
Infections Are Virulent and Inhibit the Human Malaria Parasite in
Endosymbiotic Wolbachia bacteria are potent modulators of pathogen infection and transmission in multiple naturally and artificially infected insect species, including important vectors of human pathogens. Anopheles mosquitoes are naturally uninfected with Wolbachia, and stable artificial infections have not yet succeeded in this genus. Recent techniques have enabled establishment of somatic Wolbachia infections in Anopheles. Here, we characterize somatic infections of two diverse Wolbachia strains (wMelPop and wAlbB) in Anopheles gambiae, the major vector of human malaria. After infection, wMelPop disseminates widely in the mosquito, infecting the fat body, head, sensory organs and other tissues but is notably absent from the midgut and ovaries. Wolbachia initially induces the mosquito immune system, coincident with initial clearing of the infection, but then suppresses expression of immune genes, coincident with Wolbachia replication in the mosquito. Both wMelPop and wAlbB significantly inhibit Plasmodium falciparum oocyst levels in the mosquito midgut. Although not virulent in non-bloodfed mosquitoes, wMelPop exhibits a novel phenotype and is extremely virulent for approximately 12–24 hours post-bloodmeal, after which surviving mosquitoes exhibit similar mortality trajectories to control mosquitoes. The data suggest that if stable transinfections act in a similar manner to somatic infections, Wolbachia could potentially be used as part of a strategy to control the Anopheles mosquitoes that transmit malaria.
Published in the journal:
. PLoS Pathog 7(5): e32767. doi:10.1371/journal.ppat.1002043
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1002043
Summary
Endosymbiotic Wolbachia bacteria are potent modulators of pathogen infection and transmission in multiple naturally and artificially infected insect species, including important vectors of human pathogens. Anopheles mosquitoes are naturally uninfected with Wolbachia, and stable artificial infections have not yet succeeded in this genus. Recent techniques have enabled establishment of somatic Wolbachia infections in Anopheles. Here, we characterize somatic infections of two diverse Wolbachia strains (wMelPop and wAlbB) in Anopheles gambiae, the major vector of human malaria. After infection, wMelPop disseminates widely in the mosquito, infecting the fat body, head, sensory organs and other tissues but is notably absent from the midgut and ovaries. Wolbachia initially induces the mosquito immune system, coincident with initial clearing of the infection, but then suppresses expression of immune genes, coincident with Wolbachia replication in the mosquito. Both wMelPop and wAlbB significantly inhibit Plasmodium falciparum oocyst levels in the mosquito midgut. Although not virulent in non-bloodfed mosquitoes, wMelPop exhibits a novel phenotype and is extremely virulent for approximately 12–24 hours post-bloodmeal, after which surviving mosquitoes exhibit similar mortality trajectories to control mosquitoes. The data suggest that if stable transinfections act in a similar manner to somatic infections, Wolbachia could potentially be used as part of a strategy to control the Anopheles mosquitoes that transmit malaria.
Introduction
Bacterial associates are ubiquitous among insects, including mosquitoes [1]. Wolbachia are obligate endosymbiotic bacteria that infect numerous insects, many of which are vectors of pathogenic microorganisms. Much interest has centered around Wolbachia as a means of reducing arthropod-borne disease due to the capacity of the bacteria to manipulate the reproduction of the insect host, which in turn favors their own transmission [2], [3]. However, recent studies detail that Wolbachia can directly cause pathogen interference (PI) in their invertebrate hosts, whereby infected insects are less susceptible to pathogens [4], [5], [6], [7], [8], [9]. Fitness benefits conferred by PI may partially explain the prevalence of Wolbachia strains that do not confer the more familiarly known reproductive manipulations such as cytoplasmic incompatibility. For example, some Drosophila species infected with specific Wolbachia strains have greater resistance to viral pathogens compared to their uninfected counterparts [4], [9], [10]. From an applied standpoint, mosquito vectors artificially transinfected with Wolbachia exhibit PI against diverse pathogens [5], [6], [8]. The heterologous association between Wolbachia and novel host seems to strongly induce this phenotype in mosquitoes, as the native Wolbachia strain in many vectors does not generally affect pathogen transmission [6], [8]. Wolbachia does cause a small reduction in West Nile virus titer in Culex quinquefasciatus, but this effect is subtle and is unlikely to affect the vector competence of the mosquito [7]. In Aedes aegypti, artificial Wolbachia infections suppress diverse pathogens including RNA viruses, filarial nematodes and the avian malaria parasite Plasmodium gallinaceum [5], [6], [8]. In Anopheles mosquitoes, somatic infection with the Wolbachia strain wMelPop suppresses the rodent malaria parasite P. berghei. These results show that Wolbachia-induced PI may be of use to control various vector-borne diseases [11].
Although the mechanism behind Wolbachia-induced PI is uncertain, several non-mutually exclusive hypotheses have been proposed. In wMelPop and wAlbB-transinfected Ae. aegypti, there is induction of the basal immune state of the host by the novel Wolbachia strain [5], [6], [8]. Activation of the immune state before the mosquito is challenged with pathogens may make the insect less susceptible to infection. Additionally, there is evidence for resource competition between Wolbachia and pathogens such as dengue virus, where virus was only observed in mosquito cells that were not infected with Wolbachia [6].
In addition to PI and manipulation of host reproduction, the wMelPop strain of Wolbachia causes life shortening in both Drosophila and transinfected Aedes aegypti [12], [13]. Due to the extrinsic incubation period (EIP) of many pathogens, life shortening can have a dramatic effect on reducing pathogen transmission. As such, wMelPop has been proposed to control vector-borne diseases by skewing the age structure of the mosquito population toward the younger age classes that are not old enough to transmit pathogens [14], [15]. The dual effect of life shortening and PI can act synergistically, enhancing the prospects for Wolbachia-based disease control strategies [5], [6], [12].
Although naturally uninfected, Anopheles mosquitoes are amenable to Wolbachia infection, both in vitro [16] and in the mosquito somatic tissues [17]. Somatic infection of insects allows for evaluation of Wolbachia phenotypes in the absence of a stably infected host. Recently, somatic infection by wMelPop in An. gambiae was shown to reduce P. berghei levels in conjunction with induction of several innate immune genes. However, immune up-regulation was only investigated at a single time point [11]. It is unknown whether immune induction occurs constantly throughout the life of the mosquito, whether Wolbachia infection will modulate Plasmodium species that are important for human health concerns, or whether different Wolbachia strains will induce similar phenotypes.
To address these issues, we characterized the infection dynamics of two divergent Wolbachia strains (wMelPop and wAlbB) in somatically infected An. gambiae, using fluorescence in situ hybridization (FISH) and qPCR. Host immune gene expression in response to Wolbachia infection was assessed at multiple time points throughout the lifespan of the mosquito. Wolbachia mediated PI was evaluated for the human pathogen P. falciparum. We show that the mosquito immune response to Wolbachia is dynamic, switching between induction and suppression as the mosquito ages. We examined life history traits of mosquitoes infected with the life shortening strain of Wolbachia wMelPop, before and after bloodmeals, and show that strong life shortening was only observed immediately after bloodfeeding. The results are discussed in terms of potential applications for using Wolbachia as part of a strategy for malaria control.
Results/Discussion
Using whole mosquito fluorescence in situ hybridization (FISH), we determined that the Wolbachia strain wMelPop disseminates throughout the mosquito and infects numerous tissues after somatic infection by thoracic microinjection. By 30 days post-infection, Wolbachia is ubiquitous in the abdomen, where it primarily resides within cells of the fat body, and in cells that adhere to the Malpighian tubules, which are most likely hemocytes that have phagocytized Wolbachia. The fat body and hemocytes are major immune tissues within the mosquitoes and infection of these tissues could potentially affect immune processes. Previously it had been demonstrated that Wolbachia could replicate within Anopheles mosquitoes, however the cellular orientation of the infection was unknown [17]. The occurrence of Wolbachia within fat body and hemocyte cells demonstrate conclusively that Wolbachia have the capacity to enter, replicate and survive intracellularly in specific somatic tissues within Anopheles. This observation is supported by in vitro experimentation where Wolbachia has established infections in Anopheles cell culture [16]. Wolbachia are also observed to infect the head of the insect, possibly in the brain or pericerebral fat body. Infection is also observed within the mouthparts and sensory organs of the mosquito (Figure 1) – whether these Wolbachia are free in the hemolymph or contained within circulating hemocytes remains to be determined. The distribution of Wolbachia in somatically infected An. gambiae in part resembles that of the stably infected Aedes aegypti [6], [12]. One noticeable difference between the two mosquito species is the lack of infection in the Anopheles midgut and germline (Figure S1).

Although adult microinjection has successfully been adapted to transinfect multiple insect species [18], [19], [20], no evidence was found for entry of wMelPop into the An. gambiae germline. Previously, adult injection was successfully used to re-infect D. melanogaster with wMel, and to establish infection in Ae. aegypti with wAlbA and wAlbB [18], [20]. Laodelphax striatellus, which naturally harbors wStri, was co-infected with wRi using adult microinjection [19], while wStri has been transferred to Nilaparvata lugens by nymphal injection [21]. In D. melanogaster, Wolbachia was localized to the somatic stem cell niche in the germarium [20], while in both Ae. aegypti and L. striatellus, progeny of microinjected females were infected suggesting entry of Wolbachia into the germline [18], [19]. In contrast, and similar to our results, somatic infection of Bombyx mori was successful after microinjection of Wolbachia into immature life stages, but germline infection was not established [22]. Using FISH, no signal was detected in mature ovaries or immature ovarioles in Anopheles (Figure S1).
The lack of infection of the An. gambiae germline may go some way to explain the unique biology of the Anopheles genera, which is naturally uninfected in nature and seems to be impervious to Wolbachia transinfection despite numerous attempts. There are many possibilities that may explain the lack of infection in the ovary. While Wolbachia can survive intracellularly in Anopheles mosquitoes, the ovarian milieu may be inhospitable to the bacteria. Alternatively, ovarian cell receptors that Wolbachia utilizes may be too divergent in Anopheles, preventing entry into the ovary. Infection itself may cause reproductive ablation. Amhed and Hurd [23] demonstrated that apoptosis in ovarian follicular epithelial cells occurs when the melanization response or humoral antimicrobial activity is induced in An. gambiae. Alternatively, constraints to infection may be related to the bacteria. It is evident that Wolbachia can adapt to new host backgrounds [24], and certain strains of Wolbachia may be more or less suitable for infection establishment. Experiments that address these hypotheses may provide a mechanistic basis for the inability of Wolbachia to infect the Anopheles germline and may provide clues that could ultimately lead to transinfection of this genus.
Quantitative PCR (qPCR) analysis demonstrated that Wolbachia multiples within the mosquito. Since we do not know whether Wolbachia are polyploid, results are presented as Wolbachia genomes per host genome. After microinjection, there is an initial decrease in bacterial density before Wolbachia replicates to increase in abundance (Figure 2). These results are in concordance with Jin et al [17] who used standard PCR to assess somatic infection dynamics of the wMelpop Wolbachia strain. Here, we quantify both wMelpop and wAlbB infection with qPCR and find both these Wolbachia strains display a similar infection pattern, although wAlbB densities are several orders of magnitude lower than wMelPop. This is not unexpected as wMelPop, an over replicating strain, replicates faster than wAlbB in the mosquito (Figure 2) and is initially extracted from cell culture and microinjected into the mosquito at higher densities. It is also possible that the ploidy of wMelPop is higher than wAlbB.

In contrast to Ae. aegypti stably infected with Wolbachia, we see that the immune response in Anopheles after somatic infection is dynamic. At 3 days post infection there is minimal effect on gene expression. Infection by wMelPop and wAlbB moderately suppress Serpin6. wMelPop moderately suppresses cactus, the negative regulator of the Toll pathway, while wAlbB moderately induces Caspar, the negative regulator of the IMD pathway. At 6 days post-infection, Caspar is suppressed by wMelPop in conjunction with up-regulation of Rel2 and cecropin, as well as modestly up-regulating cactus. This time period is coincident with the initial clearing of infection measured by qPCR (Figure 2), and is similar to observations by Kambris and colleagues [11] who observed immune up-regulation (including strong cecropin induction) at a similar time point (8 days post-infection). wAlbB infected mosquitoes display a different profile at this time point, with gene expression not significantly affected. However, at 10 post-infection, the pattern changes to dramatic down-regulation of many immune-related host genes in response to both Wolbachia strains, including FBN9, Heat shock 70, CLIP7A, TEP15 and the transcription factors Rel1 and Rel2 (Figure 3). This time period corresponds with Wolbachia replication in the mosquito (Figure 2), suggesting that Wolbachia may be actively manipulating host gene expression to mediate the infection and replication process. In several instances, suppression of host gene expression by wAlbB is greater compared to wMelPop, suggesting there are strain-specific responses in addition to differences related to bacterial density. This down-regulation is in agreement with regulation patterns observed in vitro, where the Wolbachia strains wAlbB and wRi suppressed many host genes (including genes associated with innate immunity) in cultured An. gambiae Sua5B cells [25]. By 15 days post infection, the response is mixed, with some genes up-regulated and some down-regulated in a Wolbachia strain-specific manner (Figure 3).

After somatic infection, P. falciparum oocyst development was significantly reduced (40–60%) by both wMelPop and wAlbB compared to the Mos55 (Anopheles cell extract) injected control. We observed similar results using both low gametocytemic and high gametocytemic Plasmodium cultures (Figure 4). In the low gametocytemic replicate, infection prevalence (percentage of mosquitoes with one or more oocysts per midgut) was statistically reduced in wMelPop-injected mosquitoes (Mos55 : 75%, N = 65; wMelPop: 33%, N = 21; wAlbB: 60%, N = 45; d.f. = 2, Cramer's V = 0.39, P = 0.002). Infection prevalence did not differ statistically in the high gametocytemic replicates (Mos55 : 90%, N = 50; wMelPop: 83%, N = 35; wAlbB: 84%, N = 55). No correlation was observed between Wolbachia density and Plasmodium oocyst load for either Wolbachia strain (Figure S2), suggesting that the reduction of Plasmodium is not directly related to Wolbachia density (i.e. mosquitoes with high oocyst levels did not necessarily have the lowest Wolbachia titers).

While wMelPop moderately induces the mosquito immune system at 6 days post-infection, by 10 days post-injection, the majority of tested immune genes were down-regulated by both Wolbachia strains (Figure 3). These time points correlate to when Plasmodium is developing within the mosquito midgut. Although, Kambris et al [11] provide evidence that wMelPop-mediated immune up-regulation induces PI in Anopheles against P. berghei, our data suggest that the mosquito immune response to Wolbachia is more dynamic. The modulation of the later immune response suggests mechanisms other than stimulation of basal immunity may be involved in PI in An. gambiae. Alternatively, immune up-regulation around the initial infection period when ookinetes are invading the midgut may be sufficient for a decrease in Plasmodium load. Possibly these different mechanisms may be acting in concert. A more thorough analysis of global immune regulation in response to Wolbachia infection throughout the life of the insect may clarify this issue.
In our Plasmodium experiments, we noted higher mortality of wMelPop-injected mosquitoes compared to wAlbB or cell homogenate-injected treatments. Our previous data suggested that somatic infections of wMelPop were not virulent to Anopheles gambiae [17]. However, in those experiments mosquitoes were not allowed access to blood. We therefore considered the hypothesis that wMelPop-induced virulence in Anopheles gambiae was conditional on bloodfeeding.
Mosquitoes were injected with wMelPop or with uninfected cell culture homogenate as previously described, held for 7 days, then were offered a human bloodmeal with or without P. falciparum parasites through a membrane feeder. After bloodfeeding, fed mosquitoes were separated from unfed mosquitoes and their mortality trajectories assessed. We observed that prior to bloodfeeding, there were no dramatic differences in mortality between infected and uninfected mosquitoes, similar to previous observation. However, wMelPop-infected mosquitoes exhibited a dramatic increase in mortality between 12–24 h post-bloodmeal. After 3 days approximately 80% of the mosquitoes died. After this period, the mortality trajectories of the two treatments become similar again (Figure 5). Infection with Plasmodium made no difference in the mortality phenotypes. Interestingly, we also noted that when comparing Wolbachia levels to Plasmodium oocyst levels, Wolbachia titers were much lower in assayed wMelPop-infected mosquitoes compared to wAlbB mosquitoes (Figure S2), suggesting that mosquitoes with high wMelPop titers did not survive long enough to be assayed for Plasmodium infection. These data show that wMelPop is virulent to An. gambiae, but the virulence phenotype is different than that described for Ae. aegypti and Drosophila [12], [13]. Instead of a general increase in lifetime mortality rates, we observe an acute increase in mortality directly related to bloodmeal acquisition and/or digestion.
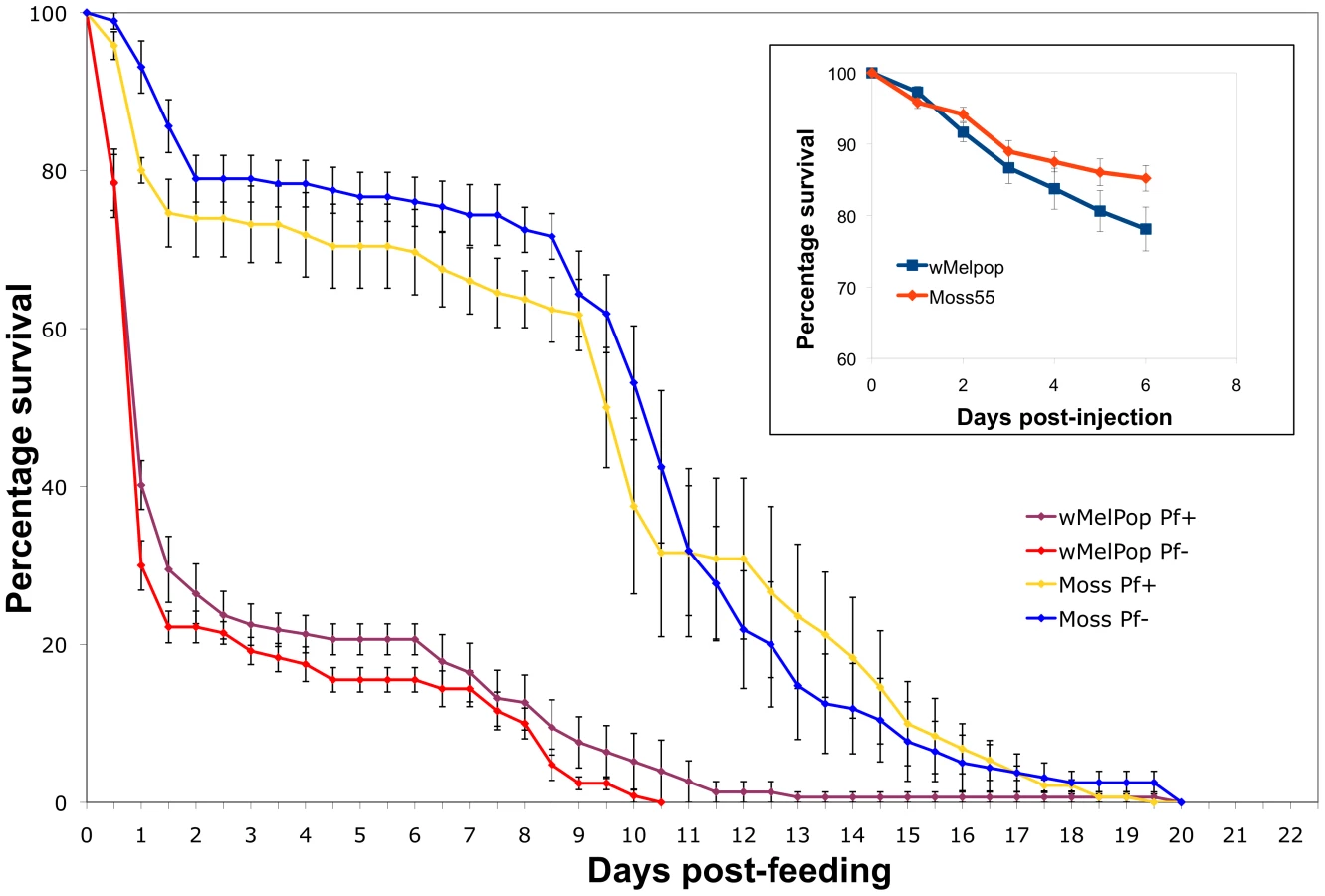
Post bloodmeal, multiple developmental and metabolic processes occur which drastically alter mosquito physiology. Alteration of any of these processes by Wolbachia may potentially induce mortality. In cultured Anopheles Sua5B cells, Wolbachia infection down-regulates host expression of multiple antioxidant genes, including peroxiredoxin, superoxide dismutase and glutathione S transferase [25]. In bloodfed mosquitoes, antioxidant transcripts are up-regulated post bloodmeal [26], [27], [28], [29], [30]. A blood meal also increases iron levels, which are a precursor to reactive oxygen species (ROS). In other systems, Wolbachia has been seen to influence the expression of ferritin and plays a role in iron metabolism [31], [32]. We hypothesize that modulated levels of ROS within the mosquito may be the cause of post bloodmeal mortality. Lending credence to this hypothesis is the observation of increased mortality post-bloodmeal in An. gambiae after silencing of anti-oxidant genes [33]. The more striking mortality observed in this study may be due to down-regulation of numerous genes. Additionally, blood feeding is known to spark a proliferation of bacteria within the insect [1]. In Ae. aegypti, the expansion of gut bacteria post blood meal is attributed to a reduction in ROS, which can result in death of the mosquito [34]. Here, pathogenicity may be directly linked to wMelPop levels or indirectly by Wolbachia influencing the density of other bacteria. Alternatively, the effect of wMelPop on other physiological processes that occur after a blood meal (such as vitellogenesis or nutrient metabolism) may cause fitness costs, as seen in Ae. aegypti where wMelPop affects reproductive output when mosquitoes were fed on non-human hosts [35]. If stable Anopheles infections behave in a similar manner to somatic infections, this acute mortality phenotype could inhibit CI-induced drive of wMelPop into mosquito populations, and provide a selection pressure against the life-shortening phenotype as a large proportion of mosquitoes may die before producing offspring. These potential pitfalls could be offset by the use of this phenotype in a population suppression strategy, or the use of non-virulent Wolbachia strains such as wAlbB.
The use of Wolbachia to control arthropod-borne disease has been postulated for some time. Previous ideas centered on the use of Wolbachia as a gene drive agent, however now it is evident that Wolbachia can also inhibit pathogen development in insects [4], [5], [6], [7], [8], [9]. The obvious limitation to this approach for malaria control is the failure to create a Wolbachia infected Anopheles line, and this still remains a massive challenge in the field of Wolbachia biology. Here we have shown that An. gambiae mosquitoes somatically infected by two strains (wMelPop or wAlbB) are less susceptible to the major human malaria parasite P. falciparum. Using FISH and qPCR, we determined that Wolbachia has ubiquitous distribution in many mosquito tissues and replicates within the Anopheles host. As one oocyst is capable of producing many sporozoites, it would be interesting to determine if sporozoite number is reduced by Wolbachia considering the vast tissue distribution in somatically infected mosquitoes. The results suggest that An. gambiae stably infected with Wolbachia may have reduced ability to maintain transmission of Plasmodium by multiple strain-dependent mechanisms.
Materials and Methods
Ethics statement
Anonymous expired human blood was obtained from a local blood bank for use in mosquito blood feeding experiments.
Wolbachia culture and mosquito infection
Wolbachia was cultured and extracted from infected Anopheles cells as previously described [16], [36]. An. gambiae mosquitoes (Keele strain) were reared as described [16]. Two days post emergence, adult female mosquitoes were anesthetized on ice and injected with Wolbachia according to previously established methodology [17]. Post injection, mosquitoes were incubated at 19°C for 2 days for recovery then maintained at 28°C.
Fluorescence in situ hybridization (FISH)
FISH was performed on wMelPop infected mosquitoes 30 days post injection following the experimental procedure outlined by Koga et al. [37]. Briefly, mosquitoes were fixed in acetone for 3 months, legs were removed and mosquitoes were secondarily fixed in Carnoy's solution. To minimize autofluorescence, mosquitoes were transferred to 10% hydrogen peroxide in 6% alcohol for 5 days. After rehydration in PBST (1–2 hours), tissues were pre-hybridized followed by hybridization with the Wolbachia specific probe overnight [38]. Samples were washed in PBST 3 times to remove excess probe, counterstained with SYTOX green (Invitrogen) and visualized by epifluorescent and confocal microscopy. Individual channel images are available as Supplementary data (Figure S3). FISH controls included 1) no probe controls, 2) competition controls in which unlabeled oligonucleotides were added to the hybridization buffer to suppress the fluorescent signals and 3) RNase digestion controls, in which prior to hybridization RNAs in the insect materials were removed by RNase A treatment (Figure S4).
Quantitative PCR (qPCR) for Wolbachia density and host gene expression
DNA or RNA was extracted from somatically infected mosquitoes using DNAzol (Molecular Research Center, Inc., Cincinnati, OH) or RNeasy mini kits (Qiagen) for estimation of Wolbachia density and quantification of host gene expression respectively. qPCR to determine the density of wMelPop in whole mosquitoes was completed by amplifying the single copy gene WD_0550 [24], while wAlbB was amplified with modified GF and BR primers which specifically bind to the wsp gene [18]. Ten mosquitoes were assay at each time point for each strain to estimate Wolbachia density, while 5 mosquitoes were used for host gene expression per time point. The relative abundance of each Wolbachia strain was determined after normalization to the mosquito single-copy S7 gene [39]. For host gene expression, RNA was DNase treated (Ambion) and cDNA synthesized using superscript III (Invitrogen) following manufactures guidelines. qPCR was completed using a Rotor gene Q (Qiagen) using the Rotor gene SYBR green PCR kit (Qiagen) according to manufactures guidelines. qPCRs were completed in triplicate. PCR primers are listed in Table S1. Melt curve analysis was completed on all PCRs. In Wolbachia density experiments, data were analyzed by Kruskal-Wallis test using the Connover-Inman method for pairwise contrasts between time points. For host gene expression experiments, significance was assessed by Mann-Whitney U test compared to mosquitoes injected with uninfected Mos55 cell culture homogenate (control). Tested mosquito genes were identified in a microarray screen of Wolbachia-regulated Anopheles genes in cultured cells [25]. Additional analyses were conducted using REST [40] and qGENE [41] software.
Plasmodium falciparum mosquito infections
2-day old female mosquitoes were intrathoracically injected with wMelPop or wAlbB (purified from cell culture) as described [17] or with uninfected Mos55 cell culture homogenate (control). Seven days post-injection, mosquitoes were offered a Plasmodium-infected blood meal. Prior to blood feeding, mosquitoes were starved overnight. The gametocytemia of infected blood meals was approximately 0.3% and 1% for low and high titer infections, respectively. After blood feeding, unfed mosquitoes were removed. P. falciparum NF-54 gametocyte cultures were washed and mosquitoes were fed infected blood warmed to 37°C through a membrane feeder [42]. Post feeding, unfed mosquitoes were removed and blood-fed An. gambiae were incubated at 24°C for 7 days. Midguts of mosquitoes were dissected, stained with 0.2% mercurochrome and oocysts enumerated using a light contrast microscope (Olympus). The Wolbachia density of each mosquito carcass was determined by qPCR as described above. The experiment was replicated 3 times. Replicate one was a high-gametocytemic culture, while replicates two and three had low gametocytemia. The variances of the data for replicates two and three did not differ statistically and were pooled for analysis (squared ranks test, P>0.05) while replicate one was analyzed separately. Data were analyzed by Kruskal-Wallis test using the Dwass method for pairwise comparisons.
wMelPop mortality experiments
An. gambiae female adults were injected with wMelPop or uninfected Mos55 cell culture homogenate (control) and fed a P. falciparum gametocyte infected or uninfected blood meal as previously described. Unfed mosquitoes were separated from fed mosquitoes. Mosquitoes were reared at 24°C at a density of approximately 30 mosquitoes per cup (4 cups per treatment) and monitored twice daily for survival. Dead mosquitoes were removed from the experiment every 12 hours. The entire experiment was repeated twice. Data were analyzed by Kaplan-Meier analysis. Statistical significance was assessed by Kruskal-Wallis test using the Dwass method for pairwise comparisons.
Supporting Information
Zdroje
1. DongYManfrediniFDimopoulosG 2009 Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog 5 e1000423
2. WerrenJHBaldoLClarkME 2008 Wolbachia: master manipulators of invertebrate biology. Nat Rev Microbiol 6 741 751
3. StouthamerRBreeuwerJAHurstGD 1999 Wolbachia pipientis: microbial manipulator of arthropod reproduction. Annu Rev Microbiol 53 71 102
4. HedgesLMBrownlieJCO'NeillSLJohnsonKN 2008 Wolbachia and virus protection in insects. Science 322 702 702
5. KambrisZCookPEPhucHKSinkinsSP 2009 Immune activation by life-shortening Wolbachia and reduced filarial competence in mosquitoes. Science 326 134 136
6. MoreiraLAIturbe-OrmaetxeIJefferyJALuGPykeAT 2009 A Wolbachia symbiont in Aedes aegypti limits infection with Dengue, Chikungunya, and Plasmodium. Cell 139 1268 1278
7. GlaserRLMeolaMA 2010 The native Wolbachia endosymbionts of Drosophila melanogaster and Culex quinquefasciatus increase host resistance to West Nile virus infection. PLoS ONE 5 e11977
8. BianGXuYLuPXieYXiZ 2010 The endosymbiotic bacterium Wolbachia induces resistance to Dengue virus in Aedes aegypti. PLos Pathog 6 e1000833
9. TeixeiraLFerreiraAAshburnerM 2008 The bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster. PLoS Biol 6 e1000002
10. OsborneSELeongYSO'NeillSLJohnsonKN 2009 Variation in antiviral protection mediated by different Wolbachia strains in Drosophila simulans. PLoS Pathog 5 e1000656
11. KambrisZBlagboroughAMPintoSBBlagroveMSCGodfrayHCJ 2010 Wolbachia stimulates immune gene expression and inhibits Plasmodium development in Anopheles gambiae. PLoS Pathog 6 e1001143
12. McMenimanCJLaneAMCassBNFongAWCSidhuM 2009 Stable introduction of a life-shortening Wolbachia infection into the mosquito Aedes aegypti. Science 323 141 144
13. MinKTBenzerS 1997 Wolbachia, normally a symbiont of Drosophila, can be virulent, causing degeneration and early death. Proc Natl Acad Sci USA 94 10792 10796
14. CookPMcMenimanCJO'NeillSL 2008 Modifying insect population age structure to control vector-borne disease. Adv Exp Med Biol 627 126 140
15. SinkinsSPO'NeillSL 2000 Wolbachia as a vehicle to modify insect populations. HandlerAFJamesAA Insect Transgenesis: Methods and Applications New York CRC Press 271 287
16. RasgonJLRenXPetridisM 2006 Can Anopheles gambiae be infected with Wolbachia pipientis? Insights from an in vitro system. Appl Environ Microbiol 72 7718 7722
17. JinCRenXRasgonJL 2009 The virulent Wolbachia strain wMelPop efficiently establishes somatic infections in the malaria vector Anopheles gambiae. Appl Environ Microbiol 75 3373 3376
18. Ruang-AreerateTKittayapongP 2006 Wolbachia transinfection in Aedes aegypti: A potential gene driver of dengue vectors. Proc Natl Acad Sci USA 103 12534 12539
19. KangLMaXCaiLLiaoSSunL 2003 Superinfection of Laodelphax striatellus with Wolbachia from Drosophila simulans. Heredity 90 71 76
20. FrydmanHMLiJMRobsonDNWieschausE 2006 Somatic stem cell niche tropism in Wolbachia. Nature 441 509 512
21. KawaiSMatsumotoYGotohTNodaH 2009 Transinfection of Wolbachia in planthoppers: nymphal injection of cultured Wolbachia and infection dynamics. Environ Entomol 38 1626 1633
22. KageyamaDNaritaSNodaH 2008 Transfection of feminizing Wolbachia endosymbionts of the butterfly, Eurema hecabe, into the cell culture and various immature stages of the silkmoth, Bombyx mori. Microb Ecol 56 733 741
23. AhmedAMHurdH 2006 Immune stimulation and malaria infection impose reproductive costs in Anopheles gambiae via follicular apoptosis. Microbes Infect 8 308 315
24. McMenimanCJLaneAMFongAWCVoroninDAIturbe-OrmaetxeI 2008 Host adaptation of a Wolbachia strain after long-term serial passage in mosquito cell lines. Appl Environ Microbiol 74 6963 6969
25. HughesGLRenXRamirezJLSakamotoJMBaileyJA 2011 Wolbachia infections in Anopheles gambiae cells: transcriptomic characterization of a novel host-symbiont interaction. PLoS Pathog 7 e1001296
26. SandersHREvansAMRossLSGillSS 2003 Blood meal induces global changes in midgut gene expression in the disease vector, Aedes aegypti. Insect Biochem Mol Biol 33 1105 1122
27. RibeiroJM 2003 A catalogue of Anopheles gambiae transcripts significantly more or less expressed following a blood meal. Insect Biochem Mol Biol 33 865 882
28. MarinottiONguyenQKCalvoEJamesAARibeiroJM 2005 Microarray analysis of genes showing variable expression following a blood meal in Anopheles gambiae. Insect Mol Biol 14 365 373
29. HoltRA 2002 The genome sequence of the malaria mosquito Anopheles gambiae. Science 298 129 149
30. DanaANHongYSKernMKHillenmeyerMEHarkerBW 2005 Gene expression patterns associated with blood-feeding in the malaria mosquito Anopheles gambiae. BMC Genomics 6 5
31. BrownlieJCCassBNRieglerMWitsenburgJJIturbe-OrmaetxeI 2009 Evidence for metabolic provisioning by a common invertebrate endosymbiont, Wolbachia pipientis, during periods of nutritional stress. PLoS Pathog 5 e1000368
32. KremerNVoroninDACharifDMavinguiPMollereauB 2009 Wolbachia interferes with ferritin expression and iron metabolism in insects. PLoS Pathog 5 e1000630
33. MagalhaesTBrackneyDEBeierJCFoyBD 2008 Silencing an Anopheles gambiae catalase and sulfhydryl oxidase increases mosquito mortality after a blood meal. Arch Insect Biochem Physiol 68 134 143
34. OliveiraJHMGonçalvesRLSLaraFADiasFAGandaraACP 2011 Blood Meal-Derived Heme Decreases ROS Levels in the Midgut of Aedes aegypti and Allows Proliferation of Intestinal Microbiota. PLoS Pathog 7 e1001320
35. McMenimanCJHughesGLO'NeillSL 2011 A Wolbachia symbiont in Aedes aegypti disrupts mosquito egg development to a greater extent when mosquitoes feed on nonhuman versus human blood. J Med Entomol 48 76 84
36. RasgonJLGamstonCRenXX 2006 Survival of Wolbachia pipientis in cell-free medium. Appl Environ Microbiol 72 6934 6937
37. KonoMKogaRShimadaMFukatsuT 2008 Infection dynamics of coexisting beta - and gammaproteobacteria in the nested endosymbiotic system of mealybugs. Appl Environ Microbiol 74 4175 4184
38. HeddiAGrenierAMKhatchadourianCCharlesHNardonP 1999 Four intracellular genomes direct weevil biology: nuclear, mitochondrial, principal endosymbiont, and Wolbachia. Proc Natl Acad Sci USA 96 6814 6819
39. DasSRadtkeAChoiY-JMendesAMValenzuelaJG 2010 Transcriptomic and functional analysis of the Anopheles gambiae salivary gland in relation to blood feeding. BMC Genomics 11 566
40. PfafflMWHorganGWDempfleL 2002 Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res 30 e36
41. JoehanesRNelsonJC 2008 QGene 4.0, an extensible Java QTL-analysis platform. Bioinformatics 24 2788 2789
42. CarterRRanford-CartwrightLAlanoP 1993 The culture and preparation of gametocytes of Plasmodium falciparum for immunochemical, molecular, and mosquito infectivity studies. Methods Mol Biol 21 67 88
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2011 Číslo 5
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- Crystal Structure and Functional Analysis of the SARS-Coronavirus RNA Cap 2′-O-Methyltransferase nsp10/nsp16 Complex
- Lymphoadenopathy during Lyme Borreliosis Is Caused by Spirochete Migration-Induced Specific B Cell Activation
- The OXI1 Kinase Pathway Mediates -Induced Growth Promotion in Arabidopsis
- : An Emerging Cause of Sexually Transmitted Disease in Women
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy