
Toll-8/Tollo Negatively Regulates Antimicrobial Response in the Respiratory Epithelium
Barrier epithelia that are persistently exposed to microbes have evolved potent immune tools to eliminate such pathogens. If mechanisms that control Drosophila systemic responses are well-characterized, the epithelial immune responses remain poorly understood. Here, we performed a genetic dissection of the cascades activated during the immune response of the Drosophila airway epithelium i.e. trachea. We present evidence that bacteria induced-antimicrobial peptide (AMP) production in the trachea is controlled by two signalling cascades. AMP gene transcription is activated by the inducible IMD pathway that acts non-cell autonomously in trachea. This IMD-dependent AMP activation is antagonized by a constitutively active signalling module involving the receptor Toll-8/Tollo, the ligand Spätzle2/DNT1 and Ect-4, the Drosophila ortholog of the human Sterile alpha and HEAT/ARMadillo motif (SARM). Our data show that, in addition to Toll-1 whose function is essential during the systemic immune response, Drosophila relies on another Toll family member to control the immune response in the respiratory epithelium.
Published in the journal:
. PLoS Pathog 7(10): e32767. doi:10.1371/journal.ppat.1002319
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1002319
Summary
Barrier epithelia that are persistently exposed to microbes have evolved potent immune tools to eliminate such pathogens. If mechanisms that control Drosophila systemic responses are well-characterized, the epithelial immune responses remain poorly understood. Here, we performed a genetic dissection of the cascades activated during the immune response of the Drosophila airway epithelium i.e. trachea. We present evidence that bacteria induced-antimicrobial peptide (AMP) production in the trachea is controlled by two signalling cascades. AMP gene transcription is activated by the inducible IMD pathway that acts non-cell autonomously in trachea. This IMD-dependent AMP activation is antagonized by a constitutively active signalling module involving the receptor Toll-8/Tollo, the ligand Spätzle2/DNT1 and Ect-4, the Drosophila ortholog of the human Sterile alpha and HEAT/ARMadillo motif (SARM). Our data show that, in addition to Toll-1 whose function is essential during the systemic immune response, Drosophila relies on another Toll family member to control the immune response in the respiratory epithelium.
Introduction
Although the innate immune system is a primitive host defense mechanism, it involves a sophisticated repertoire of humoral and cellular responses both acting systemically and locally [1]. In recent years, the Drosophila model organism has proven to be an invaluable system in dissecting in great details the genetics and cellular mechanisms regulating the innate immunity [2]–[3]. One fundamental mechanism common to humans and Drosophila immunity involves signaling by receptors of the Toll family. Upon microbial infection, human TLRs activate the synthesis of cytokines and other regulatory molecules that stimulate the adaptive immune system [4]. In Drosophila, Toll signalling leads to the activation of the systemic immune response, which is characterized by the synthesis of AMPs by the fat body cells [5]. Upon secretion into the circulating blood, these AMPs provide systemic protection against fungi and bacteria. In the mammalian innate immune response, bacteria are directly sensed by TLRs [6]. In contrast, these microorganisms are detected in Drosophila by another class of proteins, the Peptidoglycan Recognition Proteins (PGRPs), also present in the human proteome [7]–[9]. Recognition of Lys-type peptidoglycan (PGN) (mainly found in Gram-positive bacteria cell wall) by the circulating PGRP-SA protein triggers a protease cascade involving successively Mod-Sp, Grass, and the Spätzle-Processing-Enzyme (SPE) [8], [10]–[11]. Upon activation, SPE becomes competent to transform the zymogen pro-Spätzle into an active ligand for the Toll receptor, inducing its dimerization and intracellular signalling [12]. Production of AMPs after infection by Gram-negative bacteria, is however largely independent of the Toll pathway but rather relies on another NF-κB signalling cascade named IMD [13]. Sensing of Gram-negative bacteria upstream of the IMD pathway takes place at the plasma membrane, through PGN recognition by the transmembrane PGRP-LC receptor. Binding of DAP-type PGN (present in Gram-negative bacterial cell wall) to PGRP-LC induces its dimerization, which, in turn, triggers IMD-dependent intracellular events enabling the nuclear translocation of the NF-κB transcription factor Relish [14]–[17].
Although penetration of infectious microbes into the body cavity, and consequently, activation of a systemic immune response, is a rare event, interactions between microbes and epithelia take place constantly throughout the life of all metazoan. This implies that these barrier epithelia must be armed with efficient systems for microbial detection and elimination. However, these epithelia that act as interfaces with the external environment, share some characteristics that could be seen as detrimental for the needs of an effective immune system. Indeed, they usually have large surface areas and consist of thin structural layers, thus representing ideal entry points for pathogens. In this respect, the airway epithelium is unique among all epithelia, since it has a very delicate structure and is constantly exposed to a plethora of airborne pathogens. This could explain the occurrence of a great variety of inflammatory lung diseases, including asthma, chronic obstructive pulmonary disease or cystic fibrosis [18]–[19]. Elucidation of the primary steps that lead to chronic inflammation of the mammalian lung is obstructed by the complexity of inflammatory responses in this organ. Animals with a much simpler organization, such as the fruit fly, might help us clarify the basic architecture of this epithelial immune response, thereby helping to unravel the mechanisms that lead to chronic inflammation of the airways. Although of much simpler organization, the fly's airway system shows striking similarities with the mammalian lung regarding both its architecture and its physiology [20]–[25].
In this report, we present a detailed description of the mechanisms that regulate AMP production in the Drosophila respiratory epithelium. We show that, in contrast to systemic fat body immune response, the IMD pathway can be activated non-cell autonomously in the tracheal network. We present evidence that IMD pathway activation is tightly regulated in the cells of the respiratory epithelium. We demonstrate that the molecular mechanisms underlying IMD down-regulation following infection, are different from those previously reported in the gut and in the fat body, and rely on a dialog between two antagonist pathways. The production of AMPs in the trachea is positively regulated by the IMD pathway, which is counterbalanced by a negative regulation from a signalling cassette, whose upstream receptor is a member of the Toll family, Toll-8/Tollo. Our data suggest that the Spz2/DNT1 cytokine is a putative Tollo ligand in this process, and that Ect4, the Drosophila ortholog of the human Toll/Interleukin-1 Receptor (TIR) domain-containing protein SARM mediates Tollo signalling during tracheal immune response.
Results
AMP activation followed a stereotypical pattern in the Drosophila tracheal network
The tracheal system is a relatively simple model system that has provided an important insight into the biology of branching morphology [22]. It is a tubular structure covered by a lumenal cuticular lining that forms a physical barrier against dehydration and invading microorganisms [20]. This network consists of a monolayer epithelium made up of two dorsal trunks (DT) connected to several visceral branches (VB) bringing oxygen to internal tissues. Upon infection by the entomopathogenic bacterium Erwinia carotovora carotovora (Ecc), these epithelial cells produce a cocktail of AMPs including Drosomycin, Drosocin and Attacin [26]–[27]. In order to perform a detailed spatiotemporal analysis of this epithelial response, we used a Drosomycin-GFP reporter transgene (Drs-GFP) and monitored the response of this tissue after infection. When reared on conventional medium, a few larvae showed sporadic tracheal Drs-GFP expression mainly in VB, and rarely in the posterior part of the DT, namely the posterior spiracles (PS) (Figure 1A and 1B). This basal Drs-GFP tracheal expression was qualitatively and quantitatively similar in larvae reared conventionally or under axenic conditions (data not shown). Upon infection with Ecc, Drs-GFP activation followed a somewhat stereotypical pattern. Responding larvae were categorized into three classes according to their Drs-GFP expression pattern (Figure 1A and 1C), namely, larvae expressing GFP in PS and in posterior VB only (class I), in PS and all VB (class II) and in VB and DT (class III). Kinetic experiments showed that GFP signal was first detected in PS and then spread into VB (Figure 1A, C). The reporter was only later activated in the DT and, unexpectedly, first in the anterior half and then in the entire trunk (Figure 1A). Although we appreciate that the expression patterns of reporter transgenes can slightly deviate from those of the actual AMP, the Drs-GFP expression patterns observed cannot easily be attributed to a progressive diffusion of bacterial elicitors (such as PGN) from the spiracles into the tracheal network, but rather speak for more complicated mechanisms in tracheal AMP activation.

IMD pathway activation is not strictly cell autonomous in the trachea
Next, we compared the mode of activation of AMP following forced immune pathway activation in the trachea and in another immune tissue, the fat body. Although activation of Drosomycin transcription is mainly controlled by the Toll pathway in the fat body (but can be activated by ectopic IMD pathway triggering, see later), it is strictly IMD-dependent in the trachea [5], [26], [28]. Indeed, over-activation of the IMD (UAS-PGRP-LCa, UAS-IMD), but not of the Toll pathway (UAS-spz act, Tl3), is sufficient to induce tracheal expression of Drs-GFP in non-infected larvae (Figure S1A and S1B). Concomitantly, loss-of-function mutations in IMD pathway components (Relish, PGRP-LC, IMD) prevent Drs-GFP tracheal activation in infected larvae, whereas Toll signalling mutants such as spz or Dif do show a wild-type tracheal response upon infection. To analyze whether all tracheal cells were competent to trigger AMP production upon IMD pathway activation, we induced UAS-IMD expressing clones in tracheal cells, using fat body clones as controls. Overexpression of IMD led to a strictly cell-autonomous and fully penetrant activation of both Drs-GFP and Dipt-Cherry in the fat body (Figure 2B and 2C). In the trachea, although most IMD-expressing cells showed Drs-GFP expression, a fraction did not (Figure 2A). In addition, Drs-GFP activation was not always associated with the expression of the UAS-IMD transgene (Figure 2A), suggesting that IMD pathway activation in trachea is not strictly cell autonomous. These results were confirmed by using a UAS-PGRP-LCa transgene that activated Drs-GFP both autonomously and non-autonomously in trachea cells (Figure 2E) but strictly cell-autonomously in the fat body (Figure 2F). We next addressed whether PGRP-LC function was required cell-autonomously for IMD pathway activation in the trachea upon infection. Analysis of MARCM loss-of-function clones for PGRP-LC indicates that tracheal cells mutant for PGRP-LC were totally impaired in their ability to trigger Drs-GFP expression, following Ecc infection (Figure 2D and H). These results indicate that, although PGRP-LC is essential in tracheal cells for IMD pathway triggering, IMD pathway activation in one tracheal cell can spread to neighboring cells. This contrasts with the strictly cell-autonomous IMD-dependent immune response observed in fat body cells.

Tollo is expressed apically in the Drosophila trachea cells
In order to get a further insight into the mechanisms that control AMP induction in trachea, we looked for putative immune genes expressed in this tissue. A recent report identified the repertoire of all immune genes expressed in the trachea [29]. One of the striking data of this study, [confirmed by FlyAtlas (http://flyatlas.org/)] was that, in addition to Toll itself, two other Toll family members, 18-Wheeler (Toll-2) and Tollo (Toll-8), are strongly expressed in trachea. 18-wheeler being implicated in developmental processes with indirect impacts the immune response [30], we focused our study on the putative function of the Tollo transmembrane protein in the tracheal immune response. Using Lac-Z reporter lines (data not shown) and q-RT-PCR (Figure 3A), we confirmed that Tollo mRNA is highly enriched in the tracheal epithelium, and expressed at lower levels in other tissues. To investigate the subcellular localization of the Tollo protein, we genetically associated a UAS-Tollo::Myc construct with the trachea-specific Breathless-Gal4 driver (Btl-Gal4). Anti-Myc antibody staining suggested that Tollo was localized apically at the cell membrane facing the airway lumen (Figure 3B). Double staining experiments showed that Tollo::Myc partially co-localized with the apical marker Cadherin::GFP, but was mutually exclusive with Viking::GFP, a basal membrane-associated protein (Figure 3B). These results indicate that Tollo is a protein enriched in the tracheal epithelium with an apical subcellular localization.

AMPs are specifically over-produced in infected Tollo mutant trachea
The rather restricted expression pattern of Tollo mRNA in the trachea and the apical subcellular distribution of Tollo protein prompted us to investigate its putative function in the immune response. For that purpose, we used two previously characterized hypomorphic alleles (Tollo145 and TolloR5A) together with a complete loss-of-function allele (TolloC5) that we generated by P-element mediated homologous recombination [31]–[32] (Figure S2A). All Tollo mutants were viable with no obvious developmental defects and gave rise to phenotypically normal pharate adults indicating that Tollo has no essential role in Drosophila development. We tested the ability of these Tollo mutant larvae to mount an immune response. In the absence of infection, approximately 5% of wild-type larvae showed Drs-GFP expression in VB and/or PS (Figure S3). Similar Figures were obtained with Tollo mutants suggesting that Tollo is not required to set up the basal level of AMP production in the absence of infection (Figure S3). After bacterial infection, however, the immune response was much stronger in Tollo mutants than in wild-type sibling larvae (Figure 4A and 4B and Figure S3). While we could identify the three previously described classes of Drs-GFP positive larvae in both control and Tollo mutants, the relative proportion of these was significantly different between genotypes (Figure S3). The percentage of larvae showing no GFP expression was reduced to 5–10% in Tollo mutant (compared to 40% in controls), whereas Class III larvae, which represented 15% of controls, reached up to 50% in the Tollo mutant larvae (Figure S3). Similar results were obtained with three independent Tollo alleles and in trans-heterozygous allelic combination demonstrating that this phenotype was, indeed, due to Tollo inactivation and not to other mutations on the chromosome (data not shown). The effects were not only qualitative but also quantitative. In most infected Tollo mutant larvae, Drs-GFP expression was intense, whereas it was rarely the case in controls (Figure 4A). q-RT-PCR experiments indicate that Drosomycin, Drosocin and Attacin mRNA levels were respectively increased by 6, 7 and 2.6 fold after infection in Tollo mutants compared to wild-type trachea (Figure 4B). To ensure that this effect was indeed a consequence of Tollo inactivation in the tracheal network itself, we combined the Btl-Gal4 driver with a UAS-TolloIR RNA interference construct. As shown in Figure 4A and Figure S3, larvae, in which Tollo was eliminated specifically in trachea, also showed increased Drs-GFP expression in this tissue after infection.

In addition to regulating Drosomycin expression in the trachea, the IMD pathway also controls Diptericin transcription locally in the gut and systemically in the fat body [13], [26]. To test whether Tollo acts as a general negative regulator of IMD-dependent mechanisms in other immune tissues, we analyzed the effects of inactivating Tollo on IMD pathway activation in the gut and the fat body. Using the Dipt-Cherry reporter construct for the larval stage (Figure 4D) and q-RT-PCR for both larval and adult stages (Figure 4E and 4F), we showed that Tollo was not implicated in IMD negative regulation in either tissue. This was the case for both immune responses induced by septic injury or by oral ingestion. In addition, we showed that Tollo mutants were unaffected in their ability to activate the Toll pathway during a Gram-positive bacteria-mediated systemic immune response (Figure 4F). Altogether these results demonstrate that the Tollo receptor is specifically required to dampen IMD pathway-dependent responses in the tracheal network after infection.
AMP over-production in Tollo mutants is not secondary to trachea defects
Since AMPs are induced upon cellular stress, we tested whether Drosomycin expression in Tollo mutants was a secondary consequence of a possible implication of Tollo in tracheal formation. The following reasons led us to believe that it was not the case. 1) Tollo mutant embryos gave rise to viable adults, suggesting that Tollo mutant trachea are fully functional in larvae and adults. 2) Tracheal cell morphology of Tollo and control larvae appeared similar when observed under transmission electron microscopy (Figure 5A). 3) No constitutive AMP transcription was detected in non-infected Tollo mutant larvae (Figure 4B and Figure S3). We then wondered whether Drosomycin over-activation could be linked to the presence of higher levels of potential immune elicitors in Tollo mutant and RNAi trachea. This could be due to the presence of higher bacterial load in the trachea. However, as shown in Figure 5B and Figure S4B, the number of Ecc-GFP in Tollo mutant trachea was identical in Tollo mutants and in controls. Alternatively, Tollo mutant trachea could be more permeable to contaminated external fluid. To test this hypothesis, wild type and Tollo mutant larvae were incubated in the presence of a fluorescent dye, bromophenol blue. External fluid penetration inside the trachea lumen was not different in wild-type and Tollo mutant larvae (Figure 5C and 5D). This indicates that over-activation of Drosomycin in both Tollo mutant and RNAi trachea cannot be attributed to an increase in fluid penetration, and therefore putative immune elicitor load within the tracheal lumen. Altogether, these results demonstrate that the infection-dependent Drosomycin over-activation observed in Tollo mutant, is not secondary to defective trachea but rather suggests a direct implication of the Tollo protein in the regulation of IMD-dependent Drosomycin expression.

Ect4/SARM and DNT1/Spätzle 2 loss-of-function mutants phenocopy Tollo mutants
We then tried to identify the intra - and extra-cellular components that may mediate Tollo signalling in trachea. The Drosophila Toll-1 receptor and vertebrate TLR functions have all been shown to be mediated by TIR domain-containing proteins, respectively, the Drosophila dMyd88 and the mammalian Myd88, TRIF, SARM, TRAM and MAL [33]–[35]. However, the Drosophila proteome contains two TIR domain proteins, Ect4/SARM the Drosophila ortholog of vertebrate SARM and dMyd88, the latter mediating the Toll signalling during dorso-ventral axis specification and immune response [36]–[38]. In order to test whether Tollo acts through a TIR domain-containing protein, we analyzed the tracheal immune response of dMyd88 and Ect4/SARM mutants. We showed that larvae carrying Ect4/SARM mutations (Figure S2B) display a strong over-activation of Drosomycin expression (visualized with the Drs-GFP reporter transgene and quantified by q-RT-PCR) upon infection, phenotype that was not observed in dMyd88 mutant larvae (Figure 6A and 6B and Figure S5A). This suggests that Ect4/SARM is the bone fide TIR domain adaptor transducing Tollo signalling in the tracheal immune response. Whereas TLRs function as Pattern Recognition Receptors by directly binding to microbial motifs, previous work has shown than the Drosophila Toll-1 receptor is activated during both embryonic dorso-ventral axis specification and immune response by its ligand Spätzle [39]–[40]. Since spz mutant larvae did not present higher activation of Drs-GFP in the respiratory tract after infection, we believe that Spz is not a functional ligand for Tollo (Figure 6C). Since Spz-like genes are present in the fly genome, we screened them for a Tollo-like phenotype. We observed that removing Spz2 (known as DNT1) function in trachea phenocopies Tollo mutant as far as Drosomycin over-activation is concerned (Figure 6C and 6D and Figure S5B). This suggests that DNT1 could be the, or one of the ligand(s), responsible for Tollo activity in the tracheal immune response. Similarly to Tollo, q-RT-PCR data indicate that Ect4 and DNT1 are specifically acting in the tracheal epithelium and do not contribute to IMD pathway regulation in the gut and in the fat body (Figure 6B and 6D).
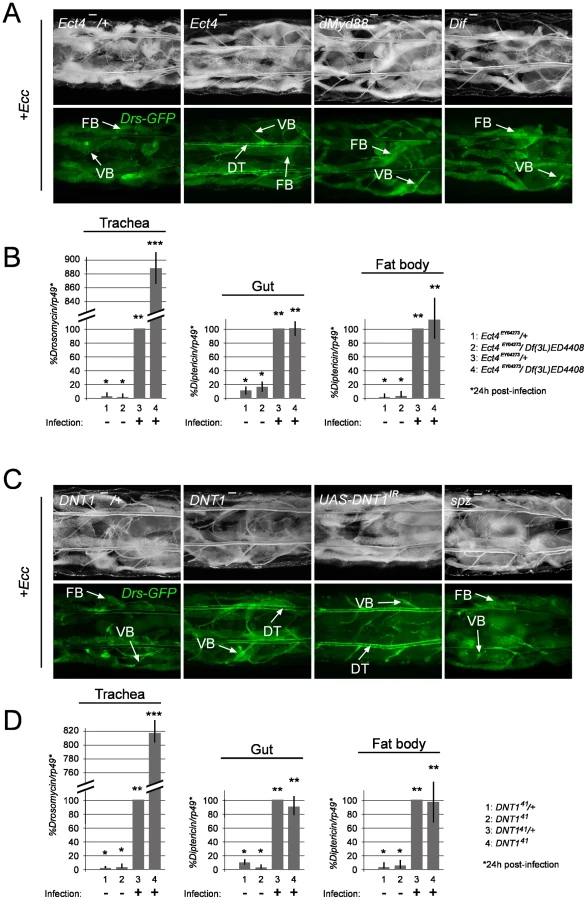
Tollo is hypostatic to IMD in the trachea
Taking into account the above results, it appears that the function of Tollo is specifically to down-regulate the IMD pathway in the tracheal cells following infection. In order to genetically place Tollo with respect to known IMD pathway components, we performed epistatic experiments. We showed that the Tollo mutant phenotype requires functional PGRP-LC receptor and intracytoplasmic adaptor IMD, since double mutant Tollo-, PGRP-LC- and imd -; Tollo- trachea did not show any signs of Drs-GFP activation after infection (Figure 4A). This epistatic relationship was confirmed by q-RT-PCR on Drosomycin mRNA (Figure 4C). In genetic terms, Tollo is hypostatic, or acts in parallel to PGRP-LC and imd. Consistently, Relish nuclear translocation monitored with an anti-Relish antibody was higher in Tollo RNAi-infected tracheal cells than in controls (Figure S6). These results suggest that Tollo is not directly involved in IMD pathway activation per se but that, in its absence, IMD pathway activation is more efficient upon infection.
Discussion
Epithelial responses are first and foremost local responses to prevent the epithelium from unnecessary immune reactions. Since the recognition steps in Drosophila respiratory epithelia involve the transmembrane receptor PGRP-LC and occur within the extracellular space, it is expected that molecular mechanisms must be at work to prevent constitutive or excessive immune response in this tissue, particularly essential for animal growth and viability. In this report, we present data demonstrating that the transmembrane receptor Tollo is part of a signalling network, whose function is to specifically down-regulate AMP production in the trachea. We show that Tollo antagonizes IMD pathway activation in the respiratory epithelium, and that DNT1/Spz2 and Ect4/SARM are putative Tollo ligand and transducer, respectively, in this process. Our data demonstrate that, in addition to the family founder Toll-1, another member of the Leucine-Rich-Repeats family of Toll proteins, is regulating the Drosophila innate immune response. Although it has been abundantly documented that every single mammalian TLR has an immune function [4], the putative implication of Toll family members, other than Toll-1 itself, in the Drosophila immune response has been a subject of controversy [41]. Data showing that Drosophila Toll-9 over-expression was sufficient to induce AMPs expression in vivo has prompted the idea that Toll-9 could maintain significant levels of anti-microbial molecules, thus providing basal protection against microbes [42]. However, our recent analysis of a complete Toll-9 loss-of-function allele has shown that this receptor is neither implicated in basal anti-microbial response nor required to mount an immune response to bacterial infection [43]. The present data are also fully consistent with a recent report showing that Toll-6, Toll-7 and Toll-8 are not implicated in systemic AMP production in flies [44], and demonstrate that a Toll family member, Tollo, is a negative regulator of local airway epithelial immune response upon bacterial infection. In contrast to Toll-1, whose activation is inducible in the fat body, Tollo pathway activation seems to be constitutive in the trachea. Despite these differences, both receptors use a member of the Spz family as ligand. Interestingly, sequence similarities, intron's size and conservation of key structural residues, indicate that Spz2/DNT1 is phylogenetically the closest family member to the Toll ligand Spz [45]. Furthermore, both Spz and Spz2/DNT1 have been shown to have neurotrophic functions in flies [46]. It would be of great interest to test whether Tollo also mediates Spz2 function in the nervous system.
Both during embryonic development and immune response, Spz is activated by proteolytic cleavage [10], [47]-[48]. This step depends upon the Easter protease that is implicated in D/V axis specification and on SPE for Toll pathway activation by microbes. Since Spz orthologs are also produced as longer precursors, they are likely to be activated by proteolysis. The fact that Tollo and Spz2 loss-of-function phenotypes correspond to excessive AMP production, suggests that in wild-type conditions, the Tollo pathway is constitutively activated by an active form of the Spz2 ligand. This situation is reminiscent to that observed in the embryonic ventral follicle cells, in which a Pipe-mediated signal induces a constitutive activation of the Easter cascade leading to Spz cleavage, Toll activation and, in turn, ventral fate acquisition [49]. It should be noted that Easter and one Pipe isoform are very strongly expressed in the trachea cells (Flyatlas), and are candidate proteins in mediating Tollo activity in the respiratory epithelia.
The fact that Ect4, but not dMyd88 mutant, loss-of-function mutant phenocopies Tollo mutant suggest that Ect4 could be the TIR domain adaptor transducing Tollo signal in the tracheal cells. Alternatively, Ect4/SARM could mediate Tollo function by interfering with IMD pathway signalling. In mammals, SARM is under the transcriptional control of TLR and negatively regulates TLR3 signalling by directly interfering with the association between the RHIM domain-containing proteins TRIF and RIP [50]. Since PGRP-LC contains a RHIM domain as TRIF, and IMD is the Drosophila counterpart of RIP, one can envisage that Drosophila SARM could act by interfering with the PGRP-LC/IMD association required for IMD pathway signalling. Similarly to its function as a negative regulator in fly immunity, SARM is the only TIR domain-containing adaptor that acts as a suppressor of TLR signalling [36], [50].
One obvious question relates to the mode of action of Tollo on IMD pathway downregulation. Two mechanisms have been recently described that result in the down-regulation of the IMD pathway. The first one regulates PGRP-LC membrane localization, and is dependent on the PIRK protein [51]–[53]. Upon infection, the intracellular PIRK protein is up-regulated and, in turn, represses PGRP-LC plasma membrane localization leading to the shutdown of the IMD signalling [53]. In infected pirk mutants, IMD-dependent AMPs are overproduced in both the gut and the fat body. In our conditions, however, inactivation of PIRK specifically in the trachea did not influence Drosomycin activation in trachea (Figure S7A). To verify whether Tollo is acting via a mechanism similar to PIRK, we looked at PGRP-LC membrane localization using a UAS-PGRP-LC::GFP construct. PGRP-LC membrane localization was identical in wild-type and Tollo mutant tracheal cells (Figure S7B). The second mechanism that modulates IMD activation, acts directly on the promoters of IMD target genes. Ha et al. (2005) have shown that the Caudal transcription factor sits on some of the IMD target promoters preventing their activation by Relish [54]. We thus tested the putative implication of Caudal in Tollo signalling by using Drs-GFP reporter transgenes containing either wild-type Caudal Responsive Elements (CDREs) or mutated versions unresponsive to Caudal activity [55]. Upon infection, Drs-GFP with mutated CDREs was activated in fat body but not in gut or trachea (Figure S7C). In conclusion, Caudal acts as a transcriptional activator, rather than a repressor, for the Drs-GFP reporter in trachea. These results indicate that Tollo does not regulate the IMD pathway via PGRP-LC membrane localization or through promoter targeting of Caudal. One challenging task for the future will be to identify the mechanism used by Tollo to counter-balance tracheal PGRP-LC activation. It has been reported that the loss of Tollo function in ectodermal cells during embryogenesis alters glycosylation in nearby differentiating neurons [31], [56]–[57]. Since the pattern of oligosaccharides expressed in a cell can influence its interactions with others and with pathogens, Tollo could function by modifying glycosylation pattern in response to microbes. It could be envisaged that Tollo mediates PGRP-LC glycosylation, and thereby reduces its ability to respond to bacterial elicitors. Further work will be required to address the above hypothesis, whereby Tollo activity and glycosylation modification could be linked in order to regulate the IMD pathway activation in trachea.
Material and Methods
Bacterial strains
The following microorganisms were used: Erwinia carotovora carotovora 15 2141 (Ecc), Erwinia carotovora carotovora 15 pOM1-GFP spectinomycinR (Ecc-GFP), Escherichia coli 1106 (E.coli) and Micrococcus luteus CIPA270 (M. luteus).
Bacterial load analysis
Bacterial load of surface sterilized individuals was quantified by plating appropriate serial dilutions of lysates obtained from 6 dissected guts or trachea (from larvae) on nutrient agar plates (Luria Bertani + spectinomycin 100 µg/ml). Biological triplicates were collected for each experimental condition at 4h and 24h after Ecc-GFP infection. Homogenization of tissues was performed using the Precellys 24 tissue homogenizer (Bertin technologies, France) and 0,75-1mm glass beads in 500 µL of LB + spectinomycin.
Drosophila melanogaster strains and maintenance
PGRP-LCDE12 is a complete deletion of the PGRP-LC locus [16]. Flies carrying this mutation are unable to activate the IMD pathway. spzrm7 is a null allele which prevents Toll pathway activation [5]. yw, Drs-GFP [27], Dpt-Cherry [58], TolloC5 (this work), TolloR5A [32], Tollo145 [59], UAS-TolloIR (VDRC #9431), UAS-Tollo::Myc [31], DNT141 [46], UAS-spz2IR (VDRC #26115), Ect4EY04273 BL#15733, Df(3L)ED4408 BL#8065, Tl3 BL#3238 (a dominant gain-of-function allele of Tl, Btl-Gal4 BL#8807, UAS-myrRFP BL#7118, act>CD2>Gal4 BL#4780, cad-EGFP BL#30875, Vkg-GFP (a gift from Michel Sémériva), hs-Gal4 BL#2077, RelishE20 [60], imd1 [5], UAS-spz act [30], dMyd88c03881 [34], UAS-PGRP-LC::GFP (a gift from François Leulier) and Dif1 [61]. Generation of the TolloC5 allele was performed as described in [62] using the two following inserted elements: d01565 and PBacf05248 [63]. Complete deletion of the Tollo gene was confirmed by sequencing genomic DNA extracted from TolloC5 mutants (molecular details upon request). Fly stocks were raised on standard cornmeal-agar medium at 25°C.
Natural infection of larvae and adults
Cells from overnight bacterial cultures were recovered by centrifugation at 4,000 g for 10 min at 4°C. The supernatant was discarded and the pellet was resuspended in fresh LB media. Cell suspensions were serially diluted in PBS, and the concentration of cells was determined by optical-density (OD) measurement. 200 µl of an overnight bacterial culture of Ecc (OD = 200) were directly added on top of feeding third instar larvae into a standard cornmeal-agar medium at 25°C. A similar volume of LB broth was used in control experiments. Larvae were monitored for Drosomycin and Diptericin transcription by fluorescence analysis using Drs-GFP and Dpt-cherry reporters respectively, and by qRT-PCR, 24h after infection. Septic injuries were performed by pricking adult males with a thin needle contaminated with M. luteus or E. coli. 200 µl of Bromophenol Blue (SIGMA # B8026) at 10 g/l were directly added on top of feeding third instar larvae.
Flip-out clones and MARCM
For Drs-GFP study, Drs-GFP;UAS-myrRFP;act>CD2>Gal4 females were crossed to either ywhsflp;;UAS-IMD or to ywhsflp;; UAS-PGRP-LCa males. For Dpt-Cherry study, ywhsflp; UAS-GFP; act>CD2>Gal4 females were crossed to Dpt-Cherry; UAS-IMD or to Dpt-Cherry, UAS-PGRP-LCa males. In both cases, larvae of the progeny were heat shocked at early-mid L3 stage (72h-96h after egg deposition, AED) and observed 24 h later. Generation of MARCM clones in trachea was performed by crossing MARCM virgin females of genotype ywhsflp;; Tub-Gal80 FRT2A en masse to the Drs-GFP; Blt-Gal4, UAS-myrRFP; PGRP-LCDE12 FRT2A line. Resulting embryos were submitted to a heat shock 4–6 hr AED for 1 hr at 38°C in a circulating water bath, and kept at 25°C until larvae reached early-mid third instar (72h-96h AED), when they were infected by Ecc and observed 24 h later.
Immunostaining on larvae
Larval tissue were dissected in PBS and fixed for 20 min in 4% paraformaldehyde on ice. After several rinses in PBT (PBS + 0.1% Triton X-100), they were blocked for 1 hr in PBT-3% BSA at 4°C and then incubated with antibody at the appropriate dilution in PBT-BSA 3% overnight at 4°C. Primary antibodies were: rabbit Anti-Relish (1∶500) or Mouse Anti-Myc (9E10 Santa Cruz at 1∶ 500). Several washes in PBT were followed by a 2 hr incubation with secondary antibody at RT (Alexa Fluor 546 goat anti-rabbit IgG and Alexa Fluor 555 goat anti-mouse IgG diluted 1∶500, Molecular Probes), then 5 washes in PBT. The tissues were finally mounted in Vectashield (Vector Laboratories) fluorescent mounting medium, with DAPI. Images were captured with a LSM 510 Zeiss confocal microscope.
Quantitative real-time PCR
Quantitative real-time PCR and SYBR Green analysis were performed as previously described [58]. Primer information can be obtained upon request. The amount of mRNA detected was normalized to control rp49 mRNA values. Normalized data was used to quantify the relative levels of a given mRNA according to cycling threshold analysis (ΔCt).
Electronic microscopy
For electron microscopic sections, third instar larvae trachea were dissected and fixed at RT in 4% PFA and 2% glutaraldehyde in 0.12 M sodium cacodylate buffer at pH 7.4 for 1 h. The trachea were then washed 3×10 min in 0.12 M sodium cacodylate buffer, post-fixed in 2% OsO4 in 0.12 M sodium cacodylate buffer for 1 h and washed again 3×10 min. Samples were subsequently dehydrated through series of ethanol gradients and infiltrated with propylene oxide, embedded in epoxy resin (Fluka, Sigma) and polymerized at 80°C. Ultrathin (80 nm) plastic sections were cut using a Leica UltraCut microtome with a diamond Diatome knife and post-stained with 2% uranyl acetate, followed by treatment with Reynolds'lead citrate, and stabilized for transmission EM by carbon coating. Examination was performed with a Zeiss Leo 912 microscope at 100 kV. Images were captured using a Gatan 792 Bioscan camera using Digital Micrograph software.
Supporting Information
{kind=link}
Zdroje
1. MedzhitovR 2007 Recognition of microorganisms and activation of the immune response. Nature 449 819 826
2. FerrandonDImlerJLHetruCHoffmannJA 2007 The Drosophila systemic immune response: sensing and signalling during bacterial and fungal infections. Nat Rev Immunol 7 862 874
3. LemaitreBHoffmannJ 2007 The host defense of Drosophila melanogaster. Annu Rev Immunol 25 697 743
4. AkiraSTakedaK 2004 Toll-like receptor signalling. Nat Rev Immunol 4 499 511
5. LemaitreBNicolasEMichautLReichhartJMHoffmannJA 1996 The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 86 973 983
6. KumarHKawaiTAkiraS 2009 Toll-like receptors and innate immunity. Biochem Biophys Res Commun 388 621 625
7. RoyetJDziarskiR 2007 Peptidoglycan recognition proteins: pleiotropic sensors and effectors of antimicrobial defences. Nat Rev Microbiol 5 264 277
8. MichelTReichhartJMHoffmannJARoyetJ 2001 Drosophila Toll is activated by Gram-positive bacteria through a circulating peptidoglycan recognition protein. Nature 414 756 759
9. LiuCXuZGuptaDDziarskiR 2001 Peptidoglycan recognition proteins: a novel family of four human innate immunity pattern recognition molecules. J Biol Chem 276 34686 34694
10. BuchonNPoidevinMKwonHMGuillouASottasV 2009 A single modular serine protease integrates signals from pattern-recognition receptors upstream of the Drosophila Toll pathway. Proc Natl Acad Sci U S A 106 12442 12447
11. LeulierFParquetCPili-FlourySRyuJHCaroffM 2003 The Drosophila immune system detects bacteria through specific peptidoglycan recognition. Nat Immunol 4 478 484
12. WeberANTauszig-DelamasureSHoffmannJALelievreEGascanH 2003 Binding of the Drosophila cytokine Spatzle to Toll is direct and establishes signaling. Nat Immunol 4 794 800
13. LemaitreBKromer-MetzgerEMichautLNicolasEMeisterM 1995 A recessive mutation, immune deficiency (imd), defines two distinct control pathways in the Drosophila host defense. Proc Natl Acad Sci U S A 92 9465 9469
14. ChoeKMLeeHAndersonKV 2005 Drosophila peptidoglycan recognition protein LC (PGRP-LC) acts as a signal-transducing innate immune receptor. Proc Natl Acad Sci U S A 102 1122 1126
15. RametMManfruelliPPearsonAMathey-PrevotBEzekowitzRA 2002 Functional genomic analysis of phagocytosis and identification of a Drosophila receptor for E. coli. Nature 416 644 648
16. GottarMGobertVMichelTBelvinMDuykG 2002 The Drosophila immune response against Gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature 416 640 644
17. Erturk-HasdemirDBroemerMLeulierFLaneWSPaquetteN 2009 Two roles for the Drosophila IKK complex in the activation of Relish and the induction of antimicrobial peptide genes. Proc Natl Acad Sci U S A 106 9779 9784
18. VercelliD 2008 Discovering susceptibility genes for asthma and allergy. Nat Rev Immunol 8 169 182
19. BarnesPJ 2008 Immunology of asthma and chronic obstructive pulmonary disease. Nat Rev Immunol 8 183 192
20. AndrewDJEwaldAJ 2010 Morphogenesis of epithelial tubes: Insights into tube formation, elongation, and elaboration. Dev Biol 341 34 55
21. GhabrialALuschnigSMetzsteinMMKrasnowMA 2003 Branching morphogenesis of the Drosophila tracheal system. Annu Rev Cell Dev Biol 19 623 647
22. LubarskyBKrasnowMA 2003 Tube morphogenesis: making and shaping biological tubes. Cell 112 19 28
23. TangHKambrisZLemaitreBHashimotoC 2008 A serpin that regulates immune melanization in the respiratory system of Drosophila. Dev Cell 15 617 626
24. WagnerCIsermannKRoederT 2009 Infection induces a survival program and local remodeling in the airway epithelium of the fly. FASEB J 23 2045 2054
25. RoederTIsermannKKabeschM 2009 Drosophila in asthma research. Am J Respir Crit Care Med 179 979 983
26. TzouPOhresserSFerrandonDCapovillaMReichhartJM 2000 Tissue-specific inducible expression of antimicrobial peptide genes in Drosophila surface epithelia. Immunity 13 737 748
27. FerrandonDJungACCriquiMLemaitreBUttenweiler-JosephS 1998 A drosomycin-GFP reporter transgene reveals a local immune response in Drosophila that is not dependent on the Toll pathway. EMBO J 17 1217 1227
28. BassetAKhushRSBraunAGardanLBoccardF 2000 The phytopathogenic bacteria Erwinia carotovora infects Drosophila and activates an immune response. Proc Natl Acad Sci U S A 97 3376 3381
29. WagnerCIsermannKFehrenbachHRoederT 2008 Molecular architecture of the fruit fly's airway epithelial immune system. BMC Genomics 9 446
30. LigoxygakisPBuletPReichhartJM 2002 Critical evaluation of the role of the Toll-like receptor 18-Wheeler in the host defense of Drosophila. EMBO Rep 3 666 673
31. SeppoAMataniPSharrowMTiemeyerM 2003 Induction of neuron-specific glycosylation by Tollo/Toll-8, a Drosophila Toll-like receptor expressed in non-neural cells. Development 130 1439 1448
32. AyyarSPistilloDCallejaMBrookfieldAGittinsK 2007 NF-kappaB/Rel-mediated regulation of the neural fate in Drosophila. PLoS One 2 e1178
33. O'NeillLABowieAG 2007 The family of five: TIR-domain-containing adaptors in Toll-like receptor signalling. Nat Rev Immunol 7 353 364
34. KambrisZBilakHD'AlessandroRBelvinMImlerJL 2003 DmMyD88 controls dorsoventral patterning of the Drosophila embryo. EMBO Rep 4 64 69
35. Tauszig-DelamasureSBilakHCapovillaMHoffmannJAImlerJL 2002 Drosophila MyD88 is required for the response to fungal and Gram-positive bacterial infections. Nat Immunol 3 91 97
36. YuanSWuKYangMXuLHuangL 2010 Amphioxus SARM involved in neural development may function as a suppressor of TLR signaling. J Immunol 184 6874 6881
37. BelindaLWWeiWXHanhBTLeiLXBowH 2008 SARM: a novel Toll-like receptor adaptor, is functionally conserved from arthropod to human. Mol Immunol 45 1732 1742
38. CouillaultCPujolNReboulJSabatierLGuichouJF 2004 TLR-independent control of innate immunity in Caenorhabditis elegans by the TIR domain adaptor protein TIR-1, an ortholog of human SARM. Nat Immunol 5 488 494
39. WeberANGangloffMMoncrieffeMCHyvertYImlerJL 2007 Role of the Spatzle Pro-domain in the generation of an active toll receptor ligand. J Biol Chem 282 13522 13531
40. GangloffMMuraliAXiongJArnotCJWeberAN 2008 Structural insight into the mechanism of activation of the Toll receptor by the dimeric ligand Spatzle. J Biol Chem 283 14629 14635
41. TauszigSJouanguyEHoffmannJAImlerJL 2000 Toll-related receptors and the control of antimicrobial peptide expression in Drosophila. Proc Natl Acad Sci U S A 97 10520 10525
42. OoiJYYagiYHuXIpYT 2002 The Drosophila Toll-9 activates a constitutive antimicrobial defense. EMBO Rep 3 82 87
43. Narbonne-ReveauKCharrouxBRoyetJ 2011 Lack of an antibacterial response defect in Drosophila Toll-9 mutant. PLoS One 6 e17470
44. YagiYNishidaYIpYT 2010 Functional analysis of Toll-related genes in Drosophila. Dev Growth Differ 52 771 783
45. ParkerJSMizuguchiKGayNJ 2001 A family of proteins related to Spatzle, the toll receptor ligand, are encoded in the Drosophila genome. Proteins 45 71 80
46. ZhuBPennackJAMcQuiltonPForeroMGMizuguchiK 2008 Drosophila neurotrophins reveal a common mechanism for nervous system formation. PLoS Biol 6 e284
47. SteinD 1995 Pattern formation: The link between ovary and embryo. Curr Biol 5 1360 1363
48. SchneiderDSJinYMorisatoDAndersonKV 1994 A processed form of the Spatzle protein defines dorsal-ventral polarity in the Drosophila embryo. Development 120 1243 1250
49. ChoYSStevensLMSteinD 2010 Pipe-dependent ventral processing of Easter by Snake is the defining step in Drosophila embryo DV axis formation. Curr Biol 20 1133 1137
50. CartyMGoodbodyRSchroderMStackJMoynaghPN 2006 The human adaptor SARM negatively regulates adaptor protein TRIF-dependent Toll-like receptor signaling. Nat Immunol 7 1074 1081
51. KleinoAMyllymakiHKallioJVanha-ahoLMOksanenK 2008 Pirk is a negative regulator of the Drosophila Imd pathway. J Immunol 180 5413 5422
52. AggarwalKRusFVriesema-MagnusonCErturk-HasdemirDPaquetteN 2008 Rudra interrupts receptor signaling complexes to negatively regulate the IMD pathway. PLoS Pathog 4 e1000120
53. LhocineNRibeiroPSBuchonNWepfAWilsonR 2008 PIMS modulates immune tolerance by negatively regulating Drosophila innate immune signaling. Cell Host Microbe 4 147 158
54. HaEMOhCTBaeYSLeeWJ 2005 A direct role for dual oxidase in Drosophila gut immunity. Science 310 847 850
55. RyuJHNamKBOhCTNamHJKimSH 2004 The homeobox gene Caudal regulates constitutive local expression of antimicrobial peptide genes in Drosophila epithelia. Mol Cell Biol 24 172 185
56. BaasSSharrowMKotuVMiddletonMNguyenK 2010 Sugar-free frosting, a homolog of SAD kinase, drives neural-specific glycan expression in the Drosophila embryo. Development 138 553 563
57. AokiKPerlmanMLimJMCantuRWellsL 2007 Dynamic developmental elaboration of N-linked glycan complexity in the Drosophila melanogaster embryo. J Biol Chem 282 9127 9142
58. CharrouxBRoyetJ 2009 Elimination of plasmatocytes by targeted apoptosis reveals their role in multiple aspects of the Drosophila immune response. Proc Natl Acad Sci U S A 106 9797 9802
59. KimSChungSYoonJChoiKWYimJ 2006 Ectopic expression of Tollo/Toll-8 antagonizes Dpp signaling and induces cell sorting in the Drosophila wing. Genesis 44 541 549
60. HedengrenMAslingBDushayMSAndoIEkengrenS 1999 Relish, a central factor in the control of humoral but not cellular immunity in Drosophila. Mol Cell 4 827 837
61. RutschmannSKilincAFerrandonD 2002 Cutting edge: the toll pathway is required for resistance to gram-positive bacterial infections in Drosophila. J Immunol 168 1542 1546
62. ParksALCookKRBelvinMDompeNAFawcettR 2004 Systematic generation of high-resolution deletion coverage of the Drosophila melanogaster genome. Nat Genet 36 288 292
63. AbdelsadikARoederT 2010 Chronic activation of the epithelial immune system of the fruit fly's salivary glands has a negative effect on organismal growth and induces a peculiar set of target genes. BMC Genomics 11 265
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2011 Číslo 10
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- Severe Acute Respiratory Syndrome Coronavirus Envelope Protein Regulates Cell Stress Response and Apoptosis
- The SARS-Coronavirus-Host Interactome: Identification of Cyclophilins as Target for Pan-Coronavirus Inhibitors
- Biochemical and Structural Insights into the Mechanisms of SARS Coronavirus RNA Ribose 2′-O-Methylation by nsp16/nsp10 Protein Complex
- Evolutionarily Divergent, Unstable Filamentous Actin Is Essential for Gliding Motility in Apicomplexan Parasites
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy