
Provides Insights into the Evolution of the Salmonellae
The genus Salmonella contains two species, S. bongori and S. enterica. Compared to the well-studied S. enterica there is a marked lack of information regarding the genetic makeup and diversity of S. bongori. S. bongori has been found predominantly associated with cold-blooded animals, but it can infect humans. To define the phylogeny of this species, and compare it to S. enterica, we have sequenced 28 isolates representing most of the known diversity of S. bongori. This cross-species analysis allowed us to confidently differentiate ancestral functions from those acquired following speciation, which include both metabolic and virulence-associated capacities. We show that, although S. bongori inherited a basic set of Salmonella common virulence functions, it has subsequently elaborated on this in a different direction to S. enterica. It is an established feature of S. enterica evolution that the acquisition of the type III secretion systems (T3SS-1 and T3SS-2) has been followed by the sequential acquisition of genes encoding secreted targets, termed effectors proteins. We show that this is also true of S. bongori, which has acquired an array of novel effector proteins (sboA-L). All but two of these effectors have no significant S. enterica homologues and instead are highly similar to those found in enteropathogenic Escherichia coli (EPEC). Remarkably, SboH is found to be a chimeric effector protein, encoded by a fusion of the T3SS-1 effector gene sopA and a gene highly similar to the EPEC effector nleH from enteropathogenic E. coli. We demonstrate that representatives of these new effectors are translocated and that SboH, similarly to NleH, blocks intrinsic apoptotic pathways while being targeted to the mitochondria by the SopA part of the fusion. This work suggests that S. bongori has inherited the ancestral Salmonella virulence gene set, but has adapted by incorporating virulence determinants that resemble those employed by EPEC.
Published in the journal:
. PLoS Pathog 7(8): e32767. doi:10.1371/journal.ppat.1002191
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1002191
Summary
The genus Salmonella contains two species, S. bongori and S. enterica. Compared to the well-studied S. enterica there is a marked lack of information regarding the genetic makeup and diversity of S. bongori. S. bongori has been found predominantly associated with cold-blooded animals, but it can infect humans. To define the phylogeny of this species, and compare it to S. enterica, we have sequenced 28 isolates representing most of the known diversity of S. bongori. This cross-species analysis allowed us to confidently differentiate ancestral functions from those acquired following speciation, which include both metabolic and virulence-associated capacities. We show that, although S. bongori inherited a basic set of Salmonella common virulence functions, it has subsequently elaborated on this in a different direction to S. enterica. It is an established feature of S. enterica evolution that the acquisition of the type III secretion systems (T3SS-1 and T3SS-2) has been followed by the sequential acquisition of genes encoding secreted targets, termed effectors proteins. We show that this is also true of S. bongori, which has acquired an array of novel effector proteins (sboA-L). All but two of these effectors have no significant S. enterica homologues and instead are highly similar to those found in enteropathogenic Escherichia coli (EPEC). Remarkably, SboH is found to be a chimeric effector protein, encoded by a fusion of the T3SS-1 effector gene sopA and a gene highly similar to the EPEC effector nleH from enteropathogenic E. coli. We demonstrate that representatives of these new effectors are translocated and that SboH, similarly to NleH, blocks intrinsic apoptotic pathways while being targeted to the mitochondria by the SopA part of the fusion. This work suggests that S. bongori has inherited the ancestral Salmonella virulence gene set, but has adapted by incorporating virulence determinants that resemble those employed by EPEC.
Introduction
Salmonella serovars are predominately pathogenic Enterobacteriaceae that are thought to have diverged from a common ancestor with Escherichia coli ∼100 million years ago [1]. The genus Salmonella currently comprises two species; S. bongori and S. enterica, with S. enterica being comprised of 6 subspecies enterica, salamae, arizonae, diarizonae, houtenae and indica [2], [3], [4], [5], [6]. These S. enterica subspecies are further subdivided into >2500 serovars. Although S. bongori have been reported to infect humans [7], [8], the species is predominantly associated with cold-blooded animals whereas serovars causing disease in humans and other warm-blooded animals mostly belong to S. enterica subspecies enterica. Since S. enterica incorporates clinically important pathogens, our knowledge about the genus Salmonella is heavily biased and there is a marked paucity of information relating to the genetic and phylogenetic makeup of S. bongori.
Even though E. coli and Salmonella are estimated to have diverged millions of years ago, their genomes still display significant similarity including extensive regions of synteny. However, in common with other Enterobacteriaceae significant diversity has been driven by horizontal gene transfer on a background of gradual genome sequence drift [9]. Many of the genes which are unique to Salmonella serovars, compared to E. coli, are found on large discrete genomic islands that include prophage elements and specialised loci termed Salmonella pathogenicity islands (SPIs) [10], [11], [12]. These Salmonella-specific functions include many genes required for the full expression of virulence and some of these were acquired by S. enterica following the split from S. bongori. For example, S. enterica encodes two complete type III secretion systems encoded by SPI-1 (T3SS-1) and SPI-2 (T3SS-2) [13], [14], [15], [16], whereas S. bongori lacks SPI-2, which is required for optimal replication within macrophages [15], [17], [18].
Several characteristics of S. bongori suggest that this species may, in evolutionary terms, lie somewhere between E. coli and S. enterica. Consequently, to prove this hypothesis we have studied multiple genotypic and phenotypic characteristics of S. bongori and compared these to S. enterica and other Enterobacteriaceae. In terms of genotype, we have determined a reference quality sequence of S. bongori 12419, originally isolated from an African frog in 1972 (Salmonella Reference Collection C strain SARC11) and prepared draft sequences of a globally and temporally diverse S. bongori collection including 21 representatives of the 23 known serovars (SV). From our data we have been able to determine inter and intra-species phylogeny and have used this to differentiate ancestral and more recently acquired virulence and metabolic functions. These data show that S. bongori possesses only a basic set of ancestral Salmonella virulence functions and lacks several metabolic pathways that define S. enterica. Nevertheless, S. bongori has not remained functionally static; it has acquired a repertoire of 12 T3SS candidate effector proteins, 10 of which are not found in other salmonellae but are significantly similar to known effectors found in enteropathogenic Escherichia coli (EPEC) strains. We herein demonstrate that representatives of these effectors are translocated and that at least one of these effectors, S. bongori outer protein H (SboH), is functionally related to the non-LEE encoded EPEC effector NleH1.
Results
Phylogenetic analysis of the species S. bongori
To place S. bongori in the context of S. enterica we produced a phylogenetic tree using the concatenated MLST gene sequences (as described in [19]) from a selection of S. enterica Sequence types (STs) covering all of the subspecies of S. enterica. The STs for S. enterica were obtained from the S. enterica MLST website (mlst.ucc.ie). The S. bongori MLST gene sequences were extracted from our sequenced strains (described in Table S1), and the EPEC MLST gene sequences were extracted from the genome sequence of strain E2348/69 (Figure 1). Despite the spatial, temporal and phenotypic diversity described within our collection, the S. bongori species forms a surprisingly tight cluster of sequence types (STs) clearly separated from the S. enterica subspecies (Figure 1). The S. bongori isolates in our collection fall into 20 STs, which include 11 novel Salmonella STs (currently S. bongori-specific; Table S1). In comparison, there were 1,419 STs identified as being part of S. enterica present in the MLST database as of the 3rd of May 2011. To investigate the diversity and population structure of S. bongori we finished and fully annotated the genome of S. bongori 12419 (also known as SARC11 [20]). We then used this genome as a reference to produce whole genome sequences for our collection of 27 further S. bongori isolates. Using the whole genome sequences we produced a phylogenetic tree using RAxML (Figure 1), following the removal of mobile genetic elements (MGE; regions excluded from this analysis are listed in Table S2). In order to determine the branch on which the root should be placed we also completed a separate mapping including S. enterica subspecies arizonae strain CDC346-86 (S. arizonae; EMBL CP000880) strain in order to provide an outgroup. When using S. arizonae to locate the root for the S. bongori tree at least three phylogenetic clusters are evident, a feature that is supported by a clustering analysis performed using the program Bayesian Analysis of Population Structure [21], [22]. One of these clusters appears to be basal to the other clusters, based on the position of the root. The clusters are separated by 15,948-22,398 SNPS (Figure S1). The level of SNP variation between the clusters is consistent with the level of SNP variation between two serovars of S. enterica. For example, 39,156 SNPs differentiate S. enterica subspecies enterica serovar Typhimurium (S. Typhimurium) strain SL1344 and S. enterica subspecies enterica serovar Enteritidis (S. Enteritidis) strain P125109 (data not shown). Within S. bongori serotype does not appear to provide a meaningful indication of phylogenetic relationships within the population (Figure 1). This feature of the dataset may imply that there is frequent lateral gene transfer amongst S. bongori strains.

It is also apparent from the genomic data that there is a larger difference in the shift in genome G+C content in S. enterica following the divergence of the salmonellae, compared with S. bongori (Figure S2). Considering S. enterica and S. bongori have been evolving over the same time period these differences are remarkable. Changes in G+C content over time are thought to reflect subtle differences in mutational bias as a consequence of different lifestyles [23]. The combined data in Figure 1 and Figure S2 suggest that there has been a greater increase in G+C content accompanying the specialisation of S. enterica subspecies into warm-blooded hosts.
Genetic flux across the salmonellae
To obtain a comprehensive view of genetic flux over time we used data from the other available Salmonella enterica genome sequences, along with our 28 sequenced S. bongori isolates. To complement this analysis, we used a pan-Salmonella microarray [24], which included S. bongori-specific probes, to look at gene presence/absence across the SARC collection where whole genome sequences are lacking (see methods). First we focussed our analysis on virulence functions that unified or distinguished S. bongori from the other salmonellae (summarised in Figure 2). Functions discussed below are conserved amongst all 28 S. bongori strains we sequenced (unless otherwise stated) and are not isolate-specific.

Salmonella pathogenicity islands
Of the 22 reported SPIs only SPI-1, SPI-4 and SPI-9 are present in S. bongori 12419 with the same gene composition as those defined in S. enterica (Summarised in Figure 2; Table 1). Consistent with previous observations SPI-3 and SPI-5 are incomplete: SPI-3 exists as two independent insertions in S. bongori, SPI-3a and SPI-3b, that appear to have fused into a single element in S. enterica (Figure 2; This study; [25]). SPI-5 has previously been shown to be a chimeric genomic island composed of two regions of markedly differing G+C content in S. enterica, region one carrying the T3SS-1 translocated effector genes sigE, sopB and pipD and region 2 encoding the T3SS-2 translocated effector gene pipB. S. bongori possesses region 1 only; there is no trace of the T3SS-2 effector gene encoded in region 2.

A significant distinguishing feature of S. bongori is the lack of SPI-2 [14], [18], [26], [27]. The site occupied by SPI-2 in S. enterica (alongside tRNA-valV) carries a ∼20 kb genomic island in S. bongori encoding a novel type VI secretion system (SPI-22; see below). The tetrathionate respiration (ttr) gene cluster which lies alongside SPI-2 in S. enterica is retained by S. bongori.
All the T3SS-2 translocated effectors are absent from S. bongori with the exception of SlrP, which in S. enterica subspecies is known to be secreted by both T3SS-1 and T3SS-2 [28]. Conversely 10 of the 12 known T3SS-1 translocated effectors are almost entirely conserved between S. enterica subspecies and S. bongori and include those that stimulate proinflammatory responses, bind actin and are important for cellular invasion (sipA, sipB, sipC, sopB, sopD and sopE2). S. bongori also carries effectors that dampen down cytoskeletal rearrangements and host signalling responses by S. enterica subspecies including avrA (reported to inhibit NF-kappa B [29]) and sptP (pseudogene). Although the S. bongori sopA gene is located at the same site as its orthologues within S. enterica it has been disrupted by an insertion which has generated a chimeric effector protein (denoted SboH; see below).
The S. bongori T3SS-1 translocated effector genes are found at exactly the same genomic loci as they are in S. enterica: carried on SPI-1 itself, SPI-5 or at identical sites in the chromosomal backbone (Table 2). This suggests that most T3SS-1 effectors were sequentially acquired prior to speciation, sopE and sspH1 being the only exceptions. The latter two effector proteins are sporadically distributed in S. enterica subspecies enterica isolates, and carried as cargo on phage [30], [31], consistent with them being more recent acquisitions.

In addition to the lack of SPI-2, S. bongori lacks the entirety of SPI-6 (encoding a type VI secretion system), SPI-13 (required for survival in chicken macrophages), SPI-14 (encoding an electron transport system) and SPI-16 (bacteriophage remnant carrying genes associated with LPS modification) making these islands unique to S. enterica (This study;[10], [32], [33]). From the in silico analysis and microarray data it is evident that SPI-6 and SPI-16 are present in all S. enterica lineages whilst SPI-13 and SPI-14 are only sporadically distributed in S. enterica (Table 1; This study [33], [34], [35]). S. bongori also lacks part of the centisome 54 island (CS54) encoding shdB ratC and ratB which are associated with survival in macrophages and longterm shedding of bacteria from the host [36].
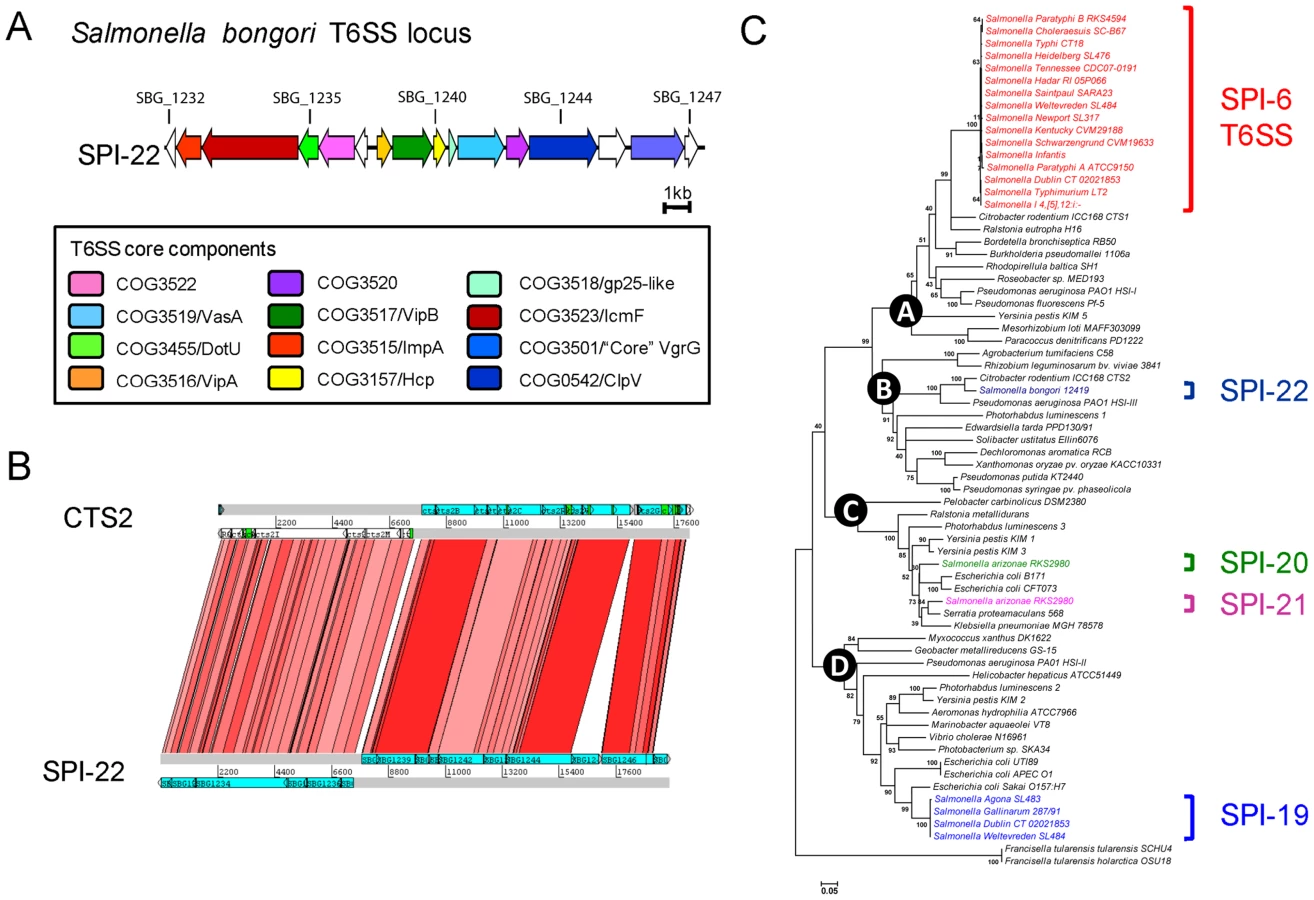
SPI-22: a novel SPI encoding a Type VI secretion system (T6SS)
There are four distinct T6SSs currently described for Salmonella, encoded on SPI-6, SPI-19, SPI-20 and SPI-21 [37], [38]. S. bongori lacks all four systems but carries a novel T6SS locus (∼20 kb in size) which we have denoted SPI-22 (Figure 3A). The T6SS genes carried on SPI-22 shares extensive similarity to the recently identified CTS2 T6SS locus of Citrobacter rodentium ICC168 [39] and the HSI-III locus of Pseudomonas aeruginosa strain PA01 known to be required for virulence (Figures 3B & 3C) [40]. SPI-22 encodes all of the core T6SS components including homologues of DotU and IcmF, necessary for secretion and membrane stabilisation of the T6SS apparatus, the ATPase ClpV, thought to provide energy to the system, as well as other essential functions associated with the T6SS apparatus including VgrG, Hcp and the Gp25-like protein (Figure 3A)[41], [42].
Novel T3SS effector proteins encoded by S. bongori
Apparently in contrast to S. enterica, in the absence of SPI-2 S. bongori has significantly expanded its repertoire of T3SS-1 effector proteins. Most of these candidate effectors are novel within Salmonella but are related to non-locus of enterocyte effacement (LEE) encoded (Nle) effector proteins found in EPEC, enterohemorrhagic E. coli (EHEC) or C. rodentium [39], [43]. These three enteric pathogens have a related infection strategy as they colonize the intestinal mucosa while causing attaching and effacing (A/E) lesions (reviewed by [44]). Of the 12 candidate T3SS-1 effector proteins of S. bongori, SboD, SboE, SboF and SboG show significant sequence similarity to NleI/G (Table 3)[45]. In addition SboC shares 57% amino acid identity with EspJ [46]. Only sboD and sboC genes have homologues in S. enterica subspecies: sboD is similar to an uncharacterized gene, STY1076, that is carried as ‘cargo’ on the S. Typhi prophage 10 [10], [47] and SboC shares 77% amino acid identity with the predicted product of SARI_00261, present at the same locus in S. arizonae.

S. bongori also harbours the first recognized Salmonella chimeric T3SS effector gene, sboH; a fusion of the 5′ 450 bps of Salmonella sopA to the 3′ 828 bps of a gene highly similar to the T3SS effector nleH1 from EPEC, EHEC and C. rodentium (Figures S3 and S4). The sboH gene is found at the same locus as the S. enterica sopA gene and so is likely to have been formed by the insertion and partial deletion of sopA by an nleH1 homologue. By homology the nleH1 portion of the gene is also incomplete, lacking the first 5′ 54 bps. The most obvious effect of this fusion is to replace the cognate export signal of NleH1 (located in the N-terminal 19 amino acids [48]) with the export signal and InvB chaperone-binding site of SopA (located in the N-terminal 45 amino acids [49]) (Figure S3).
The loss of the sopA gene may be compensated for by the presence of two other sopA-related CDSs (Table 3): the product of sboA shares 89% amino acid identity over its full length with SopA, including the export signal, the chaperone binding domain, the invariant cysteine residue and other sites conserved in the C-terminus of this family of proteins (Figure S5 [50]). The sequence conservation between SboB and SopA is limited to the N-terminal 130 amino acids (Figure S3). The remainder of the sequence of SboB is weakly similar to a number of proteins of unknown function from a range of organisms including: S. enterica subsp. arizonae (SARI_00821) and S. enterica subsp. enterica serovar Kentucky (SeKB_A1367: Genbank ABEI01000019), other bacteria including Providencia and eukaryotic proteins including a protein of unknown function from Naegleria gruberi (Amoeba; 38.4% identity [79.1% similarity] in 211 amino acid overlap).
The remaining candidate effectors include SboI, SboJ, SboK and SboL, which all share similarity with leucine rich repeat effectors from S. enterica, such as SlrP, as well as Ipa invasion plasmid antigens from Shigella. Notably SboK is more similar to leucine rich repeat (LRP) effectors found in Edwardsiella and Yersinia spp. than SlrP.
As in many other enteric pathogens, all of the novel S. bongori T3SS effector genes, except sboC, are found on intact or degenerate prophage or regions unique to S. bongori compared to other salmonellae. The exception being sboC which is found on a backbone region conserved only in S. arizonae (Table 3).
Functional analysis of the novel S. bongori effector proteins
To confirm that the candidate T3SS effector proteins could be translocated we selected representatives of all the classes we identified (Table 3) and performed a fluorescence-based ß-lactamase translocation assay [51]. This confirmed that SboA (SopA-like), SboH (SopA - NleH1 chimera), the EspJ homologue SboC, the NleG-family effector SboD and the leucine rich repeat effector SboI were all efficiently translocated into host cells in a T3SS-1 dependent manner (Figure 4A). Translocation of the effector SboI fused to four HA-tags (HAx4) was also visualized by immunofluorescence microscopy of infected cells (Figure 4B). No translocation was observed upon infection with S. bongori ΔinvA expressing SboI-HAx4 or S. bongori wild type expressing the house-keeping protein FabI fused to the HAx4-tag. FabI-HAx4 could be detected inside a few bacteria, which was also sporadically observed for SboI-HAx4 in wild type or ΔinvA strains (data not shown). In contrast, upon infection with S. bongori wild type expressing SboI-HAx4 the effector showed cytoplasmic distribution throughout strongly infected cells, and was also found surrounding a fraction of the bacteria in a ring-like staining pattern, reminiscent of a vacuolar membrane.

Since SboH is the first reported chimeric effector protein we wanted to confirm its function. The EPEC effector NleH1 was recently shown to inhibit apoptosis through a C-terminal interaction with Bax inhibitor 1 [52]. In order to determine if SboH possessed the anti-apoptotic activity of NleH1, we transfected HeLa cells with pRK5-nleH1, pRK5-sboH or a control plasmid pEGFP-N1, treated with the pro-apoptotic compounds tunicamycin (TUN) or brefeldin A and quantified the number of transfected cells showing activation of the apoptosis executioner caspase-3 by immunofluorescence microscopy. SboH prevented activation of caspase-3 by both stimuli as efficiently as NleH1 (Figure 5A & B).

The immunofluorescence analysis of transfected cells indicated that NleH1 and SboH are targeted to different subcellular locations (Figure 5A). Whereas NleH1 shows plasma membrane and perinuclear localization, SboH seemed to localize in discrete structures reminiscent of mitochondria. To substantiate this observation we stained transfected cells with the mitochondrial marker MitoTracker (Figure 5C). This demonstrated that SboH almost exclusively co-localized with the mitochondria, whereas NleH1 did not localize to the mitochondria.
To analyze the impact of SboH in S. bongori infection we performed a cell detachment assay as described by Hemrajani et al.[52]. This assay measures the loss of cells due to S. bongori infection without discriminating specific cell signaling pathways. The assay shows that a S. bongori ΔsboH mutant causes a moderate, but significant increase of 15% in cell loss (p-value<0.001) compared to S. bongori wild type (Figure 5D). Complementation of the S. bongori ΔsboH mutant with SboH restored levels of cell detachment to that of the wild-type.
Taken together, our data suggest that SboH combines features of SopA, namely the mitochondrial targeting signal, with the capability of NleH1 to inhibit tunicamycin and brefeldin A induced apoptosis. During infection, most likely through its anti-apoptotic activity, SboH reduces bacterial cytotoxicity and host cell loss.
Metabolic functions mark the evolutionary divergence of S. bongori and S. enterica
We used genome sequence data to explore the distribution of metabolic pathways and their associated genes within the Salmonella serovars. As direct comparisons of the metabolic maps of S. Typhimurium and E. coli have been reported previously [53] we focused our analysis on comparing S. bongori to other Salmonella serovars building upon data from S. bongori 12419 (Figure 2). Arguably one of the most distant comparisons we could make across the salmonellae would be between S. bongori and the acutely pathogenic, human restricted, S. enterica subspecies enterica serovar Typhi (S. Typhi). Within this comparison we found a surprisingly high degree of conservation. All of the thirty pathways known to be involved in the generation of precursor metabolites and energy for S. Typhi are present in S. bongori (Table S4). Of the 146 predicted biosynthetic pathways found in S. Typhi, including the biosynthesis of amino acids, carbohydrates, fatty acids and lipids, only 8 are missing from S. bongori. Equally, of the 78 degradative pathways carried by S. Typhi, S. bongori shares 72 and possesses only 3 unique pathways.
The unique S. bongori metabolic capabilities (compared to S. Typhi) include the degradation of complex acid sugars D-galacturonate and L-idonate, which are sporadically distributed throughout the salmonellae. However, more restricted in its distribution is the capacity to degrade lactose. S. bongori encodes β-D-galactosidase (lacZ) and the lactose operon repressor (lacI) but is missing lacY (the high affinity lactose permease) which explains why although it is a non-lactose fermenter S. bongori gives a positive result on ortho-nitrophenyl-β-D-galactopyranoside (ONPG) medium. Most members of S. enterica subspecies enterica are phenotypically non-lactose fermenters and are unable to utilise ONPG. The only other salmonellae that appear to harbour the lac operon include S. enterica subspecies arizonae and diarizonae, but these subspecies also possess lacY. Comparing S. bongori to other Enterobacteriaceae which are non-lactose fermenters but are ONPG positive there is no strong conservation in genes or the site of insertion, suggesting independent acquisition events [54], [55].
There is also evidence of metabolic streamlining in S. bongori. For example, like E. coli, S. bongori has lost the cob-pdu gene cluster (S. bongori retains only fragments of the first and last gene of the cob-pdu cluster: cobT [SBG__1882] and pduX [SBG_1882A]) and so lacks the capacity to anaerobically synthesise vitamin B12 (cobalamine) and to catabolise propanediol [56]. The cob-pdu gene cluster was thought to have been lost by many Enterobacteriaceae. It has been suggested that the cob-pdu gene cluster was subsequently reacquired by S. enterica, following its split from S. bongori [57], [58], [59], where it has been shown to be important for survival in macrophages [60] a niche in which S. bongori cannot survive [61].
Conversely there is evidence that pathways conserved in S. bongori, E. coli and wider Enterobacteriaceae, have been lost and replaced in the warm blooded-host adapted serovars of S. enterica subspecies enterica by alternative pathways that are energetically more efficient producing more ATP/mol of substrate or have differing substrate specificities.
S. bongori carries the same genes for L-tartrate or citrate fermention as those found in E. coli and most other enterics: the ttdABDT operon or citDEF. However, these have been either replaced (only remnants of the ttd genes remain in members of subspecies enterica e.g. S. Typhi strain CT18 position 3230063..3230191 and S. Enteritidis strain P125109, SEN3049A) or, in the case of citrate, augmented in S. enterica subspecies enterica by the acquisition of two operons encoding tartrate hydratase (e.g. STM3350-3359) and a second citrate lyase gene cluster (e.g. STM0052-STM0063). Both of these clusters carry a dedicated Na+ translocating oxaloacetate decarboxylase to provide reducing power. Therefore unlike the pathways found in S. bongori and E. coli these new pathways do not require a co-substrate and are energetically more efficient, producing more ATP/mol of substrate [62], [63], [64]. Moreover mutations in these new gene clusters in S. Typhimurium can be found as attenuating in genome wide mouse mutagenesis studies [65] and our microarray analysis of SARC shows that all other S. enterica subspecies resemble S. bongori by possessing the ttd cluster but lacking the alternate tartrate and citrate dissimilatory operons (data not shown). Looking more broadly across the Enterobacteriaceae, some Klebsiella pnenumoniae isolates also possesses a related second citrate lyase, sometimes located at the same site in the genome as that found in S. enterica subspecies enterica serovars. However, the gene makeup of this region in K. pnenumoniae differs slightly and we could not find this region at this site in other enterics we searched consistent with this region being sporadically acquired.
There are other examples of lineage-specific metabolic streamlining which show a more sporadic phylogenetic distribution including the C-P lyase system (phnA-P), able to breakdown a wide range of phosphonate compounds, and phosphonatase which is specific for 2-aminoethylphosphonate. Whilst S. enterica subspecies arizonae carries the entire cluster (data not shown) the S. bongori phn loci is degenerate, consisting of only phnOAB and remnants of phnP and phnN (SBG_3727 and SBG_3728A, respectively).
S. enterica subspecies enterica has also lost the majority of genes in this operon leaving only phnOAB [66], but have acquired the phosphonatase system encoded by phnVUTSRWX [66]. The explanation for this replacement in members of subspecies enterica may lie in that fact that 2-aminoethylphosphonate is found in abundance in flagellates found in the digestive tracts of ruminants such as cattle, common hosts for members of subspecies enterica [67]. Our microarray data for this cluster also shows that genes phnVUTSRWX are only present in S. enterica subspecies enterica (data not shown).
The capacity to use allantoin as a sole nitrogen source under anaerobic conditions is also phylogenetically restricted. We have previously speculated that the acquisition of the allantoin gene cluster by S. enterica subsp. enterica was linked to differences in the sequential breakdown of purines by different hosts [68], [69]: in fish, crustaceans and other invertebrates purines are broken down sequentially to ammonia and CO2 [70], [71], while genetic lesions in vertebrate species block purine catabolism at different steps leading to the accumulation of allantoin in most mammals (including rodents and domesticated animals). Our current data shows that the genes encoding allantoin degradation in Salmonella are absent from S. bongori and restricted to S. enterica subspecies enterica and salamae only.
Discussion
Our understanding of Salmonella evolution has been built largely on data from representative isolates of the relatively recently emerged Salmonella enterica subspecies enterica. S. bongori and S. enterica are thought to have diverged between 40-63.4 Myrs ago [72] and so comparing the genomes of these two distinct species provides a unique opportunity to understand the ancestral Salmonella and determine the evolutionary events that mark speciation and those that track different branch points in Salmonella evolution following this event.
We have shown that a diverse set of S. bongori isolates form a tight cluster of sequence types that, when examined on a whole genome basis, appear to comprise at least three phylogenetic groups. The G+C content in S. bongori represents a midpoint between S. enterica subspecies enterica and E. coli (Figure S2), but the phylogenetic analysis with MLST data and whole genome sequences suggests that this may be an artifact of the host specialisation into warm blooded animals that took place in the evolution of S. enterica subspecies enterica. The variation across the S. bongori species, both within and between the phylogenetic groups, contrasts sharply with that observed across S. enterica, with a comparable amount of variation across the groups in S. bongori, to that which is found between two serotypes of S. enterica subspecies enterica. The apparent lack of variation is difficult to explain since even if S. bongori had been stably maintained for a long period within its current niche significant genome diversity would still be expected, even if this was largely neutral. This reduced level of apparent diversity could be a bias of sampling, yet the isolates sequenced in this study are from a wide range of sources and are globally and temporally diverse. Although the answer to this question is still equivocal it is possible that S. bongori has been through a recent evolutionary bottleneck.
Despite their apparent evolutionary divergence, metabolic analysis showed that the biochemical maps of S. bongori and S. Typhi are very similar, suggesting that Salmonella serovars acquired many of the basic functions for an enteric lifestyle early in their evolution. This has been recently supported by the finding that the ability to use tetrathionate as an electron acceptor provides a competitive advantage to S. Typhimurium in the inflamed gut over normal flora [73]. The ttr cluster is conserved in S. bongori [74].
Where S. bongori did differ from S. enterica serovars, it generally most closely resembled E. coli and the wider Enterobacteriaceae, i.e. the presumed ancestral state. Whilst the independent acquisition of the lac operon by S. bongori is difficult to explain, for E. coli the acquisition of the lac operon may have facilitated metabolism of milk sugar and adaptation to the mammalian gut. Conversely the loss of this function from S. enterica subspecies enterica may be associated with its invasive lifestyle since recent evidence has shown that lacI expression interferes with the function of SPI-2 and attenuates virulence in macrophage [75].
This comparative analysis also highlighted metabolic traits that mark the evolution of S. enterica subspecies enterica including the differing abilities to ferment L-tartrate and citrate. These metabolic differences are already known to differentiate high and low pathogenicity Salmonella strains (S. Paratyphi B and S. Paratyphi B variant Java) [76]. Outside of Salmonella the ability to ferment citrate almost equally divides clinical Klebsiella pneumoniae biotypes into two groups and is thought to represent an adaptation to different nutrient conditions found within the host [77].
S. bongori possess a basic Salmonella virulence ‘tool kit’ consisting of SPI-1, 3a, 3b, 4, 5 and 9. Although the S. bongori SPI-3 and SPI-5 have a different structure compared to those in S. enterica these SPIs are conserved across the salmonellae and could be considered part of the Salmonella core genome. Moreover, many of these core SPIs show significant regulatory and functional interplay between the functions they encode. For example the SPI-4 adhesin SiiE is required for efficient translocation of T3SS-1 effectors in S. Typhimurium [78] and SPI-4, SPI-5 and SPI-1 genes are under joint control by the SirA/HilA global regulatory cascade [78], [79], [80], [81]. From this it is tempting to speculate that these SPIs define one of the earliest virulence networks of Salmonella. Clearly there are multiple factors missing from S. bongori which limit its ability to cause disease in warm blooded animals demonstrated by experiments that have introduced SPI-2 into S. bongori [61].
The genome of S. bongori has not remained static since divergence; we see parallels with S. enterica serovars in the functions that have been acquired following divergence. For example both S. bongori and S. enterica have independently acquired different T6SSs. It is clear that the Salmonella genus as a whole includes representatives of each of the major T6SS phylogenetic groups, thus reinforcing the long-term importance of these systems. Also like S. enterica, S. bongori has sequentially acquired a range of T3SS-1 effector proteins many of which we have shown to be translocated. The S. bongori effectors have homologues in EPEC, EHEC and C. rodentium including EspJ; which in EPEC and EHEC prevents receptor mediated phagocytosis of opsonised cells [46] and so could be important for S. bongori in resisting phagocytosis. This strategy would be well in line with previous observations of the S. bongori life style as the bacteria are not able to sustain an intracellular life style in macrophages mainly due to absence of the SPI-2 T3SS-2 and its effectors [61] and likely also because of the lack of cob-pdu operon too [60]. We also functionally characterised the effector SboH and have shown that it inhibits apoptosis in a similar manner to its EPEC homologue NleH1. Moreover, in infection SboH reduces bacterial cytotoxicity. In EPEC the anti-apoptotic activity of NleH1 has been proposed to sustain colonisation of the mucosal epithelium by reducing the ‘turn-over’ of surface enterocytes and associated any bacteria or microcolonies [52]. The acquisition of these effectors that most closely resemble those from pathogenic E. coli strains causing watery diarrhea suggests that following the split of S. bongori and S. enterica, S. bongori has adopted a specialised infection strategy which might in parts be more similar to the extracellular pathogenic E. coli than S. enterica. This infection strategy might be optimised to colonise cold-blooded reptiles, but still provides the basic armoury for S. bongori to emerge as an opportunistic pathogen of humans and animals.
Materials and Methods
Source and details of bacterial strains
The S. bongori exploited in this study included 28 isolates originating from between 1966-2004, from the USA, Africa and Europe, from hosts including humans, frogs, pigeons and reptiles as well as environmental sources including the shell of a hen's egg, cheese, fishmeal and waste water (see Table S1).
Genome sequencing assembly, mapping and phylogeny
For bacterial cultures LB medium was inoculated and grown overnight at 37°C with each isolate. Genomic DNA was extracted from 1 ml of culture by using manufacturer's instructions (Wizard Genomic DNA Purification kit from Promega).
The genome of S. bongori strain 12419 was sequenced to approximately 11-fold coverage from pUC19 (insert size 2.8–5.5 kb) and pMAQ1b_SmaI (insert size 5.5–6.0 kb) genomic shotgun libraries using big-dye terminator chemistry on ABI3700 automated sequencers. End sequences from large insert BAC libraries in pBACe3.6 (insert size 23–48 kb) were used as a scaffold. All repeat regions were bridged by read-pairs or end-sequenced polymerase chain reaction (PCR) products.
For all remaining S. bongori strains, tagged genomic library preparation and DNA sequencing (with and without multiplexing) was carried out as previously described [82]. Mapping of reads to the reference genome and SNP detection were carried out according to earlier described protocols [82]. De novo assemblies were performed by using Velvet v0.7.03 and their corresponding contigs were ordered using Abacas [82]; the resulting pseudomolecules were blasted against the reference genome to assess synteny as well as the existence of indels and novel regions. Details of mapping and assembly data output are given in Table S1. Annotation and analysis was performed using Artemis and ACT [83], [84].
Phylogenic analysis of Salmonella (shown in Figure 1) was based on the 7 concatenated MLST loci sequences, from sequences generated in this study (for the S. bongori isolates), those obtained from the Salmonella MLST Public Strains Database (http://mlst.ucc.ie/mlst/dbs/Senterica for the S. enterica STs) or obtained from genomic sequence (for EPEC strain E2348/69 [43]). The S. bongori tree was produced using a whole genome alignment generated by mapping the S. bongori samples against the finished genome of strain 12419. Trees were drawn using RAxML assuming a general time reversible site model with gamma correction [85]. In the case of the whole genome tree, phage and MGEs were removed prior to the production of the tree (see Table S2). Support for nodes was assessed by using bootstrapping (x100); SNPs were reconstructed on the tree with parsimony using accelerated transformation. BAPS analysis was performed on the SNP alignments produced from the mapping alignment, using the BAPS individual mixture model [22]. Three independent iterations of BAPS were performed (using an upper limit for the number of populations of 25, 26 and 27) to obtain the most optimal partitioning of the sample.
Defining orthologous gene-sets
To infer the orthologous genes in each pair of genomes compared: Each CDS (a) from the genome (A) was searched, using FASTA, against the CDSs of the other genome (B). If the top hit covered at least 80% of the length of both sequences with at least 30% identity, a reciprocal FASTA search of the top hit sequence (b) was launched against the CDSs of the first genome. If the reciprocal top hit was the same as the original query CDS then (a) and (b) are considered orthologous genes of (A) and (B). In a second step, in order to validate the results, we performed a BLASTN and TBLASTX comparison between the 15 genomes, visualized using ACT [83] to curate ambiguous cases, for example, gene remnants (pseudogenes), IS elements and phage-related CDSs, and to check for a syntenic relationship among the putative orthologs.
Pathway Tools
A Pathway/Genome Database (PGDB) describing the metabolic pathways of S. Typhi was created in Pathway Tools v. 13.5 (SRI International, California) using the genome sequence and annotation associated with strain CT18 [10]. This PGDB underwent manual curation and currently comprises 200 predicted metabolic pathways and over 130 predicted transport reactions. To determine the differences in S. bongori relative to S. Typhi, we mapped orthologues onto the pathways and transport reactions, subsequently removing those missing functions and adding in the functions unique to S. bongori (summarised in Figure 2 and listed in Table S4).
S. bongori deletion mutant and novel T3SS effector plasmid construction
Primers, restriction enzymes and plasmids used to create S. bongori deletion mutants and to construct expression vectors of the putative S. bongori effector proteins are listed in Table S3.
Specific gene knockouts of the invA or sboH genes were generated in S. bongori 12419 as described previously [86]. To create non-polar mutations, the kanamycin resistance cassette was removed using plasmid pCP20 leaving a scar of 84 bp [86], [87]. Deletions were confirmed by PCR and sequencing from the regions flanking the knockouts.
To obtain the vectors encoding ß-lactamase (TEM1) fusions all genes were PCR-amplified from S. bongori strain 12419 genomic DNA and PCR products digested and ligated into pCX340 [51] or pRK5, respectively. If the KpnI restriction site of pCX340 was used a new ribosome binding site (RBS) was included in the forward primer. Sequence identity of the constructs was verified by DNA sequencing. The pCX340 derivative plasmids were named pICC522 (fabI), pICC523 (sboA), pICC524 (sboC), pICC525 (sboD), pICC526 (sboI) and pICC527 (sboH), pICC611 (sopB), pICC612 (sopD).
To create a plasmid allowing the C-terminal fusion of four HA-tags to the effectors, pCX340 was digested with EcoRI and XbaI to remove the tem1 gene. Subsequently an oligo cassette encoding four HA-tags was ligated into the vector to give the plasmid pICC613. PCR products of sboI and fabI and pICC613 were digested and ligated as described for the vectors encoding TEM1 fusions to yield pICC614 (pSboI-HAx4) and pICC615 (pFabI-HAx4). The plasmid pICC616 (pSboH) allowing the inducible expression of untagged SboH was constructed by ligation of the sboH PCR product in EcoRI and XbaI digested pCX340 and the transfection vector pRK5-SboH (pICC548) by ligation of the PCR product into pRK5 (Clontech). All pICC plasmids were used to transform S. bongori strain 12419 wild type and mutant strains by electroporation.
Translocation assay
The ß-lactamase (TEM1)-translocation assay for the identification of translocated effector proteins was adapted from a protocol previously described [51]. To obtain a confluent cell layer 4.0×104 HeLa cells were seeded in 200 µL DMEM (Sigma, 1000 mg/L glucose, supplemented with 10% fetal calf serum, Glutamax (Invitrogen) and MEM non-essential amino acids (Sigma)) growth medium per well of a black wall/clear-flat bottom 96 well plate (Becton Dickinson) and cultured overnight. Prior to infection the medium was replaced with 150 µL fresh growth medium. Overnight LB broth cultures (6 µg/mL tetracycline) of S. bongori strain 12419 or the ΔinvA mutant carrying the pICC plasmids were diluted 1∶30 in LB broth (6 µg/mL tetracycline) and grown to an OD600 of 1.1–1.4 before protein expression was induced by addition of 1 mM Isopropyl β-D-1-thiogalactopyranoside (IPTG). The induced cultures were incubated until an OD600 of 1.8–2.2 was reached and diluted in Dulbecco's-PBS (D-PBS, Sigma) to a concentration of 3.75×108 bacteria/mL. To infect HeLa cells 20 µL bacterial dilution per well was added and the infection was synchronized by centrifugation (900 g, 5 min). After 1 h incubation at 37°C, 5% CO2 the cell supernatant was replaced with 100 µL Hanks' Buffered Salt Solution (Gibco, supplemented with 20 mM HEPES, 3 mM Probenecid (Sigma) pH 7.4 designated HBSS-HP), and 20 µL freshly prepared CCF2-AM ß-lactamase substrate (LiveBLAzer FRET-B/G Loading Kit, Invitrogen) were added. After 1 h 45 min incubation at room temperature in the dark the cells were washed five times with HBSS-HP. Fluorescence emission at 450 nm and 520 nm was measured from the bottom using a Fluostar Optima plate reader (excitation wavelength 410 nm, 10-nm band-pass). The translocation rate was calculated as recommended in the LiveBLAzer FRET-B/G Loading Kit manual. Briefly, emission values were first corrected by subtraction of the average background signals recorded for empty wells and the mean 450 nm/520 nm emission ratio of a triplicate of wells was calculated for each sample. The translocation rate is expressed as fold increase of the mean emission ratio 450/520 nm of each infected sample in relation to the mean emission ratio of uninfected cells. Expression of the TEM1 fusion proteins was controlled by Western blot using a mouse anti-ß-lactamase antibody (QED Bioscience Inc; data not shown).
Epifluorescence microscopy of translocated effectors
HeLa cells (1.25×105 per 24-well plate well) were seeded on coverslips and incubated in growth media overnight in a humidified atmosphere of 5% CO2 at 37 °C. S. bongori wild type or ΔinvA mutant containing pICC614 or pICC614 plasmids were grown, diluted and 50 µL dilution used for infection as described for the translocation assay. 1 h45 min –2 h post infection cells were washed three times with D-PBS, fixed with 3% paraformaldehyde (PFA), treated with 50 mM NH4Cl in D-PBS, washed three times with D-PBS, permeabilised with 0.1% (v/v) Triton X-100, washed three times with D-PBS and blocked with 2% (w/v) bovine serum albumin (BSA) and 2% (v/v) natural donkey serum in D-PBS for 1 h. The samples were stained with rabbit anti-Salmonella (O:66 Statens Serum Institute) and mouse anti-HA.11 (Covance) primary antibodies followed by Rhodamine Red X (RRX)-conjugated donkey anti-rabbit IgG and DyLight 488-conjugated donkey anti-mouse IgG (both Jackson ImmunoResearch) antibodies. Nuclei were labelled with Hoechst 33342 dye and F-actin with AlexaFluor647 phalloidin (Invitrogen). The coverslips were mounted using ProLong Gold antifade reagent (Invitrogen) and analysed on Zeiss Axio Imager Z1 or M1 immunofluorescence microscopes with Axiovision Rel 4.8 software.
Epifluorescence microscopy and caspase-3 activation assay
The experiments to compare the localization and anti-apoptotic activity of NleH1 and SboH were performed as described previously [52], [88]. Briefly, transfected HeLa cells were treated with either 5 µg/ml tunicamycin (TUN) or 10 µg/ml brefeldin A (BFA) for 18 hours, or left untreated, prior to immunofluorescence microscopy processing. The cells were fixed in 3% PFA, washed with PBS, treated with 10 mM NH4Cl, permeabilized with 0.2% (v/v) Triton X-100, washed with PBS and blocked with 1% (v/v) BSA in PBS for 1 h. Active caspase-3 and Myc-tagged effector proteins were detected using rabbit anti-cleaved caspase-3 (Cell Signalling Technology), RRX-conjugated donkey anti-rabbit IgG (Jackson ImmunoResearch) and Fluorescein Isothiocyanate (FITC)-conjugated monoclonal mouse anti-Myc (Sigma) antibodies. Nuclei were labelled with the Hoechst 33342 reagent (Invitrogen). Mitochondria were visualised using MitoTracker (Invitrogen) in accordance with manufacturer's guidelines before fixation. Samples were mounted and analysed by microscopy as described above. To determine the number of apoptotic cells 100 transfected cells were analysed in each repeat. Samples were tested in triplicate and experiments repeated a minimum of three times.
Cell detachment assay
HeLa cells (7.2×104 per 24-well plate well) were cultured overnight. S. bongori wild type, ΔinvA,ΔsboH or ΔsboH pICC616 were grown for infection as described above. Prior infection 1 mL bacterial culture was harvested by centrifugation, resuspended in the same volume of cell culture medium and used to infect HeLa cells. After 1 h cells were treated with 200 µg/ml gentamicin and incubated for 4 h. A control sample was incubated with 1 µM staurosporine (STS) for 5 h in parallel to the infection. Cells were washed 5 times with PBS and then trypsinized for 10 min. Trypsin was inactivated by addition of 700 µL growth medium. Cells were counted on a Neubauer hemocytometer. All counts were compared with the level of uninfected, untreated cells and plotted as a percentage of cells lost. Statistical analysis was done using the GraphPad InStat Version 3.06 software. The one-way ANOVA Test using Bonferroni correction was used to determine significance of the observed differences (p-values<0.001).
Accession numbers
The annotated genome sequence of Salmonella bongori strain 12419 has been deposited in the public databases under the accession numbers FR877557. The Illumina sequencing reads for all the sequences generated in this study have been deposited in the European Nucleotide Archive (ENA) under the accession numbers ERS002029 - ERS002042 (inclusive), ERS002044, ERS004246, ERS004249, ERS004170, ERS004173 - ERS004176 (inclusive), ERS004190 - ERS004193 (inclusive) and ERS004196. This is matched to strain names in Table S1. Microarray data was submitted to ArrayExpress under accession number E-TABM-931.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. DoolittleRFFengDFTsangSChoGLittleE 1996 Determining divergence times of the major kingdoms of living organisms with a protein clock. Science 271 470 477
2. BrennerFWVillarRGAnguloFJTauxeRSwaminathanB 2000 Salmonella nomenclature. J Clin Microbiol 38 2465 2467
3. CrosaJHBrennerDJEwingWHFalkowS 1973 Molecular relationships among the Salmonelleae. J Bacteriol 115 307 315
4. KauffmannF 1966 [On the history of salmonella research]. Zentralbl Bakteriol Orig 201 44 48
5. Le MinorLVeronMPopoffM 1982 [The taxonomy of Salmonella]. Ann Microbiol (Paris) 133 223 243
6. Le MinorLVeronMPopoffM 1982 [A proposal for Salmonella nomenclature]. Ann Microbiol (Paris) 133 245 254
7. NastasiAMamminaCVillafrateMRMassentiMFScarlataG 1988 Multiple typing of strains of Salmonella enterica subsp. bongori ser. 48:Z35: - isolated in southern Italy. Ann Inst Pasteur Microbiol 139 605 612
8. GiammancoGMPignatoSMamminaCGrimontFGrimontPA 2002 Persistent endemicity of Salmonella bongori 48:z(35):--in Southern Italy: molecular characterization of human, animal, and environmental isolates. J Clin Microbiol 40 3502 3505
9. McClellandMFloreaLSandersonKCliftonSWParkhillJ 2000 Comparison of the Escherichia coli K-12 genome with sampled genomes of a Klebsiella pneumoniae and three salmonella enterica serovars, Typhimurium, Typhi and Paratyphi. Nucleic Acids Res 28 4974 4986
10. ParkhillJDouganGJamesKDThomsonNRPickardD 2001 Complete genome sequence of a multiple drug resistant Salmonella enterica serovar Typhi CT18. Nature 413 848 852
11. McClellandMSandersonKESpiethJCliftonSWLatreilleP 2001 Complete genome sequence of Salmonella enterica serovar Typhimurium LT2. Nature 413 852 856
12. SchmidtHHenselM 2004 Pathogenicity islands in bacterial pathogenesis. Clin Microbiol Rev 17 14 56
13. GalanJE 1996 Molecular genetic bases of Salmonella entry into host cells. Mol Microbiol 20 263 271
14. HenselMSheaJEBaumlerAJGleesonCBlattnerF 1997 Analysis of the boundaries of Salmonella pathogenicity island 2 and the corresponding chromosomal region of Escherichia coli K-12. J Bacteriol 179 1105 1111
15. HenselMSheaJEWatermanSRMundyRNikolausT 1998 Genes encoding putative effector proteins of the type III secretion system of Salmonella pathogenicity island 2 are required for bacterial virulence and proliferation in macrophages. Mol Microbiol 30 163 174
16. CirilloDMValdiviaRHMonackDMFalkowS 1998 Macrophage-dependent induction of the Salmonella pathogenicity island 2 type III secretion system and its role in intracellular survival. Mol Microbiol 30 175 188
17. HelaineSThompsonJAWatsonKGLiuMBoyleC 2010 Dynamics of intracellular bacterial replication at the single cell level. Proc Natl Acad Sci U S A 107 3746 3751
18. OchmanHGroismanEA 1996 Distribution of pathogenicity islands in Salmonella spp. Infect Immun 64 5410 5412
19. KidgellCReichardUWainJLinzBTorpdahlM 2002 Salmonella typhi, the causative agent of typhoid fever, is approximately 50,000 years old. Infect Genet Evol 2 39 45
20. BoydEFWangFSWhittamTSSelanderRK 1996 Molecular genetic relationships of the salmonellae. Appl Environ Microbiol 62 804 808
21. CoranderJTangJ 2007 Bayesian analysis of population structure based on linked molecular information. Math Biosci 205 19 31
22. CoranderJMarttinenPSirenJTangJ 2008 Enhanced Bayesian modelling in BAPS software for learning genetic structures of populations. BMC Bioinformatics 9 539
23. SharpPMEmeryLRZengK 2010 Forces that influence the evolution of codon bias. Philos Trans R Soc Lond B Biol Sci 365 1203 1212
24. CookeFJWainJFookesMIvensAThomsonN 2007 Prophage sequences defining hot spots of genome variation in Salmonella enterica serovar Typhimurium can be used to discriminate between field isolates. J Clin Microbiol 45 2590 2598
25. Blanc-PotardABSolomonFKayserJGroismanEA 1999 The SPI-3 pathogenicity island of Salmonella enterica. J Bacteriol 181 998 1004
26. KnodlerLACelliJHardtWDVallanceBAYipC 2002 Salmonella effectors within a single pathogenicity island are differentially expressed and translocated by separate type III secretion systems. Mol Microbiol 43 1089 1103
27. HenselMNikolausTEgelseerC 1999 Molecular and functional analysis indicates a mosaic structure of Salmonella pathogenicity island 2. Mol Microbiol 31 489 498
28. TsolisRMAdamsLGFichtTABaumlerAJ 1999 Contribution of Salmonella typhimurium virulence factors to diarrheal disease in calves. Infect Immun 67 4879 4885
29. Collier-HyamsLSZengHSunJTomlinsonADBaoZQ 2002 Cutting edge: Salmonella AvrA effector inhibits the key proinflammatory, anti-apoptotic NF-kappa B pathway. J Immunol 169 2846 2850
30. MiroldSRabschWRohdeMStenderSTschapeH 1999 Isolation of a temperate bacteriophage encoding the type III effector protein SopE from an epidemic Salmonella typhimurium strain. Proc Natl Acad Sci U S A 96 9845 9850
31. EhrbarKHardtWD 2005 Bacteriophage-encoded type III effectors in Salmonella enterica subspecies 1 serovar Typhimurium. Infect Genet Evol 5 1 9
32. ShahDHLeeMJParkJHLeeJHEoSK 2005 Identification of Salmonella gallinarum virulence genes in a chicken infection model using PCR-based signature-tagged mutagenesis. Microbiology 151 3957 3968
33. VernikosGSParkhillJ 2006 Interpolated variable order motifs for identification of horizontally acquired DNA: revisiting the Salmonella pathogenicity islands. Bioinformatics 22 2196 2203
34. PorwollikSMcClellandM 2007 Determination of the gene content of Salmonella genomes by microarray analysis. Methods Mol Biol 394 89 103
35. PorwollikSBoydEFChoyCChengPFloreaL 2004 Characterization of Salmonella enterica subspecies I genovars by use of microarrays. J Bacteriol 186 5883 5898
36. KingsleyRAHumphriesADWeeningEHDe ZoeteMRWinterS 2003 Molecular and phenotypic analysis of the CS54 island of Salmonella enterica serotype typhimurium: identification of intestinal colonization and persistence determinants. Infect Immun 71 629 640
37. BlondelCJJimenezJCContrerasISantiviagoCA 2009 Comparative genomic analysis uncovers 3 novel loci encoding type six secretion systems differentially distributed in Salmonella serotypes. BMC Genomics 10 354
38. FolkessonALofdahlSNormarkS 2002 The Salmonella enterica subspecies I specific centisome 7 genomic island encodes novel protein families present in bacteria living in close contact with eukaryotic cells. Res Microbiol 153 537 545
39. PettyNKBulginRCrepinVFCerdeno-TarragaAMSchroederGN 2009 The Citrobacter rodentium genome sequence reveals convergent evolution with human pathogenic Escherichia coli. J Bacteriol 192 525 538
40. LesicBStarkeyMHeJHazanRRahmeLG 2009 Quorum sensing differentially regulates Pseudomonas aeruginosa type VI secretion locus I and homologous loci II and III, which are required for pathogenesis. Microbiology 155 2845 2855
41. ZhengJLeungKY 2007 Dissection of a type VI secretion system in Edwardsiella tarda. Mol Microbiol 66 1192 1206
42. MougousJDCuffMERaunserSShenAZhouM 2006 A virulence locus of Pseudomonas aeruginosa encodes a protein secretion apparatus. Science 312 1526 1530
43. IguchiAThomsonNROguraYSaundersDOokaT 2009 Complete genome sequence and comparative genome analysis of enteropathogenic Escherichia coli O127:H6 strain E2348/69. J Bacteriol 191 347 354
44. FrankelGPhillipsAD 2008 Attaching effacing Escherichia coli and paradigms of Tir-triggered actin polymerization: getting off the pedestal. Cell Microbiol 10 549 556
45. LiMRosenshineIYuHBNadlerCMillsE 2006 Identification and characterization of NleI, a new non-LEE-encoded effector of enteropathogenic Escherichia coli (EPEC). Microbes Infect 8 2890 2898
46. MarchesOCovarelliVDahanSCougouleCBhattaP 2008 EspJ of enteropathogenic and enterohaemorrhagic Escherichia coli inhibits opsono-phagocytosis. Cell Microbiol 10 1104 1115
47. ThomsonNBakerSPickardDFookesMAnjumM 2004 The role of prophage-like elements in the diversity of Salmonella enterica serovars. J Mol Biol 339 279 300
48. Garcia-AnguloVADengWThomasNAFinlayBBPuenteJL 2008 Regulation of expression and secretion of NleH, a new non-locus of enterocyte effacement-encoded effector in Citrobacter rodentium. J Bacteriol 190 2388 2399
49. HigashideWZhouD 2006 The first 45 amino acids of SopA are necessary for InvB binding and SPI-1 secretion. J Bacteriol 188 2411 2420
50. ZhangYHigashideWMMcCormickBAChenJZhouD 2006 The inflammation-associated Salmonella SopA is a HECT-like E3 ubiquitin ligase. Mol Microbiol 62 786 793
51. CharpentierXOswaldE 2004 Identification of the secretion and translocation domain of the enteropathogenic and enterohemorrhagic Escherichia coli effector Cif, using TEM-1 beta-lactamase as a new fluorescence-based reporter. J Bacteriol 186 5486 5495
52. HemrajaniCBergerCNRobinsonKSMarchesOMousnierA 2010 NleH effectors interact with Bax inhibitor-1 to block apoptosis during enteropathogenic Escherichia coli infection. Proc Natl Acad Sci U S A 107 3129 3134
53. AbuOunMSuthersPFJonesGICarterBRSaundersMP 2009 Genome scale reconstruction of a Salmonella metabolic model: comparison of similarity and differences with a commensal Escherichia coli strain. J Biol Chem 284 29480 29488
54. SmithHWParsellZ 1975 Transmissible substrate-utilizing ability in enterobacteria. J Gen Microbiol 87 129 140
55. WaliaSKMadhavanTChughTDSharmaKB 1987 Characterization of self-transmissible plasmids determining lactose fermentation and multiple antibiotic resistance in clinical strains of Klebsiella pneumoniae. Plasmid 17 3 12
56. RothJRLawrenceJGBobikTA 1996 Cobalamin (coenzyme B12): synthesis and biological significance. Annu Rev Microbiol 50 137 181
57. LawrenceJGRothJR 1996 Evolution of coenzyme B12 synthesis among enteric bacteria: evidence for loss and reacquisition of a multigene complex. Genetics 142 11 24
58. PorwollikSWongRMMcClellandM 2002 Evolutionary genomics of Salmonella: gene acquisitions revealed by microarray analysis. Proc Natl Acad Sci U S A 99 8956 8961
59. PrenticeMBCuccuiJThomsonNParkhillJDeeryE 2003 Cobalamin synthesis in Yersinia enterocolitica 8081. Functional aspects of a putative metabolic island. Adv Exp Med Biol 529 43 46
60. KlumppJFuchsTM 2007 Identification of novel genes in genomic islands that contribute to Salmonella typhimurium replication in macrophages. Microbiology 153 1207 1220
61. Hansen-WesterIChakravorttyDHenselM 2004 Functional transfer of Salmonella pathogenicity island 2 to Salmonella bongori and Escherichia coli. Infect Immun 72 2879 2888
62. BottM 1997 Anaerobic citrate metabolism and its regulation in enterobacteria. Arch Microbiol 167 78 88
63. WoehlkeGDimrothP 1994 Anaerobic growth of Salmonella typhimurium on L(+) - and D(-)-tartrate involves an oxaloacetate decarboxylase Na+ pump. Arch Microbiol 162 233 237
64. LutgensMGottschalkG 1980 Why a co-substrate is required for anaerobic growth of Escherichia coli on citrate. J Gen Microbiol 119 63 70
65. LawleyTDChanKThompsonLJKimCCGovoniGR 2006 Genome-wide screen for Salmonella genes required for long-term systemic infection of the mouse. PLoS Pathog 2 e11
66. JiangWMetcalfWWLeeKSWannerBL 1995 Molecular cloning, mapping, and regulation of Pho regulon genes for phosphonate breakdown by the phosphonatase pathway of Salmonella typhimurium LT2. J Bacteriol 177 6411 6421
67. McNaughtMlOwenECHenryKMKonSK 1954 The utilization of non-protein nitrogen in the bovine rumen. VIII. The nutritive value of the proteins of preparations of dried rumen bacteria, rumen protozoa and brewer's yeast for rats. Biochem J 56 151 156
68. KingsleyRAMsefulaCLThomsonNRKariukiSHoltKE 2009 Epidemic multiple drug resistant Salmonella Typhimurium causing invasive disease in sub-Saharan Africa have a distinct genotype. Genome Res 19 2279 2287
69. MatiasovicovaJAdamsPBarrowPAHradeckaHMalcovaM 2007 Identification of putative ancestors of the multidrug-resistant Salmonella enterica serovar typhimurium DT104 clone harboring the Salmonella genomic island 1. Arch Microbiol 187 415 424
70. FujiwaraSNoguchiT 1995 Degradation of purines: only ureidoglycollate lyase out of four allantoin-degrading enzymes is present in mammals. Biochem J 312 Pt 1 315 318
71. TakadaYNoguchiT 1986 Ureidoglycollate lyase, a new metalloenzyme of peroxisomal urate degradation in marine fish liver. Biochem J 235 391 397
72. McQuistonJRFieldsPITauxeRVLogsdonJMJr 2008 Do Salmonella carry spare tyres? Trends in microbiology 16 142 148
73. WinterSEThiennimitrPWinterMGButlerBPHusebyDL 2010 Gut inflammation provides a respiratory electron acceptor for Salmonella. Nature 467 426 429
74. HenselM 2000 Salmonella pathogenicity island 2. Mol Microbiol 36 1015 1023
75. EswarappaSMKarnamGNagarajanAGChakrabortySChakravorttyD 2009 lac repressor is an antivirulence factor of Salmonella enterica: its role in the evolution of virulence in Salmonella. PLoS One 4 e5789
76. KauffmannF 1955 [Differential diagnosis and pathogenicity of Salmonella java and Salmonella paratyphi B.]. Z Hyg Infektionskr 141 546 550
77. ChenYTLiaoTLWuKMLauderdaleTLYanJJ 2009 Genomic diversity of citrate fermentation in Klebsiella pneumoniae. BMC Microbiol 9 168
78. GerlachRGCláudioNRohdeMJäckelDWagnerC 2008 Cooperation of Salmonella pathogenicity islands 1 and 4 is required to breach epithelial barriers. Cell Microbiol Nov; 10 2364 2376
79. AhmerBMvan ReeuwijkJWatsonPRWallisTSHF 1999 Salmonella SirA is a global regulator of genes mediating enteropathogenesis. Mol Microbiol 31 971 982
80. De KeersmaeckerSCMarchalKVerhoevenTLEngelenKVanderleydenJ 2005 Microarray analysis and motif detection reveal new targets of the Salmonella enterica serovar Typhimurium HilA regulatory protein, including hilA itself. J Bacteriol 187 4381 4391
81. Main-HesterKLColpittsKMThomasGAFangFCLibbySJ 2008 Coordinate regulation of Salmonella pathogenicity island 1 (SPI1) and SPI4 in Salmonella enterica serovar Typhimurium. Infect Immun 76 1024 1035
82. HarrisSRFeilEJHoldenMTQuailMANickersonEK 2010 Evolution of MRSA during hospital transmission and intercontinental spread. Science 22 469 474
83. CarverTJRutherfordKMBerrimanMRajandreamMABarrellBG 2005 ACT: the Artemis Comparison Tool. Bioinformatics 21 3422 3423
84. BerrimanMRutherfordK 2003 Viewing and annotating sequence data with Artemis. Brief Bioinform 4 124 132
85. StamatakisA 2006 RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22 2688 2690
86. DatsenkoKAWannerBL 2000 One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci U S A 97 6640 6645
87. CherepanovPPWackernagelW 1995 Gene disruption in Escherichia coli: TcR and KmR cassettes with the option of Flp-catalyzed excision of the antibiotic-resistance determinant. Gene 158 9 14
88. MartinezESchroederGNBergerCNLeeSFRobinsonKS 2010 Binding to Na(+)/H(+) exchanger regulatory factor 2 (NHERF2) affects trafficking and function of the enteropathogenic Escherichia coli type III secretion system effectors Map, EspI and NleH. Cell Microbiol 12 1718 1731
89. MiaoEASchererCATsolisRMKingsleyRAAdamsLG 1999 Salmonella typhimurium leucine-rich repeat proteins are targeted to the SPI1 and SPI2 type III secretion systems. Mol Microbiol 34 850 864
90. Kujat ChoySLBoyleECGal-MorOGoodeDLValdezY 2004 SseK1 and SseK2 are novel translocated proteins of Salmonella enterica serovar typhimurium. Infect Immun 72 5115 5125
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2011 Číslo 8
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- Tumor Cell Marker PVRL4 (Nectin 4) Is an Epithelial Cell Receptor for Measles Virus
- Two Group A Streptococcal Peptide Pheromones Act through Opposing Rgg Regulators to Control Biofilm Development
- Differential Contribution of PB1-F2 to the Virulence of Highly Pathogenic H5N1 Influenza A Virus in Mammalian and Avian Species
- Recruitment of the Major Vault Protein by InlK: A Strategy to Avoid Autophagy
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy