly impaired in bak1-4 and less so in bak1-5 (Figure 3C, right). Interestingly, BRZ pre-treatment prior to BL treatment revealed a clear impairment of bak1-4 in BL-induced gene expression for both marker genes (Figure 3C). On the contrary, bak1-5 showed an induction of SAUR-AC1 comparable to wild-type (Figure 3C, right), and the induction of EXP8 appeared higher in bak1-5 than wild-type under this treatment regime (Figure 3C, left).
Defects in BR sensitivity are often revealed when mutations in potential BR signaling components or biosynthetic genes are combined with weak bri1 alleles [46]. To test if the bak1-5 mutation affects BR sensitivity in such assays, we crossed bak1-4 or bak1-5 with bri1-301 that carries a point mutation in the kinase domain of BRI1 [47]. As previously reported [18], the bak1-4 mutation increased the BR-related phenotypes of bri1-301, as measured by rosette morphology of short-day-grown plants, hypocotyl length of etiolated seedlings grown on BL - or BRZ-containing medium, and morphology of long-day grown plants (Figure 3D–3F). In contrast, the bak1-5 mutation did not aggravate the bri1-301 phenotype to the same extent in any of these assays (Figure 3D–3F). Surprisingly, as noted before with the expression of BL marker genes in bak1-5 (Figure 3C), etiolated bri1-301 bak1-5 seedlings appeared even slightly hyper-responsive to BL when compared to bri1-301 (Figure 3E).
Overall, our results clearly demonstrate that the novel allele bak1-5 is still fully sensitive to BR. This phenotype is in clear contrast with the hypo-sensitivity generally associated with bak1 loss-of-function alleles.
bak1-5 is not impaired in cell death control
To test if bak1-5 is impaired in cell death control, we crossed bak1-4 or bak1-5 with the null mutant bkk1-1 (SALK_057955) [24]. Twenty out of seventy individuals (X2 = 0.476, p = 0.49) from a bak1-4×bkk1-1 F2 segregating population died after two weeks in long-day conditions on sterile MS plates. In contrast, none of bak1-5×bkk1-1 F2 segregating seedlings (n = 76) died, and we could isolate fully viable double mutants (Figure 4). Furthermore, homozygous bak1-5 bkk1-1 plants showed no symptoms related to cell death or early senescence when grown in non-sterile soil, and this even at later stages of development (Figure S6).

The bak1-5 allele is therefore not associated with loss of cell death control.
BAK1-5 shows an enhanced interaction with the ligand-binding LRR-RKs FLS2, EFR, and BRI1
From our detailed phenotypic analysis (Figure 2, Figure 3, Figure 4), it appears that bak1-5 is specifically affected in PTI signaling. One hypothesis for the observed phenotypes could be that BAK1-5 has a reduced interaction with the PRRs FLS2 and EFR, but is still capable of interacting with the BR receptor BRI1.
To test this hypothesis, we performed co-immunoprecipitation analyses between BAK1 and these receptors. Using specific anti-FLS2 antibodies, we could detect a clear flg22-dependent complex formation between FLS2 and BAK1 in wild-type Arabidopsis seedlings (Figure 5A). Surprisingly, BAK1-5 was detected in FLS2 immunoprecipitates from non-elicited seedlings (Figure 5A). In addition, the amount of BAK1-5 in complex with FLS2 after flg22 treatment was greater than in the case of BAK1 (Figure 5A). Similar results were observed when we performed the reciprocal immunoprecipitation experiment (Figure S7).

We recently demonstrated that BAK1 also forms a ligand-dependent complex with EFR (Roux et al., submitted). Due to the lack of specific anti-EFR antibodies that could be used for immunoprecipitation experiments in Arabidopsis, we tested the interaction of epitope-tagged BAK1 or BAK1-5 with EFR after heterologous transient expression in the plant model N. benthamiana. After immunoprecipitation of BAK1-GFP using GFP-trap beads we observed a clear elf18-dependent recruitment of EFR-HA3 into the complex (Figure 5B). Interestingly, the amount of EFR-HA3 present with BAK1-5-GFP in complex after elf18 treatment was higher than with BAK1-GFP (Figure 5B).
Next, we tested the interaction of BAK1-5 with BRI1 after immunoprecipitation with specific anti-BRI1 antibodies (Figure 5C). We were able to confirm the in planta BRI1-BAK1 interaction previously reported using transgenic lines expressing epitope-tagged BRI1 and/or BAK1 proteins [18], [20]. Surprisingly, as observed with FLS2 and EFR, BAK1-5 also showed an enhanced interaction with BRI1 (Figure 5C).
Importantly, BAK1-5 still retained its interaction specificity, as it did not interact with CERK1, a LysM-RK involved in BAK1-independent chitin perception [29], [48], [49], when co-expressed as epitope-tagged proteins in N. benthamiana (Figure S8).
In contrast to our initial hypothesis, BAK1-5 has a higher affinity than BAK1 for the ligand-binding LRR-RKs FLS2, EFR and BRI1. This observation, together with the differential impact of the bak1-5 mutation on PTI signaling triggered by FLS2 and EFR, but not on BRI1-dependent responses (Figure 2 and Figure 3), indicates that the bak1-5 phenotype cannot be solely explained by differences in complex formation.
BAK1-5 is a hypoactive kinase
Since the bak1-5 mutation corresponds to a C408Y amino acid change just before the catalytic loop of the kinase domain (Figure 1C), the bak1-5 phenotype could be due to altered kinase activity.
To test potential differences in BAK1-5 kinase activity, we expressed in Escherichia coli (E. coli) the cytoplasmic domains (CD: residues 256 to 615) of BAK1 and BAK1-5, as well as the respective kinase-dead mutant variants (D416N) (indicated as BAK1* and BAK1-5*, respectively) as N-terminally tagged GST-fusion proteins and purified them using glutathione beads. In agreement with previous studies [18]–[21] we detected a strong phosphorylation of BAK1 CD on threonine/serine and tyrosine residues in vitro (Figure 6A–6B). This is due to the auto-phosphorylation of BAK1 CD during recombinant protein production and in the in vitro kinase assay as the phosphorylation status of kinase dead BAK1* CD was negligible (Figure 6A–6B). The phosphorylation status of BAK1-5 CD was slightly reduced compared to BAK1 CD but still significantly higher than that of kinase dead BAK1-5* CD (Figure 6A–6B). This is also illustrated by the fact that both BAK1 CD and BAK1-5 CD showed a mobility shift on SDS-PAGE compared to kinase inactive mutant variants (Figure 6A–6B). Next, we quantified the reduction of kinase activity of BAK1-5 by determining the auto-phosphorylation levels of BAK1 and BAK1-5 over an increasing concentration range of ATP. As shown in Figure S9, BAK1-5 has an ∼3.6-fold reduction in kinase activity as the C408Y mutation in BAK1-5 lowers its Km to ∼25 µM compared to ∼7 µM in the case of BAK1. These results demonstrate that BAK1-5 is an active kinase albeit with a slightly reduced kinase activity when compared to BAK1.

The RD kinases BRI1 and BAK1 differ from the non-RD kinases EFR and FLS2 in their phosphorylation activities
BAK1 CD and BRI1 CD are active kinases that undergo auto - and trans-phosphorylation when incubated together in vitro [18], [19], [21]. Therefore, we studied the kinase activities of FLS2 CD and EFR CD, and the trans-phosphorylation events occurring between them and the BAK1 CD.
We first analyzed FLS2 and EFR kinase activities and compared them with the kinase activity of BRI1. For this purpose, we expressed in E. coli the CDs of EFR (residues 682 to 1031), FLS2 (residues 840 to 1173) and BRI1 (residue 814 to 1196) as fusion proteins with an N-terminal maltose-binding protein (MBP) tag. As controls, we also constructed the respective kinase-dead variants EFR* CD (D849N), FLS2* CD (D997N) and BRI1* CD (D1009N). We initially intended to identify the phosphorylation status of FLS2 CD, EFR CD and BRI1 CD using phospho-site specific antibodies either recognizing phosphorylated threonine/serine or tyrosine residues. Unfortunately, we were unable to observe a signal specific to the kinase active variants of FLS2 CD and EFR CD (data not shown), therefore we restored to using radioactive [32P]-γ-ATP in in vitro kinase assays. As previously reported [50], BRI1 CD had a very strong auto - and trans-phosphorylation capacity using the artificial substrate myelin basic protein (MBP) (Figure 7A). In contrast, EFR CD possessed only minor auto-phosphorylation capacity and negligible trans-phosphorylation ability on MBP (Figure 7A). Notably, these activities were abolished in BRI1* CD and EFR* CD (Figure 7A), demonstrating that the observed phosphorylations are indeed due to the intrinsic kinase activities of these protein.

Surprisingly, we were unable to detect any FLS2 CD phosphorylation in vitro (Figure 7A), indicating that FLS2 is an extremely weak kinase. The latter result is in disagreement with previous reports that revealed phosphorylation activities in vitro for FLS2 [31], [50], [51]. As Zhou and colleagues [31] used a N-terminally His tagged FLS2 fusion protein to report FLS2 kinase activity, we also generated His-FLS2 CD. Again, as observed with MBP-FLS2 CD, we were unable to observe any phosphorylation activity (Figure S10). Under the same conditions, His-BRI1 CD displayed a strong kinase activity (Figure S10).
Consequently, it appears that in comparison BRI1 is an extremely strong kinase, EFR is a moderately good kinase, and FLS2 is almost kinase-inactive in vitro. Therefore, we focused our trans-phosphorylation studies with BAK1 and BAK1-5 on the comparison between the non-RD kinase EFR and the RD kinase BRI1.
We first confirmed in our experimental conditions that BAK1 CD was able to trans-phosphorylate BRI1* CD, and reciprocally that BRI1 CD was able to trans-phosphorylate BAK1* CD (Figure 7B). Also BAK1-5* CD was trans-phosphorylated by BRI1 CD and to a similar level compared to BAK1* CD (Figure 7B). The reduced kinase activity of BAK1-5 CD lead to a lower level of trans-phosphorylation of BRI1* CD when compared to BAK1 CD (Figure 7B).
Next, we investigated the in vitro trans-phosphorylation events surrounding EFR CD. We found that BAK1 CD was able to trans-phosphorylate EFR* CD to a level much stronger than EFR CD auto-phosphorylation (Figure 7C). This is in contrast to the BAK1-BRI1 trans-phosphorylation events in which BAK1 CD trans-phosphorylation of BRI1* CD is similar in comparison to BRI1 CD auto-phosphorylation (Figure 7B). Another striking difference was the inability of EFR CD to trans-phosphorylate BAK1* CD (Figure 7C). Importantly, BAK1-5 CD was still able to trans-phosphorylate EFR* CD and slightly enhanced the phosphorylation status of EFR CD (Figure 7C).
In summary, BAK1 trans-phosphorylates the non-RD kinase EFR, but not the reverse. In contrast, the RD-kinase BRI1 undergoes a bi-directional trans-phosphorylation with BAK1 in vitro as previously shown [19], [21]. This is particularly interesting as BAK1-5 displays a reduced trans-phosphorylation capacity for both receptors in vitro but specifically blocks signaling events mediated by the non-RD kinase EFR in vivo (Figure 2 and Figure 3).
Kinase activitiy is not required for ligand-dependent FLS2/EFR-BAK1 heteromerization
The kinase activity of BRI1 is strictly required for the ligand-induced BRI1-BAK1 complex formation [21]. To determine whether the in vivo heteromerization of BAK1 with FLS2 or EFR requires the kinase activity of either partner, we co-expressed in N. benthamiana wild-type and kinase-dead versions of FLS2, EFR and BAK1 for co-immunoprecipitation experiments.
Clear ligand-dependent complex formation between the wild-type BAK1 and FLS2 or EFR proteins could be detected (Figure 8A–8B). Co-expressing BAK1*-HA3 with either FLS2-GFP or EFR-GFP did not reduce the complex formation after PAMP treatment when immunoprecipitating FLS2 or EFR using GFP-trap beads (Figure 8). Similarly FLS2*-GFP and EFR*-GFP possessed full interaction capacity after ligand addition when co-expressed with BAK1-HA3 (Figure 8A–8B). Finally, we tested double kinase-dead receptor combinations. After ligand addition, both FLS2*-GFP and EFR*-GFP still interacted with BAK1*-HA3 as strongly as wild-type receptor combinations (Figure 8A–8B).
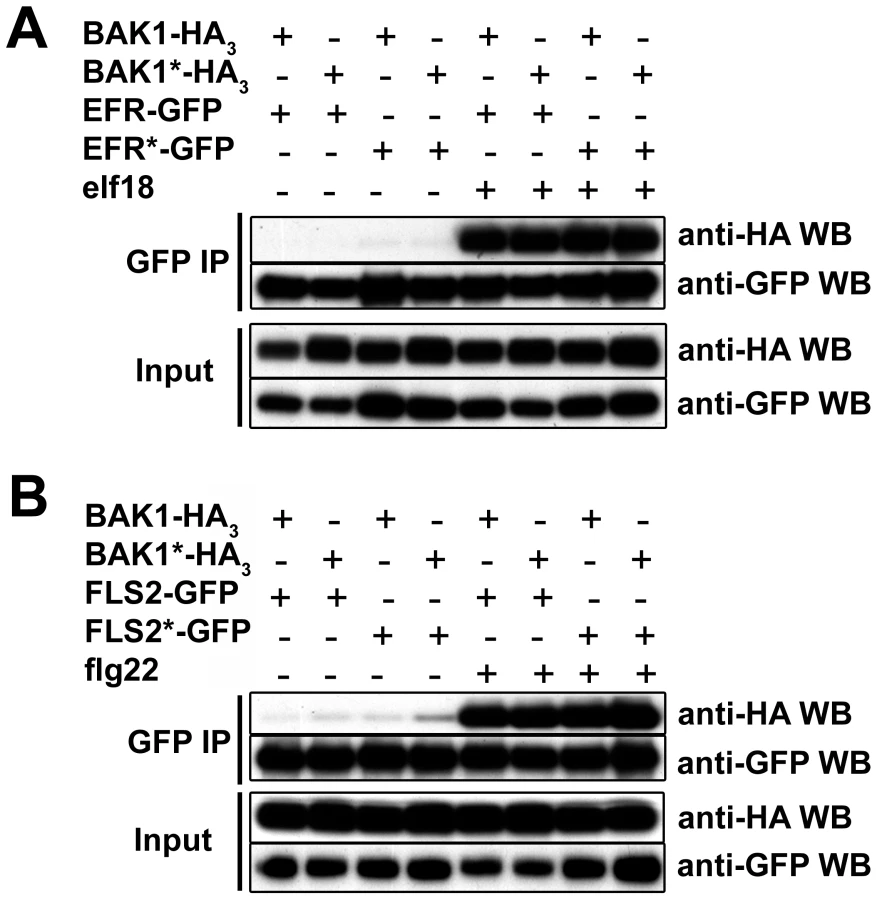
Thus, the kinase activities of neither FLS2/EFR nor BAK1 are required for their ligand-induced heteromerization.
The kinase activity of BAK1-5 is required for the bak1-5 phenotype
We tested if the kinase activity of BAK1-5 is required for the bak1-5 phenotype. As bak1-5 has the strongest differential phenotype with elf18 response when compared to bak1-4, we concentrated on EFR-dependent responses to address this question.
We created stable transgenic lines in the bak1-4 background expressing BAK1, BAK1*, BAK1-5 and BAK1-5* under the native regulatory sequence of BAK1 (Figure S11). The wild-type allele of BAK1 was able to rescue the reduced and delayed elf18-induced ROS burst of bak1-4 (Figure 9A). As previously shown (Figure 1E), expression of BAK1-5 in bak1-4 recapitulated the bak1-5 phenotype (Figure 9B). Interestingly, the expression of the kinase inactive BAK1* in bak1-4 led to a further decrease in elf18-induced ROS burst (Figure 9B), revealing a dominant-negative effect of BAK1* and demonstrating the importance of BAK1 kinase activity for downstream signaling. Strikingly, the expression of BAK1-5* in bak1-4 led to a similar dominant-negative effect as BAK1* but did not fully suppress elf18-induced ROS burst as observed in bak1-5 or when BAK1-5 was expressed in bak1-4 (Figure 9A). Similar results were observed in the SGI assay (Figure 9B).

These two observations demonstrate that BAK1-5 requires its kinase activity to quench EFR-dependent signaling. More importantly, it strongly suggests that the differential impact of the bak1-5 mutation on different signaling pathways is linked to phosphorylation.
Discussion
Plants need to correctly process diverse exogenous and endogenous information. For this purpose they rely heavily on surface localised ligand-binding RKs and regulatory RLKs. In recent years, the importance of the regulatory RLK BAK1 became apparent, as it is involved in several independent signaling pathways, namely BR responses, innate immunity and cell death control [14]. It was however unclear whether the regulatory role and the importance of BAK1 in these different biological processes are similar. Here, we clearly demonstrated that BAK1 differentially regulates these pathways in a phosphorylation-dependent manner.
Phosphorylation-dependent differential regulation of BAK1-dependent BR and PTI signaling pathways
We found that bak1-5 mutant plants are impaired in all early and late elf18 - and flg22-triggered responses tested (Figure 1 and Figure 2). Yet, bak1-5 possesses full signaling capacity for BR signaling (Figure 3). This is in contrast with previously described bak1 loss-of-function mutant alleles that are partially impaired in early and late flg22-triggered responses, but only in early responses triggered by elf18 (Figure 2; Figure S1) [26], [27]. Importantly, bak1 loss-of-function mutants are also weakly impaired in BR signaling (Figure 3; Figure S5) [18], [19]. Our initial working hypothesis for the differential regulation of BR and PTI signaling in bak1-5 was based on a potential differential interaction of BAK1-5 with the different ligand-binding RKs. However, this simple hypothesis did not hold true as BAK1-5 displays an enhanced interaction with all three ligand-binding RKs tested, namely FLS2, EFR and BRI1 (Figure 5). Therefore, we investigated the kinase activity of BAK1-5 and were able to show that BAK1-5 possesses considerable kinase activity albeit slightly reduced compared to BAK1. Importantly, BAK1-5 was still able to trans-phosphorylate both BRI1 and EFR in vitro (Figure 7B–7D). This raises the alternative hypothesis that the reduced kinase activity of BAK1-5 is sufficient to support BR but not PTI signaling. Yet several observations do not support this hypothesis. First, there is no direct correlation between the in vitro kinase activity of BAK1 mutant variants and their ability to complement either the compromised flg22-triggered SGI of bak1-4 bkk1-1 or the growth retardation phenotype of bri1-5 [21]. BAK1(T449A) is able to complement both phenotypes but has a reduced kinase activity compared to BAK1(T450A) that is not able to complement either phenotype [21]. Interestingly, BAK1-5 possesses a stronger kinase activity than BAK1(T449A) (data not shown) further substantiating this observation. Second, plants expressing the hypo-active kinase variant BAK1(Y610F) are blocked only in BR signaling but not flg22-triggered SGI [34] thereby displaying an opposite phenotype to bak1-5 plants even though both BAK1 variants are compromised in their overall kinase activity. Therefore, the quantitative kinase output of BAK1 is not the determining factor per se that enables BAK1 to function in PTI - or BR-signaling (Table S1). Third, in bak1-5 plants PTI signaling is not simply more impaired than in bak1-4 loss of function mutants but rather differentially regulated. This is exemplified in the differential MPK activation in bak1-5 plants whereby MPK3 and 6 but not MPK4 are fully activated 15 mins after ligand-treatment (Figure 2B). Fourth, BAK1-5 requires its kinase activity to fully suppress elf18-triggered ROS-burst in vivo (Figure 9B).
Altogether, this leads to the new hypothesis that BAK1-5 differentially regulates PTI - and BR-signaling pathways by discriminative auto-phosphorylation and/or trans-phosphorylation of the main-ligand binding receptors. Therefore, the qualitative kinase output of BAK1 defines its signal competence in respect to PTI - or BR - signaling pathways.
In the case of bak1-5 mutant plants, the differential auto-phosphorylation of BAK1-5 could theoretically already lead to a differential interaction surface for potential downstream signaling components. Alternatively (or concomitantly), BAK1-5 could trans-phosphorylate specific residues on EFR and FLS2 that would affect interactions with positive and/or negative regulators, such as BIK1 and related proteins [50], [52]. Phosphorylation of specific phosphosites in the intra-cellular juxta-membrane region and C-terminal tail of mammalian RTKs and Ser/Thr RKs are known to regulate signal complex composition, sub-cellular localization, receptor degradation, and therefore the initiation, amplitude, complexity and/or duration of the signal [53], . Interestingly, the rice PRR XA21 also seems to be under phosphorylation-dependent negative regulation. The ATPase XB24 interacts with XA21 in vivo, promotes XA21 auto-phosphorylation in vitro, and is a negative regulator of XA21-mediated immunity [55].
As observed previously for bak1 loss-of-function [23] and bak1-4 bkk1-1 plants expressing the phosphosite mutant variant BAK1(Y610F) [34], the basal expression level of several defence marker genes was significantly reduced in bak1-5 (Figure S3). Since BAK1-5 showed a reduced Tyr phosphorylation level in vitro (Figure 6B) BAK1-5 may be unable to auto-phosphorylate on Y610 and that BAK1 normally regulates basal gene expression via phosphorylation of this specific amino acid. Alternatively, the overall reduced kinase activity of both BAK1-5 and BAK1(Y610F) may lead to a lower constitutive basal defence signaling either induced by epiphytic bacteria and/or caused by spontaneous kinase activity [53].
Regulation of BAK1-dependent cell death control
bak1-5 is not impaired in cell death control, as bak1-5 bkk1-1 double mutant is viable and do not show any cell death or early senescence phenotypes (Figure 4 and Figure S6). This peculiarity is currently difficult to interpret, as the role of BAK1 and BKK1 in inducible and constitutive cell death control is still unclear. It was initially speculated that BAK1 and BKK1 might negatively control a ligand-binding RK perceiving a potential endogenous “survival” ligand [23], . Another LRR-RLK, BIR1, interacts with BAK1 in vivo and is strictly required for cell death control [25]. An alternative model is suggested by the constitutive cell death phenotype of bak1-4 bkk1-1 seedlings that is partially dependent on salicylic acid [24], is light-dependent [56], and the fact that the bir1-1 cell death phenotype is partially reverted by high temperatures and mutations in PAD4 and EDS1 [25], components classically associated with R protein-mediated hyper-sensitive response [57]. The integrity and/or activity of a multimeric complex containing BAK1, BKK1 and BIR1 may be “guarded” by an R protein. The absence of BAK1 and BKK1, or BIR1, would thus trigger constitutive cell death and explain the mutant seedling lethality even in sterile conditions. Interestingly though, the kinase activity of BAK1 seems to be important for cell death control, as kinase-dead variants of BAK1 cannot rescue the bak1-4 bkk1-1 lethality [21]. In this respect, it is not surprising that bak1-5 bkk1-1 is fully viable as only kinase inactive variants of BAK1 were previously shown to be unable to complement the bak1-4 bkk1-1 lethality phenotype [21], [34].
Differential regulation of RD and non-RD kinases
The differential impact of bak1-5 on BRI1-dependent and FLS2/EFR-dependent signaling could also be related to a more general differential regulation of RD versus non-RD kinases. RD kinases carry an arginine (Arg) before the conserved catalytic core Asp, and generally are activated by phosphorylation in the activation loop. The phospho-groups interact with a positively-charged pocket containing the Arg and most likely re-orient residues within the catalytic loop, ATP-binding pocket and/or facilitate peptide substrate binding [13]. In contrast, non-RD kinases do not require phosphorylation of the activation loop to adopt an active confirmation. They are regulated by different mechanisms such as relief of auto-inhibition by C-terminal extensions [58], Tyr phosphorylation in the P+1 loop [59], or are constitutively active kinases [60]. In several cases the kinase activity of non-RD kinases was shown to be at least partially dispensable for some of their functions [35], [61] suggesting a role as scaffolds. However, EFR and FLS2 require kinase activity for signaling, which implies that they do not function solely as scaffolding proteins.
The RD-kinase BRI1 was far more active in vitro in our conditions than the non-RD kinases EFR and FLS2 showing strong auto - and trans-phosphorylation capacities (Figure 7A–7B). EFR did possess some degree of auto-phosphorylation (Figure 7A, 7C), but no trans-phosphorylation capacity either towards the artificial kinase substrate MBP (Figure 7A), or towards the physiologically-relevant BAK1 kinase domain (Figure 7C). Surprisingly, we were unable to detect any in vitro activity for FLS2 CD (residues 840 to 1173) neither as N-terminal MBP-tag nor His-tag fusion protein, especially in comparison to the strong BRI1 kinase activity (Figure 7 and Figure S10). This is in contradiction to previous observations that report kinase activity of FLS2 in vitro [31], [50], [51], but is in agreement with recent publications reporting only residual kinase activity of FLS2 CD (residues 832 to 1173) and stating that recombinant FLS2 possess only weak kinase activity impeding analysis of trans-phoshorylation events in vitro [52]. Notably, close sequence analysis of the FLS2 kinase domain revealed a low conservation of the otherwise highly conserved Gly-rich loop [GxGxxG] in subdomain I, which is involved in the correct positioning of the substrate ATP [62]. Particularly, the replacement of the second invariant Gly by a Ser (S879) in FLS2 is predicted to lead to a dramatic reduction in kinase activity, as mutation of the corresponding Gly in the model Ser/Thr kinase cAPK reduces the kinase activity by 50-fold [63].
In contrast to the situation with BRI1 and BAK1, no trans-phosphorylation of BAK1 by EFR could be observed in vitro (Figure 7B–7C). Yet, BAK1 is capable of trans-phosphorylating EFR in vitro (Figure 7C). Of course, we cannot exclude that FLS2 and EFR kinase domains are only fully activated in vivo after extracellular ligand binding via conformation changes mediated by the trans-membrane domain, which is missing in the in vitro system.
Consistent with their low activity in vitro, so far no phosphosites could be identified by mass spectrometry on recombinant EFR or FLS2 CDs, even when co-incubated with BAK1 (data not shown). Even in the case of the well-studied non-RD kinase XA21, all studied phosphorylation sites were initially found by targeted mutagenesis and not by mass spectrometry analysis [64], [65]. The identification of specific phosphosites underlying the positive or negative regulation of EFR and FLS2 therefore remain a real technical challenge.
Nevertheless, the kinase activities and some potential phosphosites of FLS2 and EFR are important for several downstream signaling events. A kinase-dead version of EFR (EFR*) is unable to confer elf18-triggered ROS burst when transiently expressed in N. benthamiana (Figure S12). A K898M mutation in the FLS2 kinase domain abolished MPK3 and MPK6 activation by flg22 after transient over-expression in fls2 mutant protoplasts [66]. Targeted mutagenesis of potential phosphosites in FLS2 revealed that T867, T1040 and T1072 are required for its full functionality [67]. However, it was not investigated if these sites are required for kinase activity, are auto-phosphorylation sites, or whether they represent trans-phophorylation targets of BAK1.
Overall, the striking difference between the kinase activities of the two RD kinases BRI1 and BAK1 compared to the non-RD kinases EFR and FLS2 suggests a different regulatory mechanism between these two kinase classes. A highly conserved Thr residue in the intracellular juxta-membrane domain reveals a differential regulation of the overall kinase activity of RD and non-RD kinase by a single site. Accordingly, T705 of the non-RD kinase XA21 is essential for in vitro auto-phosphorylation, interaction with downstream signaling components, and for XA21-mediated resistance [65]. Similarly, a mutation of the corresponding residue in FLS2 (T867) compromised its function in planta [67]. However, in the case of the RD kinase BRI1 the phosphorylation of the corresponding Thr (T880) is not required for its function [20]. These results suggest that the regulation of auto-phosphorylation of non-RD kinases by phosphosites in the intra-cellular juxta-membrane region may play an important role in the recruitment of downstream signaling components, as suggested in [65].
Another difference between RD and non-RD RK seems to be the requirement of kinase activity for complex formation with the RD-RLK BAK1. We found that the kinase activity of neither interaction partner is required for the ligand-induced interaction of FLS2 or EFR with BAK1 (Figure 8). Optimal ligand-dependent heteromerization could even be induced between double mutant combinations of FLS2* or EFR* with BAK1* (Figure 8). These results obtained after transient over-expression in N. benthamiana nicely complement previous pharmacological studies in A. thaliana cell cultures [30]. Treatment of cell cultures with the broad-range kinase inhibitor K252a did not block FLS2-BAK1 complex formation, but totally inhibited phosphorylation of either of the interaction partners. Ligand-dependent conformational changes thus seem sufficient to trigger heteromerization between the non-RD kinases EFR and FLS2 with BAK1. Therefore, the interaction of EFR and FLS2 with BAK1 is a requirement rather than a consequence of detectable phosphorylation. This situation is in stark contrast with the absolute requirement of the BRI1 kinase activity for the ligand-induced complex formation with BAK1 in planta [21].
Conclusions
BAK1 is able to dictate specificity of downstream signaling as BAK1-5 nearly totally blocked FLS2 - and EFR-mediated PTI signaling but barely influenced cell death control and BRI1-mediated BR signaling. Based on these results and the recent work from Schulze and colleagues [30], we propose a model for the mechanisms underlying EFR/FLS2 heteromerization with BAK1, and the role of BAK1 in the establishment of PTI signaling. EFR and FLS2 most likely exist in close proximity with BAK1 at the plasma membrane in loose pre-formed complexes due to their near instantaneous oligomerization after ligand binding [30]. Conformational changes triggered by ligand binding lead to the stabilization of the complex. This interaction is kinase-independent, but may lead to the activation of the EFR/FLS2 kinase activity by BAK1 via trans-phosphorylation events. Phosphorylation of specific residues on EFR/FLS2 and/or BAK1 leads to the recruitment of downstream signaling components that dictate the specificity of the signaling output. In this model, BAK1 is not a simple enhancer of the kinase activity of the ligand-binding RKs, but is an integral part of the signaling pathway.
Future studies need to carefully address the role of kinase activity of non-RD kinases for PTI signaling and final defence outcomes. Therefore, careful qualitative and quantitative analyses guided by mass-spectrometry of the phosphorylation status of BAK1, BAK1-5, FLS2 and EFR in vitro and in vivo will shed more light onto the complex regulatory mechanisms of these two model non-RD PRRs by the regulatory RLK BAK1. These studies are however technically challenging, as unlike BRI1, the kinase activity of EFR is very weak and that of FLS2 is practically negligible at least in vitro.
Methods
Plant material and methods
All mutants and transgenic lines used in this study were in the background of A. thaliana ecotype Columbia (Col-0). The Arabidopsis plants were grown on soil or MS salt medium (Duchefa), 1% sucrose and 1% agar with a 10 H or 16 H photoperiod at 20–22°C. The third backcross of bak1-5 with Col-0 was used for all experiments.
The mutants bak1-4, bkk1-1, bri1-301 were previously described [24], [27], [47]. The double mutants bak1-4 bkk1-1, bak1-5 bkk1-1, bak1-4 bri1-301, and bak1-5 bri1-301 were generated by crossing and genotyped using the primers listed in Table S2.
bak1-5 marker design
For bak1-5 homozygous mutant identification a dCAPS marker was designed using dCAPS Finder 2.0 [68]. The genomic region around the bak1-5 mutation was PCR amplified using Taq polymerase (Qiagen) and the primers listed in Table S2. The corresponding product was cut with RsaI (NEB) and bak1-5 derived PCR products contained an additional RsaI site in addition to the internal restriction control site.
Generation of transgenic plants
The genomic fragment of BAK1, including the promoter and the coding region, in pDONR201 (Invitrogen) was a gift from B. Kemmerling [23]. The corresponding point mutations for BAK1*, BAK1-5, and BAK1-5* were introduced by point mutagenesis PCR using primers given in Table S2. The PCR product was digested with 1.5 µl DpnI (NEB) overnight and subsequently transformed into Escherichia coli DH5α. The presence of the corresponding mutations and the integrity of the genomic fragments were verified by sequencing. The correct clones were used to transfer the inserts into pGWB2 [69] using GATEWAY LR CLONASE II enzyme (Invitrogen). The resulting constructs were verified by restriction analysis and electroporated into Agrobacterium tumefaciens strain AglI.
All constructs were transformed into Arabidopsis mutant bak1-4 using the floral dipping method [70]. Transformants were selected on MS agar medium containing 40 µg/ml hygromycin.
In vitro protein analysis
Molecular cloning
The kinase domain of BAK1 in the pGEMTeasy vector was a gift from Sacco de Vries [71]. The corresponding point mutations for BAK1*, BAK1-5, and BAK1-5* were introduced as described above using primers given in Table S2. The inserts of sequence verified clones were transferred into pGEX-4T1 using EcoRI and XhoI (NEB) to generate N-terminal GST fusion constructs.
The kinase domain of BRI1, FLS2 and EFR were PCR amplified using the primers given in Table S2. The resulting PCR products were cloned either into pOPINM or pOPINF [72] using the IN-FUSION reagent (Clontech) to obtain N-terminal MBP or His fusion constructs, respectively. The resulting constructs were verified by restriction analysis and sequencing. The corresponding point mutations of BRI1*, FLS2*, and EFR* were obtained as described above using primers given in Table S2.
Recombinant protein purification
Recombinant fusion proteins were produced in E. coli BL21 (Novagen), extracted using BugBuster reagent (Novagen) containing 1 µl/ml Benzoase (Novagen), 1 KU/ml Lysozyme (Novagen) and 150 µl/ml protease inhibitor cocktail set II (Novagen) and the soluble fraction was used to enrich for fusion proteins. GST-tagged fusion proteins (GST-BAK1, GST-BAK1*, GST-BAK1-5, GST-BAK1-5*) were enriched using Glutathione Sepharose Fast Flow (GE Healthcare) according to the manufactures protocol. MBP-tagged fusion proteins (MBP-BRI1, MBP-BRI1*, MBP-FLS2, MBP-FLS*, MBP-EFR, MBP-EFR*) were enriched using Amylose Resin (NEB) according to manufactures protocol. His-tag fusion proteins (His-BRI1, His-BRI1*, His-EFR, His-EFR*) were enriched using His-Bind Resin (Novagen) according to the manufactures protocol. After elusion fusion proteins were adjusted to the same concentration in 10% glycerol solution and stored at −20°C until usage.
In vitro kinase assay
The fusion proteins were incubated in 30 µl kinase buffer (50 mM Tris, pH 7.5, 10 mM MgCl2, 10 mM MnCl2, 1 mM DTT) in the presence of only 1 µM unlabeled ATP or 1 µM unlabeled ATP and 183 kB of [32P]-γ-ATP for 30 min at 30°C with shaking at 900 rpm. The reactions were stopped by adding 2xLDS loading buffer (Invitrogen). The phosphorylation status of fusion proteins was analyzed by audioradiography after separation of one-fourth of the in vitro kinase assay by SDS-PAGE followed by western blotting, if not indicated otherwise. In autophosphorylation assays 1 µg fusion protein for MBP - and GST-tagged proteins and 5 µg for His-tagged proteins was incubated with 1 µg of MBP (Fluka). In transphosphorylation assays 1 µg of each fusion protein was used.
For Km determination, in vitro kinase assays were performed as previously described [73]. Post electrophoresis, proteins were transferred onto PVDF membranes. Subsequently, the membranes were subjected to autoradiography using a FUJI Film FLA5000 PhosphorImager (Fuji, Tokyo, Japan) to estimate relative activities.
Phosphorylation site analysis
The indicated amount of fusion proteins (GST-BAK1, GST-BAK1*, GST-BAK1-5, GST-BAK1-5*) were separated by SDS-PAGE and blotted onto PVDF membrane (Biorad). The immunoblots were blocked in 5% (w/v) BSA (Sigma) in TBS-Tween (0.1%) for 1–2 H. Phospho-Serine/Threonine sites were detected using anti-p-Thr (1∶1000, Cell Signaling Technology) overnight, followed by anti-mouse-HRP conjugated secondary antibodies (1∶5000, Sigma). Phospho-Tyrosine sites were detected using anti-p-Tyr (1∶2000, Cell Signaling Technology) overnight, followed by anti-rabbit-HRP conjugated secondary antibodies (1∶5000, Sigma).
qRT-PCR
14-days-old seedlings grown for five days on MS plates and than transferred to liquid MS were used for all gene induction studies. RNA was extracted using RNeasy Plant Mini kit (Qiagen) followed by DNase-treatment using Turbo DNA-free (Ambion) and quantified with a Nanodrop spectrophotometer (Thermo scientific). cDNA was synthesized from 2.5 µg total RNA using SuperScript III reverse transcriptase (Invitrogen). SybrGreen master mix (Sigma) was used for qPCR reactions.
For defence gene induction analysis a triplicate of two seedlings each was treated either with water, 100 nM elf18 or 100 nM flg22 for 0, 30, 60 and 180 min and pooled before harvesting. Gene expression of At2g17740 (DC1-domain containing protein), At5g57220 (CYP81F2) and At1g51890 (LRR-RLK) was monitored by qPCR analysis. The expression of each marker gene was normalized to the internal reference gene At4g05320 (UBQ10) and plotted relative to the Col-0 steady-state expression level.
For BR gene expression analysis a triplicate of two seedlings each was treated with either mock solvent control or 2.5 µM BRZ (Sigma) for 16 H over night. The next morning samples were further treated with mock solvent control or 200 nM brassinolide (SRICI) for another three hours before being pooled for harvesting. Gene expression of At2g40610 (EXP8) and At4g38850 (SAUR-ACI) was monitored by qPCR analysis. The expression of each gene was normalized to the internal reference gene At5g15400 (U-box containing protein) and plotted relative to the Col-0 double mock treated expression level.
Hypocotyl growth assay
Freshly harvested seeds were surface sterilized and stratified in sterile water at 4°C for 4–6 days in the dark. Individual seeds were put on ½ MS containing 0.8% phytoagar (Duchefa) without hormone, with 100 nM BL or with 100 nM BRZ and left up-right in the dark at 20–22°C. Hypocotyl length was measured after 5-day incubation.
Bacterial infection assays
The P. syringae pv. tomato DC3000 COR− (Pto DC3000 COR−) [40] strain was grown in overnight culture in Kings B medium supplemented with appropriate antibiotics. Cells were harvested by centrifugation and pellets re-suspended in sterile water to OD600 = 0.2. Immediately prior to spraying, Silwett L-77 was added to bacteria to a concentration of 0.04% (v/v). Bacteria were sprayed onto leaf surfaces until run-off and plants covered for 3 days. Samples were taken using a cork-borer (2 mm) to cut leaf discs from 2 leaves per plant and 4 plants per genotype. Leaf discs were ground in water, diluted and plated on TSA medium with appropriate selection. Plates were incubated at 28°C and colonies counted 2 days later.
MAP kinase assay
14-days-old seedlings were grown for five days on MS plates and than transferred to liquid MS. Triplicates of two seedlings each were treated with water, 100 nM elf18 or 100 nM flg22 for 0, 5 and 15 min before being pooled for harvest. Seedlings were ground to fine powder in liquid nitrogen and solubilised in better lacus buffer [50 mM Tris-HCl pH 7.5; 100 mM NaCl; 15 mM EGTA; 10 mM MgCl2; 1 mM NaF; 1 mM Na2MoO4.2H2O; 0.5 mM NaVO3; 30 mM β-glycerophosphate; 0.1% IGEPAL CA 630; 100 nM calyculin A (CST); 0.5 mM PMSF; 1% protease inhibitor cocktail (Sigma, P9599)]. The extracts were centrifuged at 16,000×g, the supernatant cleared by filtering through Miracloth and 4xLDS loading buffer (Invitrogen) added. 40 µg of total protein was separated by SDS-PAGE and blotted onto PVDF membrane (Biorad). Immunoblots were blocked in 5% (w/v) BSA (Sigma) in TBS-Tween (0.1%) for 1–2 H. The activated MAP kinases were detected using anti-p42/44 MAPK primary antibodies (1∶1000, Cell Signaling Technology) overnight, followed by anti-rabbit-HRP conjugated secondary antibodies (Sigma).
Seedling growth inhibition
Fresh harvested seeds were surface sterilized, sown on MS media, stratified for 2 days at 4°C in the dark and put in the light. Five-day-old seedlings were transferred into liquid MS with or without the indicated amount of peptide and incubated for eight further days. Dry weight of six replicates per treatment was measured using a precision scale (Sartorius) and blotted relative to untreated control.
ROS burst assay
Eight leaf discs (4 mm diameter) of at least four 3–4 week plants were sampled using a cork borer and floated over night on sterile water. The following day the water was replaced with a solution of 17 mg/ml (w/v) luminol (Sigma) and 10 mg/ml horseradish peroxidase (Sigma) containing 100 nM elf18 or 100 nM flg22. Luminescence was captured either using a Varioskan Flash (Thermo Scientific) multiplate reader or Photek camera (East Sussex, UK). The amount of relative light units might differ depending on the light capturing apparatus used.
Transient expression in N. benthamiana
The whole coding sequence without the stop codon of FLS2, EFR, BAK1 and BAK1-5 was PCR amplified using the primers given in Table S2 and cloned into the pENTR-D/TOPO vector using the pENTR Directional TOPO cloning kit (Invitrogen). The resulting clones were verified by restriction analysis and sequence. The kinase dead variants FLS2*, EFR* and BAK1* were generated by point mutagenesis using the primers given in Table S2 and sequence verified. The coding sequences of FLS2, FLS2*, EFR and EFR* were transferred into pEarleyGate103 [74] using the method described for Gateway vectors generating C-terminal GFP-His-tag fusion constructs under the 35S promoter. The coding sequence of BAK1, BAK1* and BAK1-5 were transferred into pGWB14 generating C-terminal HA-tag fusion constructs under the 35S promoter. The CERK1p::CERK1-3xHA construct was previously published [75]. The EFRp::EFR-3xHA construct, containing own promoter plus coding region, was described previously with the exception of using epiGreenB5 as binary vector [36]. All resulting constructs were verified by restriction analysis and transformed into A. tumefaciens strain GV3101.
The constructs of BAK1p::BAK1-eGFP or BAK1p::BAK1-5-eGFP, containing own promoter plus coding regions, were PCR amplified using primers given in Table S2. The resulting constructs were cloned into pCR-Blunt-II-TOPO (Invitrogen) and verified by sequencing. The inserts were released by digesting with BsmBI and BamHI (NEB) and ligated into epiGreenB(eGFP) digested with EcoRI and BamHI (NEB). Resulting constructs were verified by restriction analysis transformed into A. tumefaciens strain AglI containing the pSOUP helper plasmid.
A. tumefaciens containing the indicated constructs were grown in L medium supplemented with the appropriate antibiotics overnight. Cultures were spun down and resuspended in 10 mM MgCl2 to a final O.D.600 = 0.2-0.5. The indicated cultures were mixed 1∶1 and syringe infiltrated into 3-week-old N. benthamiana leaves. After 2 dpi whole leaves were vacuum infiltrated with water or 100 nM of the indicated peptide, incubated for 5 min and harvested by freezing in liquid nitrogen.
Protein extraction and immunoprecipitation in N. benthamiana
Leaves were ground to fine powder in liquid nitrogen and 5 ml extraction buffer [50 mM Tris-HCl pH 7.5; 150 mM NaCl; 10% glycerol; 10 mM DTT; 10 mM EDTA; 1 mM NaF; 1 mM Na2MoO4.2H2O; 1% (w/v) PVPP; 1% (v/v) P9599 protease inhibitor cocktail (Sigma); 1% (v/v) IGEPAL CA-630 (Sigma)] added. Samples were cleared by centrifugation at 16.000×g for 15 min at 4°C and adjusted to 2 mg/ml total protein concentration. Immunoprecipitation were performed on 1.5 ml total protein by adding 20 µl GFPTrap-A beads (Chromotek) and incubation at 4°C for 3–4 H. Beads were washed 4 times with TBS containing 0.5% (v/v) IGEPAL CA-630, immunoprecipitates eluted with 30 µl 2xLDS (Invitrogen) and heating at 70°C for 10 min.
SDS-PAGE and immunoblotting
SDS-gels were prepared with either 7.5 or 10% cross-linking. Gels were run at 80/150 V and proteins electroblotted onto PVDF membrane at 235 mA (Biorad). Membranes were rinsed in TBS and blocked in 5% (w/v) nonfat milk powder in TBST 0.1% (w/v) for 1 H. Primary antibodies were diluted in blocking solution to the following concentration and incubated overnight: anti-GFP (AMS Biotechnology) 1∶5000; anti-BAK1 1∶500; anti-HA-HRP (Santa Cruz) 1∶2000; anti-FLS2 1∶1000; anti-BRI1 1∶1000. Membranes were washed 3 times in TBST 0.1% (w/v) before 1 hour incubation with secondary antibodies anti-rabbit-HRP (Sigma) 1∶5000 or anti-rabbit-HRP (Ebioscience) 1∶5000. Signals were visualized using chemiluminescent substrate (Lumigen ECL, GE Healthcare) before exposure to film (AGFA CP-BU).
Protein extraction and immunoprecipitation in Arabidopsis
Leaves were ground to fine powder in liquid nitrogen and extraction buffer [50 mM Tris-HCl pH 7.5; 150 mM NaCl; 10% glycerol; 5 mM DTT; 2 mM EDTA; 1 mM NaF; 1 mM Na2MoO4.2H2O; 1 mM PMSF (Sigma); 5 mM Na3VO4, 1% (v/v) P9599 protease inhibitor cocktail (Sigma); 1% (v/v) IGEPAL CA-630 (Sigma)] added. Samples were cleared by centrifugation at 16.000×g for 15 min at 4°C and adjusted to 2 mg/ml total protein concentration. Immunoprecipitations were performed on 1.5 ml total protein by adding 20 µl true-blot anti-rabbit Ig beads (Ebioscience), 15 µl antibody and incubation at 4°C for 3–4 H. Beads were washed 4 times with TBS containing 0.5% (v/v) IGEPAL CA-630, immunoprecipitates eluted with 50 µl 2xLDS (Invitrogen) and heated at 70°C for 10 min.
Supporting Information
Zdroje
1. ShiuSHKarlowskiWMPanRTzengYHMayerKF
2004
Comparative analysis of the receptor-like kinase family in
Arabidopsis and rice.
Plant Cell
16
1220
1234
2. OuyangSQLiuYFLiuPLeiGHeSJ
2010
Receptor-like kinase OsSIK1 improves drought and salt stress
tolerance in rice (Oryza sativa) plants.
Plant J
3. De SmetIVossUJurgensGBeeckmanT
2009
Receptor-like kinases shape the plant.
Nat Cell Biol
11
1166
1173
4. BollerTFelixG
2009
A renaissance of elicitors: perception of microbe-associated
molecular patterns and danger signals by pattern-recognition
receptors.
Annu Rev Plant Biol
60
379
406
5. CitriAYardenY
2006
EGF-ERBB signalling: towards the systems level.
Nat Rev Mol Cell Biol
7
505
516
6. JorissenRNWalkerFPouliotNGarrettTPWardCW
2003
Epidermal growth factor receptor: mechanisms of activation and
signalling.
Exp Cell Res
284
31
53
7. WardCWLawrenceMCStreltsovVAAdamsTEMcKernNM
2007
The insulin and EGF receptor structures: new insights into
ligand-induced receptor activation.
Trends Biochem Sci
32
129
137
8. BoseRZhangX
2009
The ErbB kinase domain: structural perspectives into kinase
activation and inhibition.
Exp Cell Res
315
649
658
9. JuraNEndresNFEngelKDeindlSDasR
2009
Mechanism for activation of the EGF receptor catalytic domain by
the juxtamembrane segment.
Cell
137
1293
1307
10. Red BrewerMChoiSHAlvaradoDMoravcevicKPozziA
2009
The juxtamembrane region of the EGF receptor functions as an
activation domain.
Mol Cell
34
641
651
11. MorandellSStasykTSkvortsovSAscherSHuberLA
2008
Quantitative proteomics and phosphoproteomics reveal novel
insights into complexity and dynamics of the EGFR signaling
network.
Proteomics
8
4383
4401
12. JohnsonLNNobleMEOwenDJ
1996
Active and inactive protein kinases: structural basis for
regulation.
Cell
85
149
158
13. NolenBTaylorSGhoshG
2004
Regulation of protein kinases; controlling activity through
activation segment conformation.
Mol Cell
15
661
675
14. ChinchillaDShanLHePde VriesSKemmerlingB
2009
One for all: the receptor-associated kinase BAK1.
Trends Plant Sci
14
535
541
15. PostelSKufnerIBeuterCMazzottaSSchwedtA
2009
The multifunctional leucine-rich repeat receptor kinase BAK1 is
implicated in Arabidopsis development and immunity.
Eur J Cell Biol
16. HechtVVielle-CalzadaJ-PHartogMVSchmidtEDLBoutilierK
2001
The Arabidopsis Somatic Embryogenesis Receptor Kinase 1 Gene Is
Expressed in Developing Ovules and Embryos and Enhances Embryogenic
Competence in Culture.
Plant Physiol
127
803
816
17. AlbrechtCRussinovaEKemmerlingBKwaaitaalMde VriesSC
2008
Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR KINASE proteins serve
brassinosteroid-dependent and -independent signaling
pathways.
Plant Physiol
148
611
619
18. NamKHLiJ
2002
BRI1/BAK1, a receptor kinase pair mediating brassinosteroid
signaling.
Cell
110
203
212
19. LiJWenJLeaseKADokeJTTaxFE
2002
BAK1, an Arabidopsis LRR receptor-like protein kinase, interacts
with BRI1 and modulates brassinosteroid signaling.
Cell
110
213
222
20. WangXGosheMBSoderblomEJPhinneyBSKucharJA
2005
Identification and functional analysis of in vivo phosphorylation
sites of the Arabidopsis BRASSINOSTEROID-INSENSITIVE1 receptor
kinase.
Plant Cell
17
1685
1703
21. WangXKotaUHeKBlackburnKLiJ
2008
Sequential transphosphorylation of the BRI1/BAK1 receptor kinase
complex impacts early events in brassinosteroid signaling.
Dev Cell
15
220
235
22. JeongYJShangYKimBHKimSYSongJH
2010
BAK7 displays unequal genetic redundancy with BAK1 in
brassinosteroid signaling and early senescence in
arabidopsis.
Mol Cells
29
259
266
23. KemmerlingBSchwedtARodriguezPMazzottaSFrankM
2007
The BRI1-associated kinase 1, BAK1, has a
brassinolide-independent role in plant cell-death control.
Curr Biol
17
1116
1122
24. HeKGouXYuanTLinHAsamiT
2007
BAK1 and BKK1 regulate brassinosteroid-dependent growth and
brassinosteroid-independent cell-death pathways.
Curr Biol
17
1109
1115
25. GaoMWangXWangDXuFDingX
2009
Regulation of cell death and innate immunity by two receptor-like
kinases in Arabidopsis.
Cell Host Microbe
6
34
44
26. HeeseAHannDRGimenez-IbanezSJonesAMHeK
2007
The receptor-like kinase SERK3/BAK1 is a central regulator of
innate immunity in plants.
Proc Natl Acad Sci U S A
104
12217
12222
27. ChinchillaDZipfelCRobatzekSKemmerlingBNurnbergerT
2007
A flagellin-induced complex of the receptor FLS2 and BAK1
initiates plant defence.
Nature
448
497
500
28. KrolEMentzelTChinchillaDBollerTFelixG
2010
Perception of the Arabidopsis danger signal peptide 1 involves
the pattern recognition receptor AtPEPR1 and its close homologue
AtPEPR2.
J Biol Chem
285
13471
13479
29. ShanLHePLiJHeeseAPeckSC
2008
Bacterial effectors target the common signaling partner BAK1 to
disrupt multiple MAMP receptor-signaling complexes and impede plant
immunity.
Cell Host Microbe
4
17
27
30. SchulzeBMentzelTJehleAKMuellerKBeelerS
2010
Rapid heteromerization and phosphorylation of ligand-activated
plant transmembrane receptors and their associated kinase
BAK1.
J Biol Chem
285
9444
9451
31. XiangTZongNZouYWuYZhangJ
2008
Pseudomonas syringae effector AvrPto blocks innate immunity by
targeting receptor kinases.
Curr Biol
18
74
80
32. XiangTZongNZhangJChenJChenM
2011
BAK1 is not a target of the Pseudomonas syringae effector
AvrPto.
Mol Plant Microbe Interact
24
100
107
33. KinoshitaTCano-DelgadoASetoHHiranumaSFujiokaS
2005
Binding of brassinosteroids to the extracellular domain of plant
receptor kinase BRI1.
Nature
433
167
171
34. OhMHWangXWuXZhaoYClouseSD
2010
Autophosphorylation of Tyr-610 in the receptor kinase BAK1 plays
a role in brassinosteroid signaling and basal defense gene
expression.
Proc Natl Acad Sci U S A
107
17827
17832
35. DardickCRonaldP
2006
Plant and animal pathogen recognition receptors signal through
non-RD kinases.
PLoS Pathog
2
e2
doi:10.1371/journal.ppat.0020002
36. NekrasovVLiJBatouxMRouxMChuZH
2009
Control of the pattern-recognition receptor EFR by an ER protein
complex in plant immunity.
Embo J
28
3428
3438
37. BoudsocqMWillmannMRMcCormackMLeeHShanL
2010
Differential innate immune signalling via Ca(2+) sensor
protein kinases.
Nature
464
418
422
38. FiilBKPetersenKPetersenMMundyJ
2009
Gene regulation by MAP kinase cascades.
Curr Opin Plant Biol
12
615
621
39. HePShanLLinNCMartinGBKemmerlingB
2006
Specific bacterial suppressors of MAMP signaling upstream of
MAPKKK in Arabidopsis innate immunity.
Cell
125
563
575
40. MelottoMUnderwoodWKoczanJNomuraKHeSY
2006
Plant stomata function in innate immunity against bacterial
invasion.
Cell
126
969
980
41. ThompsonMJMeudtWJMandavaNBDutkySRLusbyWR
1982
Synthesis of brassinosteroids and relationship of structure to
plant growth-promoting effects.
Steroids
39
89
105
42. NagataNAsamiTYoshidaS
2001
Brassinazole, an inhibitor of brassinosteroid biosynthesis,
inhibits development of secondary xylem in cress plants (Lepidium
sativum).
Plant Cell Physiol
42
1006
1011
43. MussigCLissoJColl-GarciaDAltmannT
2006
Molecular analysis of brassinosteroid action.
Plant Biol (Stuttg)
8
291
296
44. GodaHSawaSAsamiTFujiokaSShimadaY
2004
Comprehensive comparison of auxin-regulated and
brassinosteroid-regulated genes in Arabidopsis.
Plant Physiol
134
1555
1573
45. WangZYNakanoTGendronJHeJChenM
2002
Nuclear-localized BZR1 mediates brassinosteroid-induced growth
and feedback suppression of brassinosteroid biosynthesis.
Dev Cell
2
505
513
46. KimTWWangZY
2010
Brassinosteroid signal transduction from receptor kinases to
transcription factors.
Annu Rev Plant Biol
61
681
704
47. XuWHuangJLiBLiJWangY
2008
Is kinase activity essential for biological functions of
BRI1?
Cell Res
18
472
478
48. MiyaAAlbertPShinyaTDesakiYIchimuraK
2007
CERK1, a LysM receptor kinase, is essential for chitin elicitor
signaling in Arabidopsis.
Proc Natl Acad Sci U S A
104
19613
19618
49. WanJZhangXCNeeceDRamonellKMCloughS
2008
A LysM receptor-like kinase plays a critical role in chitin
signaling and fungal resistance in Arabidopsis.
Plant Cell
20
471
481
50. LuDWuSGaoXZhangYShanL
2010
A receptor-like cytoplasmic kinase, BIK1, associates with a
flagellin receptor complex to initiate plant innate
immunity.
Proc Natl Acad Sci U S A
107
496
501
51. Gomez-GomezLBauerZBollerT
2001
Both the extracellular leucine-rich repeat domain and the kinase
activity of FSL2 are required for flagellin binding and signaling in
Arabidopsis.
Plant Cell
13
1155
1163
52. ZhangJLiWXiangTLiuZLalukK
2010
Receptor-like cytoplasmic kinases integrate signaling from
multiple plant immune receptors and are targeted by a Pseudomonas syringae
effector.
Cell Host Microbe
7
290
301
53. LemmonMASchlessingerJ
2010
Cell signaling by receptor tyrosine kinases.
Cell
141
1117
1134
54. BirchmeierCBirchmeierWGherardiEVande WoudeGF
2003
Met, metastasis, motility and more.
Nat Rev Mol Cell Biol
4
915
925
55. ChenXChernMCanlasPERuanDJiangC
2010
An ATPase promotes autophosphorylation of the pattern recognition
receptor XA21 and inhibits XA21-mediated immunity.
Proc Natl Acad Sci U S A
107
8029
8034
56. HeKGouXPowellRAYangHYuanT
2008
Receptor-like protein kinases, BAK1 and BKK1, regulate a
light-dependent cell-death control pathway.
Plant Signal Behav
3
813
815
57. DoddsPNRathjenJP
2010
Plant immunity: towards an integrated view of plant-pathogen
interactions.
Nat Rev Genet
11
539
548
58. KobeBHeierhorstJFeilSCParkerMWBenianGM
1996
Giant protein kinases: domain interactions and structural basis
of autoregulation.
Embo J
15
6810
6821
59. MayansOvan der VenPFWilmMMuesAYoungP
1998
Structural basis for activation of the titin kinase domain during
myofibrillogenesis.
Nature
395
863
869
60. NolenBYunCYWongCFMcCammonJAFuXD
2001
The structure of Sky1p reveals a novel mechanism for constitutive
activity.
Nat Struct Biol
8
176
183
61. MeylanETschoppJ
2005
The RIP kinases: crucial integrators of cellular
stress.
Trends Biochem Sci
30
151
159
62. BossemeyerD
1994
The glycine-rich sequence of protein kinases: a multifunctional
element.
Trends Biochem Sci
19
201
205
63. TaylorSSRadzio-AndzelmEMadhusudanChengXTen EyckL
1999
Catalytic subunit of cyclic AMP-dependent protein kinase:
structure and dynamics of the active site cleft.
Pharmacol Ther
82
133
141
64. XuWHWangYSLiuGZChenXTinjuangjunP
2006
The autophosphorylated Ser686, Thr688, and Ser689 residues in the
intracellular juxtamembrane domain of XA21 are implicated in stability
control of rice receptor-like kinase.
Plant J
45
740
751
65. ChenXChernMCanlasPEJiangCRuanD
2010
A conserved threonine residue in the juxtamembrane domain of the
XA21 pattern recognition receptor is critical for kinase autophosphorylation
and XA21-mediated immunity.
J Biol Chem
66. AsaiTTenaGPlotnikovaJWillmannMRChiuWL
2002
MAP kinase signalling cascade in Arabidopsis innate
immunity.
Nature
415
977
983
67. RobatzekSChinchillaDBollerT
2006
Ligand-induced endocytosis of the pattern recognition receptor
FLS2 in Arabidopsis.
Genes Dev
20
537
542
68. NeffMMTurkEKalishmanM
2002
Web-based primer design for single nucleotide polymorphism
analysis.
Trends Genet
18
613
615
69. NakagawaTKuroseTHinoTTanakaKKawamukaiM
2007
Development of series of gateway binary vectors, pGWBs, for
realizing efficient construction of fusion genes for plant
transformation.
J Biosci Bioeng
104
34
41
70. CloughSJBentAF
1998
Floral dip: a simplified method for Agrobacterium-mediated
transformation of Arabidopsis thaliana.
Plant J
16
735
743
71. KarlovaRBoerenSvan DongenWKwaaitaalMAkerJ
2009
Identification of in vitro phosphorylation sites in the
Arabidopsis thaliana somatic embryogenesis receptor-like
kinases.
Proteomics
9
368
379
72. BerrowNSAldertonDSainsburySNettleshipJAssenbergR
2007
A versatile ligation-independent cloning method suitable for
high-throughput expression screening applications.
Nucleic Acids Res
35
e45
73. NtoukakisVMucynTSGimenez-IbanezSChapmanHCGutierrezJR
2009
Host inhibition of a bacterial virulence effector triggers
immunity to infection.
Science 2009
324
784
787
74. EarleyKWHaagJRPontesOOpperKJuehneT
2006
Gateway-compatible vectors for plant functional genomics and
proteomics.
Plant J
45
616
629
75. Gimenez-IbanezSHannDRNtoukakisVPetutschnigELipkaV
2009
AvrPtoB targets the LysM receptor kinase CERK1 to promote
bacterial virulence on plants.
Curr Biol
19
423
429
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2011 Číslo 4
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Survival Motor Neuron Protein Regulates Stem Cell Division, Proliferation, and Differentiation in
- PTG Depletion Removes Lafora Bodies and Rescues the Fatal Epilepsy of Lafora Disease
- Evolution of Vertebrate Transient Receptor Potential Vanilloid 3 Channels: Opposite Temperature Sensitivity between Mammals and Western Clawed Frogs
- An Evolutionary Genomic Approach to Identify Genes Involved in Human Birth Timing
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy