
Expression of Genes Involves Exchange of the Histone Variant H2A.Z at the Promoter
Plasmodium falciparum employs antigenic variation to evade the human immune response by switching the expression of different variant surface antigens encoded by the var gene family. Epigenetic mechanisms including histone modifications and sub-nuclear compartmentalization contribute to transcriptional regulation in the malaria parasite, in particular to control antigenic variation. Another mechanism of epigenetic control is the exchange of canonical histones with alternative variants to generate functionally specialized chromatin domains. Here we demonstrate that the alternative histone PfH2A.Z is associated with the epigenetic regulation of var genes. In many eukaryotic organisms the histone variant H2A.Z mediates an open chromatin structure at promoters and facilitates diverse levels of regulation, including transcriptional activation. Throughout the asexual, intraerythrocytic lifecycle of P. falciparum we found that the P. falciparum ortholog of H2A.Z (PfH2A.Z) colocalizes with histone modifications that are characteristic of transcriptionally-permissive euchromatin, but not with markers of heterochromatin. Consistent with this finding, antibodies to PfH2A.Z co-precipitate the permissive modification H3K4me3. By chromatin-immunoprecipitation we show that PfH2A.Z is enriched in nucleosomes around the transcription start site (TSS) in both transcriptionally active and silent stage-specific genes. In var genes, however, PfH2A.Z is enriched at the TSS only during active transcription in ring stage parasites. Thus, in contrast to other genes, temporal var gene regulation involves histone variant exchange at promoter nucleosomes. Sir2 histone deacetylases are important for var gene silencing and their yeast ortholog antagonises H2A.Z function in subtelomeric yeast genes. In immature P. falciparum parasites lacking Sir2A or Sir2B high var transcription levels correlate with enrichment of PfH2A.Z at the TSS. As Sir2A knock out parasites mature the var genes are silenced, but PfH2A.Z remains enriched at the TSS of var genes; in contrast, PfH2A.Z is lost from the TSS of de-repressed var genes in mature Sir2B knock out parasites. This result indicates that PfH2A.Z occupancy at the active var promoter is antagonized by PfSir2A during the intraerythrocytic life cycle. We conclude that PfH2A.Z contributes to the nucleosome architecture at promoters and is regulated dynamically in active var genes.
Published in the journal:
. PLoS Pathog 7(2): e32767. doi:10.1371/journal.ppat.1001292
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1001292
Summary
Plasmodium falciparum employs antigenic variation to evade the human immune response by switching the expression of different variant surface antigens encoded by the var gene family. Epigenetic mechanisms including histone modifications and sub-nuclear compartmentalization contribute to transcriptional regulation in the malaria parasite, in particular to control antigenic variation. Another mechanism of epigenetic control is the exchange of canonical histones with alternative variants to generate functionally specialized chromatin domains. Here we demonstrate that the alternative histone PfH2A.Z is associated with the epigenetic regulation of var genes. In many eukaryotic organisms the histone variant H2A.Z mediates an open chromatin structure at promoters and facilitates diverse levels of regulation, including transcriptional activation. Throughout the asexual, intraerythrocytic lifecycle of P. falciparum we found that the P. falciparum ortholog of H2A.Z (PfH2A.Z) colocalizes with histone modifications that are characteristic of transcriptionally-permissive euchromatin, but not with markers of heterochromatin. Consistent with this finding, antibodies to PfH2A.Z co-precipitate the permissive modification H3K4me3. By chromatin-immunoprecipitation we show that PfH2A.Z is enriched in nucleosomes around the transcription start site (TSS) in both transcriptionally active and silent stage-specific genes. In var genes, however, PfH2A.Z is enriched at the TSS only during active transcription in ring stage parasites. Thus, in contrast to other genes, temporal var gene regulation involves histone variant exchange at promoter nucleosomes. Sir2 histone deacetylases are important for var gene silencing and their yeast ortholog antagonises H2A.Z function in subtelomeric yeast genes. In immature P. falciparum parasites lacking Sir2A or Sir2B high var transcription levels correlate with enrichment of PfH2A.Z at the TSS. As Sir2A knock out parasites mature the var genes are silenced, but PfH2A.Z remains enriched at the TSS of var genes; in contrast, PfH2A.Z is lost from the TSS of de-repressed var genes in mature Sir2B knock out parasites. This result indicates that PfH2A.Z occupancy at the active var promoter is antagonized by PfSir2A during the intraerythrocytic life cycle. We conclude that PfH2A.Z contributes to the nucleosome architecture at promoters and is regulated dynamically in active var genes.
Introduction
Plasmodium falciparum causes the majority of malaria-induced morbidity and mortality, resulting in approximately 860,000 deaths each year [1]. Plasmodium parasites have a complicated life cycle, during which they differentiate into several morphologically distinct asexual and sexual stages in the human host and the mosquito vector.
Disease occurs during the repeated cycles of invasion and asexual replication of the parasite inside human erythrocytes. A central mechanism of malaria pathogenesis is the ability of the infected erythrocytes (IE) to sequester at vascular sites by cytoadherence to host receptors. Through this process the parasite avoids clearance by splenic macrophages and contributes to severe malaria complications such as cerebral and placental malaria [2]. Sequestration is mediated by the P. falciparum erythrocyte membrane protein 1 (PfEMP1) variant antigens, which are expressed on the IE surface and are encoded by approximately 60 genes of the var multigene family [3], [4], [5], [6]. Only a single var gene is expressed at a time [7], [8] and switching between expression of different PfEMP1 variants alters both the cytoadherence phenotype and the antigenic profile of the IE, resulting in antigenic variation [9]. These processes are critical to immune evasion of P. falciparum and chronic infection [10], [11], [12].
During its asexual, intaerythrocytic life cycle, P. falciparum employs stringent regulatory mechanisms to achieve stage-specific gene expression [13], [14]. The multiple layers of regulation that mediate morphological and physiological adaptations, include specific transcription factors and repressors [15], [16], [17], [18], translational repression [19], [20], post-translational protein modifications (reviewed [21]) and epigenetic mechanisms [22], [23]. Recent studies assessing the global structure of chromatin during the asexual intra-erythrocytic developmental cycle (IDC) have shown that chromatin undergoes massive reorganization in P. falciparum, emphasizing the significance of epigenetic control in the parasite [24].
Epigenetic gene regulation confers a heritable state of gene expression and is typically mediated by changes in chromatin structure without a change in DNA sequence. The fundamental units of chromatin are nucleosomes and are formed by ∼146 bp of DNA wrapping around an octamer of histones. The canonical nucleosome components are two H2A/H2B dimers and an H3/H4 tetramer. In most eukaruyotes an additional subunit, H1, links nucleosomes, but this subunit is missing in Plasmodium species. Chromatin can exist as compact, silent heterochromatin and as open, transcriptionally competent euchromatin. These different physical states are essentially determined by the composition and distribution of nucleosomes, post-translational modification of histones, presence of chromatin associated trans-factors and by the covalent modification of DNA, although this latter mechanism has not yet been shown in Plasmodium. The Plasmodium genome encodes a broad set of common chromatin remodelling and modifying factors, including many that are novel and that may be employed in unique epigenetic mechanisms [25], [26], [27].
The organisation of P. falciparum chromatin has unique features that differ markedly from other eukaryotes. P. falciparum intergenic regions, including promoters, display a global nucleosome depletion [24], [28]. Consequently, and unlike in other eukaryotes, nucleosome occupancy at P. falciparum promoters does not correlate inversely with steady-state mRNA levels [29], [30], [31]. Genome-wide analysis of histone modifications has shown that the blood stage genome of P. falciparum exists in an unusually euchromatic state conferred by the euchromatin marks trimethylated lysine 4 of histone 3 (H3K4me3) and acetylated lysine 9 of histone 3 (H3K9ac) [22], [32]. In contrast to the situation in yeast and other eukaryotes, the presence of neither H3K4me3 nor H3K9ac enrichment seems to correlate with gene activity throughout most of the intraerythrocytic cycle, except for late stage schizont parasites when these marks are enriched at the 5′ ends of active genes [22]. P. falciparum heterochromatin, defined by the histone post-translational modification H3K9me3 and its cognate trans-factor heterochromatin protein 1 (HP1), is restricted to subtelomeric and several chromosome internal domains in P. falciparum that contain gene families including the majority of var genes [22], [33], [34], [35], [36].
The var genes present both in subtelomeric and central chromosomal positions form clusters at the nuclear periphery [37], [38], [39] and expression of a var gene appears to require it to leave the heterochromatic cluster and relocate to a specific perinuclear expression site [35], [38], [40], [41]. The variegated, monoallelic expression of var genes is controlled by epigenetic mechanisms [42]. To become activated in ring stages, a var gene must acquire the histone marks H3K4me3 and H3K9ac in its promoter [34], [35]. The var promoter is the only cis element required for monoallelic exclusive transcription [8] but promoter activity of the var gene intron is important for var gene silencing [43], [44], [45], [46] and additional cis sequence elements contribute to the rate of var gene switching [47]. A critical role of histone acetylation in var gene regulation was proven by the upregulation of numerous var genes and the loss of monoallelic var gene expression that occurred when either of two Sir2 histone deacetylase genes were disrupted [41], [48].
Alternative histones can replace canonical histones through ATP-dependent deposition to create structurally and functionally specialized chromatin domains [49]. While the importance of histone modifications to P. falciparum gene regulation is apparent, the role of alternative histones has not yet been investigated. H2A.Z is an H2A variant, which is essential for viability in most organisms apart from yeast and has been highly conserved through evolution [50], [51], [52], [53]. H2A.Z has been implicated in the regulation of very diverse processes such as heterochromatin formation, chromosome stability and segregation, proliferation and transcriptional activation or repression [49]. Its role in transcriptional regulation has been clearly established across species, but the underlying mechanisms are not well understood and reported effects of H2A.Z deposition are contradictory. Genome wide analyses consistently revealed that H2A.Z is enriched in nucleosomes near the transcriptional start site (TSS) in RNA polymerase II promoters [54], [55]. In human cells and in the protist parasite Toxoplasma gondii, H2A.Z enrichment at promoters (and enhancers in humans) correlates with gene activity [54], [56], whereas in yeast H2A.Z was found to occupy the promoters of active as well as poised genes from where it is lost with active transcription [55], [57].
In this study, we provide the first characterization of the alternative histone H2A.Z in P. falciparum. Using immunofluoresecence analysis (IFA), co-immunoprecipitation (Co-IP) and chromatin immunoprecipitation (ChIP) in conjunction with quantitative real time PCR (qPCR), we show that PfH2A.Z is enriched in the promoter of a set of developmentally regulated genes in the euchromatin compartment. In these genes promoter occupancy of PfH2A.Z does not correlate with transcription levels, suggesting that presence of PfH2A.Z is important for providing a transcriptionally competent chromatin structure at the promoter but does not directly relate to promoter activity. In var genes by contrast, PfH2A.Z promoter occupancy is strongly associated with transcriptional activity, as PfH2AZ is periodically enriched in the active var gene and is depleted in silent var genes, which is consistent with our IFA evidence that H2A.Z is absent from the subtelomeric heterochromatin compartment. This balance is distorted in cells in which the histone deacetylase Sir2A has been disrupted. Thus, these data suggest that var gene silencing requires expulsion of PfH2A.Z from the var promoter and that this involves histone deacetylation.
Results
The P. falciparum H2A.Z variant
The P. falciparum H2A.Z variant (PfH2A.Z) is encoded by the gene PFC0920w [58]. Sequence alignments show that PfH2A.Z shares 56.4% amino acid identity with the S. cerevisiae H2A.Z variant Htz1 and 67.3% identity with the human and mouse H2A.Z proteins (Figure S1), whereas conservation between other organisms was reported to be 70%–90% [59]. The major divergences affect the extended and highly charged N-terminus of PfH2A.Z, which is characterized by an accumulation of lysine residues and a three-fold repetition of the peptide sequence GGKV (position 9–20). Mass spectrometric evidence indicates that the N-terminus of PfH2A.Z can be acetylated in seven lysine residues, which would partially neutralize the positive charge [60]. In loop 1, two threonine residues implicated in mediating H2A.Z/H2A.Z self-interactions and prohibiting dimerization with H2A [61] are changed to isoleucine and serine (PfH2A.Z Ile 65 and Ser 66), respectively, and a conserved positively charged residue is substituted by aspartic acid (PfH2A.Z Asp 68) introducing a negative charge in this region. In the C-terminal docking domain all amino acids critical for interaction with the H3/H4 dimer are conserved [61].
Developmental expression of PfH2A.Z during the asexual life cycle
To analyse PfH2A.Z, antiserum against recombinant full length PfH2A.Z was generated. The serum specifically recognizes PfH2A.Z but not H2A and cross-reacts with human H2A.Z, which has a different N terminal acetylation pattern, indicating reactivity of the antibody with the non-acetylated, conserved C-terminal domains (Figure 1A). We further showed that anti-PfH2A.Z immunoprecipitates both acetylated and non-acetylated forms of PfH2A.Z because antibodies specific for an acetylated peptide present in the PfH2A.Z and H4 N-termini [61] labels immunoprecipitated PfH2A.Z by Western Blot (Figure 1B).

We found that PfH2A.Z is present throughout the asexual life cycle (Figure 1C & D). In comparison to the canonical histone H3, PfH2A.Z abundance increases significantly in the schizont stage (Figure 1C). This is consistent with an increase in PfH2A.Z mRNA observed in schizonts [58]. We confirmed that PfH2A.Z is localized in the nucleus in all asexual stages by immunofluorescence analyses (IFA) and confocal microscopy. An area stained with the DNA dye DAPI but devoid of PfH2A.Z labelling was consistently observed, indicating that PfH2A.Z is enriched towards one side of the nucleus (Figure 1D). 3D reconstruction verified this polarized localization of PfH2A.Z (Video S1) and transgenic P. falciparum ectopically expressing PfH2A.Z-GFP fusion proteins corroborated the sub-nuclear distribution of PfH2A.Z (Figure S2). A similar cap-like pattern has previously been reported for the euchromatic histone mark H3K4me3 [62].
PfH2A.Z is present in the euchromatin compartment
H2A.Z has been shown to contribute to diverse biological processes associated with different chromatin compartments, such as gene activation and poising [63], [64], [65], [66], chromosome segregation [67], [68] and heterochromatin structure [69], [70], [71], [72], [73]. To investigate the chromatin association of PfH2A.Z we performed co-localization experiments with well-characterized chromatin marks. Double staining showed good overlap between PfH2A.Z and the euchromatin marks H3K4me3 and H3K9ac (Figure 2A), which are enriched across the P. falciparum genome [22]. In contrast, PfH2A.Z staining was distinct from the subtelomeric heterochromatin marks H3K9me3 and HP1 (Figure 2A). Consistent with these results, immunoelectron microscopy indicated that PfH2A.Z was not restricted to the nuclear periphery, where inactive subtelomeric and internal var genes cluster. However, its distribution appeared concentrated in certain subnuclear compartments, frequently at the border of the electron lucent and electron dense nuclear material that is presumed to represent euchromatin and heterochromatin, respectively (Figure 2B) [38].

In mammalian cells it was shown that H2A.Z-containing nucleosomes preferentially carry the euchromatic mark H3K4me3 [73], [74]. Consistent with this, we found by co-immunoprecipitation experiments with anti-PfH2A.Z that mononucleosomes containing PfH2A.Z are highly enriched in H3K4me3 (Figure 2C). Together, these data support that PfH2A.Z is functionally linked to euchromatin but is largely depleted from subtelomeric heterochromatin.
PfH2A.Z is enriched near the transcription start site (TSS) of genes irrespective of transcriptional status
To identify the genomic target sites of PfH2A.Z and investigate how the presence of PfH2A.Z correlates with transcriptional activity, we performed chromatin immunoprecipitation (ChIP) followed by quantitative PCR in ring stage parasites, trophozoites and schizonts. H2A.Z is enriched in nucleosomes surrounding the transcription start site (TSS) in other organisms, therefore at least two quantitative (q) PCR reactions were performed for each gene to amplify regions near the predicted TSS upstream of the start codon (ups) as well as in the open reading frame (orf).
Our results show that PfH2A.Z is enriched in all three stages in the promoter regions of candidate genes that are differentially regulated throughout the life cycle (Figure 3A, S3A). In contrast to PfH2A.Z, H2A showed equal distribution in the promoter and the open reading frame of the investigated genes (Figure 3B). In rings, promoter enrichment of PfH2A.Z was highest in genes that are constitutively expressed (e.g. HSP70 and casein kinase) or induced during asexual intra-erythrocytic differentiation (e.g. schizont genes MSP2 and Eba175), but also clearly apparent in silent genes (e.g. sporozoite genes CSP and SSP2). In trophozoites and schizonts, PfH2A.Z enrichment in the promoter showed similar levels across all genes.

To determine whether the level of PfH2A.Z promoter occupancy in the examined genes correlated with gene expression, mRNA levels were quantified by q-RT-PCR (Figure S4). The ups/orf ratio of PfH2A.Z enrichment was determined and plotted against the relative expression levels. No significant correlation between PfH2A.Z promoter occupancy and transcription level could be observed at any stage (Spearman correlation, p>0.5).
To further investigate the relationship between PfH2A.Z and euchromatic and heterochromatic histone marks, ChIP was performed in parallel for PfH2A.Z as well as H3K4me3, H3K9ac and H3K9me3. PfH2A.Z and the two euchromatic marks, H3K4me3, H3K9ac all showed significant enrichment in the ups region when compared to the orf in ring and schizont stage parasites, in contrast the levels of the heterochromatic mark H3K9me3 were the same in ups and orf (Figure S5). This is consistent with previous work on the histone marks [22] and supports our finding that PfH2A.Z and H3K4me3 are present in the same nucleosomes (Figure 2C). Furthermore, we found that the PfH2A.Z enrichment level in the upstream region of genes positively correlates with both euchromatic marks (P<0.0001), whereas a negative correlation was evident between PfH2A.Z and H3K9me3 (p<0.0001) (Figure S6).
Together, these results demonstrate that PfH2A.Z is enriched near the TSS in genes, independently of their transcriptional activity, and that PfH2A.Z enrichment near the TSS correlates with enrichment of the euchromatin marks H3K4me3 and H3K9ac.
PfH2A.Z enrichment in the var gene promoter is developmentally regulated
The P. falciparum histone deacetylases Sir2A and Sir2B silence subtelomeric and central var genes [41], [48], [75] and in S. cerevisiae H2A.Z antagonises subtelomeric gene silencing by Sir2 [53]. Therefore we investigated whether PfH2A.Z was involved in the transcriptional control of var genes. We harvested chromatin and RNA from a parasite culture that had been selected for the expression of a single var gene encoding VAR2CSA by panning on chondroitin sulphate A (CSA). To map the position of PfH2A.Z along the var2csa gene, we designed seven qPCR reactions spanning this var locus. The TSS of var2csa has previously been mapped to −1475 bp upstream of the start codon in the FCR3 parasite line [34] and is predicted to be located at approximately −1200 bp in 3D7 (PlasmoDB). Three primer pairs amplified regions in the non-coding upstream region (-1500 bp, −1000 bp, −575 bp), and four primer pairs targeted areas along the coding region.
ChIP analysis showed that in ring stage parasites, when var2csa transcription peaks (Figure S4) [76], PfH2A.Z is strongly enriched in the areas flanking the predicted TSS (−1500 and −1000 bp), but not further downstream (−575 bp) or within the open reading frame (ATG, DBL3, DBL6) (Figure 4A, S3B). As the parasites progress through the trophozoite to the schizont stage var transcription declines and PfH2A.Z enrichment around the TSS decreases. No enrichment of PfH2A.Z was detectable around the TSS of the var2csa gene at schizont stage. This stage-specific deposition of PfH2A.Z in the var gene promoter contrasts with the continuous presence of PfH2A.Z in the promoters of the limited number of other genes we analysed (Figure 3A) indicating that PfH2A.Z deposition differs in its temporal regulation in var genes.

To directly compare PfH2A.Z occupancy of an active and inactive var promoter we performed ChIP experiments on unselected 3D7 parasites that do not transcribe the var2csa gene (var2csa OFF), and var2csa expressing parasites (var2csa ON) at ring stage. Transcription levels were verified by q-RT-PCR (Figure S4). In contrast to the strong enrichment of PfH2A.Z in the active var promoter, the alternative histone was clearly not enriched when var2csa was not transcribed. These results demonstrate that PfH2A.Z promoter occupancy in var2csa strongly correlates with transcription (Figure 4B).
To verify our observation that PfH2A.Z occupancy is restricted to the active var TSS in a second var gene, 3D7 parasites were selected on ICAM1 and the gene PFL0020w was identified as the dominant transcript by q-RT-PCR and Northern Blot analysis (Figure S7 A, B). ChIP analysis across the gene confirmed increased PfH2A.Z occupancy near the PFL0020w TSS in ICAM1 selected parasites as compared to non-selected parasites at ring stage. In line with our results with var2csa, no enrichment was evident in schizonts (Figure S7 C, D).
PfH2A.Z is enriched in the var intron
Var genes possess a second promoter, which is situated in the conserved intron. From this promoter, truncated sense and antisense transcripts are synthesized which are thought to contribute to heterochromatin structure and var gene silencing [77]. To investigate a possible association of PfH2A.Z with the var intron promoter, we designed primer pairs targeting the var introns and repeated the ChIP experiment in var2csa expressing and non-expressing ring stage parasites (Figure 4C). Consistent with the previous observations, PfH2A.Z was enriched around the TSS of the active var2csa gene (var2csa ON), but not the TSS of inactive var2csa (var2csa OFF) nor the TSS of the other two silent var genes PF08_0141 (var41) and PFL0020w (var20). In contrast to the upstream region, all three var genes were moderately enriched in PfH2A.Z in the introns (Figure 4C). This was also evident in the ICAM selected parasite line (Figure S7 C&D). Analysis of intron/orf pairs from ten different var genes confirmed that the enrichment of PfH2A.Z in the intron was statistically significant (Figure 4D).
PfH2A.Z deposition is maintained at the promoters of active var genes in mature Sir2AKO, but not Sir2BKO parasites
Because H2A.Z may function as a barrier to prevent the spread of Sir2-mediated silencing in yeast [53] we further investigated the relationship between Sir2 and PfH2A.Z in the control of var gene expression in P. falciparum. ChIP and expression analyses were performed on ring and schizont stage parasites in which Sir2A or Sir2B had been disrupted (3D7Δsir2A and 3D7Δsir2B) [41], [48]. By q-RT-PCR we first monitored the expression profiles of all var genes in ring stages of both 3D7Δsir2 parasite lines (Figure S8). With the aim to understand how each Sir2 paralogue influences PfH2A.Z deposition and how this correlates with var transcription, we selected five var genes that were highly expressed and five var genes that were lowly expressed in 3D7Δsir2A parasites for further analysis, all of which had previously been shown to be regulated by Sir2A [41], [48]. We used the same strategy to choose ten var genes previously shown to be regulated by Sir2B [48] for analysis in 3D7Δsir2B. We then analysed PfH2A.Z deposition by ChIP and qPCR in upstream and coding regions of these var genes in knock out and wild type parasites. The ups/orf ratios were determined and compared between 3D7Δsir2A or 3D7Δsir2B and 3D7 parasites, respectively (Figure 5).

Although ups/orf ratios were generally quite low in ring stages, PfH2A.Z enrichment at var promoters was significantly greater in the highly expressed var genes in the 3D7Δsir2A and B lines than in the 3D7 control (Mann-Whitney, p = 0.0232 (Δsir2A) and p = 0.0152 (Δsir2B)) (Figure 5A & C). In schizonts, no enrichment of PfH2A.Z upstream of active Sir2B regulated genes was detected (Figure 5D). This result is consistent with a role of PfH2A.Z in active transcription of var genes. However, interestingly PfH2A.Z occupancy in the upstream region of var genes was maintained and even elevated at schizont stage in 3D7Δsir2A parasites at significantly higher levels than in 3D7 (Mann-Whitney, p = 0.0159) (Figure 5B), although var gene expression is down-regulated in mature 3D7Δsir2 parasites (data not shown) [48]. This result indicates a link between Sir2A and loss of PfH2A.Z and implicates PfH2A.Z with maintenance of the integrity of the heterochromatin/euchromatin boundary at Sir2A regulated loci.
To further investigate the atypical PfH2A.Z enrichment in Sir2A regulated var genes in 3D7Δsir2A schizonts we analysed by ChIP the relationship between enrichment of PfH2A.Z and H3K9me3 or H3K4me3, respectively (Figure S9). Similar to actively transcribed var genes in immature wildtype parasites (Figure S6), we found a positive correlation between PfH2A.Z and H3K4me3 enrichment (Spearman correlation, p = 0.0005) and a.negative correlation between PfH2A.Z and H3K9me3 (Spearman correlation, p = 0.0182), which was consistent with the previously described depletion of H3K9me3 in var genes upregulated in 3D7Δsir2A parasites [35].
Discussion
The P. falciparum H2A.Z ortholog
H2A.Z is essential in many eukaryotes including Trypanosoma brucei, Tetrahymena thermophilus, Drosophila, Xenopus, and vertebrates [50], [52], [59], [78], [79] and is one of the structurally most conserved histones throughout evolution [80]. However, Plasmodium H2A.Z differs significantly from its orthologues, particularly in its extended N-terminus which contains seven lysine residues that can be acetylated [58], [60], as opposed to five in humans and four in yeast. In T. thermophilus the N-terminus of H2A.Z has been implicated in directly interacting with the DNA [81]. Acetylation of at least one lysine residue is essential for viability, and probably acts by reducing the positive charge and thereby weakening H2A.Z–DNA interactions [82]. Interestingly, T. thermophilus H2A.Z encodes a repeated GGK motif similar to the one observed in PfH2A.Z, suggesting that a similar mechanism may apply for regulating PfH2A.Z-DNA interactions in the malaria parasite.
By western blot we observed increased PfH2A.Z abundance in the schizont stage relative to H3. Interestingly, PfH2A.Z accumulation coincides with expression of the ATP-dependent chromatin remodeling factor PfSwr1 (unpublished data), orthologues of which mediate the post-replicative H2A.Z incorporation into nucleosomes in yeast and humans [83], [84], [85]. The Plasmodium genome becomes densely packed with nucleosomes in the late schizont stages [24], and the concurrent increase in PfH2A.Z may reflect a rising requirement for the alternative histone in the intergenic regions. H2A.Z facilitates intra-molecular folding of nucleosomal arrays into a 30 nm fibre [86], so it may play a role in chromatin condensation in the mature schizonts.
PfH2A.Z and genes in the euchromatin compartment
Enrichment of H2A.Z in RNA polymerase II promoters has been conserved through the evolution of eukaryotes as diverse as yeast, humans and the protist parasites T. brucei and T. gondii [29], [53], [54], [55], [56], [57], [78], [87], [88]. Our ChIP analysis showed that P. falciparum conforms to this pattern, with an enrichment of PfH2A.Z, but not H2A, in the upstream regions of genes (Figures 3, 4). Consistent with this finding, we co-precipitated PfH2A.Z and H3K4me3 (Figure 2), which is enriched in 5′-upstream regions of several organisms including P. falciparum [22], [54], [89]. Our study also indicates that the pattern of PfH2A.Z occupancy at euchromatic gene promoters remains relatively stable throughout the IDC (Figure 3), in contrast to the reported fluctuations in levels of the transcriptionally permissive histone modifications H3K4me3 and H3K9ac [22]. Further substantiating an association of PfH2A.Z with promoter architecture, we showed that PfH2A.Z occupation in upstream regions correlates with H3K4me3 and H3K9ac enrichment (Figures S5 & 6).
With the exception of var genes, PfH2A.Z enrichment did not correlate with mRNA levels in our experiments. A global enrichment of H2A.Z at active and inactive promoters has also been observed in yeast [55], whereas some other studies reported a negative correlation with transcription and proposed that H2A.Z poises inducible silent genes for activation and is subsequently evicted during transcription [29], [57], [88]. In humans, H2A.Z is either present in active gene promoters in differentiated cells [54], [90] or marks poised gene promoters in hematopoietic stem cells [91]. These conflicting results demonstrate that the role of H2A.Z in transcription is very complex and the underlying mechanisms remain enigmatic.
In both yeast and humans H2A.Z assists in RNA polymerase II recruitment [90], [92]. Components of the preinitiation complex are pre-assembled in some erythrocytic stage P. falciparum promoters regardless of gene activity [93], similar to the PfH2A.Z enrichment at the TSS shown here. Thus PfH2A.Z may contribute to the open chromatin structure necessary for preinitiation complex formation. But how could PfH2A.Z modulate gene activity despite global occupancy at promoters? Mass spectrometric evidence that PfH2A.Z can be heavily acetylated at multiple lysine residues in the N-terminus suggests that the neutralization of positive charges by lysine acetylation may facilitate an open chromatin structure making the DNA more accessible [58], [60]. In our experiments, total PfH2A.Z occupancy was monitored because our antiserum did not differentiate between acetylated and non-acetylated forms of PfH2A.Z (Figure 1). Acetylation may thus provide a functional switch necessary to promote transcription, as has been suggested for yeast and humans [64], [66], [94], [95]. Histone acetylation normally promotes gene activation, whereas sumoylation and ubiquitination are post-translational modification associated with recruitment of histone deacetylase complexes and transcriptional repression [96], [97]. PfH2A.Z has six potential sumoylation sites [98] and may also be ubiquitinated in the C-terminal domain, although this has not yet been shown. In mammalian cells, mono-ubiquitinated H2A.Z is enriched in the facultative heterochromatin that constitutes the inactive X-chromosome [73]. Thus, acetylation, ubiquitination and sumoylation represent interesting candidate regulators of PfH2A.Z function at promoters.
In Arabidopsis thaliana and yeast it has recently been shown that H2A.Z mediates a thermo-sensory response and facilitates differentiation processes by regulating transcription [99]. This is thought to be due to reduced DNA wrapping of H2A.Z containing nucleosomes at higher temperatures, resulting in a relaxed chromatin structure that permits of transcription. It is tempting to speculate that PfH2A.Z may function as a similar physical switch to control gene expression in response to temperature change, for example during fever or as P. falciparum is transmitted between its two hosts.
Apart from PfH2A.Z, P. falciparum encodes two other histone variants, H2Bv and H3.3 [58], which have previously been implicated in transcriptional regulation. H2Bv is a variant unique to protist parasites and has so far only been characterized in T. brucei and T. gondii [56], [78], [87]. In both parasites H2Bv pairs with H2A.Z. While H2A.Z/H2Bv nucleosomes are enriched in the promoters of active genes in T. gondii [56], they showed a global association with PolII transcription start sites in T. brucei [87]. Nucleosomes containing H2A.Z and H3.3 have been reported to be less stable than nucleosomes containing H2A.Z and canonical H3 in humans [100], and nucleosomes composed of both variant histones are present in active promoters, enhancers and insulators. It was postulated that instability of H2A.Z/H3.3 composite nucleosomes at the TSS might facilitate access to transcription factors [101]. H3.3 differs only by three amino acids from canonical H3 so the antibodies directed against H3 in this study precipitated both variants. The functional cooperation of alternative histones in P. falciparum gene regulation will be an interesting field of future research.
PfH2A.Z and var genes
In many organisms, including Plasmodium, subtelomeric genes are subject to special mechanisms of gene regulation. This occurs via the unique subtelomeric heterochromatin which provides a specialized architecture to control variegated expression of gene family members (reviewed in [102], [103]). A link between H2A.Z and subtelomeric gene regulation has been established in S. cerevisiae as disruption of the H2A.Z orthologue htz1 results in the down regulation of many subtelomeric genes [53]. Consistent with its proposed role as an anti-silencing factor for subtelomeric genes in yeast, we found PfH2A.Z to be enriched in the subtelomeric var2csa and pfl0020w promoters when these genes were active, but not when they were silent (Figures 4, S7). This was supported by significant enrichment of PfH2A.Z in active var genes in the two Sir2 KO lines (Figure 5). Thus, PfH2A.Z is a novel component contributing to the promoter architecture of active var genes together with the euchromatin factors H3K4me3 and H3K9ac [34]. In contrast to the other genes we examined, var genes exhibited significant temporal modulation in PfH2A.Z enrichment at the promoter. Loss of PfH2A.Z from the var promoter is observed from the trophozoite stage on, which is when DNA replication begins, and thus is consistent with S-phase dependent silencing of var genes [43]. While canonical histones are deposited into chromatin at the replication fork, deposition of histone variants can occur post-replication by ATP-dependent enzyme complexes. This allows dynamic changes to the chromatin structure at the promoter during differentiation (reviewed in [49]). Based on this knowledge we speculate that PfH2A.Z is lost from the var promoter during DNA replication, possibly due to limited stability of PfH2A.Z containing nucleosomes, and is deposited at the var promoter in rings. The loss of PfH2A.Z from the active var gene promoter upon temporal silencing suggests that, unlike in subtelomeric genes in yeast [57], [104], PfH2AZ is not involved in epigenetic memory of var genes.
Our ChIP and qPCR experiments along the entire var gene revealed that PfH2A.Z enrichment peaks at positions directly surrounding the predicted TSS of the active var promoter. This pattern is consistent with yeast where H2A.Z is highly enriched in the two nucleosomes surrounding a nucleosome free-region at the TSS, and with humans where it is restricted to the two nucleosomes in the −1 and +1 position as well as at the TSS itself [55], [101]. The distinctive pattern of PfH2A.Z enrichment we observed within the var intron corroborates the existence of a promoter-like chromatin structure at this site. The var gene intron contains a bi-directional promoter from which sense and antisense transcripts are synthesized in both active and inactive var genes during late erythrocytic stages, indicating that this mechanism might function in the control of transcriptional timing rather than monoallelic expression [38], [77]. The impact of the regulatory activity of the var gene intron upon local chromatin structure has been previously indicated through nucleosome depletion within the intron [24], [28]. While PfH2A.Z occupancy at the var upstream promoter could only be detected when var2csa was actively transcribed, enrichment at the conserved intron promoter occured in both active and inactive var genes. This is consistent with the previously observed lack of correlation between activity of the var intron promoter and of the upstream var promoter [38], [77].
PfH2A.Z and Sir2
In yeast, H2A.Z acts as a boundary element that antagonizes the spread of Sir2-mediated heterochromatin into euchromatic areas [53], [105]. This is consistent with our findings by IFA, CoIP and ChIP revealing that PfH2A.Z is associated with regions of euchromatin but is absent from the heterochromatin compartment that is characterized by H3K9me3 and HP1 deposition and contains silent var genes (Figures 2, 3, 4). Interestingly, our electron microscopy analysis suggests enrichment of PfH2A.Z at the border between electron light and electron dense nuclear material (Figure 2), which has previously been interpreted to represent eu - and heterochromatin, respectively [38]. This raises the prospect that PfH2A.Z may also form a barrier to the spread of Sir2-mediated heterochromatin. This is supported by our finding that in 3D7Δsir2A parasites PfH2A.Z is enriched in the upstream region of var genes that are highly transcribed not only when the var genes are active in ring stages but also when they are silent in mature schizonts (Figure 5 & [41], [48]). In contrast PfH2A.Z is enriched in the upstream region of highly expressed var genes in 3D7Δsir2B parasites only while the var genes are active in the ring stages. Sir2A-regulated var genes occupy the heterochromatin/euchromatin boundary at both chromosome internal and subtelomeric clusters; in contrast Sir2B regulates the most telomere proximal upsB type var genes that are separated from the rest of the chromosome by the Sir2A regulated subtelomeric upsA var genes [41], [48]. This raises the intriguing possibility that antagonism between Sir2A and PfH2A.Z is involved in maintaining the heterochromatin/euchromatin boundary but Sir2B is not. Future experiments testing the enrichment of PfH2A.Z at boundary sites using ChIP will address this question.
Possibly PfSir2A is not only involved in maintaining var gene silencing in heterochromatin by removing activating histone acetylations such as H3K9ac [35], but also assists in the temporary expulsion of PfH2A.Z from the active var promoter in mature parasites (Model in Figure 6). This could occur indirectly through recruitment of the ATP-dependent chromatin remodeling machinery responsible for histone variant exchange, or directly through Sir2-mediated deacetylation of PfH2A.Z. Sir2 plays such a direct role in regulating H2A.Z levels in human myocytes where over-expressed Sir2 deacetylates H2A.Z which in turn leads to ubiquitination and proteasome-dependent degradation of H2A.Z [106].
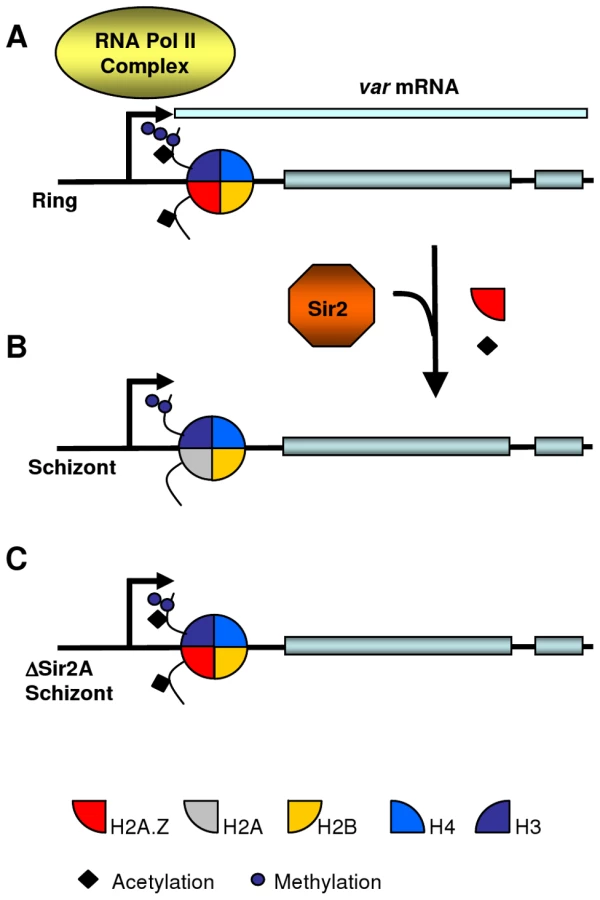
Conclusion
Epigenetic gene regulation contributes significantly to the transcriptional control of fundamental mechanisms such as differentiation and antigenic variation of the malaria parasite (reviewed in [107]). Here, we identify the alternative histone PfH2A.Z as a component of nucleosomes in the promoters of euchromatic genes, suggesting it may be involved in gene regulation. Our report provides the first description of histone variant exchange in P. falciparum, as we demonstrate a temporal modulation of PfH2A.Z occupancy in var genes. This mechanism is disturbed in parasites in which the histone deacetylase Sir2A is disrupted, suggesting a functional link between Sir2A and regulation of PfH2A.Z dynamics at the var promoter.
Materials and Methods
Parasite lines and culture
P. falciparum lines 3D7, 3D7ΔSir2A [41], 3D7ΔSir2B [41] and 3D7HP1GFP [33] were cultured in RPMI medium supplemented with 5% heat-inactivated human serum 0.25% albumax [108]. Synchronicity was maintained by weekly treatment with 5% sorbitol. Selection of var2CSA expressing parasites was performed by panning on plastic dishes coated with 50 µg/ml bovine trachea CSA (Sigma), as described previously [108].
Recombinant protein expression and immunization
The full length PfH2A.Z (PFC0920w) sequence was amplified from 3D7 cDNA using oligonucleotides PfH2A.ZBamHIFor: 5′-GGGATCCGGATGGAAGTTCCAGGAAAAGT and PfH2A.ZEcoRIRev: 5′-GAATTCTTATTGAGCTGTTGGGGGAAGTG. The PCR product was cloned into pGEX-5X-3 (GE Healthcare). PfH2A.ZGST fusion protein was expressed in BL21 cells in Luria Broth and induced with 1mM IPTG. Bacteria pellets were resuspended in Bugsbuster reagent and benzonase (Novagen) and the soluble proteins were purified on GST-Bind Resin (Novagen). Antibodies were generated in rabbits by the Walter and Eliza Hall Institute Monoclonal Antibody Facility (Bundoora, Vic. Australia).
Antibodies
Primary antibodies employed in ChIP assays in this study were rabbit anti-PfH2A.Z, rabbit anti-H3 (Abcam Ab1791), rabbit anti-H2A (Millipore 07-146), rabbit anti-H2B (Abcam Ab1790), non-immune rabbit IgG (Abcam Ab 46540) and pre-immune rabbit serum. Primary antibodies used for IFA co-localization and WB were rabbit anti-H3K9me3 (Abcam Ab8898), rabbit anti-H3K4me3 (Millipore 04-745) and rabbit anti-H3K9ac (Millipore 06-942). Secondary antibodies for IFA were goat anti-rabbit AlexaFluor488 or chicken anti-rabbit AlexaFluor594 (Molecular Probes). Secondary antibodies for WB were goat anti-rabbit HRP (Invitrogen).
Parasite lysates for Western Blot analysis (WB)
Parasites were harvested in 8 hour intervals at 5–10% parasitemia. Lysates were generated by saponin lysis of cultures and extraction of the resulting parasite pellets with 2 x SDS PAGE loading buffer. Equivalents of 5×107 IE were separated by SDS-PAGE on 10% Bis-Tris gels (Invitrogen) and analysed by Western Blotting as described previously [109].
Immunofluorescence analysis (IFA)
IFA was performed on paraformaldehyde/glutaraldehyde-fixed cultures as described previously [110]. For co-localization studies, histone modifications were first labelled using chicken anti-rabbit AlexaFluor594 (1∶1000) (Molecular Probes) as a secondary antibody. PfH2A.Z was subsequently detected with affinity purified rabbit anti-PfH2A.Z (10 ng/µl) directly labelled using the Zenon Rabbit IgG labelling kit (Invitrogen) according to the manufacturers instructions. IE were mounted onto slides using ProLong antifade (Invitrogen), left overnight to cure and analysed with an Olympus FV1000 Confocal Laser Scanning Microscope and the FluoView software.
Immunoelectron microscopy
Parasites were fixed in 1% glutaraldehyde for 1 h at 4°C, dehydrated in increasing ethanol concentrations, then embedded in LR Gold resin (Electron Microscopy Sciences, Fort Washington, PA). Ultrathin (90–100 nm) sections were cut using a Leica Ultracut R microtome, labeled with rabbit anti-PfH2A.Z and goat-anti-rabbit IgG conjugated to 12 nm colloidal gold (Jackson ImmunoResearch Laboratories). Sections were poststained with uranyl acetate and lead citrate and observed using a Philips CM120 BioTwin Transmission Electron Microscope.
Chromatin immunoprecipitation (ChIP)
Chromatin was isolated at three time points during the intra-erythrocytic developmental cycle (IDC) from early ring (6–14 hpi), trophozoite (24–32 hpi) and schizont (36–44 hpi) stage parasites. Parasite cultures were cross-linked with 1% paraformaldehyde for 10 min at 37°C and the reaction subsequently quenched with 125 mM glycine. After one wash with PBS parasites were released from IE by saponin lysis, and nuclei were isolated by incubation for 30 min on ice in lysis buffer (10 mM Hepes pH 7.9, 10 mM KCl, 0.1 mM EDTA, 0.1 mM EDTA, 1 mM DTT, 1x EDTA-free protease inhibitor cocktail (Roche)) followed by dounce homogenization (Pestle B). 0.25% NP40 was added to the parasite suspension prior to homogenization. Nuclei were pelleted by centrifugation at 21,000×g for 10 min at 4°C and resuspended in SDS lysis buffer (1% SDS, 10 mM EDTA, 50 mM Tris pH 8.1, 1 x EDTA-free protease inhibitor cocktail). Chromatin was sheared into 200–1000 bp fragments by sonication for 2×8 min at 30 sec intervals using a Bioruptor UCD-200 (Diagenode) and diluted 1∶10 in ChIP dilution buffer (0.01% SDS, 1.1% Triton X-100, 1.2 mM EDTA, 16.7 mM Tris pH 8.1, 150 mM NaCl).
Immunoprecipitation was performed with the EZ ChIP Kit (Millipore). For each IP, 1×109 ring stage parasites, 2.5×108 trophozoites or 1×108 schizonts were used. Optimal antibody dilutions were determined empirically and ranged between 1∶50 and 1∶200.
Real time qPCR analysis
Primers amplifying upstream regions near the TSS (ups) and in the open reading frame (orf) were designed using the Primer Express software (Applied Biosystems) (Table S1). For genes whose TSS had not previously been experimentally determined and reported, predictions were obtained from the MAPP algorithm implemented at PlasmoDB [111]. Immunoprecipitated DNA and input DNA was quantified by real time qPCR (Applied Biosystems 7900HT) with SYBR PCR master mix (Applied Biosystems). Optimal PCR conditions were determined for each primer pair using serial dilutions of gDNA. PCR was performed in duplicates and melting curves were analysed after each run to confirm the specificity of the amplification. ChIP recoveries were normalized for input signals (ΔCt) and corrected for values obtained with non-immune control antibodies (ΔΔCt). The site specific enrichment of histones was calculated as the 2−ΔΔCt value. To correct for differences in nucleosome density, enrichment of PfH2A.Z and H2A was expressed as a ratio over H3. As in the course of the study different H3 antibody lots were used to precipitate biological replicates, the H3 recoveries and therefore ratios varied considerably between experiments. However, the pattern of histone variant enrichment across genes was reproducible between experiments (Figure 3, 4 & S4).
For statistical analysis of PfH2A.Z enrichment in 3D7ΔSir2A and 3D7 parasites, the ratio of PfH2A.Z enrichment near the TSS and in the open reading frame (ups/orf ratio) was calculated. Paired t-test was performed using the GraphPad Prism software (Version 4).
Mononucleosome co-immunoprecipitation
Mononucleosomes were prepared from freshly isolated nuclei by MNase (NEB) digestion with 20 KU MNase per 1×109 IE and extraction with salt-free buffers [33]. Mononucleosomes from 2.5×108 IE were incubated with 10 µl of antiserum overnight at 4°C and precipitated with 30 µl protein G agarose (Millipore). After extensive washing proteins were eluted with 2 x SDS PAGE loading buffer and analysed by SDS-PAGE and Western Blotting.
Transcription analysis
Total RNA was harvested in parallel to the chromatin preparation by lysis of pelleted IEs in 20 pellet volumes of TRIzol (Invitrogen). RNA was purified as described previously [112] and cDNA was generated using Superscript III Reverse Transcriptase (Invitrogen). Quantitative RT-PCR was performed as described previously [47] using gene specific primers targeting the open reading frame, listed in Table S1. The level of each sequence in cDNA was determined relative to its level in a constant quantity of 3D7 strain gDNA and the amounts of cDNA and gDNA were normalised using the housekeeping gene arginyl-tRNA synthetase or the sbp1 gene by 2−ΔΔCt analysis (Applied Biosystems user bulletin 2).
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. World Health Organization 2009 World Malaria Report 2009. Geneva WHO Press 27
2. BeesonJG
BrownGV
2002 Pathogenesis of Plasmodium falciparum malaria: the roles of parasite adhesion and antigenic variation. Cell Mol Life Sci 59 258 271
3. BiggsBA
GoozeL
WycherleyK
WilkinsonD
BoydAW
1990 Knob-independent cytoadherence of Plasmodium falciparum to the leukocyte differentiation antigen CD36. J Exp Med 171 1883 1892
4. SuXZ
HeatwoleVM
WertheimerSP
GuinetF
HerrfeldtJA
1995 The large diverse gene family var encodes proteins involved in cytoadherence and antigenic variation of Plasmodium falciparum-infected erythrocytes. Cell 82 89 100
5. BaruchDI
PasloskeBL
SinghHB
BiX
MaXC
1995 Cloning the P. falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell 82 77 87
6. GardnerMJ
HallN
FungE
WhiteO
BerrimanM
2002 Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 419 498 511
7. DzikowskiR
FrankM
DeitschK
2006 Mutually exclusive expression of virulence genes by malaria parasites is regulated independently of antigen production. PLoS Pathog 2 e22
8. VossTS
HealerJ
MartyAJ
DuffyMF
ThompsonJK
2006 A var gene promoter controls allelic exclusion of virulence genes in Plasmodium falciparum malaria. Nature 439 1004 1008
9. SmithJD
ChitnisCE
CraigAG
RobertsDJ
Hudson-TaylorDE
1995 Switches in expression of Plasmodium falciparum var genes correlate with changes in antigenic and cytoadherent phenotypes of infected erythrocytes. Cell 82 101 110
10. BiggsBA
GoozeL
WycherleyK
WollishW
SouthwellB
1991 Antigenic variation in Plasmodium falciparum. Proc Natl Acad Sci U S A 88 9171 9174
11. BiggsBA
AndersRF
DillonHE
DavernKM
MartinM
1992 Adherence of infected erythrocytes to venular endothelium selects for antigenic variants of Plasmodium falciparum. J Immunol 149 2047 2054
12. RobertsDJ
CraigAG
BerendtAR
PinchesR
NashG
1992 Rapid switching to multiple antigenic and adhesive phenotypes in malaria. Nature 357 689 692
13. BozdechZ
LlinasM
PulliamBL
WongED
ZhuJ
2003 The transcriptome of the intraerythrocytic developmental cycle of Plasmodium falciparum. PLoS Biol 1 E5
14. Le RochKG
ZhouY
BlairPL
GraingerM
MochJK
2003 Discovery of gene function by expression profiling of the malaria parasite life cycle. Science 301 1503 1508
15. BalajiS
BabuMM
IyerLM
AravindL
2005 Discovery of the principal specific transcription factors of Apicomplexa and their implication for the evolution of the AP2-integrase DNA binding domains. Nucleic Acids Res 33 3994 4006
16. De SilvaEK
GehrkeAR
OlszewskiK
LeonI
ChahalJS
2008 Specific DNA-binding by apicomplexan AP2 transcription factors. Proc Natl Acad Sci U S A 105 8393 8398
17. YudaM
IwanagaS
ShigenobuS
MairGR
JanseCJ
2009 Identification of a transcription factor in the mosquito-invasive stage of malaria parasites. Mol Microbiol 71 1402 1414
18. FlueckC
BartfaiR
NiederwieserI
WitmerK
AlakoBT
2010 A major role for the Plasmodium falciparum ApiAP2 protein PfSIP2 in chromosome end biology. PLoS Pathog 6 e1000784
19. MairGR
BraksJA
GarverLS
WiegantJC
HallN
2006 Regulation of sexual development of Plasmodium by translational repression. Science 313 667 669
20. MairGR
LasonderE
GarverLS
Franke-FayardBM
CarretCK
2010 Universal features of post-transcriptional gene regulation are critical for Plasmodium zygote development. PLoS Pathog 6 e1000767
21. ChungDW
PontsN
CervantesS
Le RochKG
2009 Post-translational modifications in Plasmodium: more than you think! Mol Biochem Parasitol 168 123 134
22. Salcedo-AmayaAM
van DrielMA
AlakoBT
TrelleMB
van den ElzenAM
2009 Dynamic histone H3 epigenome marking during the intraerythrocytic cycle of Plasmodium falciparum. Proc Natl Acad Sci U S A 106 9655 9660
23. ChaalBK
GuptaAP
WastuwidyaningtyasBD
LuahYH
BozdechZ
2010 Histone deacetylases play a major role in the transcriptional regulation of the Plasmodium falciparum life cycle. PLoS Pathog 6 e1000737
24. PontsN
HarrisEY
PrudhommeJ
WickI
Eckhardt-LudkaC
2010 Nucleosome landscape and control of transcription in the human malaria parasite. Genome Res 20 228 238
25. CoulsonRM
HallN
OuzounisCA
2004 Comparative genomics of transcriptional control in the human malaria parasite Plasmodium falciparum. Genome Res 14 1548 1554
26. BischoffE
VaqueroC
2010 In silico and biological survey of transcription-associated proteins implicated in the transcriptional machinery during the erythrocytic development of Plasmodium falciparum. BMC Genomics 11 34
27. TempletonTJ
IyerLM
AnantharamanV
EnomotoS
AbrahanteJE
2004 Comparative analysis of apicomplexa and genomic diversity in eukaryotes. Genome Res 14 1686 1695
28. WestenbergerSJ
CuiL
DhariaN
WinzelerE
2009 Genome-wide nucleosome mapping of Plasmodium falciparum reveals histone-rich coding and histone-poor intergenic regions and chromatin remodeling of core and subtelomeric genes. BMC Genomics 10 610
29. SchonesDE
CuiK
CuddapahS
RohTY
BarskiA
2008 Dynamic regulation of nucleosome positioning in the human genome. Cell 132 887 898
30. LeeCK
ShibataY
RaoB
StrahlBD
LiebJD
2004 Evidence for nucleosome depletion at active regulatory regions genome-wide. Nat Genet 36 900 905
31. MavrichTN
JiangC
IoshikhesIP
LiX
VentersBJ
2008 Nucleosome organization in the Drosophila genome. Nature 453 358 362
32. CuiL
MiaoJ
FuruyaT
LiX
SuXZ
2007 PfGCN5-Mediated Histone H3 Acetylation Plays a Key Role in Gene Expression in Plasmodium falciparum. Eukaryot Cell 6 1219 1227
33. FlueckC
BartfaiR
VolzJ
NiederwieserI
Salcedo-AmayaAM
2009 Plasmodium falciparum heterochromatin protein 1 marks genomic loci linked to phenotypic variation of exported virulence factors. PLoS Pathog 5 e1000569
34. Lopez-RubioJJ
GontijoAM
NunesMC
IssarN
Hernandez RivasR
2007 5′ flanking region of var genes nucleate histone modification patterns linked to phenotypic inheritance of virulence traits in malaria parasites. Mol Microbiol 66 1296 1305
35. Lopez-RubioJJ
Mancio-SilvaL
ScherfA
2009 Genome-wide analysis of heterochromatin associates clonally variant gene regulation with perinuclear repressive centers in malaria parasites. Cell Host Microbe 5 179 190
36. Perez-ToledoK
Rojas-MezaAP
Mancio-SilvaL
Hernandez-CuevasNA
DelgadilloDM
2009 Plasmodium falciparum heterochromatin protein 1 binds to tri-methylated histone 3 lysine 9 and is linked to mutually exclusive expression of var genes. Nucleic Acids Res 37 2596 2606
37. Freitas-JuniorLH
BottiusE
PirritLA
DeitschKW
ScheidigC
2000 Frequent ectopic recombination of virulence factor genes in telomeric chromosome clusters of P. falciparum. Nature 407 1018 1022
38. RalphSA
Scheidig-BenatarC
ScherfA
2005 Antigenic variation in Plasmodium falciparum is associated with movement of var loci between subnuclear locations. Proc Natl Acad Sci U S A 102 5414 5419
39. MartyAJ
ThompsonJK
DuffyMF
VossTS
CowmanAF
2006 Evidence that Plasmodium falciparum chromosome end clusters are cross-linked by protein and are the sites of both virulence gene silencing and activation. Mol Microbiol 62 72 83
40. DzikowskiR
LiF
AmulicB
EisbergA
FrankM
2007 Mechanisms underlying mutually exclusive expression of virulence genes by malaria parasites. EMBO Rep 8 959 965
41. DuraisinghMT
VossTS
MartyAJ
DuffyMF
GoodRT
2005 Heterochromatin silencing and locus repositioning linked to regulation of virulence genes in Plasmodium falciparum. Cell 121 13 24
42. ScherfA
Hernandez-RivasR
BuffetP
BottiusE
BenatarC
1998 Antigenic variation in malaria: in situ switching, relaxed and mutually exclusive transcription of var genes during intra-erythrocytic development in Plasmodium falciparum. Embo J 17 5418 5426
43. DeitschKW
CalderwoodMS
WellemsTE
2001 Malaria. Cooperative silencing elements in var genes. Nature 412 875 876
44. CalderwoodMS
Gannoun-ZakiL
WellemsTE
DeitschKW
2003 Plasmodium falciparum var genes are regulated by two regions with separate promoters, one upstream of the coding region and a second within the intron. J Biol Chem 278 34125 34132
45. Gannoun-ZakiL
JostA
MuJ
DeitschKW
WellemsTE
2005 A silenced Plasmodium falciparum var promoter can be activated in vivo through spontaneous deletion of a silencing element in the intron. Eukaryot Cell 4 490 492
46. FrankM
DzikowskiR
CostantiniD
AmulicB
BerdougoE
2006 Strict pairing of var promoters and introns is required for var gene silencing in the malaria parasite Plasmodium falciparum. J Biol Chem 281 9942 9952
47. DuffyMF
ByrneTJ
CarretC
IvensA
BrownGV
2009 Ectopic recombination of a malaria var gene during mitosis associated with an altered var switch rate. J Mol Biol 389 453 469
48. TonkinCJ
CarretCK
DuraisinghMT
VossTS
RalphSA
2009 Sir2 paralogues cooperate to regulate virulence genes and antigenic variation in Plasmodium falciparum. PLoS Biol 7 e84
49. TalbertPB
HenikoffS
2010 Histone variants—ancient wrap artists of the epigenome. Nat Rev Mol Cell Biol 11 264 275
50. FaastR
ThonglairoamV
SchulzTC
BeallJ
WellsJR
2001 Histone variant H2A.Z is required for early mammalian development. Curr Biol 11 1183 1187
51. ClarksonMJ
WellsJR
GibsonF
SaintR
TremethickDJ
1999 Regions of variant histone His2AvD required for Drosophila development. Nature 399 694 697
52. LiuX
LiB
GorovskyMa
1996 Essential and nonessential histone H2A variants in Tetrahymena thermophila. Mol Cell Biol 16 4305 4311
53. MeneghiniMD
WuM
MadhaniHD
2003 Conserved histone variant H2A.Z protects euchromatin from the ectopic spread of silent heterochromatin. Cell 112 725 736
54. BarskiA
CuddapahS
CuiK
RohTY
SchonesDE
2007 High-resolution profiling of histone methylations in the human genome. Cell 129 823 837
55. RaisnerRM
HartleyPD
MeneghiniMD
BaoMZ
LiuCL
2005 Histone variant H2A.Z marks the 5′ ends of both active and inactive genes in euchromatin. Cell 123 233 248
56. DalmassoMC
OnyangoDO
NaguleswaranA
SullivanWJJr
AngelSO
2009 Toxoplasma H2A Variants Reveal Novel Insights into Nucleosome Composition and Functions for this Histone Family. J Mol Biol 392 33 47
57. ZhangH
RobertsDN
CairnsBR
2005 Genome-wide dynamics of Htz1, a histone H2A variant that poises repressed/basal promoters for activation through histone loss. Cell 123 219 231
58. MiaoJ
FanQ
CuiL
LiJ
2006 The malaria parasite Plasmodium falciparum histones: organization, expression, and acetylation. Gene 369 53 65
59. IouzalenN
MoreauJ
MechaliM
1996 H2A.ZI, a new variant histone expressed during Xenopus early development exhibits several distinct features from the core histone H2A. Nucleic Acids Res 24 3947 3952
60. TrelleMB
Salcedo-AmayaAM
CohenAM
StunnenbergHG
JensenON
2009 Global histone analysis by mass spectrometry reveals a high content of acetylated lysine residues in the malaria parasite Plasmodium falciparum. J Proteome Res 8 3439 3450
61. SutoRK
ClarksonMJ
TremethickDJ
LugerK
2000 Crystal structure of a nucleosome core particle containing the variant histone H2A.Z. Nat Struct Biol 7 1121 1124
62. IssarN
RalphSA
Mancio-SilvaL
KeelingC
ScherfA
2009 Differential sub-nuclear localisation of repressive and activating histone methyl modifications in P. falciparum. Microbes Infect 11 403 407
63. AdamsRR
MaiatoH
EarnshawWC
CarmenaM
2001 Essential roles of Drosophila inner centromere protein (INCENP) and aurora B in histone H3 phosphorylation, metaphase chromosome alignment, kinetochore disjunction, and chromosome segregation. J Cell Biol 153 865 880
64. BruceK
MyersFA
MantouvalouE
LefevreP
GreavesI
2005 The replacement histone H2A.Z in a hyperacetylated form is a feature of active genes in the chicken. Nucleic Acids Res 33 5633 5639
65. LarochelleM
GaudreauL
2003 H2A.Z has a function reminiscent of an activator required for preferential binding to intergenic DNA. EMBO J 22 4512 4522
66. SantistebanMS
KalashnikovaT
SmithMM
2000 Histone H2A.Z regulats transcription and is partially redundant with nucleosome remodeling complexes. Cell 103 411 422
67. KroganNJ
BaetzK
KeoghMC
DattaN
SawaC
2004 Regulation of chromosome stability by the histone H2A variant Htz1, the Swr1 chromatin remodeling complex, and the histone acetyltransferase NuA4. Proc Natl Acad Sci U S A 101 13513 13518
68. RangasamyD
GreavesI
TremethickDJ
2004 RNA interference demonstrates a novel role for H2A.Z in chromosome segregation. Nat Struct Mol Biol 11 650 655
69. RangasamyD
BervenL
RidgwayP
TremethickDJ
2003 Pericentric heterochromatin becomes enriched with H2A.Z during early mammalian development. EMBO J 22 1599 1607
70. SwaminathanJ
BaxterEM
CorcesVG
2005 The role of histone H2Av variant replacement and histone H4 acetylation in the establishment of Drosophila heterochromatin. Genes Dev 19 65 76
71. FanJY
RangasamyD
LugerK
TremethickDJ
2004 H2A.Z alters the nucleosome surface to promote HP1alpha-mediated chromatin fiber folding. Mol Cell 16 655 661
72. GreavesIK
RangasamyD
RidgwayP
TremethickDJ
2007 H2A.Z contributes to the unique 3D structure of the centromere. Proc Natl Acad Sci U S A 104 525 530
73. SarcinellaE
ZuzartePC
LauPN
DrakerR
CheungP
2007 Monoubiquitylation of H2A.Z distinguishes its association with euchromatin or facultative heterochromatin. Mol Cell Biol 27 6457 6468
74. DryhurstD
IshibashiT
RoseKL
Eirin-LopezJM
McDonaldD
2009 Characterization of the histone H2A.Z-1 and H2A.Z-2 isoforms in vertebrates. BMC Biol 7 86
75. Freitas-JuniorLH
Hernandez-RivasR
RalphSA
Montiel-CondadoD
Ruvalcaba-SalazarOK
2005 Telomeric heterochromatin propagation and histone acetylation control mutually exclusive expression of antigenic variation genes in malaria parasites. Cell 121 25 36
76. DuffyMF
ByrneTJ
ElliottSR
WilsonDW
RogersonSJ
2005 Broad analysis reveals a consistent pattern of var gene transcription in Plasmodium falciparum repeatedly selected for a defined adhesion phenotype. Mol Microbiol 56 774 788
77. EppC
LiF
HowittCA
ChookajornT
DeitschKW
2009 Chromatin associated sense and antisense noncoding RNAs are transcribed from the var gene family of virulence genes of the malaria parasite Plasmodium falciparum. RNA 15 116 127
78. LowellJE
KaiserF
JanzenCJ
CrossGA
2005 Histone H2AZ dimerizes with a novel variant H2B and is enriched at repetitive DNA in Trypanosoma brucei. J Cell Sci 118 5721 5730
79. van DaalA
ElginSC
1992 A histone variant, H2AvD, is essential in Drosophila melanogaster. Mol Biol Cell 3 593 602
80. van DaalA
WhiteEM
ElginSC
GorovskyMA
1990 Conservation of intron position indicates separation of major and variant H2As is an early event in the evolution of eukaryotes. J Mol Evol 30 449 455
81. EisenJA
CoyneRS
WuM
WuD
ThiagarajanM
2006 Macronuclear genome sequence of the ciliate Tetrahymena thermophila, a model eukaryote. PLoS Biol 4 e286
82. RenQ
GorovskyMA
2001 Histone H2A.Z acetylation modulates an essential charge patch. Mol Cell 7 1329 1335
83. KoborMS
VenkatasubrahmanyamS
MeneghiniMD
GinJW
JenningsJL
2004 A protein complex containing the conserved Swi2/Snf2-related ATPase Swr1p deposits histone variant H2A.Z into euchromatin. PLoS Biol 2 E131
84. WongMM
CoxLK
ChriviaJC
2007 The chromatin remodeling protein, SRCAP, is critical for deposition of the histone variant H2A.Z at promoters. J Biol Chem 282 26132 26139
85. MizuguchiG
ShenX
LandryJ
WuWH
SenS
2004 ATP-driven exchange of histone H2AZ variant catalyzed by SWR1 chromatin remodeling complex. Science 303 343 348
86. FanJY
GordonF
LugerK
HansenJC
TremethickDJ
2002 The essential histone variant H2A.Z regulates the equilibrium between different chromatin conformational states. Nat Struct Biol 9 172 176
87. SiegelTN
HekstraDR
KempLE
FigueiredoLM
LowellJE
2009 Four histone variants mark the boundaries of polycistronic transcription units in Trypanosoma brucei. Genes Dev 23 1063 1076
88. GuillemetteB
BatailleAR
GevryN
AdamM
BlanchetteM
2005 Variant histone H2A.Z is globally localized to the promoters of inactive yeast genes and regulates nucleosome positioning. PLoS Biol 3 e384
89. PokholokDK
HarbisonCT
LevineS
ColeM
HannettNM
2005 Genome-wide map of nucleosome acetylation and methylation in yeast. Cell 122 517 527
90. HardyS
JacquesPE
GevryN
ForestA
FortinME
2009 The euchromatic and heterochromatic landscapes are shaped by antagonizing effects of transcription on H2A.Z deposition. PLoS Genet 5 e1000687
91. CuiK
ZangC
RohTY
SchonesDE
ChildsRW
2009 Chromatin signatures in multipotent human hematopoietic stem cells indicate the fate of bivalent genes during differentiation. Cell Stem Cell 4 80 93
92. AdamM
RobertF
LarochelleM
GaudreauL
2001 H2A.Z is required for global chromatin integrity and for recruitment of RNA polymerase II under specific conditions. Mol Cell Biol 21 6270 6279
93. GopalakrishnanAM
NyindodoLA
Ross FergusM
Lopez-EstranoC
2009 Plasmodium falciparum: Preinitiation complex occupancy of active and inactive promoters during erythrocytic stage. Exp Parasitol 121 46 54
94. MillarCB
XuF
ZhangK
GrunsteinM
2006 Acetylation of H2AZ Lys 14 is associated with genome-wide gene activity in yeast. Genes Dev 20 711 722
95. ThambirajahAA
DryhurstD
IshibashiT
LiA
MaffeyAH
2006 H2A.Z stabilizes chromatin in a way that is dependent on core histone acetylation. J Biol Chem 281 20036 20044
96. IizukaM
SmithMM
2003 Functional consequences of histone modifications. Curr Opin Genet Dev 13 154 160
97. OuyangJ
GillG
2009 SUMO engages multiple corepressors to regulate chromatin structure and transcription. Epigenetics 4 440 444
98. IssarN
RouxE
MatteiD
ScherfA
2008 Identification of a novel post-translational modification in Plasmodium falciparum: protein sumoylation in different cellular compartments. Cell Microbiol 10 1999 2011
99. KumarSV
WiggePA
2010 H2A.Z-containing nucleosomes mediate the thermosensory response in Arabidopsis. Cell 140 136 147
100. JinC
FelsenfeldG
2007 Nucleosome stability mediated by histone variants H3.3 and H2A.Z. Genes Dev 21 1519 1529
101. JinC
ZangC
WeiG
CuiK
PengW
2009 H3.3/H2A.Z double variant-containing nucleosomes mark ‘nucleosome-free regions’ of active promoters and other regulatory regions. Nat Genet 41 941 945
102. MerrickCJ
DuraisinghMT
2006 Heterochromatin-mediated control of virulence gene expression. Mol Microbiol 62 612 620
103. Hernandez-RivasR
Perez-ToledoK
Herrera SolorioAM
DelgadilloDM
VargasM
2010 Telomeric heterochromatin in Plasmodium falciparum. J Biomed Biotechnol 2010 290501
104. BricknerDG
CajigasI
Fondufe-MittendorfY
AhmedS
LeePC
2007 H2A.Z-mediated localization of genes at the nuclear periphery confers epigenetic memory of previous transcriptional state. PLoS Biol 5 e81
105. ZhouBO
WangSS
XuLX
MengFL
XuanYJ
2010 SWR1 Complex Poises Heterochromatin Boundaries for Antisilencing Activity Propagation. Mol Cell Biol 30 2391 2400
106. ChenIY
LypowyJ
PainJ
SayedD
GrinbergS
2006 Histone H2A.z is essential for cardiac myocyte hypertrophy but opposed by silent information regulator 2alpha. J Biol Chem 281 19369 19377
107. CuiL
MiaoJ
2010 Chromatin-mediated epigenetic regulation in the malaria parasite Plasmodium falciparum. Eukaryot Cell 9 1138 1149
108. NoviyantiR
BrownGV
WickhamME
DuffyMF
CowmanAF
2001 Multiple var gene transcripts are expressed in Plasmodium falciparum infected erythrocytes selected for adhesion. Mol Biochem Parasitol 114 227 237
109. PetterM
BonowI
KlinkertMQ
2008 Diverse expression patterns of subgroups of the rif multigene family during Plasmodium falciparum gametocytogenesis. PLoS ONE 3 e3779
110. TonkinCJ
van DoorenGG
SpurckTP
StruckNS
GoodRT
2004 Localization of organellar proteins in Plasmodium falciparum using a novel set of transfection vectors and a new immunofluorescence fixation method. Mol Biochem Parasitol 137 13 21
111. BrickK
WatanabeJ
PizziE
2008 Core promoters are predicted by their distinct physicochemical properties in the genome of Plasmodium falciparum. Genome Biol 9 R178
112. KyesS
PinchesR
NewboldC
2000 A simple RNA analysis method shows var and rif multigene family expression patterns in Plasmodium falciparum. Mol Biochem Parasitol 105 311 315
113. LavstsenT
SalantiA
JensenAT
ArnotDE
TheanderTG
2003 Sub-grouping of Plasmodium falciparum 3D7 var genes based on sequence analysis of coding and non-coding regions. Malar J 2 27
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2011 Číslo 2
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- Genetic Mapping Identifies Novel Highly Protective Antigens for an Apicomplexan Parasite
- Type I Interferon Signaling Regulates Ly6C Monocytes and Neutrophils during Acute Viral Pneumonia in Mice
- Infections in Cells: Transcriptomic Characterization of a Novel Host-Symbiont Interaction
- The ESCRT-0 Component HRS is Required for HIV-1 Vpu-Mediated BST-2/Tetherin Down-Regulation
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy