
The Vi Capsular Polysaccharide Enables Serovar Typhi to Evade Microbe-Guided Neutrophil Chemotaxis
To control a bacterial infection, neutrophils have to first migrate toward the microbe and then ingest and kill the intruder. Since S. Typhi has a greater propensity than S. Typhimurium to disseminate from its port of entry, we investigated whether both pathogens differ in their ability to evade neutrophil chemotaxis. Remarkably, S. Typhi, but not S. Typhimurium, inhibited neutrophil chemotaxis. Elucidation of the underlying mechanism revealed that microbe-guided chemotaxis proceeded through a C5a-dependent mechanism, which could be blocked by the Vi capsular polysaccharide of S. Typhi. These data suggest that the chemotactic chase of neutrophils is a host defense mechanism operational during gastroenteritis, but not during the initial stages of typhoid fever.
Published in the journal:
. PLoS Pathog 10(8): e32767. doi:10.1371/journal.ppat.1004306
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1004306
Summary
To control a bacterial infection, neutrophils have to first migrate toward the microbe and then ingest and kill the intruder. Since S. Typhi has a greater propensity than S. Typhimurium to disseminate from its port of entry, we investigated whether both pathogens differ in their ability to evade neutrophil chemotaxis. Remarkably, S. Typhi, but not S. Typhimurium, inhibited neutrophil chemotaxis. Elucidation of the underlying mechanism revealed that microbe-guided chemotaxis proceeded through a C5a-dependent mechanism, which could be blocked by the Vi capsular polysaccharide of S. Typhi. These data suggest that the chemotactic chase of neutrophils is a host defense mechanism operational during gastroenteritis, but not during the initial stages of typhoid fever.
Introduction
Salmonella enterica serovar Typhi (S. Typhi) is a strictly human-adapted pathogen associated with a disseminated febrile illness, termed typhoid fever (reviewed in [1]). In contrast, S. enterica serovar Typhimurium causes an infection that manifests as a localized gastroenteritis in immunocompetent individuals (Reviewed in [2]). Several S. Typhi virulence mechanisms that are absent from S. Typhimurium have been implicated to explain the differences in the disease presentation, including expression of the typhoid toxin [3], [4], altered flagellin gene regulation [5]–[7], altered invasion gene regulation [8]–[10] and expression of the virulence-associated (Vi) capsular polysaccharide [11]–[17]. In addition, S. Typhimurium genes that are absent from S. Typhi can contribute to differences in the outcome of host microbe interaction. These include the gtgE gene, encoding a type III secreted effector protein [18] and the fepE gene, which encodes a regulator of O-antigen chain length [19].
One of the host factors important for limiting dissemination of S. Typhimurium in humans appears to be neutrophils. The idea that neutrophils help prevent dissemination is supported by the finding that neutropenic individuals have an increased risk of developing invasive bloodstream infections with non-typhoidal Salmonella serovars (NTS) [20], [21]. Furthermore, severe malarial hemolysis impairs resistance to S. Typhimurium infection by impairing the neutrophil oxidative burst in a mouse model [22], which might explain the increased susceptibility of patients with severe pediatric malaria to develop bloodstream infections with NTS, most commonly S. Typhimurium [23]. Thus, while neutrophils help to limit dissemination of S. Typhimurium into the bloodstream in immunocompetent individuals, S. Typhi breaches this barrier to cause typhoid fever, suggesting that both pathogens differ in their ability to evade neutrophil-dependent host defense mechanism.
To eliminate a threat, neutrophils must migrate toward the microbial intruder and perform phagocytosis. Opsonophagocytosis is initiated when bacterial surfaces activate complement through the alternative pathway (reviewed in [24]). The complement component 3 (C3) cleavage product C3b is generated continuously at a low level. In the presence of bacteria, an internal thioester group of C3b reacts with a hydroxyl group on the bacterial surface to form an ester bond [25], [26]. The covalently surface bound C3b binds factor B to form C3bB, which is cleaved to yield the C3 convertase C3bBb (Fig. S1). In turn, surface bound C3 convertase promotes accelerated cleavage of C3 into C3a and C3b, thereby promoting opsonization by amplifying C3b deposition through ester bond formation with hydroxyl groups present on the bacterial surface. Finally, fragmentation of C3b leads to formation of C3bi, which is bound by complement receptor (CR)3, a phagocytic receptor expressed on the surface of neutrophils (reviewed in [27]). While it is well established that S. Typhi can overcome opsonophagocytosis [12], [16], it is not known whether S. Typhi and S. Typhimurium differ in their ability to evade neutrophil chemotaxis. Here we used single-cell experiments [28], [29] to study the chemotactic response of neutrophils toward S. Typhi and S. Typhimurium.
Results
Chemotactic responses of human neutrophils are obstructed by S. Typhi, but not by S. Typhimurium or E. coli Nissle 1917
To study bacterial-guided neutrophil chemotaxis, an initially quiescent human neutrophil was picked up at the tip of a micropipette. Bacteria were then immobilized by a laser optical-trap (laser tweezers) and brought stepwise into close proximity of the neutrophil in the presence of human serum (Fig. S2A). We investigated three organisms that interact differently with the human intestinal mucosa: a commensal organism residing in the intestinal lumen (Escherichia coli strain Nissle 1917), an invasive enteric pathogen associated with a localized gastroenteritis (S. Typhimurium) and the causative agent of an invasive disseminated infection termed typhoid fever (S. Typhi). Bringing E. coli (Fig. 1A, Video S1) or S. Typhimurium (Fig. 1B, S2B, Video S2) into a certain distance of a human neutrophil induced a vigorous chemotactic response, characterized by formation of a cellular pseudopod, which protruded toward the bacteria and responded quickly to their relocation. In striking contrast, S. Typhi did not elicit any chemotactic response by human neutrophils (Fig. 1C, S2C, Video S3).
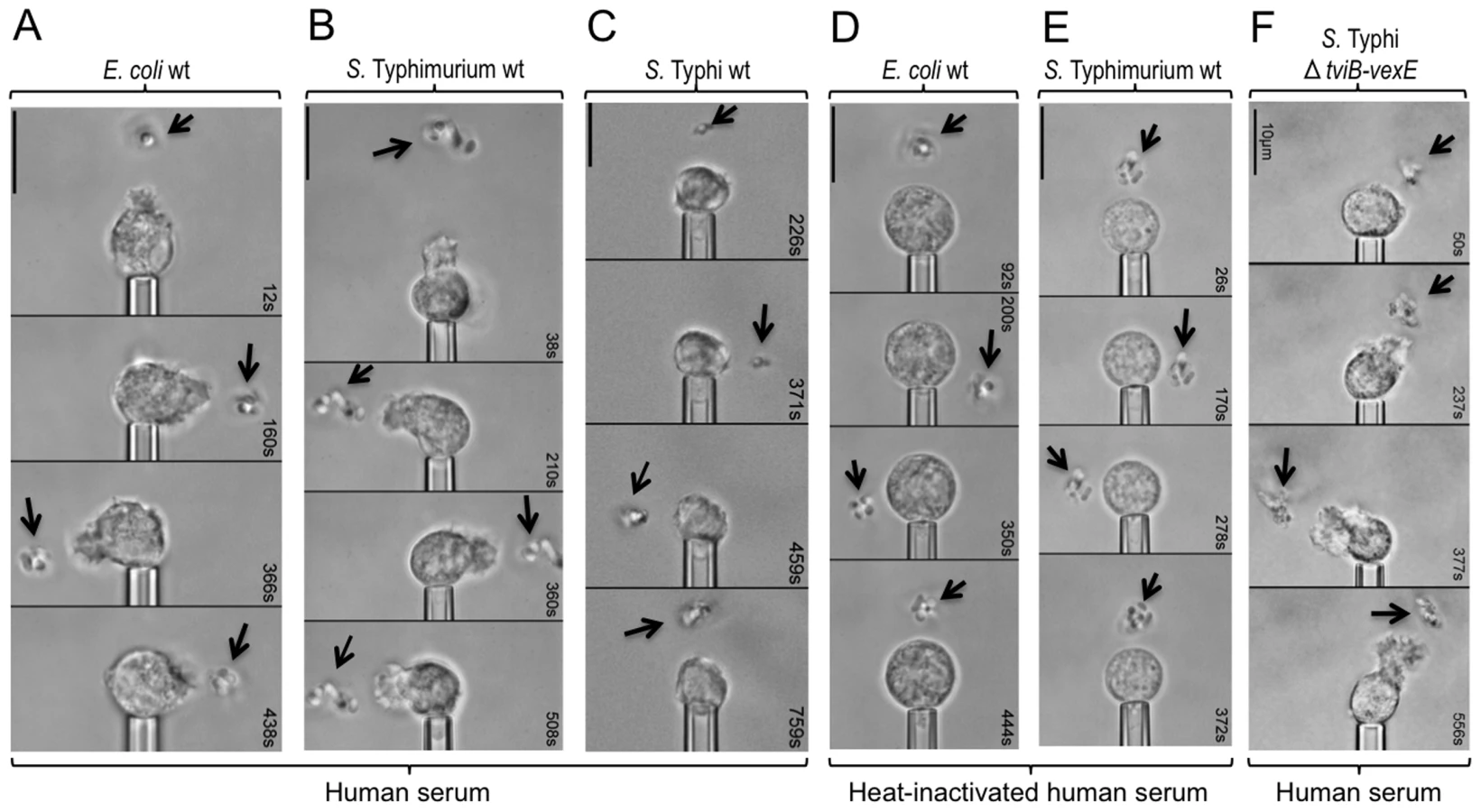
Bacterial-guided chemotactic responses of human neutrophils are complement-dependent
We reasoned that the chemotactic response toward E. coli or S. Typhimurium could be due to the release of N-formyl peptides from the bacterial surface, which are known neutrophil chemoattractants [30]. Alternatively, bacterial surface carbohydrates might activate complement by the alternative pathway, thereby giving rise to complement fragment C5a, a potent neutrophil chemoattractant [31]. Only the latter mechanism depends on the presence of serum proteins, the activity of which can be abrogated by heat inactivation of serum. Specifically, one protein in the alternative pathway of complement activation that is heat-sensitive is factor B (reviewed in [32]). As a result, heat inactivation of serum prevents the generation of C3 convertase (i.e. surface bound C3bBb), which promotes cleavage of C3 into C3a and C3b (Fig. S1). A downstream consequence of preventing formation of C3 convertase is that C3b can no longer bind surface bound C3bBb to form C5 convertase (i.e. surface bound C3bBbC3b), which promotes cleavage of C5 into C5a and C5b. Through this chain of events, heat inactivation of serum abrogates the formation of C5a. Interestingly, in the presence of heat-inactivated human serum, neither E. coli (Fig. 1D, Video S4) nor S. Typhimurium (Fig. 1E, Video S5) elicited a chemotactic response from human neutrophils in single-cell experiments.
To further investigate the mechanism of bacterial-guided neutrophil chemotaxis we used a standard Boyden chamber assay [33]. In this assay, the migration of human neutrophils from the upper compartment of the Boyden chamber [33] into a bottom reservoir that contains human serum is measured in the presence or absence of chemoattractants. The presence in the bottom chamber of either S. Typhimurium, IL-8 or synthetic N-formyl peptide (N-formyl-L-methionyl-L-leucyl-L-phenylalanine, fMLP) elicited migration of significantly (P<0.01) larger numbers of human neutrophils into the bottom reservoir than the vehicle control (Fig. 2A). To investigate the contribution of complement we used Futhan, which specifically binds factor Bb in C3 convertase (i.e. surface bound C3bBb) and C5 convertase (i.e. surface bound C3bBbC3b), thereby inhibiting the production of C3a and C5a (Fig. S1) [34], [35]. Addition of the complement inhibitor Futhan blunted the S. Typhimurium-induced neutrophil migration into the bottom chamber (P<0.01) (Fig. 2A). In contrast, migration of human neutrophils elicited by IL-8 or N-formyl peptide was not significantly inhibited by the addition of Futhan (P>0.05). Collectively, these data suggested that neutrophil chemotaxis elicited by S. Typhimurium was mediated by a complement-dependent mechanism, which was distinct from the complement-independent mechanisms of neutrophil chemotaxis elicited by IL-8 or N-formyl peptide.

The Vi capsular polysaccharide of S. Typhi impairs complement-dependent chemotactic responses of neutrophils in vitro
We next wanted to investigate the mechanism by which S. Typhi obstructs neutrophil chemotaxis. One major difference to the closely related S. Typhimurium is that S. Typhi expresses the Vi capsular polysaccharide [36]. Expression of the Vi capsular polysaccharide in S. Typhi is encoded by the viaB locus on Salmonella Pathogenicity island 7 (SPI7), a DNA region that is absent from the S. Typhimurium genome [37]. The viaB locus contains genes for the regulation (tviA), the biosynthesis (tviBCDE) and the export (vexABCDE) of the Vi capsular polysaccharide (Fig. 2B) [38]. To investigate a possible role of the Vi capsular polysaccharide in evading neutrophil chemotaxis we used single-cell experiments with human neutrophils (Fig. S2A) to compare chemotactic responses elicited by the capsulated S. Typhi wild type (Ty2) with those elicited by an isogenic strain lacking capsule expression due to deletion of the capsule biosynthesis and export genes (ΔtviB-vexE mutant) (Fig. 2C) [5]. In striking contrast to the capsulated S. Typhi wild-type, which did not elicit neutrophil chemotaxis (Fig. 1C, S2C, Video S3), human neutrophils readily extended chemotactic pseudopodia towards the non-capsulated S. Typhi ΔtviB-vexE mutant (Fig. 1F, S2C, Video S6).
To further investigate whether expression of the Vi capsular polysaccharide inhibits neutrophil chemotaxis, we monitored migration of human neutrophils from the upper compartment of a Boyden chamber [33] into a bottom reservoir that contained human serum inoculated with vehicle control or different bacterial strains. The presence of the non-capsulated S. Typhi ΔtviB-vexE mutant in the bottom chamber elicited migration of significantly (P<0.05) larger numbers of human neutrophils than the presence of the capsulated S. Typhi wild-type strain (Fig. 2D). Furthermore, neutrophil migration towards bacteria was impaired in the presence of the complement inhibitor Futhan. Collectively, these data suggested that S. Typhi obstructs complement-dependent chemotactic responses of human neutrophils in vitro by expressing the Vi capsular polysaccharide. We next investigated whether the Vi-capsular polysaccharide also obstructs chemotactic responses of murine neutrophils. While the capsulated S. Typhi wild type did not elicit chemotactic responses from murine neutrophils (Fig. S3A, Video S7), murine neutrophils extended chemotactic pseudopodia towards the non-capsulated S. Typhi ΔtviB-vexE mutant (Fig. S3B, Video S8).
Expression of the Vi capsular polysaccharide reduces complement activation by the alternative pathway [12], [16], as indicated by diminished fixation of complement fragment C3b on the surface of the capsulated S. Typhi wild-type strain compared to a non-capsulated S. Typhi mutant (ΔtviB-vexE mutant) (Fig. 2E and 2F). To further investigate whether chemotactic responses were dependent on C3, we performed single-cell experiments with serum and/or neutrophils from mice with C3-deficiency [39]. In serum from C3-deficient mice (bred on a C57BL/6 background), neutrophils from C3-deficient mice did not exhibit a chemotactic response toward the non-capsulated S. Typhi ΔtviB-vexE mutant (Fig. 3A, Video S9). In contrast, neutrophils from C3-deficient mice extended chemotactic pseudopodia toward the non-capsulated S. Typhi ΔtviB-vexE mutant when the experiment was performed in serum from wild type (C57BL/6) mice, which contains functional C3 protein (Fig. 3B, Video S10). As expected, chemotactic responses were also observed when both serum and neutrophils were derived from wild type (C57BL/6) mice (Fig. 3C, Video S11).

We next investigated chemotactic responses of neutrophils from C5aR-deficient mice (bred on a BALB/c background), which exhibit a specific defect in complement-dependent chemotaxis [40]. C5aR (also known as CD88) induces chemotactic responses by binding the serum component C5a, a soluble cleavage product of C5, which is generated during activation of the complement cascade (Fig. S1) [31]. Unlike neutrophils from wild-type mice (BALB/c) (Fig. 3D, Video S12), neutrophils from C5aR-deficient mice did not exhibit any chemotactic responses toward the non-capsulated S. Typhi ΔtviB-vexE mutant (Fig. 3E, Video S13). Collectively, these results showed that similar to what was observed with human neutrophils (Fig. 2D), chemotactic responses of murine neutrophils toward the non-capsulated S. Typhi ΔtviB-vexE mutant were complement-dependent.
To directly investigate the contribution of C5a to neutrophil chemotaxis, we monitored migration of human neutrophils from the upper compartment of a Boyden chamber into a bottom reservoir that either contained vehicle control or a neutralizing mouse anti-human C5a antibody. The presence of S. Typhimurium in the bottom chamber elicited neutrophil migration, which was significantly (P<0.01) reduced in the presence of anti-C5a antibody (Fig. 3F). In single cell experiments, anti-C5a antibody markedly blunted the chemotactic response of human neutrophils towards S. Typhimurium (Fig. S4). Furthermore, expression of the Vi capsular antigen reduced (P<0.05) generation of C5a during incubation of S. Typhi in human serum (Fig. 3G). Collectively, the outcome of these in vitro experiments suggested that the chemotactic responses of neutrophils towards bacteria were C5a-dependent.
Neutrophil chemotaxis is required for bacterial uptake in vivo
Since in vitro experiments with murine neutrophils recapitulated our results obtained with human neutrophils, we proceeded to investigate whether the Vi capsular polysaccharide could inhibit neutrophil chemotaxis in vivo using a mouse model. S. Typhi is a human restricted pathogen and is rapidly cleared from organs of mice [16]. However, since chemotactic chasing by neutrophils is an early event that precedes clearance of bacteria, we reasoned that this aspect of host microbe interaction could be studied in mice at early time points after infection. The capsulated S. Typhi wild-type strain and the non-capsulated S. Typhi ΔtviB-vexE mutant were transformed with a plasmid encoding green fluorescence protein (GFP) and injected intraperitoneally into mice (BALB/c). Both GFP-labeled bacterial strains were recovered in similar numbers from the peritoneal cavity one hour after infection (Fig. S5A). Neutrophils present in the peritoneal lavage one hour after infection were analyzed by flow cytometry (Fig. S6). Both GFP-labeled bacterial strains elicited the influx of similar numbers of neutrophils into the peritoneal cavity (Fig. S5B and S5C). However, a significantly larger fraction of neutrophils were associated with the non-capsulated S. Typhi ΔtviB-vexE mutant than with the capsulated S. Typhi wild-type strain (Fig. 3H and 3I). These data suggested that expression of the Vi capsular polysaccharide impaired the ability of neutrophils to take up bacteria in vivo.
To determine whether these differences were due to a capsule-mediated inhibition of neutrophil chemotaxis, the experiment was repeated in C5aR-deficient mice, which are defective for complement-dependent chemotactic responses (Fig. 3E) [40]. When C5aR-deficient mice were infected with the capsulated S. Typhi wild-type strain or the non-capsulated S. Typhi ΔtviB-vexE mutant, there was a marked overall reduction in the fraction of neutrophils associated with GFP-labeled bacteria. Furthermore, no differences were observed in C5aR-deficient mice between the fractions of neutrophils associated with capsulated or non-capsulated bacterial strains (Fig. 3H and 3I), indicating that in the absence of complement-dependent chemotaxis, the Vi capsular polysaccharide had no detectable influence on bacterial association with neutrophils early after infection. Collectively, these data suggested neutrophils engulf bacteria in the peritoneal cavity through a C5Ra-dependent mechanism that can be obstructed by S. Typhi through expression of the Vi capsular polysaccharide.
Discussion
Neutrophil migration into an infected tissue is initially guided by chemoattractants, such as interleukin (IL)-8, a chemokine produced by host cells after they detect the presence of bacteria using pathogen recognition receptors (PRRs). Neutrophils that follow a gradient of IL-8 to enter a site of infection will ultimately reach host cells producing this chemokine. In Boyden chamber assays, IL-8 can contribute to host cell migration, because this chemokine can be released from neutrophils upon contact with bacteria [41]. However, early microscopic observations suggest that chemotactic chasing of Staphylococcus aureus by neutrophils is bacterial-guided [42]. Investigation of bacterial factors possibly involved in this process suggests that purified C5a and N-formyl-l-methionyl-l-phenylalanine both can cause a change in neutrophil shape from a spherical to a polarized configuration [43], [44]. Some studies on the mechanism that triggers chemotaxis towards intact bacterial cells suggest a contribution of C5a [45], [46], while others suggest that a release of N-formyl peptides from the surface of intact bacterial cells is involved [41], [47]–[51]. Thus, it is not clear from the literature whether C5a emanating from the microbial surface, the release of N-formyl peptides from the bacterial cell or a combination of both mechanisms account for Salmonella-guided neutrophil chemotaxis. Here we addressed this question using microscopic observations of single cell experiments. Our results show that the formation of chemotactic pseudopodia towards intact cells of S. Typhimurium or non-capsulated S. Typhi was solely dependent on complement.
The question of whether either complement or N-formyl peptides are responsible for Salmonella-guided neutrophil chemotaxis is relevant because expression of the Vi capsular polysaccharide prevents complement deposition on the bacterial surface [12], [16], but is not known to inhibit detection of N-formyl peptides by neutrophils. The Vi capsular polysaccharide is a homopolymer composed of α-1,4 (2-deoxy)-2-N-acetyl-3-O-acetylgalacturonic acid, which is devoid of free hydroxyl-groups that would be available for C3b deposition (Fig. S1) [52]. In other words, by covering its surface with Vi capsular polysaccharide, S. Typhi inhibits activation of complement through the alternative pathway by preventing covalent binding of C3b to the bacterial cell. The fepE gene, which is required for expression of very long O-antigen chains containing an estimated 100 copies of a oligosaccharide repeat unit, is interrupted in the S. Typhi genome by a stop codon. Restoration of the fepE gene in S. Typhi results in expression of very-long O-antigen chains, which in turn results in enhanced C3 fixation [19], indicating that capsular material must extend beyond the O-antigen chains to effectively inhibit complement activation. Thus, inhibition of complement activation is not a property inherent to capsules, but might be a function of their length and carbohydrate composition. For example, the capsular polysaccharide of Cryptococcus neoformans prevents neither complement activation nor neutrophil chemotaxis [53]. In contrast, the Vi capsular polysaccharide of S. Typhi inhibits complement-dependent processes, such as opsonophagocytosis [12], [16]. Here we show that inhibition of complement activation by the Vi capsular polysaccharide also abrogated bacterial-guided neutrophil chemotaxis, because this process was solely complement dependent. Our supplementary videos illustrate that the Vi capsular polysaccharide can act as a “cloaking device” that makes S. Typhi practically “invisible” to neutrophils. By using an animal model we were able to show that C5aR-dependent chemotaxis is essential for a rapid association of bacteria with neutrophils. Obstruction of C5aR-dependent chemotaxis by the Vi capsular polysaccharide thus aids in evading neutrophil-dependent host defense mechanisms.
Our results suggest that one of the differences between gastroenteritis and typhoid fever is that the pathogen causing the latter disease evades neutrophil chemotaxis. A correlation between blocked neutrophil chemotaxis and increased vulnerability to bacterial dissemination is also observed during gonorrhea, where delayed stimulation of complement-dependent neutrophil chemotaxis in vitro is associated with dissemination of Neisseria gonorrhoeae infection [46]. Furthermore, C5aR mediates mucosal defense in a mouse model of Pseudomonas aeruginosa lung infection [40]. The inhibition of neutrophil chemotaxis reported here likely cooperates with other functions ascribed to the Vi capsular polysaccharide, including an obstruction of osponophagocytosis [12], [16] and a blunting of the respiratory burst of neutrophils [11], to help S. Typhi overcome neutrophil-dependent mucosal barrier functions. However, the increased ability of S. Typhi to cause bacteremia in immunocompetent individuals likely involves additional Vi capsular polysaccharide-independent mechanisms, such as changes in flagella gene expression, which have been implicated in increasing bacterial dissemination in animal models [6], [7].
Materials and Methods
Ethics statement
All animal experiments were performed according to USDA guidelines and approved by the Institutional Animal Care and Use Committee at the University of California at Davis. The University of California at Davis Institutional Review Board approved the protocol for obtaining blood draws for this study and written informed consent was obtained from all individuals.
Bacterial strains and culture conditions
The S. Typhi wild-type isolate Ty2 (ATCC 19430) was obtained from the American Type Culture Collection. A derivative of Ty2 carrying a ΔtviB-vexE deletion (SW74) has been described previously [5]. To label bacteria with GFP, S. Typhi strains were transformed with plasmid pDW5 [54]. All bacterial strains were maintained in −80°C freezer stocks and were streaked on Luria-Bertani (LB) agar plates (15 g/l agar, 10 g/l tryptone, 5 g/l yeast extract, 10 g/l NaCl). To induce optimal expression of Vi capsular polysaccharide, strains were grown overnight in modified LB broth (10 g/l tryptone, 5 g/l yeast extract) with shaking at 37°C.
Animal experiments
Six to eight week old female mice obtained from The Jackson Laboratory were used for this study.
Mice were injected intraperitoneally with 1×107 colony forming units (CFU)/animal suspended in 0.1 ml phosphate buffered saline (PBS). Mice were euthanized and peritoneal lavages were collected an hour post injection by flushing the peritoneal cavity with 5 ml PBS.
To collect murine neutrophils for single-cell chemotaxis experiments, mice were euthanized and peripheral blood was collected by cardiac puncture. Neutrophils were then isolated using a neutrophil enrichment kit (Stemcell Technologies, Vancouver, Canada) using instructions provided by the manufacturer.
Single-cell experiments
Neutrophils were suspended in chemotaxis medium (1.5×106 cells/ml in Hank's Balanced Salt Solution with 10% autologous serum, unless mentioned otherwise). In some experiments, PBS (vehicle control) or mouse monoclonal anti-human C5a antibody (ab11876, Abcam) dissolved in PBS was added at a final concentration of 10 µg/ml. To generate bacterial micro-agglutinates, 109 CFU/ml were suspended in PBS containing 0.02% sodium azide. Rabbit anti-Vi serum (for Ty2) or rabbit anti-O9 serum (for ΔtviB-vexE mutant) were added to the bacterial suspension at a 1∶1 ratio to generate micro-agglutinates. The micropipette setup has been described previously [29]. Briefly, micropipettes with inner diameter of 2–3 µm were mounted on a motorized 3-axis manipulator that was controlled by a custom-written software. Neutrophils and bacteria were dispensed into a microscopic chamber (Figure S1). Neutrophils and bacteria were lifted above the chamber bottom by aspiring them on a micropipette or by using optical tweezers. The micropipette-held bacterial micro-agglutinate or bacteria immobilized by optical tweezers were gradually brought towards the micropipette-held neutrophil and the chemotactic response was recorded. Each experiments was repeated with cells from at least 4 different donors.
Boyden chamber assay
Human neutrophils were isolated from peripheral blood of healthy adult donors using Cytoselect Cell Migration Assay kit (Cellbiolabs Inc., San Diego, CA). Neutrophils were suspended in HBSS at the concentration of approximately 1.5×106 cells/ml and 300 µl were loaded into the top chamber of a 24-well Boyden chamber plate (Cell Biolabs inc., San Diego, CA). Sterile PBS or a bacterial suspension (1.5×107 CFU/ml) in chemotaxis medium was added to the bottom chamber. In some experiments, the complement inhibitor Futhan (6-amidino-2-naphthyl p-guanidinobenzoate dimethanesulfonate) [34] was added to the bottom chamber at a final concentration of 50 µg/ml. Human IL-8 (100 ng/ml, R&D Systems) or fMLP 10 nM (N-Formyl-L-methionyl-L-leucyl-L-phenylalanine, 10 nM) (Sigma, St. Louis, MO) were also added to the bottom chamber in some experiments. In some experiments, PBS (vehicle control) or mouse monoclonal anti-human C5a antibody (ab11876, Abcam) dissolved in PBS was added to the bottom chamber at a final concentration of 10 µg/ml. The plates were incubated for two hours at 37°C in tissue culture incubator. The cells that migrated to the bottom chamber were counted using a hemocytometer. Each experiments was repeated with cells from at least 3 different donors.
Flow cytometry
The peritoneal lavage containing approximately 106 cells per animal was suspended in 2 ml cold phosphate-buffered saline and stained with Aqua Live/Dead cell discriminator (Invitrogen catalog no. L34597) according to the manufacturer's instructions. Cells were then washed and resuspended in fluorescence-activated cell sorting (FACS) buffer (PBS containing 1% bovine serum albumin and 1 mM EDTA) and incubated for 15 min in blocking antibody anti-CD16/32 (eBioscience clone 93). Cells were then stained for 20 min in the dark at 4°C with optimized concentrations of anti-CD3 PE (eBioscience clone 17A2), anti-B220 PE (eBioscience clone RA3-6B2), anti-NK1.1 PE (eBioscience clone PK136), anti-Ly6C Pacific Blue (eBioscience clone HK1.4), anti-Ly6G PerCPCy5.5 (BD Pharmingen clone 1A8) and anti-CD11b APC Cy7 (Biolegend clone M1/70). Cells were washed twice with FACS buffer and subsequently fixed in 4% paraformaldehyde for 1 hour. Cells were then washed twice and resuspended in FACS buffer and analyzed using an LSR II flow cytometer (Becton Dickinson, San Jose, CA). GFP expressing bacteria were detected using 488 nm excitation. The data were analyzed by using FlowJo software (Treestar, Inc., Ashland, OR). Gates were set on singlets and then on live cells. Subsequent gates were based on Fluorescence-Minus-One and unstained controls. To confirm neutrophil morphology, LY6G+ LY6C+ cells were sorted using a MoFlo high-speed cell sorter (Dako). Cytospin samples were prepared from the sorted cells using a Shandon cytocentrifuge (Thermo Scientific, MI) and dried prior to staining with a Diff-Quik stain kit (IMEB, Inc., San Marcos, CA).
Vi capsule expression and C3b deposition was detected using flow cytometry as described previously [16], [55]. Bacterial cells were detected using the DNA specific stain propidium iodide. For detection of the Vi capsular polysaccharide, bacterial cells incubated in rabbit anti-Vi serum (Difco) were washed and stained with goat anti-rabbit FITC-conjugate (Jackson ImmunoLabs). To detect C3b deposition, bacterial cells were incubated for 30 minutes in 1% human serum. Cells were then washed and stained with FITC-conjugated goat anti-human C3b antibody (MP Biomedicals). For each sample, 50 000 events (bacterial cells) were collected using LSR II (Becton Dickinson). Data were expressed as percentage values of positive counts per total counts.
Enzyme linked immune-sorbent assay (ELISA)
For the human serum ELISA assays, S. Typhi strains were grown statically overnight at 37°C in SOB+Mg medium (20 g/liter tryptone, 5 g/liter yeast extract, 10 mM NaCl, 10 mM KCl, 10 mM MgCl2, 10 mM MgSO4) to induce expression of the Vi capsular polysaccharide. Then 1×107 CFU of S. Typhi were incubated for 15 minutes at 37°C in 10% human serum complement (Quidel) diluted in PBS. The generation of C5a was detected with the Human C5a ELISA Kit II (BD; cat #557965) using the instructions provided by the manufacturer.
Statistical analysis
To determine statistical significance between treatment groups in the animal experiments an unpaired Student t-test was used. A P value of less than 0.05 was considered to be significant.
Supporting Information
Zdroje
1. Nuccio S-P, Wangdi T, Winter SE, Baumler AJ (2013) Typhoid. In: Rosenberg E, DeLong EF, Lory S, Stackebrandt E, Thompson F, editors. The Prokaryotes. 4th ed. Berlin Heidelberg: Springer-Verlag. pp. 353–374.
2. ZhangS, KingsleyRA, SantosRL, Andrews-PolymenisH, RaffatelluM, et al. (2003) Molecular pathogenesis of Salmonella enterica serotype typhimurium - induced diarrhea. Infect Immun 71 : 1–12.
3. SpanoS, UgaldeJE, GalanJE (2008) Delivery of a Salmonella Typhi exotoxin from a host intracellular compartment. Cell Host Microbe 3 : 30–38.
4. SongJ, GaoX, GalanJE (2013) Structure and function of the Salmonella Typhi chimaeric A(2)B(5) typhoid toxin. Nature 499 : 350–354.
5. WinterSE, RaffatelluM, WilsonRP, RussmannH, BaumlerAJ (2008) The Salmonella enterica serotype Typhi regulator TviA reduces interleukin-8 production in intestinal epithelial cells by repressing flagellin secretion. Cell Microbiol 10 : 247–261.
6. WinterSE, WinterMG, GodinezI, YangH-J, RussmannH, et al. (2010) A Rapid Change in Virulence Gene Expression during the Transition from the Intestinal Lumen into Tissue Promotes Systemic Dissemination of Salmonella. PLoS Pathog 6: e1001060.
7. AtifSM, WinterSE, WinterMG, McSorleySJ, BaumlerAJ (2014) Salmonella enterica serovar Typhi impairs CD4 T cell responses by reducing antigen availability. Infect Immun 82 : 2247–2254.
8. TarteraC, MetcalfES (1993) Osmolarity and growth phase overlap in regulation of Salmonella typhi adherence to and invasion of human intestinal cells. Infect Immun 61 : 3084–3089.
9. LeclercGJ, TarteraC, MetcalfES (1998) Environmental regulation of Salmonella typhi invasion-defective mutants. Infect Immun 66 : 682–691.
10. WinterSE, WinterMG, ThiennimitrP, GerrietsVA, NuccioSP, et al. (2009) The TviA auxiliary protein renders the Salmonella enterica serotype Typhi RcsB regulon responsive to changes in osmolarity. Mol Microbiol 74 : 175–193.
11. MillerRM, GarbusJ, HornickRB (1972) Lack of enhanced oxygen consumption by polymorphonuclear leukocytes on phagocytosis of virulent Salmonella typhi. Science 175 : 1010–1011.
12. LooneyRJ, SteigbigelRT (1986) Role of the Vi antigen of Salmonella typhi in resistance to host defense in vitro. J Lab Clin Med 108 : 506–516.
13. HiroseK, EzakiT, MiyakeM, LiT, KhanAQ, et al. (1997) Survival of Vi-capsulated and Vi-deleted Salmonella typhi strains in cultured macrophage expressing different levels of CD14 antigen. FEMS Microbiol Lett 147 : 259–265.
14. WilsonRP, RaffatelluM, ChessaD, WinterSE, TukelC, et al. (2008) The Vi-capsule prevents Toll-like receptor 4 recognition of Salmonella. Cell Microbiol 10 : 876–890.
15. HanedaT, WinterSE, ButlerBP, WilsonRP, TukelC, et al. (2009) The capsule-encoding viaB locus reduces intestinal inflammation by a Salmonella pathogenicity island 1-independent mechanism. Infect Immun 77 : 2932–2942.
16. WilsonRP, WinterSE, SpeesAM, WinterMG, NishimoriJH, et al. (2011) The Vi capsular polysaccharide prevents complement receptor 3-mediated clearance of Salmonella enterica serotype Typhi. Infect Immun 79 : 830–837.
17. JansenAM, HallLJ, ClareS, GouldingD, HoltKE, et al. (2011) A Salmonella Typhimurium-Typhi Genomic Chimera: A Model to Study Vi Polysaccharide Capsule Function In Vivo. PLoS Pathog 7: e1002131.
18. SpanoS, LiuX, GalanJE (2011) Proteolytic targeting of Rab29 by an effector protein distinguishes the intracellular compartments of human-adapted and broad-host Salmonella. Proc Natl Acad Sci U S A 108 : 18418–18423.
19. CrawfordRW, WangdiT, SpeesAM, XavierMN, TsolisRM, et al. (2013) Loss of very-long O-antigen chains optimizes capsule-mediated immune evasion by Salmonella enterica serovar Typhi. MBio 4: pii: e00232-13.
20. NoriegaLM, Van der AuweraP, DaneauD, MeunierF, AounM (1994) Salmonella infections in a cancer center. Support Care Cancer 2 : 116–122.
21. TumbarelloM, TacconelliE, CaponeraS, CaudaR, OrtonaL (1995) The impact of bacteraemia on HIV infection. Nine years experience in a large Italian university hospital. J Infect 31 : 123–131.
22. CunningtonAJ, NjieM, CorreaS, TakemEN, RileyEM, et al. (2012) Prolonged neutrophil dysfunction after Plasmodium falciparum malaria is related to hemolysis and heme oxygenase-1 induction. J Immunol 189 : 5336–5346.
23. ReddyEA, ShawAV, CrumpJA (2010) Community-acquired bloodstream infections in Africa: a systematic review and meta-analysis. Lancet Infect Dis 10 : 417–432.
24. GotzeO, Muller-EberhardHJ (1971) The c3-activator system: an alternate pathway of complement activation. J Exp Med 134 : 90–108.
25. LawSK, LevineRP (1977) Interaction between the third complement protein and cell surface macromolecules. Proc Natl Acad Sci U S A 74 : 2701–2705.
26. TackBF, HarrisonRA, JanatovaJ, ThomasML, PrahlJW (1980) Evidence for presence of an internal thiolester bond in third component of human complement. Proc Natl Acad Sci U S A 77 : 5764–5768.
27. HorwitzMA (1982) Phagocytosis of microorganisms. Rev Infect Dis 4 : 104–123.
28. HeinrichV, LeeCY (2011) Blurred line between chemotactic chase and phagocytic consumption: an immunophysical single-cell perspective. J Cell Sci 124 : 3041–3051.
29. MankovichAR, LeeCY, HeinrichV (2013) Differential effects of serum heat treatment on chemotaxis and phagocytosis by human neutrophils. PLoS One 8: e54735.
30. SchiffmannE, CorcoranBA, WahlSM (1975) N-formylmethionyl peptides as chemoattractants for leucocytes. Proc Natl Acad Sci U S A 72 : 1059–1062.
31. MantheyHD, WoodruffTM, TaylorSM, MonkPN (2009) Complement component 5a (C5a). Int J Biochem Cell Biol 41 : 2114–2117.
32. Muller-EberhardHJ (1975) Complement. Annu Rev Biochem 44 : 697–724.
33. BoydenS (1962) The chemotactic effect of mixtures of antibody and antigen on polymorphonuclear leucocytes. J Exp Med 115 : 453–466.
34. IkariN, SakaiY, HitomiY, FujiiS (1983) New synthetic inhibitor to the alternative complement pathway. Immunology 49 : 685–691.
35. InagiR, MiyataT, MaedaK, SugiyamaS, MiyamaA, et al. (1991) FUT-175 as a potent inhibitor of C5/C3 convertase activity for production of C5a and C3a. Immunol Lett 27 : 49–52.
36. FelixA, PittRM (1934) A new antigen of B. typhosus. Lancet 227 : 186–191.
37. ParkhillJ, DouganG, JamesKD, ThomsonNR, PickardD, et al. (2001) Complete genome sequence of a multiple drug resistant Salmonella enterica serovar Typhi CT18. Nature 413 : 848–852.
38. VirlogeuxI, WaxinH, EcobichonC, PopoffMY (1995) Role of the viaB locus in synthesis, transport and expression of Salmonella typhi Vi antigen. Microbiology 141 (Pt 12) 3039–3047.
39. FischerMB, MaM, GoergS, ZhouX, XiaJ, et al. (1996) Regulation of the B cell response to T-dependent antigens by classical pathway complement. J Immunol 157 : 549–556.
40. HopkenUE, LuB, GerardNP, GerardC (1996) The C5a chemoattractant receptor mediates mucosal defence to infection. Nature 383 : 86–89.
41. RussoTA, DavidsonBA, TopolnyckyDM, OlsonR, MorrillSA, et al. (2003) Human neutrophil chemotaxis is modulated by capsule and O antigen from an extraintestinal pathogenic Escherichia coli strain. Infect Immun 71 : 6435–6445.
42. MellyMA, ThomisonJB, RogersDE (1960) Fate of staphylococci within human leukocytes. J Exp Med 112 : 1121–1130.
43. SmithCW, HollersJC, PatrickRA, HassettC (1979) Motility and adhesiveness in human neutrophils. Effects of chemotactic factors. J Clin Invest 63 : 221–229.
44. ZhelevDV, AlteraifiAM, ChodniewiczD (2004) Controlled pseudopod extension of human neutrophils stimulated with different chemoattractants. Biophys J 87 : 688–695.
45. MusherDM, Hague-ParkM, GyorkeyF, AndersonDC, BaughnRE (1983) The interaction between Treponema pallidum and human polymorphonuclear leukocytes. J Infect Dis 147 : 77–86.
46. DensenP, MacKeenLA, ClarkRA (1982) Dissemination of gonococcal infection is associated with delayed stimulation of complement-dependent neutrophil chemotaxis in vitro. Infect Immun 38 : 563–572.
47. SordelliDO, DjafariM, GarciaVE, FontanPA, DoringG (1992) Age-dependent pulmonary clearance of Pseudomonas aeruginosa in a mouse model: diminished migration of polymorphonuclear leukocytes to N-formyl-methionyl-leucyl-phenylalanine. Infect Immun 60 : 1724–1727.
48. IjiriY, MatsumotoK, KamataR, NishinoN, OkamuraR, et al. (1994) Suppression of polymorphonuclear leucocyte chemotaxis by Pseudomonas aeruginosa elastase in vitro: a study of the mechanisms and the correlation with ring abscess in pseudomonal keratitis. Int J Exp Pathol 75 : 441–451.
49. LiY, LoikeJD, EmberJA, ClearyPP, LuE, et al. (2002) The bacterial peptide N-formyl-Met-Leu-Phe inhibits killing of Staphylococcus epidermidis by human neutrophils in fibrin gels. J Immunol 168 : 816–824.
50. SouthgateEL, HeRL, GaoJL, MurphyPM, NanamoriM, et al. (2008) Identification of formyl peptides from Listeria monocytogenes and Staphylococcus aureus as potent chemoattractants for mouse neutrophils. J Immunol 181 : 1429–1437.
51. MurphyH, CoganT, HumphreyT (2011) Direction of neutrophil movements by Campylobacter-infected intestinal epithelium. Microbes Infect 13 : 42–48.
52. HeynsK, KiesslingG (1967) Strukturaufklarung des Vi-antigens aus Citrobacter freundii (E. coli) 5396/38. Carbohydrate Res 3 : 340–353.
53. LaxaltKA, KozelTR (1979) Chemotaxigenesis and activation of the alternative complement pathway by encapsulated and non-encapsulated Cryptococcus neoformans. Infect Immun 26 : 435–440.
54. CummingsLA, WilkersonWD, BergsbakenT, CooksonBT (2006) In vivo, fliC expression by Salmonella enterica serovar Typhimurium is heterogeneous, regulated by ClpX, and anatomically restricted. Mol Microbiol 61 : 795–809.
55. RaffatelluM, SantosRL, ChessaD, WilsonRP, WinterSE, et al. (2007) The capsule encoding the viaB locus reduces interleukin-17 expression and mucosal innate responses in the bovine intestinal mucosa during infection with Salmonella enterica serotype Typhi. Infect Immun 75 : 4342–4350.
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2014 Číslo 8
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- Disruption of Fas-Fas Ligand Signaling, Apoptosis, and Innate Immunity by Bacterial Pathogens
- Ly6C Monocyte Recruitment Is Responsible for Th2 Associated Host-Protective Macrophage Accumulation in Liver Inflammation due to Schistosomiasis
- Host Responses to Group A Streptococcus: Cell Death and Inflammation
- Pathogenicity and Epithelial Immunity
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy