
Cyclic di-GMP-dependent Signaling Pathways in the Pathogenic Firmicute
Listeria monocytogenes is ubiquitously present in the environment, highly adaptable and tolerant to various stresses. L. monocytogenes is also a foodborne pathogen associated with the largest foodborne outbreaks in recent US history. Signaling pathways involving the second messenger c-di-GMP play important roles in increased stress survival of proteobacteria and mycobacteria, yet roles of c-di-GMP signaling pathways in L. monocytogenes have remained unexplored. Here, we identified and systematically characterized functions of the proteins involved in c-di-GMP synthesis, degradation and sensing. We show that elevated c-di-GMP levels in L. monocytogenes result in synthesis of a previously unknown exopolysaccharide that promotes cell aggregation, inhibits motility in semi-solid media, and importantly, enhances bacterial tolerance to commonly used disinfectants as well as desiccation. These properties of the exopolysaccharide may increase listerial survival in food processing plants as well as on produce during transportation and storage. Elevated c-di-GMP levels also grossly diminish listerial invasiveness in enterocytes in vitro, and impair bacterial accumulation in selected mouse organs during oral infection.
Published in the journal:
. PLoS Pathog 10(8): e32767. doi:10.1371/journal.ppat.1004301
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1004301
Summary
Listeria monocytogenes is ubiquitously present in the environment, highly adaptable and tolerant to various stresses. L. monocytogenes is also a foodborne pathogen associated with the largest foodborne outbreaks in recent US history. Signaling pathways involving the second messenger c-di-GMP play important roles in increased stress survival of proteobacteria and mycobacteria, yet roles of c-di-GMP signaling pathways in L. monocytogenes have remained unexplored. Here, we identified and systematically characterized functions of the proteins involved in c-di-GMP synthesis, degradation and sensing. We show that elevated c-di-GMP levels in L. monocytogenes result in synthesis of a previously unknown exopolysaccharide that promotes cell aggregation, inhibits motility in semi-solid media, and importantly, enhances bacterial tolerance to commonly used disinfectants as well as desiccation. These properties of the exopolysaccharide may increase listerial survival in food processing plants as well as on produce during transportation and storage. Elevated c-di-GMP levels also grossly diminish listerial invasiveness in enterocytes in vitro, and impair bacterial accumulation in selected mouse organs during oral infection.
Introduction
Cyclic dimeric GMP (c-di-GMP) [1] is one of the most common bacterial second messengers. Over the last ten years our understanding of c-di-GMP-mediated signal transduction pathways has rapidly expanded (reviewed in [2]–[6]). However, this expansion has been dominated by studies of Proteobacteria, and to a lesser extent Actinobacteria and Spirochetes, while studies of c-di-GMP signaling in Firmicutes have lagged behind. In the Proteobacteria, elevated levels of intracellular c-di-GMP are associated with inhibition of motility and increased synthesis of biofilm components, e.g. exopolysaccharides (EPS), pili and/or surface adhesins. In pathogens that propagate extracellularly, elevated c-di-GMP levels have been found generally detrimental for acute infections (reviewed in [2], [5]–[7]), although individual components of c-di-GMP signaling networks may play different roles during various stages of infection [8], [9]. In contrast, during chronic infections, c-di-GMP-induced biofilms greatly increase pathogen survival in vivo [10]–[12]. In intracellular proteobacterial pathogens, c-di-GMP signaling pathways are required for full-scale virulence in those species that form biofilm-like intracellular structures [13]–[15], but appear to be detrimental, at least in some species, that do not form such structures [16].
In this study, we used the foodborne pathogen Listeria monocytogenes [17] as a model to gain insight into c-di-GMP-based regulation in Firmicutes in general. L. monocytogenes is widespread in the environment. It has been isolated from soil, silage, groundwater, sewage and vegetation and actively grows at a broad range of temperatures (from 0 to 44°C), oxygen levels, pH (from 4.4 to 9.6), and salt concentrations (up to 10% w/v NaCl), and is capable of utilizing a variety of carbohydrates as well as other organic molecules as carbon sources. Listeriosis is a relatively infrequent disease but it has the highest mortality rate, ∼20%, among foodborne diseases in the developed world. The complications of listeriosis, common in immunocompromised patients, include encephalitis, meningitis, and stillbirths or infection of the central nervous system in newborns [17]–[20].
Common sources of listerial contamination include unpasteurized milk and milk products, raw meat, and packaged cooked meat products. In plants processing meat and milk products listerial biofilms can persist for years and even decades and cause repetitive contamination of processed foods [21], [22]. In recent years, listeriosis caused by contaminated fresh produce has become a significant concern. According to The Centers for Disease Control and Prevention, the 2011 outbreak caused by Listeria-contaminated cantaloupes resulted in 33 deaths and was the largest foodborne disease outbreak in US history in almost 90 years [23], [24]. Our understanding of how listeria attach and grow on the surfaces of produce is surprisingly poor, and so is our knowledge of the mechanisms ensuring long-term listerial survival. EPS is one of the common components that facilitate bacterial attachment to plant surfaces and increases their tolerance to desiccation and disinfection [25], [26], both of which are critical parameters for food safety. However, the ability of L. monocytogenes to synthesize EPS has remained controversial [27], [28].
In Proteobacteria, EPS synthesis is commonly induced via c-di-GMP signaling pathways, yet studies of such pathways in Firmicutes are just beginning to emerge [29]–[32]. It is peculiar that distribution of c-di-GMP signaling pathways in Firmicutes is very uneven. Several major genera of pathogenic firmicutes, Staphylococci, Streptococci and Enterococci, lack these altogether. However, staphylococci retain remnants of c-di-GMP signaling enzymes, which are involved in biofilm regulation but are no longer associated with c-di-GMP [33]. On the other extreme of the spectrum are certain clostridial species, e.g. Clostridium difficile, that have numerous enzymes involved in c-di-GMP synthesis and hydrolysis [29]. A recent study by Tamayo and colleagues [30] showed that elevated levels of c-di-GMP inhibited motility and induced cell aggregation in C. difficile. The c-di-GMP-dependent riboswitches from C. difficile expressed in a heterologous host were shown to affect gene expression in a c-di-GMP-dependent manner. One riboswitch is located upstream of the C. difficile flagellar biosynthesis operon [34]; the other one is part of the riboswitch-ribozyme system predicted to control adhesin gene expression [35]. Other recent studies [31], [32] characterized enzymes involved in c-di-GMP synthesis and degradation in Bacillus subtilis and uncovered the role of c-di-GMP in regulating motility and biofilm formation in this species.
Here, we present a genome-wide view of c-di-GMP signaling in L. monocytogenes. We used bioinformatics analysis to identify genes involved in c-di-GMP synthesis, degradation and signal transduction, and subsequently applied genetic and biochemical approaches to characterize functions of these genes in EPS synthesis, motility inhibition, tolerance to disinfection and desiccation, invasiveness in mammalian cells, and virulence in a mouse model of listeriosis.
Results
Bioinformatic analysis of the c-di-GMP signaling system in Listeria
C-di-GMP is synthesized by diguanylate cyclases (DGCs), which contain GGDEF domains [36], [37], and degraded by c-di-GMP-specific phosphodiesterases (PDEs), which contain either EAL [38]–[40] or HD-GYP [41] catalytic domains. The currently sequenced strains of L. monocytogenes, and the majority of related listerial species encode four GGDEF domain proteins, three EAL domain proteins and no HD-GYP domain proteins (Pfam database [42]) (Fig. 1).

Our sequence analysis predicted that three of the four GGDEF proteins from L. monocytogenes EGD-e, Lmo1911 (DgcA), Lmo1912 (DgcB) and Lmo2174 (DgcC), contain conserved residues associated with DGC activity, and therefore they likely possess DGC activities [4], [43]. The three predicted DGCs have similar domain architectures with a GGDEF domain preceded by either six or eight transmembrane helices (Fig. 1). This domain architecture suggests that c-di-GMP synthesis is regulated by external signals or signals derived from the cell wall or cytoplasmic membrane. The three proteins share approximately 30% identity to each other over their entire lengths, and may have resulted from ancient gene duplications. The EAL domain proteins in strain EGD-e, Lmo0131 (PdeB), Lmo1914 (PdeC) and Lmo0111 (PdeD), have conserved residues required for c-di-GMP binding and hydrolysis [4], [44], and therefore were expected to possess PDE activities (Fig. 1). These putative PDEs contain only single EAL domains suggesting their cytoplasmic localization.
The dgcA and dgcB genes are codirectional and separated from each other by 20 bp, which indicates that they likely form an operon. The pdeC gene appears to belong to the same dgcA-dgcB-lmo1913-pdeC (lmo1911-1914) operon. Tiling microarray expression data support an operonal structure of this gene cluster [45]. The intervening gene, lmo1913, encodes a protein of unknown function. Based on structural predictions, Lmo1913 belongs to the six-hairpin glycosidase superfamily [46] (Fig. 1). Therefore, DgcA, DgcB and PdeC may represent a signaling module involved in c-di-GMP synthesis and degradation, and this module may be involved in controlling synthesis of an unknown EPS.
The GGDEF domain of the fourth GGDEF protein, Lmo0531, is clearly degenerate. The signature GG(D/E)EF motif in Lmo531 is 208DKDDA, which should make this protein incapable of c-di-GMP synthesis (Fig. 1). Five amino acids upstream of the signature motif is an RxxD motif that represents a part of a c-di-GMP-binding sequence known as an I-site [43], [47]. Therefore, we hypothesize that Lmo0531 acts as a c-di-GMP receptor/effector protein similar to the I-site containing degenerate GGDEF domain proteins described earlier [48], [49]. It is peculiar that Lmo0531 is the only c-di-GMP receptor that can be predicted based on genome sequence analysis [2].
To test functions of the predicted L. monocytogenes DGC and PDE proteins and a single identifiable c-di-GMP receptor, we cloned and expressed these genes in E. coli indicator strains that respond to changes in intracellular c-di-GMP concentrations in a predictable fashion, and, where necessary, purified proteins to test their activities in vitro.
L. monocytogenes PdeB-D proteins possess c-di-GMP PDE activities
We expressed L. monocytogenes pdeB, pdeC and pdeD in E. coli MG1655 ΔyhjH. This mutant lacks a major c-di-GMP PDE, YhjH [50], [51], and as a result, is impaired in motility in semi-solid agar [52]. We found that expression of any one of the pde genes was sufficient to partially restore swim zones of MG1655 ΔyhjH in semi-solid agar (Fig. 2A). These results are consistent with the possibility that all three proteins, PdeB, PdeC, and PdeD, function as c-di-GMP PDEs. However, overexpressed but enzymatically inactive EAL domain proteins that retain the ability to bind (but not to hydrolyze) c-di-GMP also can lower intracellular c-di-GMP concentration thus mimicking the phenotypes of overexpressed PDEs [53].

To resolve the ambiguity regarding the enzymatic activity of the PdeB-D proteins, we purified each protein and tested its ability to hydrolyze c-di-GMP in vitro. The PdeB and PdeD proteins were overexpressed and purified as N-terminal His6-tagged fusions. Since the His6-tagged PdeC fusion proved to be insoluble, PdeC was purified as a fusion to maltose-binding protein (MBP) (Fig. 2B). The ability of purified PdeB, PdeC, or PdeD to hydrolyze c-di-GMP was assessed by measuring the substrate and products of reactions over time using HPLC, as described previously [38]. Fig. 2C shows that all three recombinant proteins possess c-di-GMP PDE activities in vitro.
L. monocytogenes DgcA-C proteins possess DGC activities
The functionality of putative L. monocytogenes DGC proteins was assessed by monitoring swim zone sizes in semi-solid agar. The three dgc genes were cloned into the pBAD/Myc-His vector under the control of an arabinose-inducible promoter. Each of the three dgc genes decreased, to various degrees, the sizes of the swim zones of strain MG1655, which is highly motile in the absence of heterologous DGCs (Fig. 3A).

To exclude the possibility of nonspecific motility inhibition (e.g., due to protein toxicity), we assessed a second c-di-GMP-dependent phenotype that is independent of motility inhibition. In E. coli BL21 (DE3), c-di-GMP induces synthesis of curli fimbriae that can be detected by staining with Congo red dye [47]. As shown in Fig. 3B, BL21 (DE3) strains expressing each of the three Dgc proteins individually exhibited more intensely colored colonies on Congo red agar compared to the negative control expressing an empty vector. Together, these results support the prediction that the DgcA-C proteins possess DGC activity.
L. monocytogenes phenotypes associated with perturbed intracellular c-di-GMP levels
Having established that L. monocytogenes EGD-e possesses functional components for c-di-GMP-mediated signaling, we examined phenotypes associated with elevated and decreased intracellular c-di-GMP levels. To perturb c-di-GMP levels, we expressed in the EGD-e strain two c-di-GMP metabolizing enzymes characterized by us previously, i.e., DGC (Slr1143 from Synechocystis sp. [37]) and PDE (YhjH from E. coli [51]), and assessed their role in swimming motility and EPS production. The use of heterologous proteins allowed us assess the effects of changing intracellular c-di-GMP levels without undesired changes in protein-protein interactions that may have been occured if we were to overexpress listerial DGC and PDE enzymes.
L. monocytogenes uses flagella for motility [54]. Expression of Slr1143 blocked swimming of strain EGD-e in semi-solid agar, whereas expression of YhjH had no effect (Fig. 4A top). Expression of Slr1143 also resulted in more pigmented L. monocytogenes colonies on Congo red agar, whereas expression of YhjH had no observable phenotype (Fig. 4B, sectors 10 versus 1 and 9). Later in this work we show that YhjH is expressed and functional as a PDE in L. monocytogenes. Therefore, we interpreted the lack of a phenotype associated with YhjH overexpression as an indication that intracellular c-di-GMP levels in strain EGD-e are already low, and that c-di-GMP does not play a significant role under the conditions used in these assays. Since L. monocytogenes is not known to synthesize pili, and the genome of strain EGD-e has no candidate pili genes, we hypothesized that Congo red staining was indicative of EPS production. An EPS has been suspected in some naturally occurring L. monocytogenes isolates [28]. Further, Tiensuu and colleagues have recently observed Congo red staining rings within L. monocytogenes colonies exposed to dark-light cycles [55], however the nature of the Congo red-binding extracellular polymer was not investigated.

Construction and characterization of the L. monocytogenes dgc and pde mutants
Having identified two phenotypes associated with elevated c-di-GMP levels, we proceeded to inactivate, individually and in combination, the L. monocytogenes pdeB-D genes. Based on the inhibition of swim zones in semi-solid agar by the heterologous DGC, Slr1143, we expected pdeB-D mutations to result in smaller swim zones. However, inactivation of individual pde genes did not significantly affect swim zone sizes (Fig. 4C). Inactivation of pairs of pde genes produced relatively minor decreases in swim zones sizes, while simultaneous deletion of all three pde genes, ΔpdeB/C/D, produced a mutant severely impaired in swimming in semi-solid agar (Fig. 4C). This phenotype is similar to the phenotype of the wild type EGD-e expressing the heterologous DGC, Slr1143 (Fig. 4A top). These results suggest that the PDEs have at least partially overlapping functions in degrading intracellular c-di-GMP.
As expected, expression of Slr1143 in the triple ΔpdeB/C/D mutant did not affect the already inhibited motility any further (Fig. 4A bottom). However, expression of YhjH in this mutant fully restored the swim zone to the size of the wild-type strain, thus showing that YhjH is expressed and functional in L. monocytogenes (Fig. 4A bottom), and that motility inhibition in semi-solid agar was due to elevated c-di-GMP levels in the triple ΔpdeB/C/D mutant.
We proceeded to test the effects of L. monocytogenes pde mutations on Congo red binding. The triple ΔpdeB/C/D mutant showed significant accumulation of Congo red (Fig. 4B, sector 2 versus 1), similar to the wild type strain expressing Slr1143 (Fig. 4B, sector 10). Expression of YhjH, but not Slr1143, in the triple ΔpdeB/C/D mutant, inhibited Congo red accumulation (Fig. 4B, sector 6 versus 5 or 7). Individual pde mutants did not affect Congo red staining, while among double mutants, the pdeB/C mutant showed some staining (Figure S1).
In addition to Congo red binding, the colonies of the ΔpdeB/C/D mutant were found to have rough edges, compared to smooth-edged colonies of the wild type strain (Fig. 4D). The observed changes in colony morphology in the ΔpdeB/C/D mutant were not as pronounced as the wrinkled or rough colony morphologies reported in the proteobacterial species overexpressing EPS [56]–[59], however, when combined with enhanced Congo red binding, these changes are indicative of EPS production.
Inactivation of the dgc genes, individually or in combination, resulted in no observable phenotypes, just like the expression of YhjH in the wild type strain produced no phenotype. We therefore conclude that c-di-GMP plays little, if any role, in strain EGD-e grown under these laboratory conditions.
Bioinformatics-based identification of the putative EPS biosynthesis pssA-E operon in L. monocytogenes
We searched the L. monocytogenes genome for EPS biosynthesis genes potentially responsible for c-di-GMP-induced Congo red binding. The lmo0527-0531 operon, designated here pssA-E (polysaccharide synthesis) (Fig. 1), emerged as the prime candidate for this role based on the following reasoning. The last gene of the operon, pssE (lmo0531), encodes a degenerate GGDEF domain protein, hypothesized to function as a c-di-GMP receptor (Fig. 1). If this assumption is correct, PssE may be involved in a c-di-GMP-dependent activation of EPS synthesis, similar to activation of cellulose [1], [51], alginate [60] and Pel EPS synthesis [48] in Proteobacteria. An additional reason to implicate the pssA-E cluster in EPS biosynthesis was based on the presence of the putative glycosidase gene, lmo1913, in the dgcA-dgcB-lmo1913-pdeC operon that encodes enzymes for synthesis and hydrolysis of c-di-GMP (Fig. 1). Glycosidases counterbalance glycosyltransferases and are integral components of EPS synthesis and degradation apparati.
The pssA-E operon appears to encode enzymes associated with biosynthesis of poly--1,6-N-acetyl-D-glucosamine (PNAG) or poly--1,4-D-glucopyranose (cellulose), either of which is capable of binding Congo red, or yet another EPS. The key player in this operon is PssC (Lmo0529), which is predicted to function as type 2 glycosyltransferase responsible for the polymerization reaction. PssC shows the highest (∼30%) identity (over an ∼300 amino acid region) to the N-acetylglycosyltransferases involved in PNAG synthesis from S. aureus (IcaA) and Yersinia pestis (HmsR) [61], [62]. However, no other genes found in the staphylococcal ica or yersinial hms gene clusters are present in the pssA-E operon. Instead, the gene downstream of pssC, pssD (lm0530), encodes an ortholog of the BcsB subunit of bacterial cellulose synthases [63]. The BcsB proteins have thus far been associated exclusively with cellulose synthases, yet they are involved in the membrane passage of the polysaccharide polymer not its synthesis [64], therefore BcsB can possibly accommodate polymers of different composition than cellulose. It is noteworthy that the glycosyltransferases catalyzing cellulose synthesis also belong to type 2 glycosyltransferases, like the PNAG synthases [63]. Further, PssC shares ∼25% identity with the type 2 glycosyltransferase BcsA of the cellulose synthase complex of Rhodobacter sphaeroides [64]. The almost equal similarity of the listerial glycosyl transferase to PNAG - and cellulose synthases makes predictions of the composition of the listerial EPS unreliable.
The pssA-E gene cluster is indeed responsible for listerial EPS synthesis
To test the involvement of the pssA-E gene cluster in EPS biosynthesis, we deleted the predicted glycosyltransferase gene, pssC, in the ΔpdeB/C/D background. We found that the constructed ΔpdeB/C/D ΔpssC mutant no longer bound Congo red (Fig. 4B, sector 4). This result supports the hypothesis that the pssA-E operon is responsible for c-di-GMP-induced EPS biosynthesis. To verify it further, we tested another bioinformatics-based prediction, i.e. that inactivation of pssE in the ΔpdeB/C/D background will also impair EPS synthesis. Indeed, the constructed ΔpdeB/C/D ΔpssE mutant did not bind Congo red either (Fig. 4B, sector 3). We conclude that PssE, a putative c-di-GMP receptor, plays a critical role in EPS synthesis. Complementation of the ΔpdeB/C/D ΔpssC and ΔpdeB/C/D ΔpssE mutants with individually cloned pssC and pssE, respectively, restored Congo red binding (data not shown) verifying that the ΔpssC and ΔpssE mutations were responsible for the mutant phenotypes. The ΔpssC and ΔpssE mutations in the ΔpdeB/C/D mutant background reversed the rough colony phenotype back to a smooth appearance (Fig. 4D and data not shown).
Biochemical evidence that the PssE protein is a c-di-GMP receptor
To test the prediction that the PssE protein acts as a c-di-GMP receptor, we overexpressed its GGDEF domain containing the I-site as an MBP fusion (MBP-GGDEFpssE), purified this protein (Fig. 5A) and analyzed its ability to bind c-di-GMP in vitro using equilibrium dialysis. MBP-GGDEFpssE was found to bind c-di-GMP with an apparent Kd of 0.79±0.17 µM (Fig. 5B). This value falls within the range of physiologically relevant intracellular c-di-GMP concentrations measured in other bacteria that are believed to be in the submicromolar to low micromolar range [5]. The binding capacity of the MBP-GGDEFpssE protein, Bmax, was calculated to be 2.03±0.12 µM c-di-GMP (µM protein)−1 indicating that each PssE molecule can bind two c-di-GMP molecules at saturation. This result is consistent with the observation of an intercalated c-di-GMP dimer bound to the I-sites of crystallized GGDEF domain proteins [2], [3]. Therefore, PssE is a bona fide c-di-GMP receptor that is predicted to transfer the c-di-GMP signal to activate synthesis of the listerial Pss EPS.

C-di-GMP-induced listerial EPS promotes cell aggregation but plays limited role in biofilm formation on abiotic surfaces
PNAG and cellulose increase biofilm formation by the proteobacterial species on abiotic surfaces. To test the effect of c-di-GMP-induced EPS in L. monocytogenes, we performed a conventional Crystal violet dye-binding assay that measures the biomass of cells attached to the wells of microtiter plates following removal of liquid cultures [65]. Surprisingly, we did not observe an increase in biofilm levels in the ΔpdeB/C/D mutant, compared to the wild type, when these strains were grown in LB medium (where biofilm formation of strain EGD-e is low). We observed only a marginal increase in surface-attached biofilm levels in LB supplemented with glycerol (where biofilms are greatly stimulated) (Fig. 6A). Interestingly, this increase in biofilm levels was observed in all ΔpdeB/C/D strains grown in LB plus glycerol, whether or not they produced EPS (Fig. 6A). These results suggest that, instead of the anticipated stimulation of biofilms, listerial EPS may actually inhibit biofilm formation, at least under certain conditions. They also implicate a c-di-GMP-activated non-EPS component in biofilm stimulation. Similar to the results on polystyrene surfaces, the ΔpdeB/C/D mutant produced no more biofilm in LB medium on glass or metal (aluminum foil or steel coupons) surfaces than did the wild type (data not shown).

We noticed that incubation of the ΔpdeB/C/D mutant (but not the wild type, ΔpdeB/C/D ΔpssC or ΔpdeB/C/D ΔpssE mutants) in liquid glucose-rich minimal HTM medium resulted in cell clumping (Fig. 6B). This suggests that listerial EPS strengthens intercellular interactions but not bacterial interactions with abiotic surfaces. The pssC and pssE gene deletions in the ΔpdeB/C/D background completely abolished clumping, just like they decreased Congo red binding in BHI plates. This result confirms that listerial EPS is responsible for clumping.
C-di-GMP-dependent EPS impairs L. monocytogenes motility in semi-solid agar
Since the EPS producing ΔpdeB/C/D mutant was impaired in swimming in semi-solid agar (Fig. 4C), we set out to explore the effect of EPS on motility. Surprisingly, inactivation of EPS synthesis by the pssC or pssE mutations, restored swimming of the ΔpdeB/C/D mutant in semi-solid agar to the wild-type levels (Fig. 4E). It therefore appears that swimming in semi-solid agar was impaired exclusively due to listerial EPS.
To gain additional insight into this issue, we analyzed the motility of the wild type, the ΔpdeB/C/D mutant and the ΔpdeB/C/D ΔpssC mutant in liquid medium where clumping is minimal and not detectable by the naked eye. Phase contrast microscopic observations revealed that single cells of the ΔpdeB/C/D and ΔpdeB/C/D ΔpssC mutants were as motile as the wild-type cells. These results favor the scenario whereby EPS accumulated on cell surfaces results in cell aggregation, which inhibits spreading of the cells in semi-solid agar.
C-di-GMP-induced EPS significantly enhances L. monocytogenes tolerance to disinfectants and desiccation
EPS is known to protect bacteria from environmental insults (27). Here we evaluated the role of L. monocytogenes EPS in providing tolerance to disinfection and desiccation. The wild-type strain EGD-e as well as its ΔpdeB/C/D mutant synthesizing EPS and grown under clump-forming conditions were subjected to selected disinfectants commonly used in the food-processing industry and produce storage facilities: sodium hypochlorite (bleach), benzalkonium chloride (a quaternary ammonium compound) and hydrogen peroxide [66]. To distinguish between the contribution of EPS versus EPS-independent c-di-GMP-responsive agents, we included in the tests the ΔpdeB/C/D ΔpssC mutant characterized by elevated intracellular c-di-GMP levels but defective in EPS production.
The sodium hypochlorite treatment applied here was highly effective in killing the wild-type strain, but not the EPS producing ΔpdeB/C/D mutant, whose survival was approximately >106-fold higher than the survival of the wild type (Fig. 6C). Tolerance of the ΔpdeB/C/D mutant to hydrogen peroxide and benzalkonium chloride treatments was also highly, approximately 102-fold, higher, compared to the wild type or the EPS-deficient ΔpdeB/C/D ΔpssC mutant (Fig. 6C). These observations suggest that the c-di-GMP-induced EPS is a critical factor responsible for increased tolerance to these agents.
EPS also enhanced survival of L. monocytogenes to long-term desiccation. In this experiment, the liquid-grown cultures were centrifuged, and the pellets were kept in a desiccator for 7 or 21 days. We found that the desiccation survival rates of the EPS producing ΔpdeB/C/D strain were significantly higher, compared to those of the wild type or the ΔpdeB/C/D ΔpssC mutant (Fig. 6D). These results suggest that EPS provides superior protection not only against various commonly used disinfectants in food processing plants but also to desiccation, which may enhance listerial survival during food transportation and storage.
Elevated c-di-GMP levels inhibit L. monocytogenes invasion into mammalian cells
As a foodborne pathogen, L. monocytogenes is expected to use gut epithelial cells for primary invasion [17]–[20]. We examined the consequences of elevated c-di-GMP levels on bacterial invasion into HT-29 human colon adenocarcinoma cells. As shown in Fig. 7A, the strains with elevated c-di-GMP levels were significantly impaired in invasion, whether elevated c-di-GMP was caused by expression of the heterologous DGC, Slr1143, or by the ΔpdeB/C/D mutations. Consistent with the inhibitory role of c-di-GMP, invasion was increased, by approximately 2-fold, in the L. monocytogenes strain expressing a c-di-GMP PDE, YhjH.
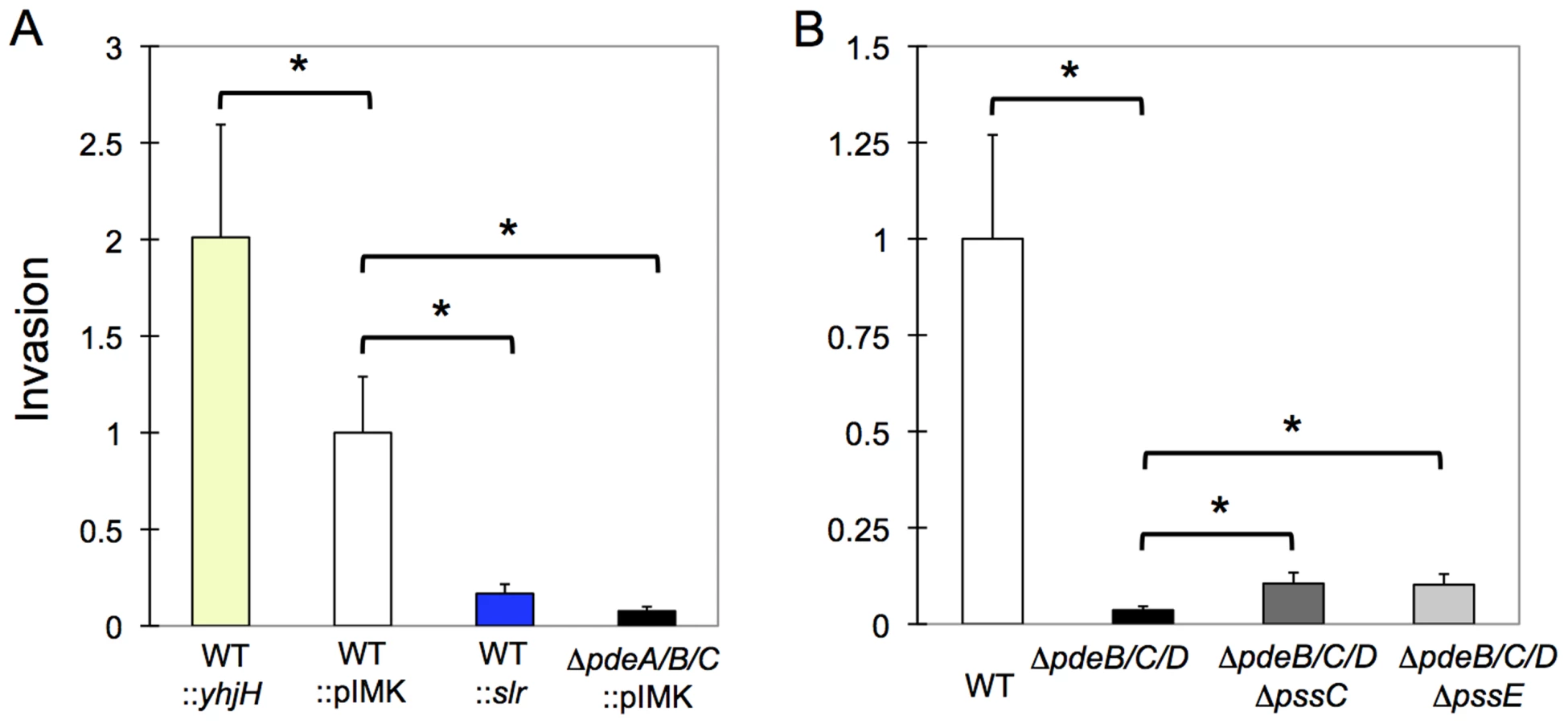
Next, we tested what role the c-di-GMP-induced EPS may have played in invasion inhibition. We observed that the ΔpdeB/C/D ΔpssC and ΔpdeB/C/D ΔpssE mutants showed approximately 2–2.5-fold greater invasiveness compared to the ΔpdeB/C/D mutant (Fig. 7B), but remained approximately 10-fold less invasive than the wild type strain. These results suggest that while EPS moderately inhibits invasion, the major reason for the defective invasion is a c-di-GMP-induced component(s) different from EPS. The nature of this component(s) and the mechanisms through which it inhibits listerial invasion remain to be investigated.
Elevated c-di-GMP levels reduce systemic spread of L. monocytogenes in mice infected via an oral route
To assess the role of c-di-GMP signaling in vivo, we used a newly developed mouse model of foodborne listeriosis [67]. Groups of BALB/c/By/J mice were fed either wild-type EGD-e or the ΔpdeB/C/D mutant, and the bacterial load in various tissues was assessed 60 h post infection. There was no significant difference in colonization of the ileum, colon or spleen at this time point (Fig. 8). However, the ΔpdeB/C/D triple mutant was significantly impaired in colonizing both the liver and the gallbladder. The decreased bacterial load in the liver appears to be linked to EPS, since the ΔpssC mutation in the ΔpdeB/C/D background restored the bacterial load to the wild-type level (Fig. 8). In fact, no significant differences in bacterial loads in the liver were observed when the same L. monocytogenes strains were injected intravenously (Fig. S2), suggesting that increased levels of c-di-GMP may alter the ability of the bacteria to disseminate from the gut.

Discussion
We predicted that the L. monocytogenes EGD-e genome encodes three GGDEF domain DGCs, one inactive GGDEF domain protein and three EAL domain PDEs, and verified this prediction by a combination of genetic and biochemical tests. Interestingly, all of the enzymes involved in c-di-GMP metabolism are highly conserved not only in the genomes of L. monocytogenes isolates but also in other Listeria species, e.g. L. innocua, L. ivanovii, L. seeligeri, and L. welshimeri. The high conservation of these proteins implies that c-di-GMP signaling pathways play important roles in the evolutionary success of Listeria. Such conservation is striking in light of the flexibility in the organization of c-di-GMP signaling pathways observed in other Firmicutes. For example, in the genus Bacillus, the number of enzymes involved in c-di-GMP synthesis and hydrolysis varies from three to eleven; it varies from eight to forty in the genus Clostridium (http://www.ncbi.nlm.nih.gov/Complete_Genomes/SignalCensus.html).
We discovered that c-di-GMP regulation affects at least two targets in L. monocytogenes. One of these targets is a novel EPS (Fig. 4B, 4D, 6B). This finding resolves the long-standing controversy regarding the ability of listeria to produce EPS [27], [28]. The second (and possibly additional) target of c-di-GMP regulation, whose identity remains unknown, appears to be responsible for the drastic inhibition of listerial invasiveness in mammalian cells (Fig. 7), modest stimulation of biofilm formation on abiotic surfaces in LB supplemented with glycerol (Fig. 6A) and lower pathogen accumulation in certain mouse organs following oral infection (Fig. 8).
Here, we revealed that the c-di-GMP induced EPS is synthesized by the pssA-E operon. The composition of the listerial EPS is difficult to predict because, while some Pss proteins share similarity to the components of cellulose synthases and PNAG synthases, other components are unique. Interestingly, in contrast to cellulose or PNAG, both of which promote biofilm formation on abiotic surfaces [61], [62], [64], [68], listerial EPS either does not affect or inhibits biofilm formation on abiotic surfaces. Instead, it promotes cell aggregation, in minimal media (Fig. 6B). These observations favor the hypothesis that the composition of listerial EPS is different from cellulose or PNAG.
In this study, we identified the mechanism through which c-di-GMP activates EPS synthesis in L. monocytogenes. C-di-GMP binds to the I-site receptor PssE, whose gene is located in the pss operon, and whose function is essential for EPS biosynthesis. Bacterial cellulose synthases studied thus far are activated via c-di-GMP-binding PilZ domains linked to the C-termini of BcsA subunits [1], [51], [64]. The PNAG synthase of E. coli is activated by c-di-GMP binding to two subunits, PgaD and PgaC, one of which, PgaD, is proteolytically degraded in the absence of c-di-GMP [69]. Perhaps the most similar c-di-GMP-dependent mechanism to that operating in L. monocytogenes Pss synthase involves the Pseudomonas aeruginosa Pel EPS synthase, which is activated via an I-site c-di-GMP receptor protein [48], [70].
We showed that the L. monocytogenes Pss EPS is responsible for multiple phenotypes, i.e., cell aggregation, decreased motility in semi-solid media, moderate inhibition of invasiveness in mammalian cells, and drastically elevated tolerance to disinfectants and desiccation. The latter effects of c-di-GMP-induced listerial EPS are particularly noteworthy in light of the increasing frequency of listerial outbreaks associated with produce. EPS may contribute to enhanced survival of listeria on produce surfaces during washing with disinfectants as well as during transporation and storage of listeria-contaminated produce. It is also possible that listerial EPS contributes to bacterial survival in food-processing facilities. It will be interesting to investigate whether listerial strains from the recent outbreaks synthesize EPS, and what role, if any, it may have played in their survival and disease causing abilities.
C-di-GMP-induced motility inhibition is common in Proteobacteria. One of the best-understood mechanisms of c-di-GMP-induced motility inhibition involves YcgR, the PilZ-domain c-di-GMP backstop brake that operates in E. coli and related enteric bacteria. YcgR binds to the flagellar switch complex and, at elevated c-di-GMP concentrations, introduces a rotational bias that decreases the frequency of flagella reversals and therefore, the frequency of changes in swimming direction [52], [71], [72]. The smooth, almost unidirectional, swimming results in bacteria being trapped in blind alleys of semi-solid agar [73]. YcgR may also slow down rotating flagella [74]. A similar mechanism has been proposed for a PilZ domain protein in B. subtilis, however important details have yet to be elucidated [31]. A different mechanism of c-di-GMP-induced motility inhibition was described in Caulobacter crescentus, where a PilZ domain receptor affects the abundance of a flagellum assembly regulatory subunit [75]. B. subtilis has yet another mechanism of motility inhibition that involves a bifunctional protein EpsE that acts as a glycosyl transferase involved in EPS synthesis and as a molecular clutch that disengages the flagellum rotor from the membrane-localized energy-supplying stator [76], [77]. Whether an EpsE-like clutch operates in L. monocytogenes remains unknown, however it is clear that no clutch or break is induced by c-di-GMP because liquid-grown cells show no obvious motility defects. The most striking observation is that inactivation of Pss synthesis is sufficient to restore motility in semi-solid agar. Therefore, listerial spreading in semi-solid agar appears to be inhibited due to cell aggregation and possibly flagella trapping in the EPS. Recently, a similar mechanism has been described in S. enterica, which at high c-di-GMP levels, secretes cellulose [78].
Listerial EPS inhibits bacterial invasiveness in mammalian cells, however, only modestly, 2–2.5-fold, whereas an as yet unidentified c-di-GMP pathway is responsible for a much larger component of invasiveness inhibition. The composition of this new c-di-GMP signaling pathway remains unknown. In this regard, it is noteworthy that PssE is the only c-di-GMP receptor that can be predicted based on the genome sequence analysis. Listeria lack other identifiable c-di-GMP receptor proteins or c-di-GMP-sensing riboswitches (reviewed in [2], [3], [6]).
In addition to uncovering the role of c-di-GMP in vitro, we tested its role in virulence using a recently developed food borne mouse disease model that closely mimics human infection (67). We found that elevated c-di-GMP levels decreased listerial infection in the liver, and that this defect could be restored by abolishing EPS biosynthesis. Thus, it is possible that c-di-GMP induced EPS impairs the ability of L. monocytogenes to either efficiently disseminate from the intestine or to replicate and spread from cell-to-cell in hepatocytes. While we observed a significant defect in the ability of the ΔpdeB/C/D mutant to invade HT-29 colon carcinoma cells in vitro, there was no difference in the ability of the ΔpdeB/C/D mutant to colonize the murine intestines, compared to the wild type strain. Thus, increased c-di-GMP levels may impair the direct invasion of intestinal epithelial cells mediated by InlA/E-cadherin interactions, but does not significantly impede the ability of L. monocytogenes to translocate across the gut mucosa barrier, presumably because the bacteria use alternate mechanisms of invasion. L. monocytogenes can transcytose across M cells, specialized epithelial cells that are found both overlying Peyer's patches and scattered elsewhere throughout the epithelium [79]–[81]. It is also possible that specialized subsets of dendritic cells in the intestinal lamina propria can engulf L. monocytogenes by extending dendrites into the gut lumen, a process that has been demonstrated during oral S. enterica infection [82], [83].
Another issue pertaining to this study concerns the role of cyclic dinucleotides as bacterial biomarkers recognized by the innate immune system and in the stimulation of the host intracytoplasmic surveillance response. Recently, the listerial second messenger c-di-AMP, which is structurally related to c-di-GMP, has been shown to be secreted into the cytosol of infected mammalian cells where it triggers interferon (IFN) production via the STING-signaling cascade [84]–[86]. A robust IFNβ response promotes the growth of L. monocytogenes administered intravenously [87]. We showed here that the ΔpdeB/C/D mutant, which likely has the highest c-di-GMP production achievable during L. monocytogenes intracellular growth, was overall less infective following oral infection. This suggests that elevated c-di-GMP levels play a negative role in L. monocytogenes virulence, in an apparent contrast with the role of c-di-AMP [87]. However, it is premature to draw definitive conclusions because (i) while c-di-AMP secretion enhances intracellular growth and spread of L. monocytogenes, overproduction of c-di-AMP suppresses listerial virulence [88], and (ii) individual c-di-GMP signaling pathways often play different, even opposite, roles in host-pathogen interactions [reviewed in ref. 2]. Therefore, more detailed analysis of c-di-GMP synthesis and secretion during listerial intracellular growth will be required to figure out the roles of c-di-GMP signaling pathways in L. monocytogenes virulence.
In addition to questions regarding intracellular growth and spread of listeria in different organs, our study raises numerous other questions pertaining to c-di-GMP signaling in listeria. How does c-di-GMP signaling inhibit cell invasion? What signals control c-di-GMP synthesis? Can c-di-GMP signaling pathways be manipulated to inhibit listerial cell invasion? What is the composition of the Pss EPS? Does this EPS contribute to listerial colonization of produce surfaces? Is it made by the L. monocytogenes strains from recent produce-associated outbreaks? Does it affect survival of such strains during disinfection in food processing facilities and desiccation during transportation and storage? These questions will have to be addressed in the future.
Materials and Methods
Ethics statement
This work was performed in accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health. All procedures were approved by the Institutional Animal Care and Use Committee (IACUC) at the University of Kentucky (permit number A-3336-01).
Bacterial strains, plasmids and growth conditions
The bacterial strains and plasmids used in this study are listed in Table 1. The primers used in this study are listed in Table S1. E. coli was routinely grown in LB medium supplemented with appropriate antibiotics at 25, 30 or 37°C, as indicated. L. monocytogenes was grown in Brain Heart Infusion (BHI) medium (Difco), HTM (minimal medium containing 3% glucose) [89] or LB, supplemented with appropriate antibiotics at 25, 30, 37 or 42°C, as indicated.

Plasmid and mutant construction
Genomic DNA of L. monocytogenes EGD-e was purified from bacterial cells using a Bactozol kit (Molecular Research Center, OH). L. monocytogenes genes were PCR amplified using genomic DNA, Vent DNA polymerase (New England Biolabs), and gene-specific primers (Table S1). PCR fragments were gel purified with the Gel Purification kit (Qiagen), digested with the appropriate restriction enzymes, and cloned into vector pMAL-c2x (New England Biolabs) in strain DH5α or into vector pET23a (Invitrogen) in strain BL21(DE3) containing pLysS (Invitrogen).
In-frame deletions in the pdeB/C/D, dgcA/B/C, pssC and pssE genes were generated by site-directed mutagenesis by splice-by-overlap extension PCR. The PCR products containing genes with in-frame deletions were cloned into the temperature-sensitive shuttle vector pKSV7 [90]. The recombinant sequences were used to replace the corresponding wild type sequences in the chromosome of the L. monocytogenes EGD-e strain by allelic exchange, as previously described [90]. L. monocytogenes was electroporated as described earlier [91].
Motility and Congo red dye binding assays
The analysis of swimming in semi-solid agar was performed essentially as described [51]. Briefly, 2 µl of overnight cultures was inoculated onto soft agar plates containing 0.25% agar, 1% tryptone and 0.5% NaCl. Diameters of the swimming zones were assessed after 6-h incubation at 37°C for E. coli and 12–18-h incubation at 30°C for L. monocytogenes.
For Congo red binding assays, LB (E. coli) or BHI (L. monocytogenes) agar plates containing 40–80 µg ml−1 Congo red were incubated at 30°C for 48–72 h.
Biofilm assays
Surface-adhered biofilm formation was assayed in a 96-well format using a modified version of a previously published protocol [92]. Briefly, overnight cultures grown in BHI at 30°C (A600, 2.5–3.5) were diluted into freshly made BHI, LB or LB supplemented with 3% glycerol to an initial A600 of 0.05–0.1, and 150 µl aliquots of each culture were inoculated into each of six wells. Biofilms attached to wells were measured following growth for 1–6 days at 30°C. Biofilms were stained with a 0.1% aqueous solution of Crystal violet dye, which was subsequently dissolved in 33% acetic acid and quantified by measurement of A595 [65].
Protein overexpression and purification
For purification of PdeB::His6 and PdeD::His6, isopropyl-β-D-thiogalactopyranoside (IPTG) (final concentration, 0.2 mM) was added to exponentially (A600, 0.6–0.7) growing cultures of E. coli BL21 (DE3) pLysS containing appropriate overexpression plasmids. After 2 to 4 h of induction at 30°C, the cells were chilled to 4°C and collected by centrifugation. The cell pellets were resuspended in buffer (pH 8.0) containing 300 mM NaCl, 50 mM NaH2PO4, and 10 mM imidazole and protease inhibitors (phenylmethylsulfonyl fluoride and P8849 protease inhibitor cocktail) at the concentrations specified by the manufacturer (Sigma-Aldrich). The cell suspensions were passed through a French press mini-cell (Spectronic Instruments, NJ), followed by a brief sonication using a Sonifier 250 (Branson Ultrasonics, CT). The crude cell extracts were centrifuged at 15,000× g for 45 min. Soluble protein fractions were collected and mixed with preequilibrated Ni2+ resin (Qiagen) for 1 h at 4°C, which was placed into a column and extensively washed with the resuspension buffer containing 20 mM imidazole. The proteins were subsequently eluted using 200 mM imidazole. The buffer was exchanged with PDE buffer [38] using desalting columns according to the instructions of the manufacturer (Pierce Biotechnology). Protein purity was assessed by SDS-PAGE and protein concentration was determined using a BCA protein assay kit (Pierce Biotechnology).
For purification of MBP::PdeC and MBP::GGDEFpssE fusions, IPTG (final concentration, 0.2 mM) was added to exponentially (A600, 0.6–0.8) growing E. coli DH5α containing appropriate plasmids. After 2-h induction, cells were collected by centrifugation. Cell pellets were resuspended in a buffer containing 200 mM NaCl, 0.5 mM EDTA, 5 mM MgCl2, 20 mM Tris-HCl (pH 7.6), and 5% glycerol that also contained protease inhibitors. Following cell disruption and clearing of the crude cell extracts, as described above, soluble protein fractions were mixed with pre-equilibrated amylose resin (New England Biolabs) for 1 h at 4°C, which was subsequently extensively washed with the resuspension buffer. MBP fusions were eluted with maltose and the buffer was exchanged for PDE or c-di-GMP binding assay [51] buffer using desalting columns.
PDE assays
Assays were performed essentially as described by Schmidt et al. [38]. Briefly, a PDE enzyme (1–5 µM) was added to PDE reaction buffer (final volume, 100 µl) containing 250 µM c-di-GMP, and the reaction was allowed to proceed at 37°C. Aliquots were withdrawn at various time points; the reaction was stopped by addition of CaCl2 (final concentration, 10 mM), and the sample was boiled for 3 min and centrifuged. The supernatant was then filtered through a 0.22-µm filter, and the reaction products were analyzed by reversed-phase HPLC (Summit HPLC system; Dionex) using a Supelcosil LC-18-T column (Sigma-Aldridge). The buffer system and gradient elution program were described previously [37].
Equilibrium dialysis
Equilibrium dialysis experiments were performed as described earlier [51]. Briefly, MBP-GGDEFPssE (20 µM) was injected into one of the two chambers of a Dispo-Biodialyzer cassette (10 kD cutoff, The Nest Group, MA) filled with dialysis buffer. c-di-GMP (concentrations from 1 to 50 µM) was injected into the opposite cell of the cassette. The cassettes were maintained for 25 h at room temperature under agitation, after which samples from each chamber were withdrawn, boiled for 3 min, centrifuged, and supernatants were filtered through a 0.22-µm microfilter. The nucleotide concentrations were quantified by HPLC. Binding constants were calculated by the GraphPad Prism software, version 4.03 (GraphPad Software, San Diego, CA) using a nonlinear regression model.
Invasion assay
L. monocytogenes invasion properties were analyzed using a gentamicin-based assay with HT-29 human colon adenocarcinoma cell monolayers in 24-well plates, essentially as described [93], [94]. Briefly, overnight cultures of L. monocytogenes grown in BHI at 37°C were centrifuged, washed and resuspended in DMEM medium. Monolayers of HT-29 cells were inoculated with 100 µl of the L. monocytogenes suspensions (∼5×108 CFU ml−1) at a multiplicity of infection of 100 and incubated for 1.5 h at 37°C in a 7% CO2 atmosphere. The monolayers were then washed and incubated in the presence of 100 µg gentamicin ml−1 (final concentration) for 1.5 h. Following this incubation, the cell monolayers were washed again and lysed with 0.1% Triton X-100. Appropriate dilutions were plated on BHI plates for enumeration of intracellular bacteria. Each experiment was done in triplicate, and experiments were performed at least three times independently. Statistical analysis was performed by using Tukey's test at p of <0.05.
Foodborne infection of mice
Female BALB/c/By/J mice were purchased from The Jackson Laboratory (Bar Harbor, ME) at 5 weeks of age and used in experiments when they were 6–9 weeks old. Mice were maintained in a specific pathogen free facility at the University of Kentucky and all procedures were performed in accordance with IACUC guidelines. Aliquots of early stationary phase bacteria were prepared and stored at −80°C. To prepare the inoculum, aliquots were thawed on ice, cultured standing in BHI broth for 1.5 h at 30°C, washed once in PBS and then suspended in 5 µl of melted, salted butter (Kroger) and used to saturate a 2–3 mm piece of white bread (Kroger). Infection by the natural feeding route was carried out at night as described [67]. Briefly, mice were given unrestricted access to water but denied food for 22 h, then placed in an empty cage, and given 5–10 minutes to pick up the contaminated bread piece and eat all of it. Mice were then returned to their original cages with raised wire flooring to prevent coprophagy, and normal mouse chow was replenished.
Processing of tissue samples
Colon contents were removed by squeezing with sterile forceps and then flushing with 8 ml of PBS through a 25 g needle. Washed tissues were cut longitudinally and homogenized for 1 min in 2 ml of sterile water using a PowerGen 1000 homogenizer (Fisher) at 60% power. The total number of cell-associated (adherent plus intracellular) L. monocytogenes cells was determined by plating serial dilutions on BHI agar supplemented with 15 g LiCl l−1 and 10 g glycine l−1 (BHI/L+G). Colonies were counted after 48 h incubation at 37°C. This selective agar inhibited the growth of most intestinal microbiota; suspect colonies were confirmed to be L. monocytogenes by plating on CHROMagar Listeria plates (Becton Dickinson). Spleens and livers were harvested aseptically and homogenized for 30 sec in 2 ml of sterile water. Gallbladders were ruptured with sterile scissors in a microfuge tube containing 1 ml of sterile water and vortexed for 30 sec. Dilutions of each tissue were plated on BHI/L+G agar.
Disinfection and desiccation tolerance
Solutions of sodium hypochlorite, hydrogen peroxide and benzalkonium chloride (Sigma-Aldrich and Sigma Life Sciences) were prepared in sterile phosphate-buffered saline with disinfection concentrations of 1600 ppm, 200 mM, and 100 ppm, respectively. Cultures were grown in HTM with 3% glucose at 37°C for 24 h, at which point small uniform clumps are formed by the ΔpdeB/C/D strain. Aliquots (250 µl; 108 cfu/ml) of these cultures were mixed with disinfectants at 1∶1 vol ratios in small glass tubes that also contained 0.1 g of acid washed glass beads (Sigma Life Sciences). Following a 10-min exposure to disinfectants at room temperature, D/E neutralizing broth (500 µl; Difco) was added [95]. Samples were vigorously vortexed (for 5 min) and clumps of the ΔpdeB/C/D strain were dispersed due to the action of the glass beads. Serial dilutions were plated on BHI agar and colonies were counted following a 48-h incubation at 37°C.
To assess desiccation tolerance, strains were grown as described above. One milliliter of cultures (5×108 cfu/ml) was centrifuged in 1.5 ml eppendorf microtubes containing 0.1 g glass beads. After supernatant removal, the tubes were stored at room temperature in a desiccator jar containing anhydrous calcium sulfate. After 7 and 21 days, the pellets were resuspended in phosphate buffered saline, vigorously vortexed, and plated on BHI agar. Colonies were counted following 48-h incubation at 37°C.
Supporting Information
Zdroje
1. RossP, WeinhouseH, AloniY, MichaeliD, OhanaP, et al. (1987) Regulation of cellulose synthesis in Acetobacter xylinum by cyclic diguanylic acid. Nature 325 : 279–81.
2. RömlingU, GalperinMY, GomelskyM (2013) Cyclic di-GMP: The first 25 years of a universal bacterial second messenger. Microbiol Mol Biol Rev 77 : 1–52.
3. KrastevaPV, GiglioKM, SondermannH (2012) Sensing the messenger: The diverse ways that bacteria signal through c-di-GMP. Protein Sci 21 : 929–48.
4. SondermannH, ShikumaNJ, YildizFH (2012) You've come a long way: c-di-GMP signaling. Curr Opin Microbiol 15 : 140–6.
5. BoydCD, O'TooleGA (2012) Second messenger regulation of biofilm formation: breakthroughs in understanding c-di-GMP effector systems. Annu Rev Cell Dev Biol 28 : 439–62.
6. MillsE, PultzIS, KulasekaraHD, MillerSI (2011) The bacterial second messenger c-di-GMP: mechanisms of signalling. Cell Microbiol 13 : 1122–9.
7. TamayoR, PrattJT, CamilliA (2007) Roles of cyclic diguanylate in the regulation of bacterial pathogenesis. Annu Rev Microbiol 61 : 131–48.
8. KulesekaraH, LeeV, BrencicA, LiberatiN, UrbachJ, et al. (2006) Analysis of Pseudomonas aeruginosa diguanylate cyclases and phosphodiesterases reveals a role for bis-(3′-5′)-cyclic-GMP in virulence. Proc Natl Acad Sci USA 103 : 2839–44.
9. AhmadI, LamprokostopoulouA, Le GuyonS, StreckE, BarthelM, et al. (2011) Complex c-di-GMP signaling networks mediate transition between virulence properties and biofilm formation in Salmonella enterica serovar Typhimurium. PLoS One 6: e28351.
10. StarkeyM, HickmanJH, MaL, ZhangN, De LongS, et al. (2009) Pseudomonas aeruginosa rugose small colony variants have adaptations likely to promote persistence in the cystic fibrosis lung. J Bacteriol 191 : 3492–3503.
11. MaloneJG, JaegerT, SpanglerC, RitzD, SpangA, et al. (2010) YfiBNR mediates cyclic di-GMP dependent small colony variant formation and persistence in Pseudomonas aeruginosa. PLoS Pathog 6: e1000804.
12. ByrdMS, PangB, HongW, WaligoraEA, JuneauRA, et al. (2011) Direct evaluation of Pseudomonas aeruginosa biofilm mediators in a chronic infection model. Infect Immun 79 : 3087–95.
13. LaiTH, KumagaiY, HyodoM, HayakawaY, RikihisaY (2008) Anaplasma phagocytophilum PleC histidine kinase and PleD diguanylate cyclase two-component system and role of cyclic di-GMP in host-cell infection. J Bacteriol 191 : 693–700.
14. KumagaiY, MatsuoJ, HayakawaY, RikihisaY (2010) Cyclic di-GMP signaling regulates invasion by Ehrlichia chaffeensis of human monocytes. J Bacteriol 192 : 4122–33.
15. KumagaiY, MatsuoJ, ChengZ, HayakawaY, RikihisaY (2011) c-di-GMP signaling regulates intracellular aggregation, sessility, and growth of Ehrlichia chaffeensis. Infect Immun 79 : 3905–12.
16. LeviA, FolcherM, JenalU, ShumanHA (2011) Cyclic diguanylate signaling proteins control intracellular growth of Legionella pneumophila. mBio 2: e00316–10.
17. CossartP (2011) Illuminating the landscape of host-pathogen interactions with the bacterium Listeria monocytogenes. Proc Natl Acad Sci U S A 108 : 19484–91.
18. FreitagNE, PortGC, MinerMD (2009) Listeria monocytogenes - from saprophyte to intracellular pathogen. Nat Rev Microbiol 7 : 623–28.
19. DrevetsDA, BronzeMS (2008) Listeria monocytogenes: epidemiology, human disease, and mechanisms of brain invasion. FEMS Immunoi Med Microbiol 53 : 151–65.
20. AllerbergerF, WagnerM (2010) Listeriosis: a resurgent foodborne infection. Clin Microbiol Infection 16 : 16–23.
21. MoretroT, LangsrudS (2004) Listeria monocytogenes: biofilm formation and persistence in food-processing environments. Biofilms 1 : 107–21.
22. OrsiRH, BorowskyML, LauerP, YoungSK, NusbaumC, et al. (2008) Short-term genome evolution of Listeria monocytogenes in a non-controlled environment. BMC Genomics 9 : 539.
23. Centers for Disease Control and Prevention. Multistate Outbreak of Listeriosis Linked to Whole Cantaloupes from Jensen Farms, Colorado. http://www.cdc.gov/listeria/outbreaks/cantaloupes-jensen-farms/082712/index.html
24. McCollumJT, CronquistAB, SilkBJ, JacksonKA, O'ConnorKA, et al. (2013) Multistate outbreak of listeriosis associated with cantaloupe. N Engl J Med 369 : 944–53.
25. RameyBE, KoutsoudisM, von BodmanSB, FuquaC (2004) Biofilm formation in plant-microbe associations. Curr Opin Microbiol 7 : 602–09.
26. FlemmingHC, WingenderJ (2010) The biofilm matrix. Nat Rev Microbiol 8 : 623–33.
27. RenierS, HébraudM, DesvauxM (2011) Molecular biology of surface colonization by Listeria monocytogenes: an additional facet of an opportunistic Gram-positive foodborne pathogen. Environ Microbiol 13 : 835–50.
28. HoelzerK, PouillotR, DennisS (2012) Listeria monocytogenes growth dynamics on produce: A review of the available data for predictive modeling. Foodborne Pathog Dis 9 : 661–73.
29. BordeleauE, FortierLC, MalouinF, BurrusV (2011) c-di-GMP turn-over in Clostridium difficile is controlled by a plethora of diguanylate cyclases and phosphodiesterases. PLoS Genet 7: e1002039.
30. PurcellEB, McKeeRW, McBrideSM, WatersCM, TamayoR (2012) Cyclic diguanylate inversely regulates motility and aggregation in Clostridium difficile. J Bacteriol 194 : 3307–16.
31. ChenY, ChaiY, GuoJH, LosickR (2012) Evidence for cyclic di-GMP-mediated signaling in Bacillus subtilis. J Bacteriol 194 : 5080–90.
32. GaoX, MukherjeeS, MatthewsPM, HammadLA, KearnsDB, DannCE3rd (2013) Functional characterization of core components of the Bacillus subtilis c-di-GMP signaling pathway. J Bacteriol 195 : 4782–92.
33. HollandLM, O'DonnellST, RyjenkovDA, GomelskyL, SlaterSR, et al. (2008) A staphylococcal GGDEF domain protein regulates biofilm formation independently of c-di-GMP. J Bacteriol 190 : 5178–89.
34. SudarsanN, LeeER, WeinbergZ, MoyRH, KimJN, et al. (2008) Riboswitches in eubacteria sense the second messenger cyclic di-GMP. Science 321 : 411–13.
35. LeeER, BakerJL, WeinbergZ, SudarsanN, BreakerRR (2010) An allosteric self-splicing ribozyme triggered by a bacterial second messenger. Science 329 : 845–48.
36. PaulR, WeiserS, AmiotNC, ChanC, SchirmerT, et al. (2004) Cell cycle-dependent dynamic localization of a bacterial response regulator with a novel diguanylate cyclase output domain. Genes Dev 18 : 715–27.
37. RyjenkovDA, TarutinaM, MoskvinOV, GomelskyM (2005) Cyclic diguanylate is a ubiquitous signaling molecule in bacteria: Insights into the biochemistry of the GGDEF protein domain. J Bacteriol 187 : 1792–98.
38. SchmidtAJ, RyjenkovDA, GomelskyM (2005) The ubiquitous protein domain EAL is a cyclic diguanylate-specific phosphodiesterase: enzymatically active and inactive EAL domains. J Bacteriol 187 : 4774–81.
39. ChristenM, ChristenB, FolcherM, SchauerteA, JenalU (2005) Identification and characterization of a cyclic di-GMP-specific phosphodiesterase and its allosteric control by GTP. J Biol Chem 280 : 30829–37.
40. TamayoR, TischlerAD, CamilliA (2005) The EAL domain protein VieA is a cyclic diguanylate phosphodiesterase. J Biol Chem 280 : 33324–30.
41. RyanRP, FouhyY, LuceyJF, CrossmanLC, SpiroS, et al. (2006) Cell-cell signaling in Xanthomonas campestris involves an HD-GYP domain protein that functions in cyclic di-GMP turnover. Proc Natl Acad Sci USA 103 : 6712–17.
42. Punta M, Coggill PC, Eberhardt RY, Mistry J, Tate J, et al. (2012) The Pfam protein families database. Nucleic Acids Res 40(Database issue): D290–301.
43. ChanC, PaulR, SamorayD, AmiotNC, GieseB, et al. (2004) Structural basis of activity and allosteric control of diguanylate cyclase. Proc Natl Acad Sci USA 101 : 17084–89.
44. BarendsTR, HartmannE, GrieseJJ, BeitlichT, KirienkoNV, et al. (2009) Structure and mechanism of a bacterial ligh-regulated cyclic nucleotide phosphodiesterase. Nature 459 : 1015–18.
45. Toledo-AranaA, DussurgetO, NikitasG, SestoN, Guet-RevilletH, et al. (2009) The Listeria transcriptional landscape from saprophytism to virulence. Nature 459 : 950–6.
46. GoughJ, KarplusK, HugheyR, ChothiaC (2001) Assignment of homology to genome sequences using a library of Hidden Markov Models that represent all proteins of known structure. J Mol Biol 313 : 903–19.
47. ChristenB, ChristenM, PaulR, SchmidF, FolcherM, et al. (2006) Allosteric control of cyclic di-GMP signaling. J Biol Chem 281 : 32015–32024.
48. LeeVT, MatewishJM, KesslerJL, HyodoM, HayakawaY, LoryS (2007) A cyclic-di-GMP receptor required for bacterial exopolysaccharide production. Mol Microbiol 65 : 1474–84.
49. DuerigA, AbelS, FolcherM, NicollierM, SchwedeT, et al. (2009) Second messenger-mediated spatiotemporal control of protein degradation regulates bacterial cell cycle progression. Genes Dev 23 : 93–104.
50. SimmR, MorrM, KaderA, NimtzM, RömlingU (2004) GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol Microbiol 53 : 1123–34.
51. RyjenkovDA, SimmR, RömlingU, GomelskyM (2006) The PilZ domain is a receptor for the second messenger c-di-GMP: The PilZ domain protein YcgR controls motility in entrobacteria. J Biol Chem 281 : 30310–14.
52. GirgisHS, LiuY, RyuWS, TavazoieS (2007) A comprehensive genetic characterization of bacterial motility. PLoS Genet 3 : 1644–60.
53. HobleyL, FungRK, LambertC, HarrisMA, DabhiJM, et al. (2012) Discrete cyclic di-GMP-dependent control of bacterial predation versus axenic growth in Bdellovibrio bacteriovorus. PLoS Pathog 8: e1002493.
54. PeelM, DonachieW, ShawA (1988) Temperature-dependent expression of flagella of Listeria monocytogenes studied by electron microscopy, SDS-PAGE and western blotting. J Gen Microbiol 134 : 2171–78.
55. TiensuuT, AnderssonC, RydénP, JohanssonJ (2013) Cycles of light and dark co-ordinate reversible colony differentiation in Listeria monocytogenes. Mol Microbiol 87 : 909–24.
56. YildizFH, SchoolnikGK (1999) Vibrio cholerae O1 El Tor: identification of a gene cluster required for the rugose colony type, exopolysaccharide production, chlorine resistance, and biofilm formation. Proc Natl Acad Sci U S A 96 : 4028–33.
57. ZogajX, NimtzM, RohdeM, BokranzW, RömlingU (2001) The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol Microbiol 39 : 1452–63.
58. SpiersAJ, BohannonJ, GehrigSM, RaineyPB (2003) Biofilm formation at the air-liquid interface by the Pseudomonas fluorescens SBW25 wrinkly spreader requires an acetylated form of cellulose. Mol Microbiol 50 : 15–27.
59. FriedmanL, KolterR (2004) Two genetic loci produce distinct carbohydrate-rich structural components of the Pseudomonas aeruginosa biofilm matrix. J Bacteriol 186 : 4457–65.
60. MerighiM, LeeVT, HyodoM, HayakawaY, LoryS (2007) The second messenger bis-(3-5)-cyclic-GMP and its PilZ domain-containing receptor Alg44 are required for alginate biosynthesis in Pseudomonas aeruginosa. Mol Microbiol 65 : 876–95.
61. O'GaraJP (2007) ica and beyond: biofilm mechanisms and regulation in Staphylococcus epidermidis and Staphylococcus aureus. FEMS Microbiol Lett 270 : 179–88.
62. BobrovAG, KirillinaO, PerryRD (2007) Regulation of biofilm formation in Yersinia pestis. Adv Exp Med Biol 603 : 201–10.
63. RömlingU (2002) Molecular biology of cellulose production in bacteria. Res Microbiol 153 : 205–12.
64. MorganJL, StrumilloJ, ZimmerJ (2013) Crystallographic snapshot of cellulose synthesis and membrane translocation. Nature 493 : 181–6.
65. O'TooleGA, KolterR (1998) Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: a genetic analysis. Mol Microbiol 28 : 449–61.
66. PanYJr, BreditF, KathariouS (2006) Resistance of Listeria monocytogenes biofilms to sanitizing agents in a simulated food processing environment. Appl Environ Microbiol 72 : 7711–7.
67. Bou GhanemEN, JonesGS, Myers-MoralesT, PatilPD, HidayatullahAN, D'OrazioSEF (2012) InlA promotes dissemination of Listeria monocytogenes to the mesenteric lymph nodes during food borne infection of mice. PLoS Pathog 8 e1003015.
68. ItohY, WangX, HinnebuschBJ, PrestonJF3rd, RomeoT (2005) Depolymerization of beta-1,6-N-acetyl-D-glucosamine disrupts the integrity of diverse bacterial biofilms. J Bacteriol 187 : 382–7.
69. SteinerS, LoriC, BoehmA, JenalU (2013) Allosteric activation of exopolysaccharide synthesis through cyclic di-GMP-stimulated protein-protein interaction. EMBO J 32 : 354–68.
70. WhitneyJC, HowellPL (2013) Synthase-dependent exopolysaccharide secretion in Gram-negative bacteria. Trends Microbiol 21 : 63–72.
71. PaulK, NietoV, CarlquistWC, BlairDF, HarsheyRM (2010) The c-di-GMP binding protein YcgR controls flagellar motor direction and speed to affect chemotaxis by a “backstop brake” mechanism. Mol Cell 38 : 128–39.
72. FangX, GomelskyM (2010) A posttranslational, c-di-GMP-dependent mechanism regulating bacterial flagellar motility. Mol Microbiol 76 : 1295–1305.
73. WolfeAJ, BergHC (1989) Migration of bacteria in semisolid agar. Proc Natl Acad Sci USA 86 : 6973–77.
74. BoehmA, KaiserM, LiH, SpanglerC, KasperCA, et al. (2010) Second messenger-mediated adjustment of bacterial swimming velocity. Cell 141 : 107–16.
75. ChristenM, ChristenB, AllanMG, FolcherM, JenoP, et al. (2007) DgrA is a member of a new family of cyclic diguanosine monophosphate receptors and controls flagellar motor function in Caulobacter crescentus. Proc Natl Acad Sci U S A 104 : 4112–17.
76. BlairKM, TurnerL, WinkelmanJT, BergHC, KearnsDB (2008) A molecular clutch disables flagella in the Bacillus subtilis biofilm. Science 320 : 1636–8.
77. GuttenplanSB, BlairKM, KearnsDB (2010) The EpsE flagellar clutch is bifunctional and synergizes with EPS biosynthesis to promote Bacillus subtilis biofilm formation. PLoS Genet 6: e1001243.
78. ZorraquinoV, GarcíaB, LatasaC, EcheverzM, Toledo-AranaA, et al. (2013) Coordinated cyclic-di-GMP repression of Salmonella motility through YcgR and cellulose. J Bacteriol 195 : 417–28.
79. CorrS, HillC, GahanCG (2006) An in vitro cell-culture model demonstrates internalin - and hemolysin-independent translocation of Listeria monocytogenes across M cells. Microb Pathog 41 : 241–50.
80. JensenVB, HartyJT, JonesBD (1998) Interactions of the invasive pathogens Salmonella typhimurium, Listeria monocytogenes, and Shigella flexneri with M cells and murine Peyer's patches. Infect Immun 66 : 3758–66.
81. MarcoAJ, AltimiraJ, PratsN, LópezS, DominguezL, et al. (1997) Penetration of Listeria monocytogenes in mice infected by the oral route. Microb Pathog 23 : 255–63.
82. NiessJH, BrandS, GuX, LandsmanL, JungS, et al. (2005) CX3CR1-mediated dendritic cell access to the intestinal lumen and bacterial clearance. Science 307 : 254–8.
83. RescignoM, UrbanoM, ValzasinaB, FrancoliniM, RottaG, et al. (2001) Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat Immunol 2 : 361–7.
84. WoodwardJJ, IavaroneAT, PortnoyDA (2010) c-di-AMP secreted by intracellular Listeria monocytogenes activates a host type I interferon response. Science 328 : 1703–05.
85. SauerJD, Sotelo-TrohaK, von MoltkeJ, MonroeKM, RaeCS, et al. (2011) The N-ethyl-N-nitrosourea-induced Goldenticket mouse mutant reveals an essential function of STING in the in vivo interferon response to Listeria monocytogenes and cyclic dinucleotides. Infect Immun 79 : 688–94.
86. BurdetteDL, MonroeKM, Sotelo-TrohaK, IwigJS, EckertB, et al. (2011) STING is a direct innate immune sensor of cyclic di-GMP. Nature 478 : 515–8.
87. RayamajhiM, HumannJ, PenheiterK, AndreasenK, LenzLL (2010) Induction of IFN-alphabeta enables Listeria monocytogenes to suppress macrophage activation by IFN-gamma. J Exp Med 207 : 327–37.
88. SchwartzKT, CarletonJD, QuillinSJ, RollinsSD, PortnoyDA, LeberJH (2012) Hyperinduction of host beta interferon by a Listeria monocytogenes strain naturally overexpressing the multidrug efflux pump MdrT. Infect Immun 80 : 1537–45.
89. TsaiHN, HodgsonDA (2003) Development of a synthetic minimal medium for Listeria monocytogenes. Appl Environ Microbiol 69 : 6943–45.
90. SmithK, YoungmanP (1992) Use of a new integrational vector to investigate compartment-specific expression of the Bacillus subtilis spoIIM gene. Biochimie 74 : 705–11.
91. ParkSF, StewartGS (1990) High-efficiency transformation of Listeria monocytogenes by electroporation of penicillin-treated cells. Gene 94 : 129–32.
92. LemonKP, HigginsDE, KolterR (2007) Flagellar motility is critical for Listeria monocytogenes biofilm formation. J Bacteriol 189 : 4418–24.
93. JaradatZW, BhuniaAK (2003) Adhesion, invasion and translocation characteristics of Listeria monocytogenes serotypes in Caco-2 cell mouse models. Appl Environ Microbiol 69 : 3640–45.
94. MostowyS, DanckaertA, ThamTN, MachuC, GuadagniniS, et al. (2009) Septin 11 restricts InlB-mediated invasion by Listeria. J Biol Chem 284 : 11613–21.
95. RyuJ-H, BeuchatLR (2005) Biofilm formation by Esherichia coli O157:H7 on stainless steel: Effect of exopolysaccharide and curli production on its resistance to chlorine. Appl Environ Microbiol 71 : 247–54.
96. BalestrinoD, HamonMA, DortetL, NahoriMA, Pizarro-CerdaJ, et al. (2010) Single-cell techniques using chromosomally tagged fluorescent bacteria to study Listeria monocytogenes infection processes. Appl Environ Microbiol 76 : 3625–36.
97. MonkIR, GahanCG, HillC (2008) Tools for functional postgenomic analysis of Listeria monocytogenes. Appl Environ Microbiol 74 : 3921–34.
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2014 Číslo 8
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- Disruption of Fas-Fas Ligand Signaling, Apoptosis, and Innate Immunity by Bacterial Pathogens
- Ly6C Monocyte Recruitment Is Responsible for Th2 Associated Host-Protective Macrophage Accumulation in Liver Inflammation due to Schistosomiasis
- Host Responses to Group A Streptococcus: Cell Death and Inflammation
- Pathogenicity and Epithelial Immunity
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy