
Highly Differentiated, Resting Gn-Specific Memory CD8 T Cells Persist Years after Infection by Andes Hantavirus
In man, infection with South American Andes virus (ANDV) causes hantavirus cardiopulmonary syndrome (HCPS). HCPS due to ANDV is endemic in Southern Chile and much of Argentina and increasing numbers of cases are reported all over South America. A case-fatality rate of about 36% together with the absence of successful antiviral therapies urge the development of a vaccine. Although T-cell responses were shown to be critically involved in immunity to hantaviruses in mouse models, no data are available on the magnitude, specificity and longevity of ANDV-specific memory T-cell responses in patients. Using sets of overlapping peptides in IFN-γ ELISPOT assays, we herein show in 78 Chilean convalescent patients that Gn-derived epitopes were immunodominant as compared to those from the N - and Gc-proteins. Furthermore, while the relative contribution of the N-specific response significantly declined over time, Gn-specific responses remained readily detectable ex vivo up to 13 years after the acute infection. Tetramer analysis further showed that up to 16.8% of all circulating CD3+CD8+ T cells were specific for the single HLA-B*3501-restricted epitope Gn465–473 years after the acute infection. Remarkably, Gn465–473–specific cells readily secreted IFN-γ, granzyme B and TNF-α but not IL-2 upon stimulation and showed a ‘revertant’ CD45RA+CD27−CD28−CCR7−CD127− effector memory phenotype, thereby resembling a phenotype seen in other latent virus infections. Most intriguingly, titers of neutralizing antibodies increased over time in 10/17 individuals months to years after the acute infection and independently of whether they were residents of endemic areas or not. Thus, our data suggest intrinsic, latent antigenic stimulation of Gn-specific T-cells. However, it remains a major task for future studies to proof this hypothesis by determination of viral antigen in convalescent patients. Furthermore, it remains to be seen whether Gn-specific T cells are critical for viral control and protective immunity. If so, Gn-derived immunodominant epitopes could be of high value for future ANDV vaccines.
Published in the journal:
. PLoS Pathog 6(2): e32767. doi:10.1371/journal.ppat.1000779
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1000779
Summary
In man, infection with South American Andes virus (ANDV) causes hantavirus cardiopulmonary syndrome (HCPS). HCPS due to ANDV is endemic in Southern Chile and much of Argentina and increasing numbers of cases are reported all over South America. A case-fatality rate of about 36% together with the absence of successful antiviral therapies urge the development of a vaccine. Although T-cell responses were shown to be critically involved in immunity to hantaviruses in mouse models, no data are available on the magnitude, specificity and longevity of ANDV-specific memory T-cell responses in patients. Using sets of overlapping peptides in IFN-γ ELISPOT assays, we herein show in 78 Chilean convalescent patients that Gn-derived epitopes were immunodominant as compared to those from the N - and Gc-proteins. Furthermore, while the relative contribution of the N-specific response significantly declined over time, Gn-specific responses remained readily detectable ex vivo up to 13 years after the acute infection. Tetramer analysis further showed that up to 16.8% of all circulating CD3+CD8+ T cells were specific for the single HLA-B*3501-restricted epitope Gn465–473 years after the acute infection. Remarkably, Gn465–473–specific cells readily secreted IFN-γ, granzyme B and TNF-α but not IL-2 upon stimulation and showed a ‘revertant’ CD45RA+CD27−CD28−CCR7−CD127− effector memory phenotype, thereby resembling a phenotype seen in other latent virus infections. Most intriguingly, titers of neutralizing antibodies increased over time in 10/17 individuals months to years after the acute infection and independently of whether they were residents of endemic areas or not. Thus, our data suggest intrinsic, latent antigenic stimulation of Gn-specific T-cells. However, it remains a major task for future studies to proof this hypothesis by determination of viral antigen in convalescent patients. Furthermore, it remains to be seen whether Gn-specific T cells are critical for viral control and protective immunity. If so, Gn-derived immunodominant epitopes could be of high value for future ANDV vaccines.
Introduction
The family Bunyaviridae is comprised of five genera of tri-segmented negative-stranded RNA viruses, which are responsible for a considerable burden of zoonotic disease in man. While most are tick - or mosquito-borne, members of the genus Hantavirus are transmitted from chronically - and asymptomatically-infected rodents to humans via aerosols, which may derive from urine, feces or saliva. Globally hantaviruses may cause as many as 200,000 cases of human disease per year. In man, two clinical conditions may arise: hemorrhagic fever with renal syndrome, caused by the Asian and European strains (e.g. Hantaan, HTNV and Puumala, PUUV) or hantavirus cardiopulmonary syndrome (HCPS), which is caused by Sin Nombre virus (SNV) and Andes virus (ANDV), among others in the Americas. HCPS is an emerging infectious disease in North - and South America [1]-[5] and, currently, Chile represents among the most endemic regions for HCPS with more than 580 cases since 1995 [6].
As for ANDV, transmission to man is followed by infection of lung endothelial cells and, after an incubation period of 7 to 39 days [7], the development of a vascular leakage syndrome, eventually leading to massive pulmonary edema, shock and, in many cases, death. The high case-fatality ratio (mean 36%), the absence of a proven antiviral treatment or a vaccine, their mode of transmission and their potential use as weapons for bioterrorism, have rendered HCPS-causing hantaviruses Category A pathogens within NIAID's biodefense program [8]. Importantly, ANDV is the only hantavirus for which person-to-person transmission has been repeatedly documented [9]–[11].
The hantavirus virion contains a lipid-bilayer envelope into which both constituents, the Gn and Gc antigens of the heteromeric glycoprotein, are inserted via transmembrane domains. In the viral core, there are three nucleocapids each consisting of the RNA-binding N or nucleocapsid protein in complex with one of the genomic RNAs. These mRNAs encode the RNA-dependent RNA polymerase or L protein on the large or L segment (2153aa), the Gn (650aa) and Gc (490aa) glycoproteins on the middle or M segment, and the N protein (430aa) on the S segment [12].
Currently, there is a big discrepancy regarding the role of T cells in either pathogenesis or immunity of hantavirus infections. On one hand some studies in SNV-infected patients describe a correlation between the severity of HCPS and either the frequency of SNV-specific CD8+ T-cells [13] or the HLA-B35 haplotype [14], suggesting a T-cell driven pathogenesis of HCPS. On the other hand, several early reports highlight the importance of lymphocytes for immunity of mice towards hantaviruses, such as HTNV [15]–[17]. Likewise, clearance of HTNV in newborn mice was dependent on TNF-α production and cytotoxic activity of specific CD8+ T cells [18]. In addition, (HTNV-) N-protein-specific memory T-cells conferred partial protection and cross-protection towards N-expressing vaccinia virus [19] or hantaviruses [20] in mice and Syrian hamster [21],[22], respectively. In line with these findings we have recently reported that clearance of ANDV-RNA from peripheral blood cells of a patient was closely related to the appearance of cytotoxic CD8+ T cells about two months after the acute infection [23]. This observation together with the finding that we were unable to detect memory T-cells in many of the survivors of ANDV-induced HCPS (see below) led us to the hypothesis that T-cells may be crucial for protection and immunity towards ANDV rather than the pathogenesis in ANDV-infected patients. Concisely, knowledge of targeted epitopes and functional properties of ANDV-specific T-cells in ANDV-survivors may be important for both future studies in acutely ill patients and possibly for vaccine development.
Despite its importance, the knowledge of the human cellular immune response to hantaviruses is limited. To date, only few studies have assessed SNV-, HTNV - or PUUV - specific T cells [13], [24]–[28] in rather small patient cohorts and based on individual in-silico-predicted peptides and/or T-cell lines that had been expanded in vitro. Thus, the overall magnitude of human T-cell responses in vivo and the epitope-hierarchy within the memory T-cell pool in convalescent patients remains uncertain. Also, phenotype, effector functions and longevity of specific memory T-cells in humans remain to be elucidated. Greater knowledge of these matters would likely to be of high value to potential vaccine developers.
In an effort to gain insight into human cellular immunity to ANDV and to establish an immuno-hierarchy among ANDV-antigens, we carried out a study of the viral protein-specific T cell responses in 78 Chilean patients with past ANDV-infection. Our findings on the immuno-hierarchy among major structural hantavirus proteins and the frequencies as well as the functional features of CD8+ memory T cells may be of special interest for vaccine development since all attempts to induce long-lasting neutralizing humoral immunity have been unsuccessful so far [29].
Results
Gn of ANDV is highly immunogenic
In order to quantify circulating ANDV-specific T-cells ex vivo and to determine the immunodominant epitopes of ANDV, we first challenged PBMC of 78 Chilean convalescent patients (between 4 months and 13.2 years after hospitalization due to infection) with 310 overlapping peptides (distributed in 13 pools) spanning the entire N - and Gn/Gc precursor proteins [30] in IFN-γ ELISPOT assays.
Based on the criteria we used to score a sample as “positive” (see Material and Methods), 51 (66%) of the 78 patients showed significant responses against epitopes of at least one of the three viral antigens (Fig. 1A). Among these patients, 33/51 (65%) showed significant responses against epitopes of the N-protein, while 13/51 (25%) showed Gc-specific T-cells (Fig. 1B). However, 80% (41/51) of the positive patients launched significant responses towards Gn-derived epitopes. Moreover, while mean responses among the 51 individuals reached the sum of 1809 Spot Forming Units (SFU)/106PBMC when considering all viral antigens, Gn-specific responses accounted for more than half of the total response, at 973 SFU/106PBMC (Fig. 1C) as compared to 697 and 139 SFU/106 PBMC for N - and Gc-epitopes, respectively. Since all patients were BCG-vaccinated twice during childhood, we determined BCG-specific T cells (n = 10), resulting in a mean of 162 SFU/106 PBMC. Thus, Gn is the immunodominant antigen in ANDV-convalescent individuals.

Relative stability of the Gn - and Gc-specific memory T-cell pool
We next asked whether differences in the longevity of each of the specific T-cell categories could account for the relative immunodominance of Gn among ANDV antigens, e.g., whether Gn-specific cells might persist longer than did cells responsive to the other antigens. We therefore considered the numbers of circulating N-, Gn - and Gc-specific T cells of each patient in relation to the time between the patient's hospital admission due to HCPS and the timepoint at which T cells were applied to ELISPOT assays (Fig. 2A–C). Although this approach does not allow to draw conclusions on the slope of antigen-specific responses at the level of a given individual, it is possible to directly compare the different antigen-specific responses within the overall cohort over time. Similar approaches have been previously performed on a cohort of smallpox vaccinees in order to estimate longevity and half-life of cellular and humoral memory responses [31].

Interestingly, only the N-specific response (Fig. 2A) exhibited a negative and significantly descending linear regression slope over time (r = −0.11, p<0.05), as when compared to Gn - and Gc-specific responses (Fig. 2B, C) (r = 0.07 and r = 0.02, respectively). We next segregated the 51 patients with positive T-cell responses into six groups according to the time past since hospitalization due to ANDV-infection (<1, 1–2, 2–4, 4–6, 6–9 and >13 years, respectively). As can be seen in Figure 2D, up to four years after infection N-specific responses were predominant, whereas afterwards Gn-specific relative contributions to the overall T-cell response increased from approximately 40% to more than 80% (Fig. 2D). Taken together, these data suggest that Gn - and Gc-specific T-cell responses are more stably maintained as compared to N-specific responses. However, in light of the limited value of the cross-sectional data available to us, future prospective studies assessing individual T-cell responses from the acute to the convalescent phase in individual patients would be needed to determine the absolute half-life of the N-, Gn - and Gc-specific T-cell responses.
Immunodominance of Gn461–475 within the Gn carboxyl-terminus
Subsequently, results from IFN-γ ELISPOT assays revealed that regions Gn1-230 and Gn221–450 elicited a mean of 102 and 209 SFU/106 PBMC in 34% and 38% of all responsive patients, respectively (Fig. 3A). However, Gn441–650 elicited a mean response of 623 SFU/106PBMC (range 0–5506 SFU/106PBMC) in 24/51 (46%) patients. These data clearly indicate that epitopes within the carboxyl-terminus of Gn of ANDV are responsible for the immunodominance of Gn among ANDV-convalescent individuals. In contrast to the strong Gn-specific responses, Gc641–815 and Gc806–980 elicited a response in 24% (77 SFU/106PBMC) and 11% (25 SFU/106PBMC) of all patients, whereas Gc971–1140 was targeted by only 4% (30 SFU/106PBMC) of patients (data not shown).

We next stimulated cryopreserved PBMC from patients with Gn441–650-specific response using peptide pools representing Gn441–505, Gn496–560, Gn551–615 and Gn606–650, respectively. This approach revealed that region Gn441–505 comprised the major epitopes of Gn441–650 (data not shown). We then challenged cryopreserved PBMC of 11 patients with individual peptides spanning region Gn441–505. As shown in Figure 3B, only three patients (p30, p40, p59) exclusively recognized peptides Gn451–465 and Gn456–470, whereas p17 and p57 additionally recognized Gn461–475 and Gn466–480. By contrast, six patients (p6, p10, p15, p28, p32, p53) showed exclusive recognition of Gn461–475 and Gn466–480. Thus, a total of five patients recognized Gn451–465/Gn456–470, whereas eight patients recognized Gn461–475/Gn466–480. Subsequently, four additional individuals with exclusive and significant responses towards Gn461–475/Gn466–480 were identified (data not shown).
Together with a previous report from our lab [23], these epitopes are the first described within the Gn-region of hantaviruses. In addition, we determined immunodominant regions of ANDV N-protein, which included N1–70 (24.3% of responsive patients elicited a significant response) and N121–190 and N181–250 (21.2% and 19.5%, respectively) eliciting mean responses of 113–147 SFU/106PBMC (data not shown). Downmapped individual epitopes within the N-protein are summarized in Table 1.

We next wondered, which state of differentiation was expressed by the IFN-γ producing memory CD8+ T cells. Intracellular cytokine staining showed that IFN-γ+CD8+CD45RO+ T-cells expressed varying levels of CD45RA but consistently expressed a CCR7−CD28−CD27− effector memory phenotype (Fig. 3C). In line with this terminally differentiated phenotype, 25% of all IFN-γ+ T cells secreted granzyme B upon stimulation with their cognate peptide (Fig. 3D) whereas no IL-2 could be detected (data not shown). In addition, up to 45% of these IFN-γ+CD8+ memory T cells co-expressed TNF-α as determined by ICS (data not shown). These findings suggest that even years after acute ANDV infection (e.g. p32 and p40 were investigated 5.4 and 13.2 years after hospitalization, respectively), high frequencies of cytolytic memory CD8+ T-cells are maintained in the periphery.
Impact of HLA-B*35 - restricted responses on clinical outcome
As both the Gn451–465/Gn456–470 and Gn461–475/Gn466–480 epitopes share 10 amino acids in sequence within the pairs, we reasoned that one CD8+ T-cell epitope may be located within each overlapping sequence, respectively (e.g. Gn456–465 and Gn466–475). In support of this hypothesis, the analysis of HLA-A, -B, -DR and -DQ alleles revealed that 5/5 patients with response towards Gn451–465/Gn456–470 exclusively shared the HLA-A*24 allele (data not shown), whereas the HLA-B*35 allele was the only allele shared by all 12 patients recognizing Gn461–475/Gn466–480 (Fig. 4A). These data suggest the existence of two separate but neighboring CD8 T-cell epitopes in the carboxyterminal region of Gn that are restricted by HLA-A*24 and HLA-B*35, respectively. Indeed, as shown in Figure 4B, a significant response in a Gn461–475–specific T-cell line could only be detected when the HLA-B*35 allele was present on heterologous APCs (B-LCL).

It was previously suggested that severe HCPS due to SNV is associated with the HLA-B*35 allele [14] and with CD8 T-cell responses restricted to it [13]. We therefore were interested in the relation of memory T-cell responses and outcome of their ANDV infection in HLA-B*35-positive and negative patients (Fig. S1). Among all 78 patients, that is patients with (n = 51) and without (n = 27) significant memory T-cell responses, no differences in overall T-cell responses could be observed when comparing HLA-B*35-negative patients with mild or severe HCPS (Fig. S1A). By contrast, we found an about 3-fold higher overall T-cell response in HLA-B*35-positive patients with mild HCPS as compared to both HLA-B*35-positive patients with a history of severe disease and either group of HLA-B*35-negative patients (Fig. S1B). Likewise, 10/12 (83%) HLA-B*35-positive patients with significant responses to Gn461-475 had a history of mild HCPS (Fig. S1C). Thus, these data suggest that HLA-B*35-restricted memory T-cell responses are related to mild rather than to severe disease outcome.
We next sought to determine the optimal epitope of Gn461–475/Gn466–480 (Fig. 4C). Because we consistently observed stronger immune responses towards Gn461–475 (SLFSLMPDVAHSLAV) than towards Gn466–480 (MPDVAHSLAVELCVP), we reasoned that Leucine at position 465 may increase either the binding affinity or the TCR-recognition of the overlapping sequence Gn466–475 (MPDVAHSLAV). We therefore decided to generate Gn461–475–specific T-cell lines from HLA-B*3501 individuals and then challenged these cells with cleaved peptides of Gn465–475 (LMPDVAHSLAV). As shown in Figure 4C, cleavage of the carboxyterminal Leucine at position 473 led to a complete loss of epitope recognition. Similarly, elimination of the aminoterminal Methionine at position 466 was critical for epitope recognition. Most interestingly, virtually identical results were obtained when cells from HLA*B3501, HLA-B*3502 and HLA-B*3505 individuals were challenged with cleaved peptides, indicating that the Gn466–473 epitope is equally immunogenic among different HLA-B*35 subtypes.
High frequencies of memory CD8+ T cells that have the potential to exert cytotoxic function are maintained in the periphery
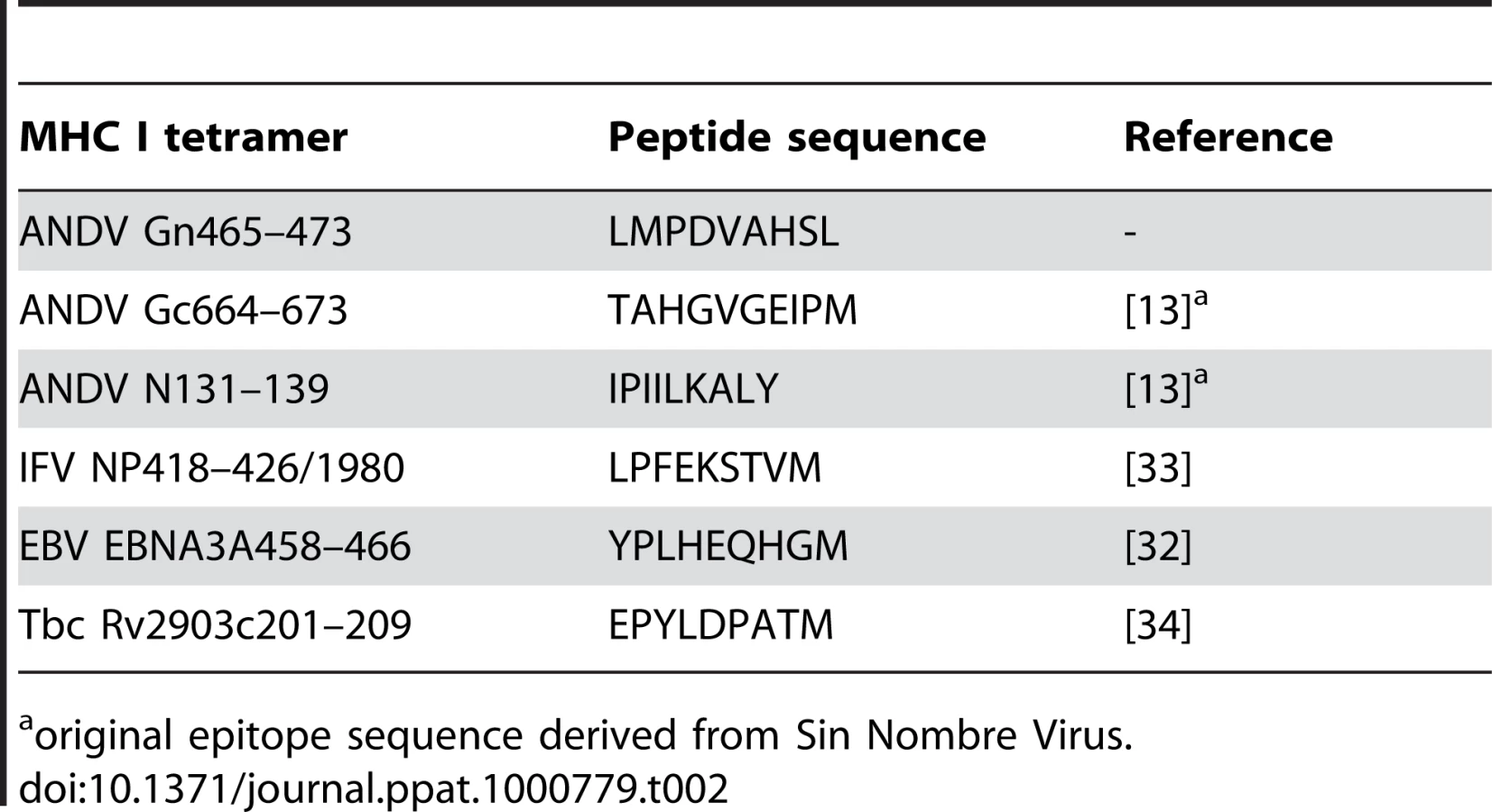
We next were interested in comparing the phenotype of Gn465–473 restricted T cells and with other HLA-B*3501-restricted virus-specific T cells in seven HLA-B*3501 positive ANDV-convalescent patients (Fig. 5A, B). The mean abundance of Gn465–473 specific T-cells was higher than those specific for N131–139 and Gc664–673 –epitopes, described by Kilpatrick et al. [13], the latent EBV-epitope EBNA3A458–466 [32], the Influenza A NP418–426 epitope [33], or the Rv2903c201–209 epitope of Mycobacterium tuberculosis, known to be recognized by BCG-vaccinated individuals [34].

In the seven HLA-B*3501-positive individuals we had detected between 0 and 4394 SFU/106 PBMC (0%–0.0044%) following stimulation with peptide Gn461–475 in IFN-γ ELISPOT assays. When normalizing the results by the percentages of CD3+CD8+ cells in these individuals (range 9.9–33.9% of PBMC, mean 19.7%), one would have expected between 0% (p45) and 0.036% (p17) of CD3+CD8+ cells being tetramer positive. However, we found 0.3% and 5.9% of all CD3+CD8+ T cells being positive for Gn465–473:HLA-B*3501 tetramer complexes, respectively. In addition, the highest frequencies for tetramer-positive cells were found in patient 10 (16.8% of CD3+CD8+ cells), whereas only 0.0111% of his CD3+CD8+ cells produced IFN-γ in ELISPOT assays. This discrepancy between both detection methods is in line with previous reports [35].
We next determined the state of differentiation of Gn465–473-specific T-cells, where a clear dichotomy was observed (Fig. 6A, B). In patients with positive responses towards Gn461–475 in IFN-γ ELISPOT assays (IFN-γ ++), Gn465–473 specific T cells were mostly CD45RA+CCR7− and significantly more of a differentiated CD28−CD27− phenotype as compared to IFN-γ− samples. In addition, we found significant differences with regard to the IL-7Rα (CD127), which is crucially involved in maintenance of memory T-cells in the periphery in the absence of cognate antigen [36]. Patients with IFN-γ+ ELISPOT results showed mainly CD127− Gn465−473 T cells, whereas T cells of IFN-γ− patients were mostly CD127+ (Fig. 6A, C). Thus, Gn465−473-specific CD8+ T cells showed a phenotype that is clearly distinct relative to that described for other self-limited diseases such as those caused by influenza A and respiratory syncytial virus but more resembled the pattern associated with latent infections, such as past exposure to CMV [37]. Because a CD28−CD27−CD127− phenotype was previously described to be a result of ongoing antigen-stimulation, as found in latent CMV infection, we next determined the expression of activation markers, such as of KLRG-1, CD69, CD38 and CD25 on Gn465–473 and Influenza A-specific T cells within the seven HLA-B*3501+ ANDV-convalescent (Fig. 6D). No significant differences could be observed between IFN-γ+ and IFN-γ− Gn465–473-specific populations or between Gn465–473 - and Influenza A NP418–426-specific T-cells.

Prospectively high frequencies of Gn-specific memory T cells, anti-N antibodies and neutralizing antibodies
In a next step we assessed whether re-exposure to viral antigens could have led to a boost in the donor's immune response. We therefore compared memory T-cell responses in patients who got infected during recreation (R-patients) with those of residents in endemic areas (E-patients) (Fig. 7A). No significant differences were observed between the two groups in those responses, although endemic patients revealed about double as many Gn-specific memory T-cells than recreational patients (mean 765 vs 361 SFU/106 PBMC) for unclear reasons. However, these results are not in line with the hypothesis of repeated viral exposure in patients who reside in endemic areas, since N - and Gc-specific responses were virtually identical in both groups.

In addition, we identified seven individuals, which had been infected during recreation (R-patients, Fig. 7B, C) and ten individuals residing in endemic areas (E-patients, Fig. 7D, E), for all of which two prospective serum samples were available. The time period between sample 1 and sample 2 was 0.3–6.9 years and 1.2–4.1 years in R - and E-patients, respectively. Surprisingly, in R - and E-patients anti-N titers raised four - to 64-fold between samples 1 and 2 in 4/7 and 5/10 patients, respectively. Most intriguingly, however, also neutralizing antibody (NAb) titers rose two - to eight-fold in 4/7 and 6/10 of R - and E-patients, respectively. Importantly, NAb titers, measured by a blinded worker, increased two - to four-fold in patients R1, R3, R5, E1 and E4 between sample timepoint 1 and 2, although in all cases sample 1 was taken months to years after the acute phase. Taken together, these results suggest that re-exposure to extrinsic, environmental virus is not responsible for the observed rise in NAb titers or high frequencies of memory T cells.
Finally, we sought to prospectively study Gn-specific T-cells in three patients. When Gn465–473-specific T cells were phenotyped over a time period of two years (Fig. 8A–C), no dynamic changes of the CD27− population could be observed, indicating that differentiated Gn465–473-specific T cells are able to stably persist at high frequencies without the need for B7:CD28 - or CD70:CD27-mediated survival signals. However, in patients E9 and E2 (Fig. 8A, C, respectively), Gn465–473-specific T cells actually increased over time, paralleling the two - to eight-fold increase in NAb titers observed in these two individuals (Fig. 7E).

Discussion
Infection with ANDV is the predominant cause for HCPS in South America. Case-fatality rates of currently 36%, person-to-person transmission and the absence of a proven effective antiviral treatment urge the development of a vaccine. Although the protective potential of neutralizing antibodies against the hantavirus surface glycoproteins Gn and Gc, but not the N-protein, was established in vitro [38]–[40] and in animal models [41]–[43], efforts to induce long-lasting neutralizing antibodies in human volunteers have been unsuccessful so far [29],[44] or remain to be proven effective and long lasting [45].
On the other hand, several early reports highlight the importance of lymphocytes for immunity of mice towards hantaviruses, such as HTNV [15]–[17]. Likewise, appearance of virus-specific CD8+ T cells with cytotoxic activity and the ability to produce IFN-γ and TNF-α was associated with clearance of HTNV in newborn mice. In contrast, HTNV infection was not cleared when TNF-α production and cytotoxic activity of specific CD8+ T cells were impaired [18]. In another report, (HTNV-) N-protein-specific CD8+ memory T-cells, induced by a DNA vaccine, conferred partial protection against re-challenge with a vaccinia virus expressing the N-protein [19]. One study in mice showed that N-protein specific T cells rather than antibodies mediated protection and cross-protection upon re-challenge with homologous and heterologous hantaviruses [20]. Finally, ANDV infection of Syrian hamsters - the sole animal model for human HCPS–could be prevented for at least 10 months by previous vaccination with ANDV N-protein [21], again indicating that protection can be achieved independently of neutralizing, Gn/Gc-specific antibodies. Most recently Safronetz et al. confirmed these findings in Syrian hamsters vaccinated with Gn-protein expressing Adenovirus vectors. Interestingly, these animals were protected from lethal ANDV infection independently of neutralizing antibodies and showed no or very low levels of ANDV-RNA up to 9 days after ANDV infection [22]. As for ANDV-infection in man, we recently showed that clearance of ANDV-RNA from peripheral blood cells was closely related to the appearance of cytotoxic CD8+ T cells, but not NAb, in a patient about two months after the acute infection [23].
Taken together, these reports suggest that cytotoxic T cells are crucially involved in clearance and protection from hantaviruses. Conversely, establishment of hantavirus-specific cytotoxic memory CD8+ T cells prior to infection, e.g. by a vaccine, may provide protective, albeit not sterilizing, immunity to the host. However, limited information on human cellular immunity to hantaviruses is available and, to date, only one study addresses ANDV-specific T-cell responses [23].
Using a panel of 310 overlapping peptides spanning the entire N-, Gn - and Gc-protein of ANDV allowed us to study most, existing T-cell epitopes in 78 convalescent survivors of ANDV infection in a non-HLA-restricted manner. In contrast to other reports with a similar approach, the majority of responses were specific for Gn - but not N-protein-derived epitopes. Thus, our results in ANDV-infected patients seem to contradict the current dogma of N-protein being the principal T-cell immunogenic hantavirus antigen [19], [20], [22], [24], [25], [27], [46]-[48]. However, in previous studies, epitope-specific T cells were detected either by in vitro expansion prior to testing or using individual peptides or tetramer complexes for ex vivo detection in a small and HLA-selected patient populations [13], [24]–[28]. Thus, differences in the experimental design, rather than its elevated immunogenicity, may explain, why we found Gn being the immunodominant antigen of ANDV, whereas no single Gn-epitope had been described for other hantaviruses. In fact, 92–96% of the amnio acid sequence of the two Gn-epitopes described herein, are conserved within the PUUV and SNV sequence, respectively. An alternative explanation may derive from the differences between our study and other studies in the timing of T-cell testing after the acute phase or differences in infection kinetics between the different hantaviruses. Specifically, as can be seen in Figure 2D, N-derived epitopes seemed to be relatively predominant up to four years after the acute infection, whereas Gn-derived epitopes were predominant in patients with a longer convalescence phase. In addition, the kinetics of NAb titers in our patients suggest that viral antigen may be present for months or years after the acute infection, a phenomenon which has not been described for other hantaviruses. Thus, differences in epitope avidity and/or precursor expansion over time may have contributed to the relative predominance of Gn-specific T-cells in our study.
Of note, 80% of all patients with detectable T-cell responses recognized Gn-derived epitopes (Fig. 1B). To our surprise, however, responses against Gn were not broad but rather focused to the carboxyterminus of Gn, namely the region of aa 451–480. Interestingly, the cytoplasmic tail of Gn has been shown to contain important virulence factors as it suppresses type 1-interferon responses in infected cells [49],[50]. On the other hand, the carboxyterminal 142 residues of pathogenic (namely of ANDV and HTNV), but not non-pathogenic hantaviruses, prone the C-terminal tail of Gn towards degradation by the proteasome, which then leads to the presentation of epitopes by MHC I molecules to CD8+ T-cells [51]. This mechanism could explain the relative immunodominance of Gn-derived epitopes seen in our study and also may represent a virulence factor of ANDV suggesting that T cells are causative for HCPS. Nonetheless, an early and vigorous cytotoxic T-cell response towards epitopes of the C-terminal Gn may also be able to restrict the virulence of ANDV infection. Indeed, among HLA-B*35-positive patients mild disease outcome seemed to be associated with stronger responses towards the Gn-carboxyterminus than in patients with severe HCPS (Fig. S1B, C). In line with this finding, a recent study in 87 Chilean ANDV-infected patients found that the HLA-B*35 allele was the most frequent allele among patients with mild disease and almost twice as frequent as in patients with severe disease [52]. Although these data seem to contradict previous reports describing both the expression of the HLA-B*35-allele [14] as well as HLA-B*35-restricted T-cell responses [13] as risk factors for severe HCPS by SNV, it remains speculative whether our results indicate an pivotal role for T cells in disease outcome. The size of the memory T-cell pool is only indirectly linked to the effector T-cell response by the original burst size [53] and therefore may not reflect the size and composition of the effector T-cell pool during the acute phase. Moreover, 33% of HLA-B*35-negative patients and 48% of patients without any detectable memory T-cell responses had a history of mild HCPS (data not shown). Likewise, 52% of patients with severe HCPS did not show memory T-cell responses (data not shown). Both seem to argue against an exclusive role of T cells for disease outcome. In addition, other hand, we also showed a discrepancy between ELISPOT and tetramer-derived T-cell frequencies (see above), which indicates the existence of IFN-γ-negative ANDV-specific T-cells. In fact, three of the seven studied HLA-B*3501 positive donors did not show significant responses in initial IFN-γ ELISPOT assays (Fig. 1) but showed substantial numbers of tetramer-positive CD3+CD8+ T cells. This is in line with previous reports comparing determination of T-cell frequencies by ELISPOT and tetramer analysis [35]. In this report tetramer analysis revealed on average ten-fold higher frequencies than IFN-γ ELISPOT assays of T cells specific for a HLA-A2-restricted HIV Gag-derived peptide. This report as well as our data suggests that the vast majority of virus-specific T cells may not readily secrete IFN-γ when stimulated by peptides in ELISPOT assays. It is also possible that these T cells were functional during the acute phase but not during convalescence. Alternatively, differences in the infectious dose (e.g. low versus high infectious dose) or the kinetics and doses of the evolving neutralizing antibodies may interfere with the functional quality of the memory T-cell pool. Taken together, additional studies with HLA-tetramers in acutely ANDV-infected patients will be necessary to better understand the role of HLA-B*35 and T-cell kinetics for ANDV disease outcome.
Another interesting aspect of our study concerns the longevity of the memory T cells in association with their highly differentiated phenotype. After more than 13 years after infection we still detected 564–2152 SFU/106 PBMC by ELISPOT, most of which were directed towards Gn-derived epitopes.
With regards to the longevity of CD8 memory T cells, our results nicely confirm a previous study by Van Epps et al. in PUUV-convalescent patients, in which up to 100–300 SFU/106 PBMC of N-epitope-specific CD8 T cells were found in three patients up to 15 years after acute infection [24],[25],[27]. However, by tetramer analyses we detected still-higher frequencies with up to 16.8% of all CD3+CD8+ T cells proving positive for the single epitope Gn465–473 while displaying a late effector memory phenotype (CD127−CD28−CD27−CCR7−). This phenotype was in line with the cells' ability to readily secrete granzyme B and TNF-α without IL-2. Surprisingly, we also found that up to 5% of tetramer+ cells, which, again in accordance with their CD28+CD27+CCR7+CD127+ phenotype, did not exert any immediate effector functions, such as IFN-γ secretion upon stimulation with their cognate epitope. While these data again show that screening by ELISPOT underestimates the real proportion of Gn-specific T cells [35], it is also tempting to speculate, albeit virtually impossible to prove, that patients with either of the observed phenotypes differ at their stage of immunity towards a possible ANDV re-challenge.
In absence of antigenic stimulation as well as of autocrine or paracrine IL-2 and co-stimulation via CD28:B7 and CD27:CD70 interaction, late effector memory T-cells are heavily prone to apoptosis. However, we herein were able to show in three HLA-B*3501+ patients that peripheral Gn465–473-specifc T cells were maintained in the periphery for at least two years, despite a consistent CD27− phenotype, thereby lacking the receptor for crucial anti-apoptotic signals provided by CD70. While the former perception was that senescent end-differentiated CD28−CD27− T cells were unable to divide, recent evidence suggests, that highly differentiated granzymeB+CD8+ memory T cells are actually dividing upon stimulation equally well as naïve CD8+ T cells [54]. In man, the CD45RA+CD28−CD27−CCR7− late effector memory phenotype has been mainly described in patients with latent CMV infection [37],[55]. In this model repetitive or latent antigen stimulation is supposed to drive CD8+ memory differentiation and/or the recruitment of new memory T-cells. However human hantavirus infections are not known to cause latent or chronic infections and we were not able to detect viral RNA in plasma or peripheral blood cells of patients with sustained and high T-cell responses (data not shown). Also, we failed to detect a fingerprint of recent antigenic stimulation of tetramer-positive cells through assessment of the expression of additional activation markers, including CD69, CD38 and CD25. In addition, expression of IL-7Rα (CD127) was described to be a critical factor for long-term survival of CD8+ memory T cells in absence of their cognate antigen. Since CD127 is usually downregulated upon antigen exposure and rapidly re-expressed after antigen clearance, it is consistent that mainly virus-specific CD127+CD8+ memory T-cells are found in studies on Influenza-, respiratory syncytial virus - and HBV - specific T-cells. By contrast, in persistently HIV-, CMV - or EBV-infected individuals T cells are maintained despite their lack of CD127 expression [56],[57]. Finally, KLRG1 is mainly expressed on antigen-experienced T-cells with immediate effector functions [55]. Thus, considering ANDV a self-limiting transient infection in man, a CD127+KLRG1− phenotype would have been expected years after the infection. However, IFN-γ +, but not IFN-γ −, Gn465–473-specific T cells expressed substantially less CD127 than their Influenza A virus (NP418–426)-specific counterparts, whereas no clear pattern could be observed regarding the KLRG1 expression. However, although no difference could be observed between ANDV (Gn465–473) and Influenza (NP418–426)–specific T-cells with regards to CD25, CD38 and CD69 expression, the lack of CD127 suggests persistent antigenic stimulation in individuals with IFN-γ+ T cells. When employing BLAST, we were not able to identify other organisms that share the amino acid sequence of Gn465–473, thereby making cross-reactivity an unlikely explanation. Consistently, no further serology testing was performed in convalescent patients.
We next hypothesized that residents of endemic areas might have received intermittent antigen-boosters due to viral re-exposure and therefore should show somewhat higher T-cell responses to all viral antigens. However, we did not find significant differences in ANDV-specific T cell numbers when comparing patients who reside in endemic areas and those, which got during recreation got infected in an endemic region (Fig. 7A), while residing in non-endemic areas.
Moreover, in the majority of prospective serum samples of ten patients from endemic regions and of seven patients from non-endemic regions, we surprisingly found an increase in both anti-N and NAb titers despite the fact that the second sample was taken years after the first samples in most cases. The fact that this was also observed in patients who never had returned to endemic regions since their primary ANDV infection, suggests that re-exposure to extrinsic (environmental) virus does not account for high antibody titers and, conversely, not for high ANDV-specific T-cell frequencies. Regarding the increase in NAb titers between sample 1 and 2, we cannot exclude that NAb titers continued to rise after sample 1 was drawn during or shortly after the acute phase. Therefore, it is possible that in these patients (Fig. 7) titers of sample 2 in fact were identical or even lower than the maximum titer achieved during the acute phase. However, NAb titers of 13/17 individuals were still relatively high (≥1∶400) at the timepoint of sample 2, that is 1.2–11.3 years after the acute infection. This argues for continuous antigen-exposure in both E - and R-patients, since NAb titers, in contrast to non-neutralizing antibodies, strictly depend on the presence of their cognate antigen. Specifically, in absence of antigen, murine NAb titers fall below the detection limit after 100–200 days [58]. Most importantly, however, we also found that NAb titers increased two - to four-fold in five patients (R1, R3, R5, E1 and E4, Fig. 7C, E) in which sample 1 was taken months to years after the acute phase. Since not only maintenance [58] but also kinetics of NAb titers heavily depend on the presence of viral surface antigens [59], these results support the hypothesis that re-exposure to viral surface (Gn/Gc) antigen is responsible for high and rising NAb titers in both R - and E-patients. Due to the fundamental differences between R - and E-patients in their risk for re-exposure to extrinsic virus, this is turn suggests that intrinsic viral antigen is responsible for the relative “immune-inflation” and the terminal differentiation of Gn-specific CD8+ T-cells. Thus, it may be that intermittent release of low doses of viral antigen from intrinsic virus (e.g. that never completely cleared from tissue reservoir(s)) is sufficient to maintain and boost of NAb titers and T-cell frequencies, whereas changes in activation marker expression on ANDV-specific T-cells are too short-lived (with the exception of low CD127 expression) to be consistently different (e.g. KLRG1) from that of Influenza A virus-specific T cells. However, as long as viral antigen or genome cannot be detected in convalescent patients as those described herein, the concept of latent or persistent ANDV infection in convalescent patients remains speculative. Future studies should therefore focus on antigen detection in tissues (e.g. surgical or post-mortem specimen) from solid (e.g. lung, kidney) and immuno-privileged (e.g. brain) organs.
Our data suggest that long-lived effector memory T cells can be maintained at high numbers in the periphery over years independently of IL-7. Notably, our findings resemble those found in murine Sendai and Influenza A virus infections, where epitope-specific T-cell clonal expansions occurred in absence of antigen throughout the CD8 memory pool [60]. As in our study, but in contrast to models of persistent infection, clonally expanded effector memory T-cells in these studies retained potent functionality despite their highly differentiated phenotype. Although we could not formally show clonal expansion for all our patients (with the exception of two individuals, see Fig. 8A, C) due to our non-prospective study design, similar underlying, but yet undefined, mechanisms may explain our findings in human ANDV infection.
The induction of a highly differentiated, resting e.g. Gn465–473–specific, memory T-cell subset might be of major interest in the context of vaccine development for several reasons. First, years after infection high numbers of these CCR7− Gn465–473–specific cells remain available for immuno-vigilance in the periphery. Second, as shown, this subset possesses the ability to readily secrete antiviral (e.g. IFN-γ, TNF-α) as well as lytic (granzyme B) effector molecules. Third, although most human studies seem to focus on epitopes restricted to HLA-A*02 because of its wide distribution among the Caucasian population (about 25%, [61]), it should be noted that within the Amerindian population, the frequency of HLA-B*35 positive individuals is about 70% higher than in Caucasians [61]. In fact, 25% (range 22–30%) of the inhabitants in ANDV endemic regions in Southern Chile express the HLA-B*35 and/or the HLA-A*02 allele, respectively [52]. Thus, for this population, HLA-B*35-restricted epitopes, like Gn465–473, might be of similar impact as HLA-A*02-restricted epitopes. However, it first has to be established in future studies (e.g. Syrian hamster models) whether and to which extend Gn-derived T-cell epitopes may contribute to protective immunity. Furthermore, although hantaviruses are not known to mutate, additional epitopes have to be identified in future studies in order to prevent failure of a T-cell based vaccine due to mutations within the Gn465–473 epitope.
Taken together, our results suggest that infection with ANDV may lead to a strong highly differentiated effector memory response. The findings concerning the predominant immunogenicity of ANDV-Gn protein may have implications for the understanding of immunity not only to ANDV, but also to other hantaviruses.
Materials and Methods
Patients, clinical classification and samples
A total of 78 patients were enrolled between 4 months and 13.2 years after hospitalization due to either mild or moderate/severe HCPS. All patients had a previous confirmed hantavirus diagnosis done in Chilean reference laboratories by IgG serology to SNV and ANDV antigens by enzyme-linked immunosorbent assay (ELISA), as previously described [62]–[64].
Mild HCPS was defined by the sole support of the patient by symptomatic therapy, including respiratory support by an oxygen mask. On the other hand, ANDV-infected patients who required intensive care by mechanic ventilation and/or anti-shock treatment with vasoactive drugs were defined as moderate/severe HCPS.
All patients included were Chilean citizens and volunteered to participate without receiving monetary incentive. Prior to enrollment all patients enrolled signed informed consent, which was previously informed by IRB committees of Clínica Alemana de Santiago, the Chilean Ministry of Health and regional IRB committees. Before enrollment, patients were extensively informed about the intention of the study by the local study nurse. Upon enrollment patients did not suffer from any signs of active disease and were only enrolled if considered healthy donors. Samples consisted in 45 cc of peripheral venous blood, using tubes containing Sodium Heparin (BD vacutainer). Samples were shipped within 24 hours to our laboratory and were processed immediately upon receipt. PBMC were isolated by Ficoll-Hypague gradient and fresh PBMC were applied to ELISPOT assays. PBMC, which were not used immediately were cryopreserved in liquid nitrogen.
ELISPOT assays
96-well filterplates (Millipore) were coated with 5 µg/ml of anti-hIFN-γ (Endogen, clone M700A) or 15 µg/ml anti-granzyme B (mabtech, clone GB10) at 4°C overnight one day prior to the assay. For granzyme B assays, prior to coating membranes were activated by incubation of the wells with 15 µl/well of 35% Ethanol for 1 minute. After washing and blocking of the plate, fresh or cryopreserved PBMC or T-cell lines were applied and incubated for 20 hours in an incubator (Nuraire) at 37°C and 5% CO2 in the presence of 310 overlapping 15mer peptides (Mimotopes, Australia) organized in 13 pools of 12 to 44 peptides (final concentration 1 µg/ml, each) of continuous sequence spanning the entire genome of the N and GPC protein of the Chilean ANDV [30]. For mapping experiments, cells were incubated with 10 µg/ml of each individual peptide. As negative controls, corresponding dilutions of DMSO (Sigma) were used, whereas a 1∶100 dilution of PHA (M form, Invitrogen) was used as positive control. After the incubation period, plates were washed and incubated with biotinylated secondary antibodies (IFN-γ: clone M701B, granzyme B: GB11) according to the manufacturer's manual. After incubation with Streptavidine-Alkaline phospahtase (Vector, at 1∶1000 for 2 hours), plates were incubated with NCIP/BPT substrate (BioRad), and analyzed using the ELI.Scan (A.EL.VIS GmbH) analyzing unit. Results were expressed as Spot Forming Units (SFU), representing the numeric difference between specific spots and the spots in the negative control (DMSO).
Focus Reduction Neutralization Test (FRNT)
All FRNT studies were carried out in an approved (C20041018-0267) biosafety level 3 laboratory. Plasma samples from the patient were serially diluted in fourfold increments, mixed with equal volumes of approximately 60 focus forming units (f.f.u.) of a human Chilean virus isolate [65] before incubation on Vero E6 cells, processed and analyzed as described before [66]. The neutralization activity of an antibody was expressed as the highest plasma dilution capable of reducing the number of foci by at least 80%.
Intracellular cytokine staining
Cryopreserved PBMC of four different patients were challenged in vitro for 1.5 hours by a previously determined individual immunogenic peptide (10 µg/ml), the corresponding DMSO dilution or PMA/ionomycin (500/50 ng/ml) in the presence of 1 µg/ml anti-CD49d (clone 9F10) and anti-CD28 (clone CD28.2) (both BD Pharmingen), respectively, and cultured for 4.5 additional hours in the presence of GolgiStop/monensin. Finally, intracellular cytokine staining was performed by fixation, permeabilization of cells and subsequent staining for surface markers and intracellular IFN-γ and TNF-α according to the manufacturer's protocol (BD, California).
Gn461–475-specific T-cell lines
2×105 PBMC/well were stimulated with 10 µg/ml of the Gn461–475 in the presence of 10 ng/ml IL-7 and 300 pg/ml IL-12 (R&D systems). On day 2 after setup and every 3–4 days 10 U/ml and 150 µg/ml IL-15 were added to the culture. On day 7 and 14, T-cells were re-stimulated using irradiated (30Gy) PBMC or irradiated autologous (100Gy) B-LCL. On day 21 T-cells were assayed in IFN-γ ELISPOT assays using truncated peptides as indicated.
Flow cytometry, antibodies and tetramers
Was performed on a 4-colour FACSCalibur (Becton Dickinson) or CyAn (Dako) and using either CellQuest (Becton Dickinson®) or Summit 4.0 (Dako) analyzing software. We used the following antibodies (all BD Pharmingen): CD3-FITC (UCHT1), CD4-FITC (SK3), CD45RA-FITC (HI100), CD27-FITC (M-T271), TNF-α-FITC (Mab11), CD45RO-PE (UCHL1), CD28-PE (L293), CD8-PercP (SK1), IFN-γ-APC (B27), CD3-APC (UCHT1). FITC-, PE - and APC - (MOPC-21) as well as PercP - (X40) conjugated mouse IgG1κ were used as isotype controls. Anti-KLRG-1-Alexa488 was kindly provided by Prof H.P. Pircher (University of Freiburg, Germany). Tetramer complexes were custom-synthesized by the NIAID tetramer facility (Gaithersburg, MD) according to the published protocol (http://research.yerkes.emory.edu/tetramer_core/protocol.html), and Gn465–473:tetramers were either APC - or PE-labeled. All other tetramer complexes were APC-labeled. Table 2 shows a summary of HLA-B*3501 tetramer complexes used in the present study.
HLA typing
The patients were genotyped for the HLA loci A, B, DRB1 and DQB1, using the SSP PCR (Sequence Specific Primer–Polymerase Chain Reaction) technique. Low and high resolution SSP kits from Dynal (Oslo, Norway) and Invitrogen Corporation (USA) were used.
Statistics and positivity criteria
For analysis of ELISPOT results for each patient an unpaired Student's t-test was applied in order to calculate significant results as compared to the internal negative (DMSO) control. To be evaluated as positive a sample (that is response to a individual or a pool of up to 40 ANDV-derived 15mer peptides) had to fulfill three criteria: (i) a significant difference (p<0.05) between sample and negative control, (ii) specific SFU had to be superior of 50/106 PBMC, (iii) value had to be above a cut-off, defined as a mean + 2xSD, which was previously established in 20 healthy controls for each peptide pool.
For differences in frequencies of tetramer cell populations an unpaired Student's t-test was applied. We further studied the association of time since ANDV infection until blood sampling with the T-cell responses against N, Gn and Gc as determined by IFN-γ ELISPOT at the time point of blood sampling. A decreasing response with increasing time since infection corresponds to a loss of T-cell memory in time and is reflected by a negative correlation. We reject the null-hypothesis of no association between time and the IFN-γ-ELISPOT response at the two-sided alpha level of 0.05. Due to skewness of the response data, we log-transformed ELISPOT responses and fitted our linear regression models on the log-transformed responses and assessed the model assumptions by inspecting residual values against time.
Supporting Information
{kind=link}
Zdroje
1. BayardV
KitsutaniPT
BarriaEO
RuedasLA
TinninDS
2004 Outbreak of hantavirus pulmonary syndrome, Los Santos, Panama, 1999–2000. Emerg Infect Dis 10 1635 1642
2. MattarS
ParraM
2004 Serologic evidence of hantavirus infection in humans, Colombia. Emerg Infect Dis 10 2263 2264
3. ToroJ
VegaJD
KhanAS
MillsJN
PadulaP
1998 An outbreak of hantavirus pulmonary syndrome, Chile, 1997. Emerg Infect Dis 4 687 694
4. WeissenbacherMC
CuraE
SeguraEL
HortalM
BaekLJ
1996 Serological evidence of human Hantavirus infection in Argentina, Bolivia and Uruguay. Medicina (B Aires) 56 17 22
5. da SilvaMV
VasconcelosMJ
HidalgoNT
VeigaAP
CanzianM
1997 Hantavirus pulmonary syndrome. Report of the first three cases in Sao Paulo, Brazil. Rev Inst Med Trop Sao Paulo 39 231 234
6. Chile MdSd 2009 El vigía 20. http://epi.minsal.cl/epi/html/frames/frame1.htm
7. VialPA
ValdiviesoF
MertzG
CastilloC
BelmarE
2006 Incubation period of hantavirus cardiopulmonary syndrome. Emerg Infect Dis 12 1271 1273
8. NIAID 2009 List of NIAID Emerging and Re-emerging Diseases. http://www3.niaid.nih.gov/topics/emerging/list.htm
9. PadulaPJ
EdelsteinA
MiguelSD
LopezNM
RossiCM
1998 Hantavirus pulmonary syndrome outbreak in Argentina: molecular evidence for person-to-person transmission of Andes virus. Virology 241 323 330
10. MartinezVP
BellomoC
San JuanJ
PinnaD
ForlenzaR
2005 Person-to-person transmission of Andes virus. Emerg Infect Dis 11 1848 1853
11. FerresM
VialP
MarcoC
YanezL
GodoyP
2007 Prospective evaluation of household contacts of persons with hantavirus cardiopulmonary syndrome in chile. J Infect Dis 195 1563 1571
12. SchmaljohnC
HjelleB
1997 Hantaviruses: a global disease problem. Emerg Infect Dis 3 95 104
13. KilpatrickED
TerajimaM
KosterFT
CatalinaMD
CruzJ
2004 Role of specific CD8+ T cells in the severity of a fulminant zoonotic viral hemorrhagic fever, hantavirus pulmonary syndrome. J Immunol 172 3297 3304
14. KosterF
VigilJ
OlsonD
TerajimaM
EnnisF
2001 Class I, II and III HLA alleles associated with severe hantavirus cardiopulmonary syndrome in the southwest US. Am J Trop Med 65 (Abstr.)
15. NakamuraT
YanagiharaR
GibbsCJJr
GajdusekDC
1985 Immune spleen cell-mediated protection against fatal Hantaan virus infection in infant mice. J Infect Dis 151 691 697
16. NakamuraT
YanagiharaR
GibbsCJJr
AmyxHL
GajdusekDC
1985 Differential susceptibility and resistance of immunocompetent and immunodeficient mice to fatal Hantaan virus infection. Arch Virol 86 109 120
17. AsadaH
TamuraM
KondoK
DohiY
YamanishiK
1988 Cell-mediated immunity to virus causing haemorrhagic fever with renal syndrome: generation of cytotoxic T lymphocytes. J Gen Virol 69 2179 2188
18. ArakiK
YoshimatsuK
LeeBH
KariwaH
TakashimaI
2003 Hantavirus-specific CD8(+)-T-cell responses in newborn mice persistently infected with Hantaan virus. J Virol 77 8408 8417
19. WooGJ
ChunEY
KimKH
KimW
2005 Analysis of immune responses against nucleocapsid protein of the Hantaan virus elicited by virus infection or DNA vaccination. J Microbiol 43 537 545
20. de Carvalho NicacioC
Gonzalez Della ValleM
PadulaP
BjorlingE
PlyusninA
2002 Cross-protection against challenge with Puumala virus after immunization with nucleocapsid proteins from different hantaviruses. J Virol 76 6669 6677
21. Padula June 13–15, 2007 Pathogenic Aspects and Protective Immune Response of Andes Hantavirus. VII International Conference on HFRS, HPS and Hantavirus.
22. SafronetzD
HegdeNR
EbiharaH
DentonM
KobingerGP
2009 Adenovirus vectors expressing hantavirus proteins protect hamsters against lethal challenge with andes virus. J Virol 83 7285 7295
23. ManigoldT
MartinezJ
LazcanoX
YeC
SchwartzS
2008 Case report: T-cell responses during clearance of Andes virus from blood cells 2 months after severe hantavirus cardiopulmonary syndrome. J Med Virol 80 1947 1951
24. Van EppsHL
SchmaljohnCS
EnnisFA
1999 Human memory cytotoxic T-lymphocyte (CTL) responses to Hantaan virus infection: identification of virus-specific and cross-reactive CD8(+) CTL epitopes on nucleocapsid protein. J Virol 73 5301 5308
25. Van EppsHL
TerajimaM
MustonenJ
ArstilaTP
CoreyEA
2002 Long-lived memory T lymphocyte responses after hantavirus infection. J Exp Med 196 579 588
26. LeeKY
ChunE
KimNY
SeongBL
2002 Characterization of HLA-A2.1-restricted epitopes, conserved in both Hantaan and Sin Nombre viruses, in Hantaan virus-infected patients. J Gen Virol 83 1131 1136
27. EnnisFA
CruzJ
SpiropoulouCF
WaiteD
PetersCJ
1997 Hantavirus pulmonary syndrome: CD8+ and CD4+ cytotoxic T lymphocytes to epitopes on Sin Nombre virus nucleocapsid protein isolated during acute illness. Virology 238 380 390
28. TuuminenT
KekalainenE
MakelaS
Ala-HouhalaI
EnnisFA
2007 Human CD8+ T cell memory generation in Puumala hantavirus infection occurs after the acute phase and is associated with boosting of EBV-specific CD8+ memory T cells. J Immunol 179 1988 1995
29. HjelleB
2002 Vaccines against hantaviruses. Expert Rev Vaccines 1 373 384
30. MeissnerJD
RoweJE
BoruckiMK
St JeorSC
2002 Complete nucleotide sequence of a Chilean hantavirus. Virus Res 89 131 143
31. HammarlundE
LewisMW
HansenSG
StrelowLI
NelsonJA
2003 Duration of antiviral immunity after smallpox vaccination. Nat Med 9 1131 1137
32. BurrowsSR
GardnerJ
KhannaR
StewardT
MossDJ
1994 Five new cytotoxic T cell epitopes identified within Epstein-Barr virus nuclear antigen 3. J Gen Virol 75 (Pt 9) 2489 2493
33. BoonAC
de MutsertG
GrausYM
FouchierRA
SintnicolaasK
2002 Sequence variation in a newly identified HLA-B35-restricted epitope in the influenza A virus nucleoprotein associated with escape from cytotoxic T lymphocytes. J Virol 76 2567 2572
34. KleinMR
HammondAS
SmithSM
JayeA
LukeyPT
2002 HLA-B*35-restricted CD8(+)-T-cell epitope in Mycobacterium tuberculosis Rv2903c. Infect Immun 70 981 984
35. GoepfertPA
BansalA
EdwardsBH
RitterGDJr
TellezI
2000 A significant number of human immunodeficiency virus epitope-specific cytotoxic T lymphocytes detected by tetramer binding do not produce gamma interferon. J Virol 74 10249 10255
36. van LeeuwenEM
de BreeGJ
RemmerswaalEB
YongSL
TesselaarK
2005 IL-7 receptor alpha chain expression distinguishes functional subsets of virus-specific human CD8+ T cells. Blood 106 2091 2098
37. AppayV
DunbarPR
CallanM
KlenermanP
GillespieGM
2002 Memory CD8+ T cells vary in differentiation phenotype in different persistent virus infections. Nat Med 8 379 385
38. LiangM
MahlerM
KochJ
JiY
LiD
2003 Generation of an HFRS patient-derived neutralizing recombinant antibody to Hantaan virus G1 protein and definition of the neutralizing domain. J Med Virol 69 99 107
39. DantasJRJr
OkunoY
AsadaH
TamuraM
TakahashiM
1986 Characterization of glycoproteins of viruses causing hemorrhagic fever with renal syndrome (HFRS) using monoclonal antibodies. Virology 151 379 384
40. KochJ
LiangM
QueitschI
KrausAA
BautzEK
2003 Human recombinant neutralizing antibodies against hantaan virus G2 protein. Virology 308 64 73
41. HooperJW
CusterDM
SmithJ
Wahl-JensenV
2006 Hantaan/Andes virus DNA vaccine elicits a broadly cross-reactive neutralizing antibody response in nonhuman primates. Virology 347 208 216
42. HooperJW
CusterDM
ThompsonE
SchmaljohnCS
2001 DNA vaccination with the Hantaan virus M gene protects Hamsters against three of four HFRS hantaviruses and elicits a high-titer neutralizing antibody response in Rhesus monkeys. J Virol 75 8469 8477
43. CusterDM
ThompsonE
SchmaljohnCS
KsiazekTG
HooperJW
2003 Active and passive vaccination against hantavirus pulmonary syndrome with Andes virus M genome segment-based DNA vaccine. J Virol 77 9894 9905
44. McClainDJ
SummersPL
HarrisonSA
SchmaljohnAL
SchmaljohnCS
2000 Clinical evaluation of a vaccinia-vectored Hantaan virus vaccine. J Med Virol 60 77 85
45. DongGM
HanL
AnQ
LiuWX
KongY
2005 Immunization effect of purified bivalent vaccine to haemorrhagic fever with renal syndrome manufactured from primary cultured hamster kidney cells. Chin Med J (Engl) 118 766 768
46. de Carvalho NicacioC
SallbergM
HultgrenC
LundkvistA
2001 T-helper and humoral responses to Puumala hantavirus nucleocapsid protein: identification of T-helper epitopes in a mouse model. J Gen Virol 82 129 138
47. ParkJM
ChoSY
HwangYK
UmSH
KimWJ
2000 Identification of H-2K(b)-restricted T-cell epitopes within the nucleocapsid protein of Hantaan virus and establishment of cytotoxic T-cell clones. J Med Virol 60 189 199
48. WangM
WangJ
ZhuY
XuZ
YangK
2009 Cellular immune response to Hantaan virus nucleocapsid protein in the acute phase of hemorrhagic fever with renal syndrome: correlation with disease severity. J Infect Dis 199 188 195
49. AlffPJ
SenN
GorbunovaE
GavrilovskayaIN
MackowER
2008 The NY-1 hantavirus Gn cytoplasmic tail coprecipitates TRAF3 and inhibits cellular interferon responses by disrupting TBK1-TRAF3 complex formation. J Virol 82 9115 9122
50. AlffPJ
GavrilovskayaIN
GorbunovaE
EndrissK
ChongY
2006 The pathogenic NY-1 hantavirus G1 cytoplasmic tail inhibits RIG-I - and TBK-1-directed interferon responses. J Virol 80 9676 9686
51. SenN
SenA
MackowER
2007 Degrons at the C terminus of the pathogenic but not the nonpathogenic hantavirus G1 tail direct proteasomal degradation. J Virol 81 4323 4330
52. FerrerCP
VialPA
FerresM
GodoyP
CuizaA
2007 Genetic susceptibility to Andes Hantavirus: Association between severity of disease and HLA alieles in Chilean patients. Revista Chilena de Infectologia 24 351 359
53. KaechSM
WherryEJ
AhmedR
2002 Effector and memory T-cell differentiation: implications for vaccine development. Nat Rev Immunol 2 251 262
54. BannardO
KramanM
FearonDT
2009 Secondary replicative function of CD8+ T cells that had developed an effector phenotype. Science 323 505 509
55. ThimmeR
AppayV
KoschellaM
PantherE
RothE
2005 Increased expression of the NK cell receptor KLRG1 by virus-specific CD8 T cells during persistent antigen stimulation. J Virol 79 12112 12116
56. PaiardiniM
CervasiB
AlbrechtH
MuthukumarA
DunhamR
2005 Loss of CD127 expression defines an expansion of effector CD8+ T cells in HIV-infected individuals. J Immunol 174 2900 2909
57. BengschB
SpangenbergHC
KerstingN
Neumann-HaefelinC
PantherE
2007 Analysis of CD127 and KLRG1 expression on hepatitis C virus-specific CD8+ T cells reveals the existence of different memory T-cell subsets in the peripheral blood and liver. J Virol 81 945 953
58. OchsenbeinAF
PinschewerDD
SierroS
HorvathE
HengartnerH
2000 Protective long-term antibody memory by antigen-driven and T help-dependent differentiation of long-lived memory B cells to short-lived plasma cells independent of secondary lymphoid organs. Proc Natl Acad Sci U S A 97 13263 13268
59. PinschewerDD
PerezM
JeetendraE
BachiT
HorvathE
2004 Kinetics of protective antibodies are determined by the viral surface antigen. J Clin Invest 114 988 993
60. ElyKH
AhmedM
KohlmeierJE
RobertsAD
WittmerST
2007 Antigen-specific CD8+ T cell clonal expansions develop from memory T cell pools established by acute respiratory virus infections. J Immunol 179 3535 3542
61. MarshS
ParhamP
BarberL
2000 The HLA Facts book. 398
62. PadulaPJ
RossiCM
Della ValleMO
MartinezPV
ColavecchiaSB
2000 Development and evaluation of a solid-phase enzyme immunoassay based on Andes hantavirus recombinant nucleoprotein. J Med Microbiol 49 149 155
63. RossiC, KT
1999 Enzyme-linked immunosorbent assay (ELISA)
LeeHW, CC
SchmaljohnCS
Manual of Hemorrhagic Fever with Renal Syndrome and Hantavirus Pulmonary Syndrome Seoul WHO Collaborating Center for Virus Reference and Research 87 91
64. FeldmannH
SanchezA
MorzunovS
SpiropoulouCF
RollinPE
1993 Utilization of autopsy RNA for the synthesis of the nucleocapsid antigen of a newly recognized virus associated with hantavirus pulmonary syndrome. Virus Res 30 351 367
65. GalenoH
MoraJ
VillagraE
FernandezJ
HernandezJ
2002 First human isolate of Hantavirus (Andes virus) in the Americas. Emerg Infect Dis 8 657 661
66. BharadwajM
NofchisseyR
GoadeD
KosterF
HjelleB
2000 Humoral immune responses in the hantavirus cardiopulmonary syndrome. J Infect Dis 182 43 48
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2010 Číslo 2
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- Caspase-1 Activation via Rho GTPases: A Common Theme in Mucosal Infections?
- Kaposi's Sarcoma Associated Herpes Virus (KSHV) Induced COX-2: A Key Factor in Latency, Inflammation, Angiogenesis, Cell Survival and Invasion
- IL-1β Processing in Host Defense: Beyond the Inflammasomes
- Reverse Genetics in Predicts ARF Cycling Is Essential for Drug Resistance and Virulence
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy