
Současný vývoj v taxonomii dermatofytů a doporučení pro pojmenovávání klinicky významných druhů
Recent Advances in Taxonomy of Dermatophytes and Recommendations for Using Names of Clinically Important Species
Taxonomy of dermatophytes has changed significantly in the past and it has never been stable.
Last extensive changes appeared after revision of the group by molecular genetic methods. This paper introduces the advances in taxonomy of dermatophytes and helps to unify the species names in use among the Czech laboratories to simplify their communication and comparison of the results with foreign literature. The complicated species concept of dermatophytes that might be applied differently and might influence the number of accepted species is discussed in the beginning. The last part is dedicated to future changes related to a new edition of the International Code of Nomenclature for algae, fungi, and plants namely to termination of a dual nomenclature of fungi and choosing the one correct name for each species.
Key words:
Atrhroderma – species concept – phylogeny – the International Code of Nomenclature – one fungus one name – polyphasic approach – Trichophyton interdigitale – Trichophyton mentagrophytes
Autoři:
V. Hubka 1,2; A. Čmoková 1; M. Skořepová 3; N. Mallátová 4; S. Dobiášová 5; P. Lysková 6; K. Mencl 7; H. Janouškovcová 8; V. Buchta 9; T. Větrovský 10; M. Kolařík 1,2
Působiště autorů:
Katedra botaniky, Přírodovědecká fakulta, Univerzita Karlova v Praze
vedoucí katedry doc. RNDr. Yvonne Němcová, Ph. D.
; Laboratoř genetiky a metabolismu hub, Mikrobiologický ústav, Akademie věd České republiky, v. v. i., Praha
vedoucí laboratoře Mgr. Miroslav Kolařík, Ph. D.
2; Centrum pro dermatomykózy, Dermatovenerologická klinika 1. lékařské fakulty Univerzity Karlovy
a Všeobecné fakultní nemocnice v Praze, přednosta kliniky prof. MUDr. Jiří Štork, CSc.
3; Pracoviště parazitologie a mykologie, Centrální laboratoře Nemocnice České Budějovice, a. s., České Budějovice
ředitel nemocnice MUDr. Břetislav. Shon
4; Oddělení bakteriologie a mykologie, Centrum klinických laboratoří, Zdravotní ústav se sídlem v Ostravě
vedoucí oddělení RNDr. Vladislav Holec
5; Laboratoř lékařské mykologie, oddělení parazitologie, mykologie a mykobakteriologie Praha
Zdravotní ústav se sídlem v Ústí nad Labem, Praha, vedoucí oddělení RNDr. Zuzana Hůzová
6; Oddělení klinické mikrobiologie, Pardubická krajská nemocnice, a. s., Pardubice
primář oddělení MUDr. et Mgr. Eva Zálabská, Ph. D.
7; Ústav mikrobiologie, Fakultní nemocnice Plzeň, přednosta ústavu RNDr. Karel Fajfrlík Ph. D.
8; Ústav klinické mikrobiologie Fakultní nemocnice a Lékařské fakulty Univerzity Karlovy, Hradec Králové
přednosta ústavu doc. RNDr. Vladimír Buchta, CSc.
9; Laboratoř environmentální mikrobiologie, Mikrobiologický ústav, Akademie věd České republiky, v. v. i., Praha
vedoucí laboratoře RNDr. Petr Baldrian, Ph. D.
10
Vyšlo v časopise:
Čes-slov Derm, 89, 2014, No. 4, p. 151-165
Kategorie:
Souborné referáty (doškolování lékařů)
Souhrn
Taxonomie dermatofytů prošla v minulosti řadou výrazných změn a nikdy nebyla příliš stabilní. Poslední rozsáhlejší změny proběhly v souvislosti s revizí skupiny molekulárně genetickými metodami. Hlavním cílem této práce je seznámit čtenáře s pokroky v taxonomii dermatofytů a prostřednictvím seznamu doporučených jmen sjednotit pojmenovávání druhů napříč českými pracovišti. To by mělo nejen usnadnit komunikaci, ale také porovnávání výsledků se zahraniční literaturou. Jedním z doporučení je například ukončení používání jména Trichophyton mentagrophytes v klinické praxi. V úvodu textu se diskutuje o problematickém konceptu druhu u dermatofytů, jehož rozdílná aplikace může mít velký vliv na počet uznávaných druhů. Závěrečná část je věnována budoucím změnám souvisejícím s novým vydáním Mezinárodního kódu nomenklatury řas, hub a rostlin. Jedná se především o ukončení duální nomenklatury hub a vybrání jediného správného jména pro každý druh. Obecně se dá říci, že je vhodné preferovat jméno nepohlavního stadia (Trichophyton, Microsporum, Epidermophyton), s výjimkou případů, kde toto jméno není k dispozici (např. Arthroderma benhamiae).
Klíčová slova:
Arthroderma – druhový koncept – fylogeneze – Mezinárodní nomenklatorický kód – jedna houba, jedno jméno – polyfázický přístup – Trichophyton interdigitale – Trichophyton mentagrophytes
ÚVOD A HISTORICKÉ SOUVISLOSTI
Taxonomie dermatofytů byla od svých počátků ve středu zájmu mikrobiologů, protože se zabývá významnými původci povrchových infekcí člověka a zvířat. Jako dermatofyty se tradičně označují rody Trichophyton, Microsporum a Epidermophyton. Zmíněná rodová jména se v duálním názvosloví (pohlavní a nepohlavní stadium nese odlišné jméno) používají pro nepohlavní stadium (anamorfu). Pro pohlavní stadium (teleomorfu) je dnes všeobecně přijímáno rodové jméno Arthroderma [113]. V průběhu svého formování prošla taxonomie dermatofytů velkými změnami a nikdy nebyla příliš stabilní. Z velké části se na tom podílela morfologická variabilita některých druhů a neúspěšná snaha vymezit základní příbuzenské vztahy uvnitř skupiny, dokonce i na rodové úrovni, kde molekulární fylogeneze přináší jiné výsledky než morfologie [34]. Koncept druhu u dermatofytů měl vždy svá specifika, zejména díky zohlednění klinického a epidemiologického aspektu. Taxonomie je totiž stále tvořena především pro její uživatele, kterým záleží na stabilitě a srozumitelnosti.
Většina klinicky významných druhů dermatofytů byla popsána v poměrně dávné minulosti, kdy ještě neexistovala pravidla a doporučení pro popis druhů zakotvená v Mezinárodním kódu nomenklatury řas, hub a rostlin (dále jen Kód), poprvé uveřejněném v roce 1905. Řada dnes dobře známých jmen je podložených jen velmi stručným popisem bez obrazové a herbářové dokumentace. Popis některých druhů (zvláště jmen navržených kolem poloviny 19. století) nebyl navíc založen ani na kultuře houby, ale jen na klinickém obrazu a vzorku přímé mikroskopie z postiženého pacienta (např. T. tonsurans). Z těchto důvodů se často nezachoval žádný původní materiál, dokonce ani kresby znázorňující typické znaky rodů a druhů dermatofytů, jak je známe dnes. Velkým milníkem v taxonomii dermatofytů proto byla kultivace původců dermatomykóz na agarových médiích, s možností sledovat makro - a mikromorfologii subkultur [87], která byla základem dalšího rozvoje taxonomie této skupiny hub.
Pro dermatofyty bylo v minulosti navrženo více než 500 nadbytečných jmen (viz databáze MycoBank; www.mycobank.org), která dnes nejsou používána. Velká část z nich však byla opuštěna ještě před érou molekulární biologie. Kvůli neexistujícímu původnímu materiálu, bylo nutné při revizi dermatofytů stanovit pro velkou část druhů nový typový materiál (neotyp), který zafixoval používání jmen a zamezil jiným výkladům v budoucnosti [29, 30, 27]. Některá rozhodnutí o synonymizaci druhů na základě prvních molekulárně genetických studií byla ale ukvapená (např. synonymizace T. equinum a T. tonsurans) a podtrhují význam polyfázického přístupu, tj. nutnosti brát v úvahu výsledky několika nezávislých metod před učiněním definitivního rozhodnutí.
Navzdory tomu, jak je medicína konzervativním oborem, je třeba s určitým časovým odstupem reflektovat změny, které v taxonomii hub probíhají. U kontroverzních a rozsáhlých změn by měl být časový odstup dostatečný (např. 10 let a více), aby byl dostatek času pro diskusi a obhájení změn ve vědecké a uživatelské komunitě, a nedocházelo tak k nadměrnému zatěžování uživatelů (např. klinických mikrobiologů) změnami. Většina zásadních taxonomických změn u dermatofytů byla učiněna již v letech 1999 a 2000. Tyto změny byly z velké části potvrzeny, zavrženy, či upřesněny v dalších nezávislých studiích, a mohou tedy být bez významnějšího rizika dalších brzkých změn začleněny do praxe. Stále ale existuje několik klinicky významných druhů, či druhových komplexů, kde zbývá taxonomické otázky dořešit. Velké změny v pojmenovávání druhů jsou nyní spojeny se zaváděním aktuálních pravidel uveřejněných v novém Kódu, které se dotkly nejcitelněji právě hub [74]. Jde hlavně o ukončení používání duální nomenklatury a zvolení jediného platného jména, které v sobě obsáhne celý životní cyklus houby.
Cílem této práce je shrnout vývoj v taxonomii dermatofytů během posledních zhruba dvou desetiletí a nastínit budoucí změny související se zaváděním jediného jména pro jednu houbu. Práce předkládá přehled v současnosti uznávaných jmen dermatofytů a sjednocuje jejich užívání napříč českými mykologickými a klinickými pracovišti, což by mělo přispět k jednoduššímu dorozumění a také snadnějšímu porovnání výsledků mezi publikacemi.
CO JE DRUH U DERMATOFYTŮ?
Definice druhu je jednou ze základních, ale zároveň nejvíce problematických otázek taxonomie dermatofytů a aplikace různých přístupů má velký dopad na počet uznávaných druhů. Pravděpodobný model evoluce dermatofytů předpokládá, že geofilní dermatofyty s rezervoárem v půdě jsou nejpůvodnější. Dlouhodobým kontaktem dermatofytů rozkládajících keratin v příbytcích živočichů s jejich obyvateli se vysvětluje adaptace těchto druhů na zvířecího hostitele (zoofilní dermatofyty) [26]. Pohlavní proces geofilních druhů a omezeného počtu zoofilních druhů probíhá často přímo v norách a hnízdech živočichů, kde je stálý dostatek keratinu [41, 109]. Dalším krokem ve vývoji dermatofytů, byla možnost přenosu zoofilních (převážně synantropních) druhů na člověka a jejich přizpůsobení se na člověka jako hlavního hostitele (antropofilní dermatofyty). Přechod k parazitickému způsobu života u antropofilních a některých zoofilních druhů (hlavně patogenů domestikovaných zvířat), vedl ke ztrátě, nebo výraznému omezení pohlavního rozmnožování [32], protože druhy ztratily prostředí vhodné k pohlavnímu procesu, který nikdy nebyl prokázán přímo na živém hostiteli [109]. Tyto druhy se navíc vyvinuly v evolučně nedávné době, genetické mezidruhové rozdíly jsou často nevýrazné [32] a také reprodukční bariéra mezi některými druhy nemusí být ještě zcela vytvořena.
Morfologický koncept druhu zahrnující makro - a mikromorfologii kultur dal tradiční, ale hrubý základ systému dermatofytů, jak ho známe dnes [19]. Postupně byly přidávány různé fyziologické testy jako perforace vlasu [2], nutriční testy [23], ureázový test [81] a další [100]. Tyto testy dále zpřesnily rozlišení některých druhů, zároveň však podpořily popsání některých nadbytečných druhů. Problematické jsou z hlediska tohoto konceptu atypické izoláty (nesporulující, s degenerovanou morfologií), které se u dermatofytů poměrně běžně vyskytují (v dnešní době často v souvislosti s léčbou předcházející odběru na kultivaci).
Ekologický koncept bere v úvahu především přirozený rezervoár druhů, a dělí tak dermatofyty na druhy geofilní a primárně patogenní (antropofilní a zoofilní) [24]. Zohlednění tohoto dělení je výhodné také pro klinickou praxi, kde již sama identifikace druhu napovídá o možných zdrojích infekce. Navíc ekologie mnohem lépe než morfologie odráží fylogenezi dermatofytů a druhy s obdobnou ekologií jsou až na výjimky fylogeneticky příbuzné (obr. 1) [34]. Spektrum hostitelů některých dermatofytů může být poměrně široké, jiné druhy jsou naopak specializované na jediného hostitele. U některých druhů je ale třeba hostitelské spektrum ještě detailněji zmapovat podle nově revidované taxonomie (viz dále). Řada hostitelsky specializovaných druhů navíc na hostiteli preferenčně napadá jen určité anatomické lokalizace a infekce se projevují různými klinickými formami (tinea capitis, t. corporis, t. pedis, apod.).

Biologický koncept chápe druh jako skupinu jedinců, kteří jsou schopní se potenciálně křížit a zároveň vykazují reprodukční izolaci od zástupců ostatních biologických druhů [73]. Pokud vznikají mezidruhoví hybridi, pak mají sníženou životaschopnost, nebo nejsou fertilní (může se projevit až u potomků vyšších generací) [12]. Většina dermatofytů je heterothalických (strategie rozmnožování dvěma oddělenými pohlavími) a opačně ladění jedinci jsou nositeli různého párovacího typu, který se arbitrárně značí + nebo -, či MAT1-1 a MAT1-2 podle přítomnosti konkrétního genu zapojeného v pohlavním procesu. Při vytvoření vhodných podmínek může být při párovacích pokusech se dvěma opačně laděnými izoláty indukována tvorba pohlavního stadia in vitro.
Biologický koncept je dobře aplikovatelný na geofilní dermatofyty [15, 79, 98, 99], u kterých odpovídá výsledkům fylogenetických studií [14, 46] a zohledňuje i morfologické rozdíly mezi druhy. Obtížněji použitelný je u některých primárně patogenních druhů, které se až na výjimky pohlavního rozmnožování „vzdaly“ kvůli patogennímu způsobu života. Genetická struktura těchto druhů je často klonální a najdeme u nich pouze jediný párovací typ, jak bylo zjištěno např. u T. rubrum [116], antropofilních kmenů T. interdigitale [102], T. tonsurans [96], T. equinum [96] a T. verrucosum [56]. Výjimkami jsou některé druhy uvnitř někdejšího komplexu T. mentagrophytes, jejichž pohlavní stadia jsou nazývána jako Arthroderma vanbreuseghemii (anamorfa T. interdigitale), A. benhamiae (Trichophyton sp.) a A. simii (T. simii). Tyto druhy si zřejmě zachovávají saprofytický cyklus v norách zvířat, kde dochází k pohlavnímu procesu. Napovídá tomu jejich neklonální genetická struktura, rovnoměrné rozložení párovacích typů a ochotná tvorba plodnic v párovacích pokusech in vitro [60, 61, 96, 106, 105].
Při velkém úsílí se při párovacích experimentech in vitro daří vytvářet mezidruhové hybridy mezi výše zmíněnými pohlavními druhy navzájem [64, 62, 63], nebo dokonce mezi některým z klonálních druhů a druhem sexuálním (např. T. rubrum a T. simii) [4]. Pokud by byl biologický koncept druhu striktně aplikován podle výsledků těchto experimentů, byl by zcela destruován koncept zoofilních a antropofilních druhů Trichophyton, jak ho dnes známe. U řady klonálních druhů je ale téměř jisté, že v přirozeném prostředí pohlavní proces neprobíhá a jednotlivé druhy si udržují svá specifika i přes teoretickou možnost křížení, která může být úspěšná v laboratorních podmínkách [32].
Fylogenetický koncept druhu vychází z monofylie, tedy ze situace, kdy druh zahrnuje všechny potomky společného předka. Druhy jsou dnes vymezovány nejčastěji na základě výsledků fylogenetických analýz počítaných z molekulárně genetických dat. Jednou z nejpoužívanější aplikací tohoto přístupu u hub je v současnosti tzv. GCPSR koncept (genealogical concordance phylogenetic species recognition) [5], který k vymezení druhu využívá DNA sekvence několika nezávislých genů. Konstrukce několika fylogenezí ze sekvencí různých genů v praxi umožní odhalit rekombinaci uvnitř pohlavně se rozmnožujících populací a určit hranice druhů. Hlavní omezení v použití tohoto konceptu u dermatofytů tedy opět nacházíme u asexuálních druhů, kde k rekombinaci nedochází [32] a každá genetická změna nalezená u některé linie by mohla být rozpoznána jako samostatný druh.
Všechny výše zmíněné koncepty mají určité výhody i nevýhody, a proto je při určování mezidruhových hranic výhodné používat kombinované přístupy. Již v minulosti se často užíval přístup kombinující morfologii, fyziologii a křížící experimenty [52, 83]. Dnes prosazovaný polyfázický přístup, bere v úvahu nejen výsledky fylogenetických studií, ale i morfologii, fyziologii a ekologii druhů. Navržení nového druhu u geofilních druhů by mělo být doplněno i křížícími experimenty s nejbližšími příbuznými druhy [10, 34, 42, 43, 46].
SOUČASNÝ POHLED NA TAXONOMII DERMATOFYTŮ A DUÁLNÍ NÁZVOSLOVÍ
Již první fylogenetické studie na skupině dermatofytů potvrdily, že se jedná o jednotnou a dobře podpořenou skupinu (monofyletickou) uvnitř čeledi Arthrodermataceae [26, 36, 68]. Morfologický koncept anamorfních rodů Trichophyton, Microsporum a Epidermophyton však neodpovídá fylogenetickým datům (viz obr. 1) [26] a druhy klasifikované jako Trichophyton a Microsporum tvoří několik oddělených linií (polyfyletické). Rod Trichophyton v užším slova smyslu (Trichophyton s. str.) zahrnuje všechny primárně patogenní druhy Trichophyton (viz obr. 1), naopak všechny ostatní druhy Trichophyton jsou geofilní a klinicky významný je jen malý počet z nich (tab. 1). Podobně i patogenní druhy Microsporum (Microsporum s. str.) tvoří oddělenou skupinu od druhů geofilních, které navíc tvoří dvě fylogeneticky oddělené skupiny (viz obr. 1) [28, 34]. Jak je zřejmé z obrázku 1, jednotlivé skupiny podpořené fylogenetickou analýzou sdílí i stejnou ekologii.

Současná taxonomie dermatofytů stále používá tzv. duální názvosloví, které bylo zakotveno i v několika předchozích vydáních Kódu, zatímco nová verze již duální názvosloví nepodporuje [74]. V praxi se duální názvosloví týká hub s komplexním životním cyklem, který zahrnuje jak pohlavní, tak nepohlavní stadium (pleomorfní houby). V tomto systému musela mít obě stadia odlišné jméno. Duální systém byl vytvořen před érou molekulární genetiky a měl nesporné výhody pro tehdejší taxonomii, která byla založená především na morfologii a ekologii. Významná část hub (např. rostlinných patogenů) tvoří pohlavní i nepohlavní stadium odděleně na různých substrátech a za různých podmínek, a proto v minulosti nebylo spojení mezi oběma stadii známo, a tak byla popsána pod různými rodovými jmény. Duální systém se stal nadbytečným s všeobecnou dostupností molekulárních metod, protože pomocí nich může být téměř jakákoliv houba (včetně druhů sterilních in vitro) zařazena na správnou pozici ve fylogenetickém systému a tedy mohou být správně určeny vztahy mezi různými stadii životního cyklu. Současný Kód proto podporuje již pouze jediné jméno pro jeden druh houby, které obsáhne celý její životní cyklus [74].
Konkrétní kroky směřující k používání jediného jména pro každý druh zatím u dermatofytů nebyly učiněny a stále je tedy aktuální duální názvosloví. Pro druhy dermatofytů, kde je známo jen nepohlavní stadium, většinou není zapotřebí žádných doporučení ohledně používání konkrétních jmen. U druhů, kde je známo pohlavní stadium, existují většinou dvě platná jména, která mají podle duálního systému stejnou váhu. Již nyní ale lze doporučit k běžnému používání jen některá jména, která odpovídají současné taxonomii dermatofytů a jejich užívání navíc redukuje změny provedené v budoucnu na minimum. Těmi jsou u dermatofytů hlavně jména nepohlavních stadií. Z kroků, které již byly podniknuty je zřejmé, že jméno Arthroderma vztahující se k pohlavnímu stadiu, nebude nadále užíváno, a proto lze již dnes jeho používání minimalizovat (kromě případů, kde nepohlavní stadium nemá jméno, nebo je jeho název méně přesný, viz dále).
Tabulka 1 obsahuje seznam doporučených jmen a těch, která lze používat jako alternativní. Alternativní jméno je vhodné používat v těch případech, kde několik různých biologických druhů (např. A. quadrifidum, A. lenticulare a A. insingulare) má pro nepohlavní stadium identické jméno (T. terrestre). V případě T. terrestre se však podle našich výsledků [44] v klinickém materiálu českých pacientů vyskytoval vždy druh A. quadrifidum, je ale možné, že v jiných geografických oblastech se mohou vyskytovat i jiné druhy tohoto komplexu. V takovém případě je vhodné uvádět obě jména např. ve formě „T. terrestre (A. quadrifidum)“, dokud nebude definitivně zvoleno vhodné jméno pro všechny tři druhy podle výsledků fylogenetických studií a nových pravidel Kódu (viz dále).
TAXONOMICKÉ ZMĚNY U KLINICKY VÝZNAMNÝCH DRUHŮ
Následující odstavce se týkají především změn, které byly učiněny v posledních dvou desetiletích s významným přispěním molekulárně genetických metod. Pro hrubou orientaci jsou užívána jména druhových komplexů, která v taxonomii dermatofytů nesou název fylogeneticky nejbližšího pohlavního stadia (viz obr. 1), i když pro klinickou praxi nemají tato označení velký význam. Dnes je uznáváno zhruba 50–60 druhů dermatofytů a tento počet je pravděpodobně téměř konečný kvůli velmi intenzivnímu výzkumu trvajícímu bezmála dvě století. Z tohoto počtu je zhruba 35 druhů známo z lidského klinického materiálu (viz tab. 1, obr. 1), ale podstatná část je vzácná, někdy známá jen ze specifických endemických oblastí, či několika málo nálezů. Během probíhající epidemiologické studie dermatofytóz v ČR byla v klinickém materiálu od českých pacientů odhalena druhová diverzita čítající 15 druhů [44, 70]. Jedná se pravděpodobně o výčet všech hlavních původců dermatofytóz v ČR, další vzácné druhy mohou být zachyceny při dlouhodobějším výzkumu, nebo jako importované nákazy. Detailnější studie dermatofytóz u zvířat na našem území chybí, ale dá se očekávat, že by spektrum druhů dále obohatily.
Z molekulárně genetických dat jasně vyplynulo, že T. mentagrophytes je komplex několika zoofilních druhů (hostitelé všech zoofilních druhů jsou shrnuti v tabulce 2). Pohlavní stadia A. vanbreuseghemii a A. benhamiae, která byla v minulosti obě přisuzována tomuto druhu, jsou od sebe fylogeneticky vzdálená a reprezentují odlišné druhy (viz obr. 1) [26, 27]. Poměrně překvapivé bylo zjištění, že A. simii je pro T. mentagrophytes s. str. nejpříbuznějším, pohlavně se rozmnožujícím druhem, přestože již v minulosti Weitzman a Padhye [112] diskutovali o blízkosti, či dokonce identitě těchto druhů. Do komplexu A. simii patří kromě zoofilního T. mentagrophytes také blízce příbuzný, i když ekologicky a morfologicky odlišný, antropofilní druh T. schoenleinii [34, 82]. Endemickými oblastmi výskytu obou druhů jsou dnes především Blízký a Střední východ, Afrika a Austrálie [6, 18, 34, 82]. Příbuznost druhů potvrzuje i podobná klinická manifestace infekcí, oba jsou mimo jiné schopné vyvolat infekce typu favus, T. schoenleinii u člověka a T. mentagrophytes u myší (kmeny dříve označované jako T. quinckeanum) a velbloudů (kmeny dříve označované jako T. sarkisovii a T. langeronii). Na našem území se T. mentagrophytes přirozeně nevyskytuje, a proto není vhodné jméno v klinické praxi nadále používat (možnost importu infekce z endemických oblastí ale samozřejmě existuje). Druhy v minulosti klasifikované morfologicky jako T. mentagrophytes a jeho variety (v pojetí jaké navrhoval např. Emmons et al. [20]) byly podle fylogenetických studií rozděleny mezi minimálně tři druhy (T. mentagrophytes s. str., T. interdigitale a T. erinacei). Většinu zoofilních infekcí, které byly i u nás tradičně přisuzovány T. mentagrophytes, působí T. interdigitale [44], který patří do komplexu A. vanbreuseghemii (viz níže). T. interdigitale v dnešním pojetí zahrnuje kmeny jak zoofilního, tak lidského původu a je v našich podmínkách velmi běžným druhem. Zoofilní kmeny jsou přitom morfologicky velmi podobné T. mentagrophytes s. str. podle nového konceptu (neotyp byl stanoven v roce 1999 [27]), i když se jedná o ekologicky a fylogeneticky odlišné druhy. Zbývající izoláty původem především z ježků, dříve klasifikované jako T. mentagrophytes var. erinacei, jsou dnes řazeny do samostatného druhu T. erinacei uvnitř komplexu A. benhamiae (viz dále). Vlastní T. simii (teleomorfa A. simii), podle kterého je pojmenován celý komplex, je poměrně málo prozkoumaný druh, z minulosti známý hlavně z primátů v Indii a přilehlých státech a také od cestovatelů, kteří navštívili tyto oblasti [52]. Ochotná tvorba pohlavního stadia v křížících experimentech upomíná spíše na některé druhy geofilních dermatofytů nebo dermatofyty napadající savce žijící v norách [101]. Je tedy možné, že primární hostitel tohoto druhu ještě nebyl objeven. Sporadické nálezy T. simii jsou známy z celého světa (USA, Brazílie, Egypt, Německo, Írán aj.) [7] a v posledních letech byl druh prokázán také z půdy a u evropských pacientů, kteří nikdy nevycestovali do zahraničí [7, 6], což vzbudilo diskuse o možné geofilní ekologii druhu [7, 34].

Komplex A. vanbreuseghemii zahrnuje klinicky významné antropofilní a zoofilní druhy T. interdigitale, T. tonsurans a T. equinum (viz obr. 1). Křížícími pokusy založenými na kmenech identifikovaných molekulárně genetickými metodami bylo zjištěno, že A. vanbreuseghemii je pohlavní stadium druhu T. interdigitale. Schopnost pohlavně se rozmnožovat si však zachovaly jen izoláty zvířecího původu, kdežto lidské izoláty reprezentují jen klon odvozený od zoofilních kmenů, který ztratil schopnost tvorby pohlavního stadia. Všechny izoláty mají jen jediný párovací typ MT+ a s referenčními párovacími kmeny A. vanbreuseghemii tvoří plodnice bez askospór [105]. Antropofilní kmeny T. interdigitale vykazují často typickou morfologii, která odpovídá dřívějšímu konceptu T. interdigitale, a zahrnuje následující znaky: bílé, vzdušné kolonie se žlutou spodinou, mikrokonidie jsou kulovité až klavátní, makrokonidie a spirální vlákna většinou chybí, nebo jsou málo časté (obr. 2). Zoofilní kmeny T. interdigitale mají nejčastěji kolonie granulární, v odstínech okrové a hnědé s hnědou nebo hnědočervenou spodinou, mikrokonidie jsou nejčastěji kulovité, doutníkovité makrokonidie a spirální vlákna jsou většinou přítomna (viz obr. 2). Nezřídka se ale vyskytují i přechodné varianty mezi oběma typy a často i jediný kmen produkuje oba typy morfologie v téže kolonii v závislosti na inokulu (viz obr. 2) [40, 76]. Morfologie zoofilních kmenů T. interdigitale asi nejlépe odpovídá izolátům dříve určovaným jako T. mentagrophytes var. granulosum [76]. Pro toto jméno ale neexistuje typový materiál a pojetí různých autorů se výrazně lišilo. Jedná se o jedno z nejvíce kontrovezních jmen vůbec a některými autory byl T. mentagrophytes var. granulosum označován jako nepohlavní stadium od A. benhamiae [3], v pojetí jiných autorů, včetně např. Kuklové et al. [67], se evidentně jednalo o zoofilní T. interdigitale. Odlišná je i klinická manifestace a většinou platí, že antropofilní kmeny T. interdigitale působí u člověka infekce manifestující se jako t. pedis a t. unguium, které mají tendenci k chronicitě. Naproti tomu zoofilní kmeny T. interdigitale působí silně zánětlivé infekce typu t. corporis (obr. 3) a jsou přenesené nejčastěji z hlodavců, psů a koček [40, 76]. Rozdílné je i rozložení těchto infekcí v populaci v závislosti na věku a pohlaví pacientů (obr. 4). Druh T. interdigitale má tedy v dnešním pojetí široké hostitelské spektrum zahrnující savce včetně člověka (tab. 2), a vyznačuje se dvěma různými morfologickými typy, jejichž rozeznávání může mít i praktický klinický význam. Vzhledem ke klonální povaze lidských izolátů je možné jejich odlišení od původem zvířecích izolátů např. pomocí sekvence ITS rDNA, které může být přínosné u kmenů s přechodnou morfologií [40]. Odlišení dalšími metodami jako PCR-fingerprinting, nebo MALDI-TOF hmotnostní spektrometrií je obtížné, nebo nemožné kvůli blízké příbuznosti obou forem [44, 77].
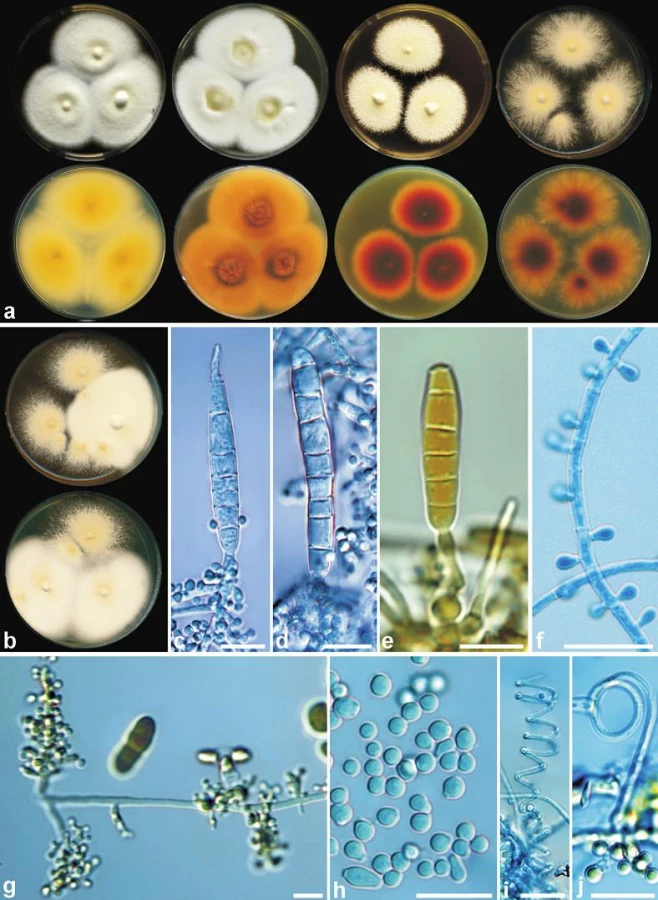


Dalšími klonálními liniemi, které se v minulosti pravděpodobně odštěpily od A. vanbreuseghemii, jsou druhy T. tonsurans a T. equinum. Oba druhy jsou velmi blízce příbuzné a z tohoto důvodu byly brzy po nástupu molekulárních metod poněkud ukvapeně synonymizovány [16, 27]. Synonymizace vzbudila poměrně bouřlivé diskuse [103, 115] a někteří autoři oba druhy obhajovali na základě jejich jedinečné ekologie a fyziologie. T. tonsurans je antropofilní druh, původce t. capitis (endothrix) a t. corporis, a je častý hlavně v méně rozvinutých oblastech světa, výjimkou je Severní Amerika a některé evropské země s velkým podílem přistěhovalců [37, 89]. Pojem t. gladiatorum byl vytvořen pro infekce t. corporis u zápasníků a dalších sportovců, které se často vyskytují v lokálních epidemiích, s nimiž se dá setkat i na našem území a v okolních státech [21, 44]. Hlavním hostitelem T. equinum je naproti tomu kůň a infekce přenesené na člověka jsou vzácné [17]. T. equinum na rozdíl od T. tonsurans vyžaduje niacin v médiu [23] a všechny kmeny T. equinum mají pouze párovací typ MT+, kdežto T. tonsurans kmeny jsou typu MT - [96, 103]. Pozdější molekulárně genetické studie potvrdily, že se druhy dají od sebe odlišit i několika záměnami v sekvencích běžně používaných genů, jako např. ITS rDNA a ß-tubulinu [84, 103], a jsou rozeznávány i MALDI-TOF hmotnostní spektrometrií [77]. Dnes panuje všeobecná shoda, že se jedná o dva odlišné druhy [17, 34].
Komplex A. benhamiae zahrnuje přinejmenším pět zoofilních druhů (viz obr. 1, tab. 2), výjimkou je pouze antropofilní T. concentricum, původce t. imbricata s hlavními endemickými oblastmi na pacifických ostrovech, v jihovýchodní Asii a Střední a Jižní Americe [9]. Nejvýznamnějším zástupcem komplexu je druh A. benhamiae, jehož nepohlavní stadium nemá v současnosti jméno (Trichophyton sp.), protože byl tento druh v minulosti spojován s domnělým nepohlavním stadiem T. mentagrophytes [3]. Pozdější fylogenetické analýzy [26, 27] i křížící experimenty ale ukázaly [106, 105], že se jedná o odlišné druhy. Rezervoárem A. benhamiae jsou hlavně hlodavci (morče, králík, činčila, křeček, apod.), méně často pes nebo kočka [66, 93, 106, 107]. V nedávné minulosti se druh rychle rozšířil v Evropě a Japonsku [11, 22, 53, 107] a v současnosti se jedná o nejvýznamnější zoofilní druh přenášený na člověka v ČR [44, 71, 95]. Stejně jako A. benhamiae, také další zoofilní druhy tohoto komplexu působí především silně zánětlivou formu t. corporis a capitis. Rezervoárem T. erinacei (dříve T. mentagrophytes var. erinacei) jsou hlavně ježci a tento druh je ve zvýšené míře zaznamenáván z Německa [76, 92] a jsou známy i případy z ČR [44]. Hostitelem druhu T. verrucosum je skot, ale nikoliv ovce, jak se dříve uvádělo. Jedna z variet druhu (T. verrucosum var. autotrophicum) uváděná z ovcí [18, 88] byla totiž synonymizována s T. interdigitale [27]. Hostitelem vzácného druhu T. bullosum jsou koně, ale přenos na člověka je vzácný [94]. Druh morfologicky připomíná T. verrucosum a byl zachycen i z léze u koně v ČR (Lysková et al., nepublikovaná data). Přirozený hostitel druhu T. eriotrephon není známý a zoofilní původ se jen předpokládá na základě klinické manifestace infekcí a fylogenetické pozice druhu (popsány byly jen 3 případy u člověka v Nizozemí a Íránu) [40, 80, 86].
Trichophyton rubrum komplex nezahrnuje žádné známé pohlavní stadium. Na základě molekulárně genetických dat byly všechny druhy a variety tohoto komplexu zahrnuty pod T. rubrum a T. violaceum (viz tab. 1) [33, 30, 35, 47-50, 55, 69, 78]. T. rubrum je tak dnes ve většině rozvinutých zemí, včetně ČR [44, 67], jednoznačně nejvýznamnějším původcem dermatofytóz vůbec [89]. Některé v minulosti popsané druhy komplexu vykazovaly unikátní morfologické (odlišnou pigmentaci kolonií, hyfy větvící se v ostrých úhlech proti směru růstu, tzv. „reflexive hyphae“; aj.), nebo fyziologické znaky (požadavky na vitaminy v médiu, pozitivní ureázový test aj.), nebo se vyznačovaly určitou převažující klinickou manifestací, či endemickým rozšířením [34]. Všechny tyto druhy jsou od sebe v podstatě neodlišitelné běžnými molekulárně genetickými metodami a pouze analýza mikrosatelitů umožňuje některé z nich rozpoznat na úrovni populace [33, 78]. To se daří např. u africké populace T. rubrum působící hlavně t. capitis a odpovídající částečně tradičnímu konceptu T. soudanense. Další populace, působící hlavně t. pedis a unguium v Evropě a Americe, odpovídá klasickému konceptu T. rubrum. Asijská populace, která působí hlavně t. corporis a t. cruris, částečně odpovídá někdejšímu konceptu T. raubitschekii [34]. Je doporučeno nadále používat pouze druhová jména T. rubrum a T. violaceum, ostatní epiteta jako např. „raubitschekii“ a „soudanense“ lze užívat jako neformální variety pro zpřesnění komunikace [34].
Komplex A. otae zahrnuje celkem tři blízce příbuzné druhy, M. canis, M. audouinii a M. ferrugineum [25, 29, 57]. Zoofilní a kosmopolitně rozšířený druh M. canis je jediným, u kterého je známé pohlavní stadium (A. otae) a jeho hostitelské spektrum (viz tab. 2) zahrnuje kočky, psy, koně, méně často i další savce [13]. Antropofilní druhy M. audouinii a M. ferrugineum jsou klonální druhy, které mají specifické endemické oblasti v Africe, respektive ve východní Asii a východní Evropě [114]. Všechny druhy komplexu jsou od sebe odlišitelné několika záměnami v sekvencích běžně používaných genů, jako jsou ITS rDNA a ß-tubulin [85], nebo fingerprintovými metodami [25, 69].
Rod Epidermophyton obsahuje podle dnešního konceptu pouze jediný druh, a sice antropofilní E. floccosum. Ostatní druhy řazené do rodu byly už v minulosti synonymizovány s E. floccosum a jinými druhy dermatofytů. Druh E. stockdaleae byl synonymizován s T. ajelloi až na základě molekulárně genetických dat [26].
Geofilní druhy se řadí podle nepohlavního stadia do rodů Trichophyton, Microsporum a Chrysosporium (pozn. některé druhy Chrysosporium bez většího klinického významu byly zařazeny mezi dermatofyty podle fylogenetických studií). U většiny z nich je známé i pohlavní stadium, které se podle posledních taxonomických změn řadí jednotně do rodu Arthroderma [113, 114]. Dnes je uznáváno zhruba 35 geofilních druhů [34], z nichž asi polovina je známá z lidského klinického materiálu (viz tab. 1). Protože geofilní druhy rodu Trichophyton a Microsporum reprezentují linie vzdálené od primárně patogenních zástupců těchto rodů (viz obr. 1), budou muset být převedeny do jiných rodů (viz dále). Na rozdíl od primárně patogenních dermatofytů nebyla diverzita geofilních dermatofytů ještě zcela vyčerpána a za posledních 10 let byly z klinického materiálu popsány druhy T. eboreum, M. mirabile, T. onychocola a M. aenigmaticum [10, 42, 43, 46]. Všechny druhy byly podpořeny polyfázickým přístupem a zachyceny u pacientů vyšetřovaných v Evropě, poslední 2 zmiňované druhy dokonce u českých pacientů [42, 43].
Také ekologie některých druhů musela být přehodnocena na základě nových poznatků. Například druh M. persicolor, dříve klasifikovaný jako zoofilní, je dnes po lepším poznání ekologie považovaný za geofilní. M. persicolor působí infekce u zvířat (často psi) a člověka [44, 45, 75] a spolu s M. gypseum a M. fulvum patří v některých oblastech k nejběžnějším dermatofytům v půdě [91]. Dále byl synonymizován geofilní druh M. vanbreuseghemii s M. gallinae považovaným za zoofilní (viz tab. 1), a proto je dnes druh M. gallinae také většinou řazen mezi geofilní dermatofyty [31, 34, 59].
BUDOUCÍ TAXONOMIE DERMATOFYTŮ: JEDNO JMÉNO PRO JEDEN DRUH
Budoucí změny ve jménech dermatofytů budou souviset nejen s výsledky fylogenetických studií, ale i novými pravidly zavedenými Kódem [74]. Jedná se zejména o ukončení duálního názvosloví u hub. Původní iniciativa One fungus, one name (jedno jméno pro jednu houbu) [38, 110], která zrušení duální nomenklatury prosazovala, nakonec dosáhla zavedení tohoto pravidla do Kódu (prakticky šlo hlavně o zrušení článku 59, který ošetřoval pojmenovávání pleomorfních hub). Taxonomie hub se nyní nachází v přechodném období, kdy je třeba přehodnotit jména všech pleomorfních hub podle těchto pravidel. Rodová jména musí být vybrána podle principů priority (datum publikace) bez ohledu na to, jestli se jedná o jméno pro anamorfu, nebo teleomorfu. Jméno teleomorfy nemá automaticky prioritu, jak navrhovaly některé předchozí systémy. Existují ale případy rodů, kde má méně často používané rodové jméno přednost na základě priority a zavedení změn striktně podle pravidel by znamenalo rozsáhlé a z hlediska uživatelů nepříjemné změny. K dispozici jsou ale nástroje, jak dobře zavedená jména udržet, kterými jsou např. konzervace, nebo zamítnutí některých jmen [39, 74]. Podle nových pravidel také není možné od ledna 2013 navrhovat odlišná jména pro anamorfu a teleomorfu při popisu nových druhů, jak bývalo zvykem u pleomorfních hub.
Duální systém nikdy nepřinášel lékařské mykologii jasný profit, na rozdíl např. od fytopatologie. Kliničtí mykologové se v kulturách téměř vždy setkávají s nepohlavním stadiem houby. To platí nejen pro dermatofyty, ale i další primární, či i oportunní patogeny člověka a zvířat [17]. S pohlavním stadiem se lze u dermatofytů až na výjimky setkat jen na přírodním materiálu, nebo je možné jeho tvorbu indukovat cílenými párovacími experimenty in vitro. Duální systém s sebou nesl i některé nejednoznačnosti. Například nebylo jasné, zda se mají vytvářet odlišná jména pro nepohlavní stadia druhů, která od sebe nejdou jasně morfologicky odlišit, i když pohlavní stadium je jednoznačně odlišné. U několika druhů tak vznikla matoucí situace, kdy např. M. gypseum bylo zároveň jménem anamorfy dvou biologických druhů, Arthroderma gypseum a A. incurvatum. Situace byla vyřešena až v nedávné době, kdy bylo vytvořeno nové jméno M. incurvatum [104]. Dnes tedy existují jasné dvojice anamorfa-teleomorfa: M. incurvatum-A. incurvatum a M. gypseum-A. gypseum. Podobná situace stále přetrvává u T. terrestre, který je zároveň anamorfou tří biologických druhů, a sice A. quadrifidum, A. lenticulare a A. insingulare.
Nový systém jmen u dermatofytů bude muset respektovat především výsledky fylogenetických studií a nová pravidla Kódu. Výsledkem by měl být systém, kde jednotlivé rody budou mít monofyletický status a každému druhu bude přiřazeno pouze jediné správné jméno. O připravovaných změnách odborná veřejnost již několik let diskutuje a tyto změny budou vycházet především z multigenové fylogeneze skupiny dermatofytů, která bude představena v blízké budoucnosti. Shrnutí budoucích změn bylo představeno v přednášce „Same fungus, new names?“ na kongresu 6th Trends in Medical Mycology (2013, Kodaň, Dánsko; prof. Yvonne Gräser). Fylogenetický strom sestrojený na základě několika genů má velmi podobnou topologii jako strom publikovaný dříve na základě sekvencí ITS rDNA [34], a převážně z těchto dat byl sestrojen i strom na obrázku 1. Výraznější změny se v budoucnu vyhnou antropo - a zoofilním dermatofytům, které tvoří monofyletické skupiny (viz obr. 1) a obsahují typové druhy rodů Trichophyton (T. tonsurans) a Microsporum (M. audouinii). Pro druh A. benhamiae bude třeba vytvořit novou kombinaci T. benhamiae, která zatím oficiálně neexistuje, i když je zřejmé, že druh patří do rodu Trichophyton. Dále se diskutuje o poměrně kontroverzním zavedení samostatného jména pro zoofilní kmeny T. interdigitale, kde je k dispozici např. jméno T. granulosum, pro které by ale musel být vhodně zvolen nový typový materiál. Jméno T. interdigitale by se pak týkalo již jen antropofilních kmenů, stejně jako v minulosti. Změna by mohla být výhodná pro klinickou praxi, kde by umožnila používat rozdílná jména pro dvě entity s různou ekologií a působící infekce s odlišnou manifestací (viz obr. 3–4). Nevýhodou takového postupu by byla nutnost sekvenace některých přechodných morfologických forem pro správnou identifikaci, protože ani řada molekulárních metod, včetně MALDI-TOF spektrometrie, antropofilní a zoofilní kmeny kvůli genetické blízkosti nerozliší [44, 77].
Na rozdíl od primárně patogenních dermatofytů se dají rozsáhlé změny jmen očekávat u geofilních dermatofytů. Celosvětově však působí geofilní druhy nejčastěji jen kolem 1 % všech dermatofytóz [89], proto tyto změny zásadně práci kliniků neovlivní. Rodová jména se budou měnit pro geofilní druhy rodů Trichophyton a Microsporum. Pro geofilní zástupce Trichophyton je k dispozici rodové jméno Keratinomyces, jehož typový druh je K. ajelloi (v současnosti T. ajelloi) [111]. Složitější situace je u geofilních druhů Microsporum, které tvoří dvě fylogeneticky odlehlé skupiny. První skupina obsahuje klinicky významné druhy jako M. gypseum a M. persicolor, pro které se nabízí v minulosti používané jméno Nannizzia (typový druh N. incurvata) [97]. Druhá skupina obsahuje jen málo klinicky významných zástupců, a pravděpodobně pro ni bude použito již dříve navržené jméno Lophophyton s typovým druhem L. gallinae (dnes Microsporum gallinae) [72].
Budoucí systém dermatofytů tak bude pravděpodobně obsahovat šest rodů. Klasické rody Trichophyton, Epidermophyton a Microsporum budou zachovány pro primárně patogenní druhy, zbývající tři rody budou obsahovat geofilní druhy. Tyto změny jsou shrnuty v pravém sloupci tabulky 1 a na obrázku 1 (rodová jména v uvozovkách).
Poděkování: Projekt byl podpořen projektem GAUK1344214.
Do redakce došlo dne 14. 7. 2014.
Adresa pro korespondenci:
Mgr. Vít Hubka
Katedra botaniky, Přírodovědecká fakulta
Univerzita Karlova v Praze
Benátská 2
128 01 Praha 2
e-mail: hubka@biomed.cas.cz
Zdroje
1. ABDEL-RAHMAN, S. M., SUGITA, T., GONZÁLEZ, G. M., et al. Divergence among an international population of Trichophyton tonsurans isolates. Mycopathologia, 2010, 169, p. 1–13.
2. AJELLO, L., GEORG, L. K. In vitro hair cultures for differentiating between atypical isolates of Trichophyton mentagrophytes and Trichophyton rubrum. Mycopathologia, 1957, 8, p. 3–17.
3. AJELLO, L., CHENG, S. L. The perfect state of Trichophyton mentagrophytes. Sabouraudia, 1967, 5, p. 230–234.
4. ANZAWA, K., KAWASAKI, M., MOCHIZUKI, T., ISHIZAKI, H. Successful mating of Trichophyton rubrum with Arthroderma simii. Med. Mycol., 2010, 48, p. 629–634.
5. AVISE, J. C., BALL, R. M. Principles of genealogical concordance in species concepts and biological taxonomy. In Futuyma, D., Antonovics, J, editors. Oxford Surveys in Evolutionary Biology. 7. Oxford: Oxford University Press; 1990, p. 45–67.
6. BEGUIN, H., PYCK, N., HENDRICKX, M., et al. The taxonomic status of Trichophyton quinckeanum and T. interdigitale revisited: a multigene phylogenetic approach. Med. Mycol., 2012, 50, p. 871–882.
7. BEGUIN, H., GOENS, K., HENDRICKX, M., et al. Is Trichophyton simii endemic to the Indian subcontinent? Med. Mycol., 2013, 51, p. 444–448.
8. BÖHME, H. Arthroderma gertleri sp. nov., die perfekte form von Trichophyton vanbreuseghemii Rioux, Jarry et Juminer. Mycoses, 1967, 10, p. 247–252.
9. BONIFAZ, A., ARCHER-DUBON, C., SAÚL, A. Tinea imbricata or Tokelau. Int. J. Dermatol., 2004, 43, p. 506–510.
10. BRASCH, J., GRÄSER, Y. Trichophyton eboreum sp. nov. isolated from human skin. J. Clin. Microbiol., 2005, 43, p. 5230–5237.
11. BRAUN, S., JAHN, K., WESTERMANN, A., BRUCH-GERHARZ, D., REIFENBERGER, P. D. J. Tinea barbae profunda durch Arthroderma benhamiae. Hautarzt, 2013, 64, p. 720–722.
12. BURKE, J. M., ARNOLD, M. L. Genetics and the fitness of hybrids. Annu. Rev. Genet., 2001, 35, p. 31–52.
13. CABAĄES, F. J. Dermatophytes in domestic animals. Rev. Iberoam. Micol., 2000, 17, p. 104–108.
14. CAMPBELL, C. K., BORMAN, A. M., LINTON, C. J., BRIDGE, P. D., JOHNSON, E. M. Arthroderma olidum, sp. nov. A new addition to the Trichophyton terrestre complex. Med. Mycol., 2006, 44, p. 451–459.
15. DAWSON, C. O., GENTLES, J. The perfect states of Keratinomyces ajelloi van-Breuseghem, Trichophyton terrestre Durie & Frey and Microsporum nanum Fuentes. Sabouraudia, 1962, 1, p. 49–57.
16. DE HOOG, G. S., GUARRO, J., GENE, J., FIGUERAS, M. J. Atlas of Clinical Fungi. 2nd edn. Utrecht: Centraalbureau voor Schimmelcultures, 2000, 1126 p.
17. DE HOOG, G. S., GUARRO, J., GENÉ, J., FIGUERAS, M. J. Atlas of Clinical Fungi. 3rd. CD-ROM ed. Utrecht: CBS-KNAW Fungal Biodiversity Centre, 2009, p.
18. DENG, S., BULMER, G., SUMMERBELL, R., et al. Changes in frequency of agents of tinea capitis in school children from Western China suggest slow migration rates in dermatophytes. Med. Mycol., 2008, 46, p. 421–427.
19. EMMONS, C. W. Dermatophytes: natural grouping based on the form of the spores and accessory organs. Arch. Derm. Syphilol., 1934, 30, p. 337–362.
20. EMMONS, C. W., BINFORD, C. H., UTZ, J. P. Medical mycology. Philadelphia: Lea & Febiger, 1970, 508 p.
21. FARI, E., GRÄSER, Y. An epidemic of tinea corporis caused by Trichophyton tonsurans among children (wrestlers) in Germany. Mycoses, 2000, 43, p. 191–196.
22. FUMEAUX, J., MOCK, M., NINET, B., et al. First report of Arthroderma benhamiae in Switzerland. Dermatology, 2004, 208, p. 244–250.
23. GEORG, L. K., CAMP, L. B. Routine nutritional tests for the identification of dermatophytes. J. Bacteriol., 1957, 74, p. 113–121.
24. GEORG, L. K. Epidemiology of the dermatophytoses sources of infection, modes of transmission and epidemicity. Ann. N Y Acad. Sci., 1960, 89, p. 69–77.
25. GRÄSER, Y., EL FARI, M., STERRY, W., PRESBER, W., TIETZ, H. Identification of common dermatophytes (Trichophyton, Microsporum, Epidermophyton) using polymerase chain reactions. Brit. J. Dermatol., 1998, 138, p. 576–582.
26. GRÄSER, Y., EL FARI, M., VILGALYS, R., et al. Phylogeny and taxonomy of the family Arthrodermataceae (dermatophytes) using sequence analysis of the ribosomal ITS region. Med. Mycol., 1999, 37, p. 105–114.
27. GRÄSER, Y., KUIJPERS, A. F. A., PRESBER, W., DE HOOG, G. S. Molecular taxonomy of Trichophyton mentagrophytes and T. tonsurans. Med. Mycol., 1999, 37, p. 315–330.
28. GRÄSER, Y., DE HOOG, G. S., KUIJPERS, A. F. A. Recent advances in the molecular taxonomy of dermatophytes. Rev. Iberoam. Micol., 2000, 17, p. 17–21.
29. GRÄSER, Y., KUIJPERS, A. F. A., EL FARI, M., PRESBER, W., DE HOOG, G. S. Molecular and conventional taxonomy of the Microsporum canis complex. Med. Mycol., 2000, 38, p. 143–153.
30. GRÄSER, Y., KUIJPERS, A. F. A., PRESBER, W., DE HOOG, G. S. Molecular taxonomy of the Trichophyton rubrum complex. J. Clin. Microbiol., 2000, 38, p. 3329–3336.
31. GRÄSER, Y. Molekulare Systematik und Evolution der Spezies der Familie Arthrodermataceae (Dermatophyten). Berlin: Humboldt-Universität; 2002.
32. GRÄSER, Y., DE HOOG, S., SUMMERBELL, R. Dermatophytes: recognizing species of clonal fungi. Med. Mycol., 2006, 44, p. 199–209.
33. GRÄSER, Y., FRÖHLICH, J., PRESBER, W., DE HOOG, S. Microsatellite markers reveal geographic population differentiation in Trichophyton rubrum. J. Med. Microbiol., 2007, 56, p. 1058–1065.
34. GRÄSER, Y., SCOTT, J., SUMMERBELL, R. The new species concept in dermatophytes – a polyphasic approach. Mycopathologia, 2008, 166, p. 239–256.
35. GUPTA, A., KOHLI, Y., SUMMERBELL, R. Exploratory study of single-copy genes and ribosomal intergenic spacers for rapid distinction of dermatophytes. Stud. Mycol., 2002, p. 87–96.
36. HARMSEN, D., SCHWINN, A., WEIG, M., BRÖCKER, E. B., HEESEMANN, J. Phylogeny and dating of some pathogenic keratinophilic fungi using small subunit ribosomal RNA. J. Med. Vet. Mycol., 1995, 33, p. 299–303.
37. HAVLICKOVA, B., CZAIKA, V., FRIEDRICH, M. Epidemiological trends in skin mycoses worldwide. Mycoses, 2008, 51 (Suppl. 4), p. 2–15.
38. HAWKSWORTH, D. L., CROUS, P. W., REDHEAD, S. A., et al. The Amsterdam declaration on fungal nomenclature. IMA Fungus, 2011, 2, p. 105–112.
39. HAWKSWORTH, D. L. Managing and coping with names of pleomorphic fungi in a period of transition. IMA Fungus, 2012, 3, p. 15–24.
40. HEIDEMANN, S., MONOD, M., GRÄSER, Y. Signature polymorphisms in the internal transcribed spacer region relevant for the differentiation of zoophilic and anthropophilic strains of Trichophyton interdigitale and other species of T. mentagrophytes sensu lato. Brit. J. Dermatol., 2010, 162, p. 282–295.
41. HUBÁLEK, Z. Keratinophilic fungi associated with free-living mammals and birds. In: Kushwaha, R.K.S., GUARRO, J, editors. Biology of Dermatophytes. Bilbao: Revista Iberoamericana de Micología; 2000, p. 93–103.
42. HUBKA, V., CMOKOVA, A., SKOREPOVA, M., MIKULA, P., KOLARIK, M. Trichophyton onychocola sp. nov. isolated from human nail. Med. Mycol., 2014, 52, p. 285–292.
43. HUBKA, V., DOBIASOVA, S., DOBIAS, R., KOLARIK, M. Microsporum aenigmaticum sp. nov. from M. gypseum complex, isolated as a cause of tinea corporis. Med. Mycol., 2014, 52, p. 387–396.
44. HUBKA, V., VĚTROVSKÝ, T., DOBIÁŠOVÁ, S., et al. Molekulární epidemiologie dermatofytóz v České republice: výsledky dvouleté studie. Čes.-slov. Derm., 2014, 89, 4, p. 167–174.
45. CHEN, W., SEIDL, H. P., RING, J., SCHNOPP, C. Two pediatric cases of Microsporum persicolor infection. Int. J. Dermatol., 2012, 51, p. 204–206.
46. CHOI, J. S., GRÄSER, Y., WALTHER, G., et al. Microsporum mirabile and its teleomorph Arthroderma mirabile, a new dermatophyte species in the M. cookei clade. Med. Mycol., 2012, 50, p. 161–169.
47. JACKSON, C. J., BARTON, R. C., EVANS, E. G. V. Species identification and strain differentiation of dermatophyte fungi by analysis of ribosomal-DNA intergenic spacer regions. J. Clin. Microbiol., 1999, 37, p. 931–936.
48. JACKSON, C. J., BARTON, R. C., KELLY, S. L., EVANS, E. G. V. Strain identification of Trichophyton rubrum by specific amplification of subrepeat elements in the ribosomal DNA nontranscribed spacer. J. Clin. Microbiol., 2000, 38, p. 4527–4534.
49. KANBE, T., SUZUKI, Y., KAMIYA, A., et al. PCR-based identification of common dermatophyte species using primer sets specific for the DNA topoisomerase II genes. J. Dermatol. Sci., 2003, 32, p. 151–161.
50. KANBE, T., SUZUKI, Y., KAMIYA, A., et al. Species - -identification of dermatophytes Trichophyton, Microsporum and Epidermophyton by PCR and PCR-RFLP targeting of the DNA topoisomerase II genes. J. Dermatol. Sci., 2003, 33, p. 41–54.
51. KANE, J., SIGLER, L., SUMMERBELL, R. C. Improved procedures for differentiating Microsporum persicolor from Trichophyton mentagrophytes. J. Clin. Microbiol., 1987, 25, p. 2449–2452.
52. KANE, J., SUMMERBELL, R., SIGLER, L., KRAJDEN, S., LAND, G. Laboratory handbook of dermatophytes: a clinical guide and laboratory handbook of dermatophytes and other filamentous fungi from skin, hair, and nails. Belmont, California: Star Publishing, 1997, 344 p.
53. KANO, R., NAKAMURA, Y., YASUDA, K., et al. The first isolation of Arthroderma benhamiae in Japan. Microbiol. Immunol., 1998, 42, p. 575–578.
54. KANO, R., NAKAMURA, Y., WATANABE, S., TSUJIMOTO, H., HASEGAWA, A. Phylogenetic relation of Epidermophyton floccosum to the species of Microsporum and Trichophyton in chitin synthase 1 (CHS1) gene sequences. Mycopathologia, 1999, 146, p. 111–113.
55. KANO, R., OKABAYASHI, K., NAKAMURA, Y., et al. Differences among chitin synthase 1 gene sequences in Trichophyton rubrum and T. violaceum. Med. Mycol., 2000, 38, p. 47–50.
56. KANO, R., YOSHIDA, E., YAGUCHI, T., et al. Mating type gene (MAT1-2) of Trichophyton verrucosum. Mycopathologia, 2014, 177, p. 103–112.
57. KASZUBIAK, A., KLEIN, S., DE HOOG, G. S., GRÄSER, Y. Population structure and evolutionary origins of Microsporum canis, M. ferrugineum and M. audouinii. Infect. Genet. Evol., 2004, 4, p. 179–186.
58. KATOH, K., STANDLEY, D. M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol., 2013, 30, p. 772–780.
59. KAWASAKI, M., AOKI, M., ISHIZAKI, H. Phylogenetic relationships of some Microsporum and Arthroderma species inferred from mitochondrial DNA analysis. Mycopathologia, 1995, 130, p. 11–21.
60. KAWASAKI, M., ANZAWA, K., TAKEDA, K., et al. Genetic and phenotypic variations among F1 progenies of Arthroderma benhamiae. Jap. J. Med. Mycol., 2008, 49, p. 103–110.
61. KAWASAKI, M., ANZAWA, K., WAKASA, A., et al. Different genes can result in different phylogenetic relationships in Trichophyton species. Jap. J. Med. Mycol., 2008, 49, p. 311–318.
62. KAWASAKI, M., ANZAWA, K., MOCHIZUKI, T., ISHIZAKI, H., M. HEMASHETTAR, B. Successful mating of a human isolate of Arthroderma simii with a tester strain of A. vanbreuseghemii. Med. Mycol. J., 2009, 50, p. 15–18.
63. KAWASAKI, M., ANZAWA, K., WAKASA, A., et al. Matings among three teleomorphs of Trichophyton mentagrophytes. Jap. J. Med. Mycol., 2010, 51, p. 143–152.
64. KAWASAKI, M. Verification of a taxonomy of dermatophytes based on mating results and phylogenetic analyses. Med. Mycol. J., 2011, 52, p. 291–295.
65. KIELSTEIN, P., WOLF, H., GRÄSER, Y., BUZINA, W., BLANZ, P. On the variability of Trichophyton verrucosum isolates from vaccinated herds with ringworm of cattle. Mycoses, 1998, 41 (Suppl. 2), p. 58–64.
66. KRAEMER, A., HEIN, J., HEUSINGER, A., MUELLER, R. Clinical signs, therapy and zoonotic risk of pet guinea pigs with dermatophytosis. Mycoses, 2013, 56, p. 168–172.
67. KUKLOVÁ, I., KUČEROVÁ, H. Dermatophytoses in Prague, Czech Republic, between 1987 and 1998. Mycoses, 2001, 44, p. 493–496.
68. LECLERC, M., PHILIPPE, H., GUEHO, E. Phylogeny of dermatophytes and dimorphic fungi based on large subunit ribosomal RNA sequence comparisons. J. Med. Vet. Mycol., 1994, 32, p. 331–341.
69. LIU, D., COLOE, S., BAIRD, R., PEDERSEN, J. Application of PCR to the identification of dermatophyte fungi. J. Med. Microbiol., 2000, 49, p. 493–497.
70. LYSKOVÁ, P., HUBKA, V., BODNÁROVÁ, J. Případ tinea corporis vyvolaný Microsporum incurvatum, geofilním druhem příbuzným M. gypseum. Čes.-slov. Derm., 2014, 89, 4, p. 187–191.
71. MALLÁTOVÁ, N., JANATOVÁ, H., KOCOURKOVÁ, K., et al. Arthroderma benhamiae jako původce tinea capitis profunda a tinea corporis u dětských pacientů. Čes.-slov. Derm., 2014, 89, 4, p. 199–204.
72. MATRUCHOT, L., DASSONVILLE, C. Recherches expérimentales sur une dermatomycose des poules et sur son parasite. Rev. Gen. Bot., 1899, 11, p. 429–444.
73. MAYR, E. Systematics and the origin of species, from the viewpoint of a zoologist. Cambridge: Harvard University Press, 1942, 334 p.
74. MCNEILL, J., BARRIE, F. R., BUCK, W. R., et al. International Code of Nomenclature for algae, fungi, and plants (Melbourne Code) adopted by the Eighteenth International Botanical Congress Melbourne, Australia, July 2011. Königstein: Regnum Vegetabile 154, Koeltz Scientific Books, 2012, 240 p.
75. MULLER, A., GUAGUťRE, E., DEGORCE-RUBIALES, F., BOURDOISEAU, G. Dermatophytosis due to Microsporum persicolor: a retrospective study of 16 cases. Can. Vet. J., 2011, 52, p. 385–388.
76. NENOFF, P., HERRMANN, J., GRÄSER, Y. Trichophyton mentagrophytes sive interdigitale? A dermatophyte in the course of time. J. Dtsch. Dermatol. Ges., 2007, 5, p. 198–202.
77. NENOFF, P., ERHARD, M., SIMON, J. C., et al. MALDI-TOF mass spectrometry-a rapid method for the identification of dermatophyte species. Med. Mycol., 2013, 51, p. 17–24.
78. OHST, T., DE HOOG, S., PRESBER, W., STAVRAKIEVA, V., GRÄSER, Y. Origins of microsatellite diversity in the Trichophyton rubrum-T. violaceum clade (dermatophytes). J. Clin. Microbiol., 2004, 42, p. 4444–4448.
79. PADHYE, A., CARMICHAEL, J. Mating reactions in the Trichophyton terrestre complex. Med. Mycol., 1973, 11, p. 64–69.
80. PAPEGAAY, J. Over pathogene huidschimmels in Amsterdam voorkomend bij den mensch. Ned. Tijdschr. Geneeskd., 1925, 69, p. 879–890.
81. PHILPOT, C. The differentiation of Trichophyton mentagrophytes from T. rubrum by a simple urease test. Med. Mycol., 1967, 5, p. 189–193.
82. PROBST, S., DE HOOG, G. S., GRÄSER, Y. Development of DNA markers to explore host shifts in dermatophytes. Stud. Mycol., 2002, p. 57–74.
83. REBELL, G., TAPLIN, D. Dermatophytes. Their recognition and identification. 2nd ed. Miami: University of Miami Press, 1970, 124 p.
84. REZAEI-MATEHKOLAEI, A., MAKIMURA, K., DE HOOG, G. S., et al. Discrimination of Trichophyton tonsurans and Trichophyton equinum PCR-RFLP and by beta-tubulin and translation elongation factor 1-alpha sequencing. Med. Mycol., 2012, 50, p. 760–764.
85. REZAEI-MATEHKOLAEI, A., MAKIMURA, K., DE HOOG, G. S., et al. Multilocus differentiation of the related dermatophytes Microsporum canis, Microsporum ferrugineum and Microsporum audouinii. J. Med. Microbiol., 2012, 61, p. 57–63.
86. REZAEI-MATEHKOLAEI, A., MAKIMURA, K., DE HOOG, S., et al. Molecular epidemiology of dermatophytosis in Tehran, Iran, a clinical and microbial survey. Med. Mycol., 2013, 51, p. 203–207.
87. SABOURAUD, R. J. A. Les teignes. Paris: Masson & Co., 1910, 855 p.
88. SCOTT, D. An outbreak of ringworm in karakul sheep caused by a physiological variant of Trichophyton verrucosum Bodin. Onderstepoort J. Vet. Res., 1975, 42, p. 49–52.
89. SEEBACHER, C., BOUCHARA, J.-P., MIGNON, B. Updates on the epidemiology of dermatophyte infections. Mycopathologia, 2008, 166, p. 335–352.
90. SHARMA, R., RAJAK, R. C., PANDEY, A. K., GRÄSER, Y. Internal Transcribed Spacer (ITS) of rDNA of appendaged and non-appendaged strains of Microsporum gypseum reveals Microsporum appendiculatum as its synonym. Antonie Leeuwenhoek, 2006, 89, p. 197–202.
91. SHARMA, R., PRESBER, W., RAJAK, R. C., GRÄSER, Y. Molecular detection of Microsporum persicolor in soil suggesting widespread dispersal in central India. Med. Mycol., 2008, 46, p. 67–73.
92. SCHAUDER, S., KIRSCH-NIETZKI, M., WEGENER, S., SWITZER, E., QADRIPUR, S. Von Igeln auf Menschen. Zoophile Dermatomykose durch Trichophyton erinacei bei 8 Patienten. Hautarzt, 2007, 58, p. 62–67.
93. SIEKLUCKI, U., OH, S. H., HOYER, L. L. Frequent isolation of Arthroderma benhamiae from dogs with dermatophytosis. Vet. Dermatol., 2014, 25, p. 39–41.
94. SITTERLE, E., FREALLE, E., FOULET, F., et al. Trichophyton bullosum: a new zoonotic dermatophyte species. Med. Mycol., 2012, 50, p. 305–309.
95. SKOŘEPOVÁ, M., HUBKA, V., POLÁŠKOVÁ, S., STARÁ, J., ČMOKOVÁ, A. Naše první zkušenosti s infekcemi vyvolanými Arthroderma benhamiae (Trichophyton sp.). Čes.-slov. Derm., 2014, 89, 4, p. 192–198.
96. STOCKDALE, P. Sexual stimulation between Arthroderma simii and other dermatophytes. In: Vanbreuseghem, R. R., De Droey, C., editors. Sexuality and pathogenicity of fungi. New York: Masson; 1981, p. 57–68.
97. STOCKDALE, P. M. Nannizzia incurvata gen. nov., sp. nov., a perfect state of Microsporum gypseum (Bodin) Guiart et Grigorakis. Sabouraudia, 1962, 1, p. 41–48.
98. STOCKDALE, P. M. The Microsporum gypseum complex (Nannizzia incurvata Stockd., N. gypsea (Nann.) comb. nov., N. fulva sp. nov.). Sabouraudia, 1964, 3, p. 114–126.
99. STOCKDALE, P. M. Nannizzia persicolor sp. nov., the perfect state of Trichophyton persicolor Sabouraud. Med. Mycol., 1967, 5, p. 355–359.
100. SUMMERBELL, R. C., ROSENTHAL, S. A., KANE, J. Rapid method for differentiation of Trichophyton rubrum, Trichophyton mentagrophytes, and related dermatophyte species. J. Clin. Microbiol., 1988, 26, p. 2279–2282.
101. SUMMERBELL, R. C. Form and function in the evolution of dermatophytes. Rev. Iberoam. Micol., 2000, 17, p. 30–43.
102. SUMMERBELL, R. C., WEITZMAN, I., PADHYE, A. A. The Trichophyton mentagrophytes complex: biological species and mating type prevalences of North American isolates, and a review of the worldwide distribution and host associations of species and mating types. Stud. Mycol., 2002, 47, p. 75–86.
103. SUMMERBELL, R. C., MOORE, M. K., STARINK-WILLEMSE, M., VAN IPEREN, A. ITS barcodes for Trichophyton tonsurans and T. equinum. Med. Mycol., 2007, 45, p. 193–200.
104. SUN, P. L., MU, C.-A., FAN, C. C., et al. Cat favus caused by Microsporum incurvatum comb. nov.: The clinical and histopathological features and molecular phylogeny. Med. Mycol., 2014, 52, p. 276–284.
105. SYMOENS, F., JOUSSON, O., PLANARD, C., et al. Molecular analysis and mating behaviour of the Trichophyton mentagrophytes species complex. Int. J. Med. Microbiol., 2011, 301, p. 260–266.
106. SYMOENS, F., JOUSSON, O., PACKEU, A., et al. The dermatophyte species Arthroderma benhamiae: intraspecies variability and mating behaviour. J. Med. Microbiol., 2013, 62, p. 377–385.
107. TAKEDA, K., NISHIBU, A., ANZAWA, K., MOCHIZUKI, T. Molecular epidemiology of a major subgroup of Arthroderma benhamiae isolated in Japan by restriction fragment length polymorphism analysis of the non-transcribed spacer region of ribosomal RNA gene. Jpn. J. Infect. Dis., 2012, 65, p. 233–239.
108. TAMURA, K., STECHER, G., PETERSON, D., FILIPSKI, A., KUMAR, S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol., 2013, 30, p. 2725–2729.
109. TANAKA, S., SUMMERBELL, R. C., TSUBOI, R., et al. Advances in dermatophytes and dermatophytosis. J. Med. Vet. Mycol., 1992, 30, p. 29–39.
110. TAYLOR, J. W. One fungus = one name: DNA and fungal nomenclature twenty years after PCR. IMA fungus, 2011, 2, p. 113–120.
111. VANBREUSEGHEM, R. Interźt theorique et practique d’un nouveau dermatophyte isolée du sol: Keratinomyces ajelloi gen. nov., sp. nov. Bull. Acad. R. Sci. Belg., Cl. Sci., 1952, 38, p. 1068–1077.
112. WEITZMAN, I., PADHYE, A. Is Arthroderma simii the perfect state of Trichophyton quinckeanum? Sabouraudia, 1976, 14, p. 65–74.
113. WEITZMAN, I., MCGINNIS, M., PADHYE, A., AJELLO, L. The genus Arthroderma and its later synonym Nannizzia. Mycotaxon, 1986, 25, p. 505–518.
114. WEITZMAN, I., SUMMERBELL, R. C. The dermatophytes. Clin. Microbiol. Rev., 1995, 8, p. 240–259.
115. WOODGYER, A. The curious adventures of Trichophyton equinum in the realm of molecular biology: a modern fairy tale. Med. Mycol., 2004, 42, p. 397–403.
116. YOUNG, C. N. Pseudo-cleistothecia in Trichophyton rubrum. Sabouraudia, 1968, 6, p. 160–162.
Štítky
Dermatologie Dětská dermatologieČlánek vyšel v časopise
Česko-slovenská dermatologie

2014 Číslo 4
- Nástroje k hodnocení závažnosti psoriázy v klinických studiích
- Systémová léčba atopické dermatitidy konečně i u dětí
- Inovace v hojení ran: krytí Zetuvit Plus Silicone Border pro optimální management exsudátu z ran
Nejčtenější v tomto čísle
- Životní jubileum profesora MUDr. Vladimíra Vašků, CSc.
- Naše první zkušenosti s infekcemi vyvolanými Arthroderma benhamiae (Trichophyton sp.)
- Případ tinea corporis vyvolaný Microsporum incurvatum, geofilním druhem příbuzným M. gypseum
- Arthroderma benhamiae jako původce tinea capitis profunda a tinea corporis u dětských pacientů
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy