
Origin and Evolution of Sulfadoxine Resistant
The Thailand-Cambodia border is the epicenter for drug-resistant falciparum malaria. Previous studies have shown that chloroquine (CQ) and pyrimethamine resistance originated in this region and eventually spread to other Asian countries and Africa. However, there is a dearth in understanding the origin and evolution of dhps alleles associated with sulfadoxine resistance. The present study was designed to reveal the origin(s) of sulfadoxine resistance in Cambodia and its evolutionary relationship to African and South American dhps alleles. We sequenced 234 Cambodian Plasmodium falciparum isolates for the dhps codons S436A/F, A437G, K540E, A581G and A613S/T implicated in sulfadoxine resistance. We also genotyped 10 microsatellite loci around dhps to determine the genetic backgrounds of various alleles and compared them with the backgrounds of alleles prevalent in Africa and South America. In addition to previously known highly-resistant triple mutant dhps alleles SGEGA and AGEAA (codons 436, 437, 540, 581, 613 are sequentially indicated), a large proportion of the isolates (19.3%) contained a 540N mutation in association with 437G/581G yielding a previously unreported triple mutant allele, SGNGA. Microsatellite data strongly suggest the strength of selection was greater on triple mutant dhps alleles followed by the double and single mutants. We provide evidence for at least three independent origins for the double mutants, one each for the SGKGA, AGKAA and SGEAA alleles. Our data suggest that the triple mutant allele SGEGA and the novel allele SGNGA have common origin on the SGKGA background, whereas the AGEAA triple mutant was derived from AGKAA on multiple, albeit limited, genetic backgrounds. The SGEAA did not share haplotypes with any of the triple mutants. Comparative analysis of the microsatellite haplotypes flanking dhps alleles from Cambodia, Kenya, Cameroon and Venezuela revealed an independent origin of sulfadoxine resistant alleles in each of these regions.
Published in the journal:
. PLoS Pathog 6(3): e32767. doi:10.1371/journal.ppat.1000830
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1000830
Summary
The Thailand-Cambodia border is the epicenter for drug-resistant falciparum malaria. Previous studies have shown that chloroquine (CQ) and pyrimethamine resistance originated in this region and eventually spread to other Asian countries and Africa. However, there is a dearth in understanding the origin and evolution of dhps alleles associated with sulfadoxine resistance. The present study was designed to reveal the origin(s) of sulfadoxine resistance in Cambodia and its evolutionary relationship to African and South American dhps alleles. We sequenced 234 Cambodian Plasmodium falciparum isolates for the dhps codons S436A/F, A437G, K540E, A581G and A613S/T implicated in sulfadoxine resistance. We also genotyped 10 microsatellite loci around dhps to determine the genetic backgrounds of various alleles and compared them with the backgrounds of alleles prevalent in Africa and South America. In addition to previously known highly-resistant triple mutant dhps alleles SGEGA and AGEAA (codons 436, 437, 540, 581, 613 are sequentially indicated), a large proportion of the isolates (19.3%) contained a 540N mutation in association with 437G/581G yielding a previously unreported triple mutant allele, SGNGA. Microsatellite data strongly suggest the strength of selection was greater on triple mutant dhps alleles followed by the double and single mutants. We provide evidence for at least three independent origins for the double mutants, one each for the SGKGA, AGKAA and SGEAA alleles. Our data suggest that the triple mutant allele SGEGA and the novel allele SGNGA have common origin on the SGKGA background, whereas the AGEAA triple mutant was derived from AGKAA on multiple, albeit limited, genetic backgrounds. The SGEAA did not share haplotypes with any of the triple mutants. Comparative analysis of the microsatellite haplotypes flanking dhps alleles from Cambodia, Kenya, Cameroon and Venezuela revealed an independent origin of sulfadoxine resistant alleles in each of these regions.
Introduction
The Thailand-Cambodia border in Southeast Asia has been an epicenter for drug resistant falciparum malaria where chloroquine (CQ) resistance emerged in the early 1960s [1], followed by sulfadoxine-pyrimethamine (SP) resistance in the late 1970s [2],[3] and mefloquine (MQ) resistance in the mid 1990s [4]. Following the emergence of MQ resistance, artemisinin-based combination therapy (ACT) consisting of artesunate (AS) and MQ was adopted as the first-line treatment against uncomplicated falciparum malaria in Thailand (1995) and Cambodia (2000) [4],[5].
Sulfadoxine-pyrimethamine (SP) combination is the most widely used antifolate to treat CQ-resistant falciparum malaria and, because of their synergistic effect, the combination is more effective than either drug used alone [6],[7],[8]. It is also one of the partner drugs of artemisinin-based combination therapy (ACT) currently being used for treatment of uncomplicated falciparum malaria in some parts of the world [9]. Importantly, SP is the only drug recommended by the World Health Organization (WHO) for intermittent preventive treatment (IPT) in pregnant women in Sub-Saharan Africa where large number of deaths occur due to malaria in pregnancy [9]. Sulfadoxine acts by inhibiting dihydropteroate synthase (dhps), an essential enzyme of the folate biosynthesis pathway [10]. Previous studies have identified mutations at five DHPS codons (S436A/F; A437G; K540E; A581G; and A613S/T) to be associated with sulfadoxine resistance in P. falciparum [11],[12],[13],[14],[15]. Kinetic studies have shown that the mutant DHPS enzyme has a reduced affinity for sulfadoxine, resulting in the development of resistance, the level of which broadly correlates with the number of mutations in dhps [13].
In Southeast Asia, at least two different triple mutant dhps alleles (AGEAA and SGEGA) have been described [16],[17],[18] whereas in South America, a single triple mutant allele SGEGA has been reported [19],[20],[21],[22],[23]. Interestingly in Africa, the SGEAA (east Africa) and AGKAA (west and central Africa) double mutants are the common dhps alleles [24],[25], with no triple mutant dhps alleles reported on this continent thus far. Similarly, the pyrimethamine-resistant quadruple mutant dhfr allele (51I/59R/108N/164L) is abundant in Southeast Asia and in low frequency in Africa [26],[27]. The triple mutant dhfr allele 51I/59R/108N is widespread in Southeast Asia and Africa, whereas in South America (mostly in the Peruvian Amazon) two other forms of the triple mutant (50R/51I/108N and 51I/108N/164L) are prevalent.
Microsatellite analysis has revealed that triple (51I/59R/108N) mutant dhfr alleles in Africa have shared ancestry with dhfr alleles from Southeast Asia suggesting that, like CQ resistance, the alleles conferring pyrimethamine resistance were also introduced into Africa from Southeast Asia [25],[26],[27],[28],[29],[30],[31],[32],[33]. However, unlike Southeast Asia where all dhfr alleles have single common ancestor [26], additional local evolution of the dhfr alleles has been reported in Africa [25],[27],[28],[34],[35]. In South America, the pyrimethamine-resistant dhfr alleles are thought to have two independent origins [20],[21],[22]. Recently, another independent origin of the double mutant dhfr allele (59R/108N) has been observed in Papua New Guinea [36]. Thus, similar to CQ resistance, at least four major distinct origins of pyrimethamine resistance have been reported worldwide.
Although the origins of CQ and pyrimethamine resistance are better understood, there is limited data related to the origins of sulfadoxine resistance globally. Previous studies have reported multiple and independent origins of double mutant dhps alleles in Africa [24],[25],[37], and one common origin for the triple mutant dhps allele in South America [20],[21],[22]. Importantly, the origins of the three different double mutants (AGKAA, SGKGA and SGEAA) and two different triple mutant (AGEAA and SGEGA) dhps alleles remain unknown in Southeast Asia. Therefore, the present study was aimed to (i) determine whether resistant dhps alleles in Cambodia are experiencing a selective sweep (ii) estimate the probable number of origins of the highly resistant dhps alleles in Cambodia and (iii) examine the evolutionary relationships between Southeast Asian (Cambodia), South American (Venezuela) and African (Cameroon and Kenya) dhps alleles.
Materials and Methods
Sample collection, DNA isolation and genotyping for dhps codons 436 to 613
Plasmodium falciparum clinical isolates were collected from patients with uncomplicated falciparum malaria from five sites in Cambodia: Pailin and Kampong Seila in the west, Chumkiri in the south, and Memut and Rattanakiri in the east (Fig. 1). Patients enrolled in this study were treated with ACT (Artesunate+Mefloquine) according to the national drug policy of Cambodia. Written informed consent was obtained from each patient before blood collection. The study was approved by Institutional Review Boards (IRBs) of the Cambodia National Ethics Committee for Health Research, the US Naval Medical Research Unit No. 2 (NAMRU-2) Jakarta, Indonesia, and the University of North Carolina at Chapel Hill (UNC), USA.

DNA was extracted from filter paper blood spots using QIAmp Mini kit (Qiagen, Valencia, CA, USA). A total of 234 P. falciparum isolates (Pailin, n = 51; Kampong Seila, n = 10; Chumkiri, n = 85; Memut, n = 43; and Rattanakiri, n = 45) were sequenced for a portion of the dhps gene covering codons 436, 437, 540, 581 and 613. Each DNA sample representing a clinical isolate was subjected to two rounds of PCR. The primary amplification was done using 5′-AACCTAAACGTGCTGTTCAA-3′ (Forward) and 5′-AATTGTGTGATTTGTCCACAA-3′ (Reverse) primers with the following cycling parameters: 5 minutes initial denaturation at 95°C followed by 35 cycles with 30 seconds denaturation at 95°C, 30 seconds annealing at 50°C, 1 minute extension at 68°C and a final 5 minute extension at 68°C. The primary amplification product was subjected to nested PCR using 5′-ATGATAAATGAAGGTGCTAG-3′ (Forward) and 5′-TCATTTTGTTGTTCATCATGT-3′ (Reverse) primers with the same cycling parameters as primary except that the annealing was done at 52°C and the number of cycles was reduced to 30. The 647 bp nested product was sequenced on an ABI 3130xl Genetic Analyzer (Applied Biosystems, Foster City, CA, USA).
Microsatellite analysis
Parasite isolates with mixed dhps sequencing electropherograms (multiple peaks at one or more nucleotide) were removed from the microsatellite analysis. Only those with single dhps genotypes were assayed for 8 neutral microsatellite loci on chromosomes 2 (GenBank UniSTS ID: C2M27, C2M29, C2M34, C2M33) and 3 (GenBank UniSTS ID: C3M40, C3M88, C3M69 and C3M39) in order to exclude any additional multiply-infected samples and to obtain an estimate of the neutral baseline heterozygosity in the population [38]. These loci have previously been used for constructing a genetic map of P. falciparum and are not known to be under the influence of any selection [38]. PCR cycling parameters for all 8 neutral loci were adapted from Nair et al [26] as described earlier [21]. Isolates containing multiple alleles at one or more loci on chromosomes 2 and/or 3 were not carried forward for analyzing microsatellites on chromosome 8 around the dhps gene. Singly-infected isolates were typed for 10 microsatellite loci, five upstream (−11 Kb, −7.5 Kb, −2.9 Kb, −1.5 Kb, −0.13 Kb) and five downstream (0.03 Kb, 0.5 Kb, 1.4 Kb, 6.4 Kb and 9 Kb) of the dhps gene. Primer sequences and PCR cycling parameters for these loci are provided as supporting information available online (Table S1). The amplified PCR products were separated on ABI 3130xl Genetic Analyzer and analyzed using GeneMapper software v3.7 (Applied Biosystems, Foster City, CA, USA).
Testing selection and genetic differentiation
To estimate the genetic variability at each of the 10 loci on chromosome 8 flanking dhps, we calculated number of alleles (A) as well as expected heterozygosity (He) per locus using GenAlEx 6.2 [39]. He was calculated using the formula (He) = [n/(n−1)][1−Σpi2], where n is the number of P. falciparum isolates genotyped for that locus and pi is the frequency of the ith allele. The sampling variance for He was calculated as 2(n−1)/n3 {2(n−2) [Σ(pi3−(Σpi2)2]}. In order to investigate whether a selective sweep had occurred around dhps alleles, we divided the entire sample set into groups based on either the number of mutations (wild type, single, double and triple mutant group) or type of mutations/genotype (e.g. SGEAA) in the dhps gene. Significant difference between the mean He of any two groups was assessed using the Mann-Whitney U test implemented in the statistical package Stata version 8.1 for Windows (Stata Corporation, College Station, TX, USA). Differences were considered significant if the calculated P value was ≤0.05 (two-tailed test). Since microsatellite loci adjacent to a gene under selection (here dhps) may behave as non-neutral because of hitchhiking, we also performed similar analyses for 8 loci on chromosomes 2 and 3 which are putatively neutral.
We estimated FST between populations (following Weir and Cockerham) [40] at 10 dhps and 8 neutral microsatellite loci using the software GDA [41]. The populations were defined as wild, single, double and triple mutants according to the number of mutations in the dhps gene. 95% confidence intervals (CI) for each estimate and the standard deviation (SD) respectively, were computed by bootstrapping (over loci) and jackknifing (over population and loci) with 1000 permutations. To test whether FST at dhps loci differed from that of the neutral loci, we used Mann-Whitney U test.
It has been shown that loci directly under selection or linked to a selected loci exhibit extremely low or high levels of genetic differentiation between populations (FST) as compared to neutral loci [42],[43],[44],[45],[46],[47],[48]. Such outlier loci can be identified by the coalescent simulations method of Beaumont and Nichols [43] where the FST distribution is plotted as a function of He across loci under an island model of migration. We used this approach to disentangle selected from unselected loci, using FDIST program implemented in the software LOSITAN [49]. Simulations were initially run (under infinite-allele model) for only 8 neutral loci to calculate the mean neutral FST value which was subsequently used as “forced mean FST” to run the second round of simulation along with all 10 dhps loci. The 95% confidence interval was achieved with 10,000 simulations of 4 expected populations with sample size of 50.
To assess whether there was underlying population structure that would bias our conclusions regarding selection, we used FSTAT 2.9.3.2 [50] to partition the variation in the population using both dhps and neutral loci. Here we considered each geographical site (Pailin, Chumkiri, Memut and Rattanakiri) as a discrete population to calculate FST (using 1000 permutations). Since there were only 7 singly-infected isolates from Kampong Seila and is located in western Cambodia, they were grouped along with Pailin.
The strength of linkage disequilibrium across all 10 loci around dhps alleles was estimated as standardized index of association (IAS), a haplotype-wide measure of linkage [51],[52] using LIAN version 3.5 [53]. IAS was calculated using the formula IAS = (VD/Ve−1)/(L−1) where VD is the observed mismatch variance, Ve is the expected mismatch variance and L is the number of loci tested. The null hypothesis of complete linkage equilibrium (IAS = 0) was tested by the Monte-Carlo simulation process using 10,000 random permutations of the data.
Determining the number of origins for dhps alleles
To examine the probable number of origins of sulfadoxine-resistant dhps alleles in Cambodia, a median-joining network was constructed using 10-loci haplotypes in NETWORK version 4.5.1.0 (http://www.fluxus-engineering.com/sharenet.htm). Median-joining networks are used for reconstructing the phylogeny of regions with reticulate evolution [54]. In order to understand the genetic relationships among the dhps alleles of Southeast Asia (Cambodia), Africa (Kenya and Cameroon) and South America (Venezuela) an eBURST (version 3) analysis [55] was performed. We used only 7 loci (−11 Kb, −7.5 Kb, −2.9 Kb, −0.13 Kb, 0.03 Kb, 0.5 Kb, 1.4 Kb) for this comparison as data for the other 3 loci were not available for African and South American isolates. Data for African and South American isolates were obtained from a previous study from our laboratory [37].
Results
Removal of multiply-infected samples
The region of the dhps gene containing codons 436, 437, 540, 581 and 613 was sequenced in 234 Cambodian P. falciparum isolates (Fig. 1). Out of these, 22 (∼9%) isolates contained mixed alleles at one or more dhps codons based on multiple peaks in the sequencing electropherograms (see Table S2 for mixed dhps genotypes) while 212 had single genotype at all 5 dhps codons based on single peaks in the sequencing electropherograms at each nucleotide. These 212 samples were subjected to neutral microsatellite (8 loci) analysis and 71 were found to contain multiple alleles at one or more neutral loci. Since it is not easy to construct unambiguous microsatellite haplotypes using the samples with multiple infections [26],[56], these 71 multiply-infected samples were excluded and only 141 single clonal samples were typed for microsatellites on chromosome 8 around the dhps gene. Therefore, we present here the dhps codon distribution for 212 isolates (Table 1) and dhps microsatellites data for 141 isolates.

A novel triple mutant dhps allele in Cambodia
Approximately 11% of the isolates, mostly from the east, had the ancestral wild type dhps allele S436A437K540A581A613 (SAKAA). None of the isolates from the west had the ancestral wild type allele (Table 1). We observed mutations at all five dhps codons implicated in sulfadoxine resistance, with striking differences in the regional distribution of dhps mutations in Cambodia. The dhps allele with the single 437G (SGKAA) mutation was seen in ∼10% of the isolates, mostly in the east. However, 437G alone or along with other dhps mutations were found in almost 89% of the isolates (Table 1). Collectively, the double mutant dhps alleles were found in nearly 27% of the isolates in either of three combinations, 436A/437G (AGKAA, 7.5% mostly in the east); 437G/581G (SGKGA, 7.5%, predominantly in the south and west); and 437G/540E (SGEAA, 11.3%, mostly in the south and east). Approximately 51% of the isolates harbored triple mutations of either 436A/437G/540E (AGEAA, 20.7%), 437G/540E/581G (SGEGA, 10.3%), the novel genotype 437G/540N/581G (SGNGA, 19.3%) or 436A/437G/613E (AGKAT, 0.47%). The alleles with triple mutations were predominant in western and southern Cambodia whereas the ancestral wild type, single, or double mutants were the predominant dhps alleles in the east (Table 1, Fig. 1). The finding of the novel triple mutant dhps allele SGNGA in high frequency (19.3%) is particularly interesting since this allele has not been reported from any other malaria endemic region of the world. The frequency of the SGNGA allele in Pailin was greater (∼45%) than the other highly resistant alleles. In addition, we found two quadruple mutant dhps alleles (AGEAT and FGEAT), albeit in low frequency (0.94%), in south Cambodia. We observed that 581G and 540E/N were always associated with 437G (SGKGA; SGEAA; SGEGA; AGEAA; SGNGA) (Table 1). Particularly, 540N was always associated with 437G and 581G.
Genetic diversity and pattern of selective sweeps around dhps alleles
To analyze genetic diversity and investigate the pattern of selective sweeps around dhps alleles, we measured the number of alleles (A) and heterozygosity (He) for each of the 10 loci. The mean number of alleles (A) as well as mean heterozygosity (He) for the wild type dhps (SAKAA) were greater (A = 8.6±4.1; He = 0.78±0.08) compared to dhps alleles harboring single (A = 5.9±3.2; He = 0.65±0.06; P = 0.052), double (A = 4.5±2.4; He = 0.56±0.07; P = 0.035), and triple mutations (A = 3.6±1.3; He = 0.39±0.07; P = 0.005) (Fig. 2A, Fig. S1A; Table S3). This observation is compatible with these mutations being selected by drug pressure as indicated by a progressive decline in A and He correlated with an increase in the number of favorable mutations in the dhps gene. Such a trend is consistent with a model of positive directional selection. To determine if the reduction in variation at dhps was possibly due to demographic effects, such as a population bottleneck, we also measured He at 8 neutral loci on chromosomes 2 and 3 that are not known to be under any selection. The mean He for the 8 neutral loci in Cambodia was significantly greater (P = 0.016) than the mean He observed at 10 dhps loci. Moreover, there was no significant difference in He at neutral loci when wild, single, double and triple mutant groups were compared (Table S3). Unlike selection which only acts on one or a few loci in a genome, demographic effects are expected to act on the entire genome, thus the decline in variation is not likely due to demographic effects [57]. As shown in Fig. 2A, the valley of reduced variation for the triple mutant dhps alleles was deeper followed by the double mutant and single mutant alleles, suggestive of stronger selection for the triple mutant alleles. With the exception of the −11 Kb locus, which generally exhibited lower diversity in both wild type and all mutant groups, there was sharp reduction in He between −2.9 and 1.4 Kb region.
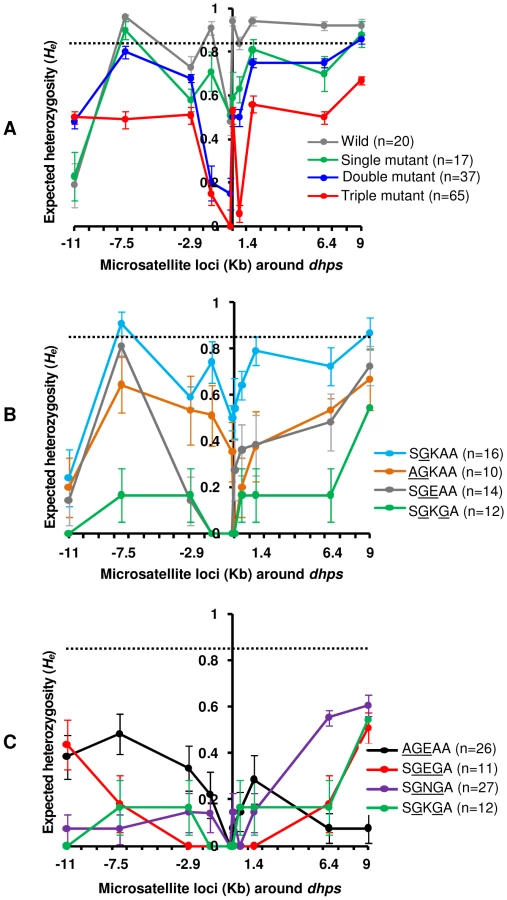
Since there were three types of double mutant dhps alleles in Cambodia, known to have different levels of in vitro resistance to sulfadoxine [13], we tested for differences in the selective sweeps around them. The double mutant SGKGA (A = 1.6±0.5; He = 0.13±0.05) showed significantly lower diversity compared to AGKAA (A = 2.8±1.2; He = 0.40±0.06; P = 0.01). However, there was no significant difference between the mean He at SGEAA (A = 2.8±1.4; He = 0.33±0.08) compared to SGKGA (P = 0.31) and AGKAA (P = 0.52) (Fig. 2B; Fig. S1B). The valley of reduced variation was also broader for SGKGA compared to the other two double mutants (Fig. 2B). All three triple mutant alleles (AGEAA: A = 2.7±1, He = 0.20±0.05; SGNGA: A = 2.3±0.8, He = 0.18±0.06; and SGEGA: A = 1.4±0.5, He = 0.13±0.06) showed lower level of diversity with no significant difference between them (Fig. 2C; Fig. S1C). Interestingly, the valley was both wider and symmetrical for the SGEGA allele as compared to the SGNGA and AGEAA alleles (Fig. 2C). This could be the result of the expansion of few lineages with the SGEGA genotype because limited genetic variability within the founding population would broaden the width of the selective sweep. Alternatively, the SGEGA may have recently evolved with insufficient time for recombination to break down potential linkage. Although the mean He for the loci surrounding SGKGA double mutant and SGEGA triple mutant alleles were almost similar (P = 0.48), there was some difference in the shape of the selection valley at these alleles (Fig. 2C).
Comparison of genetic differentiation at dhps and neutral microsatellite loci
The FST between populations (wild, single, double and triple mutant groups) were compared over 10 dhps and 8 neutral microsatellite loci as shown in Fig. 3A. As expected, FST values at all 10 dhps loci differed significantly from zero (Fig. 3A). On the other hand, three of the neutral loci also exhibited little but significant differentiation. The overall FST over the 10 dhps loci was 0.20 (SE = 0.03, CI = 0.15–0.26) and was significantly higher (Mann-Whitney U test: Z = 3.55, P = 0.0003) than the overall FST obtained over the 8 neutral loci (FST = 0.01, SE = 0.003 CI = 0.006–0.019).

The coalescent simulations method of Beaumont and Nichols [43] was used to disentangle selected loci from those behaving neutrally. The FST values for all dhps loci fell outside the 95% confidence interval of neutral expectations, indicative of being under directional selection or linked to a region of the genome under selection (Fig. 3B; see Table S4 for P value for deviation from neutral expectations). On the other hand, only one (C2M34) of the 8 neutral loci was found to lie marginally outside the 95% confidence interval (Fig. 3B; Table S4).
The pairwise FST comparisons between populations based on geography (4 regions in Cambodia) were performed using both dhps and neutral microsatellite loci (Table 2). The overall FST over the 10 dhps loci was 0.12, (SE = 0.02, CI = 0.09–0.16) and was significantly higher (Mann-Whitney U test: Z = −3.1, P = 0.0009) than the overall FST obtained over the 8 neutral loci (FST = 0.04, SE = 0.01, CI = 0.024–0.060).

Linkage disequilibrium (LD) around dhps alleles
A selective sweep reduces the amount of genetic variation at the chromosomal region containing a favorable mutation as well as at loci flanking the mutation, which leads to increased linkage disequilibrium (LD) around the target of selection. Therefore, in order to have an assessment of selection, we also measured LD as a standardized association index (IAS) between multiple loci flanking dhps alleles. There was no significant LD after Bonferroni correction (IAS = 0.019, P = 0.06) around the wild type (SAKAA) allele, whereas significant LD was observed around mutant dhps alleles. The IAS for single, double and triple mutant dhps alleles was 0.20 (P<0.01), 0.33 (P<0.01), and 0.22 (P<0.01), respectively.
Multiple origins of resistant dhps alleles in Cambodia independent from Africa and South America
We estimated the probable number of origin(s) for resistant dhps alleles in Cambodia based on the 10-loci microsatellite haplotypes around dhps (allele sizes and haplotypes information are given as supporting information in Table S5). The 10-loci haplotypes for all isolates with the wild type dhps allele were unique, showing complete linkage equilibrium between any two loci (open circles H1–H20 in Fig. 4A). The emergence of the 437G (SGKAA) mutation on multiple wild type genetic backgrounds led to an increase in the LD between loci and limited sharing of microsatellite haplotypes between the isolates (blue circles, particularly H21 to H24 in Fig. 4A). We found three major independent lineages for the double mutant dhps alleles: SGKGA, SGEAA and AGKAA in Cambodia (Fig. 4A). This was evident from the lack of shared 10-loci microsatellite haplotypes between these three alleles (Table S5). The SGKGA alleles shared identical microsatellite haplotype backgrounds, suggesting a single origin for this allele (maroon circles labeled H47 to H50 in Fig. 4A). Although a majority of the SGEAA allele shared common microsatellite haplotypes, a minority were also found to occur on diverse backgrounds (green circles in Fig. 4A). A similar scenario was also observed for the AGKAA allele (black circles in Fig. 4A). Among the triple mutants, the SGEGA and SGNGA alleles shared a common origin and appear to have emerged from the SGKGA double mutant, as indicated by their common haplotype backgrounds (Fig. 4A, 4B; Table S5). On the other hand, the AGEAA triple mutant originated from AGKAA on identical or nearly identical genetic backgrounds, suggesting multiple origins for this allele or possibly recombination events (Fig. 4A, 4B, Table S5). We did not find any evidence suggestive of the emergence of any triple mutant dhps alleles resulting from the SGEAA genetic background (Fig. 4A, 4B). Thus, our results seem to suggest multiple but limited origins for the highly resistant dhps alleles in Cambodia.

We compared the 7-loci microsatellite haplotype of the Cambodian dhps alleles with the Kenyan (SGEAA) and Cameroonian (SGKAA and AGKAA) dhps alleles (Fig. 5). In addition, we also compared backgrounds of the Cambodian alleles with the SGKGA and SGEGA alleles from Venezuela. Within Kenya, the majority of the SGEAA alleles had identical or nearly identical 7-loci microsatellite haplotypes (Fig. 5). However, a few SGEAA alleles also had unique haplotypes suggesting independent evolution of these alleles or recombination events. None of the isolates in Kenya and Cambodia shared identical 7-loci microsatellite haplotypes, suggesting local evolution of SGEAA allele in Kenya independent from Cambodia (Fig. 5). Similarly, unique haplotype profiles of the SGKAA and AGKAA alleles in Cameroon suggest multiple origins independent from Cambodia (Fig. 5). The only triple mutant found in South America (with a common founder) is SGEGA, and this allele had a completely distinct origin from the Cambodian SGKGA and SGEGA alleles. The double mutant SGKGA allele in Venezuela also had a completely distinct microsatellite haplotype compared to the Cambodian counterparts, indicating an independent evolution of dhps mutants in the South American region (Fig. 5).

Discussion
Unlike CQ and pyrimethamine resistance, there is no information available on the origins and evolutionary dynamics of sulfadoxine-resistant dhps alleles in the P. falciparum population from Southeast Asia. Therefore, the present study was undertaken to reveal the lineages of the highly resistant dhps alleles in this region. Samples in this study were analyzed from Pailin on the Thailand-Cambodia border as well as from four other regions in eastern and southern Cambodia. The isolates from the east predominantly had wild type (SAKAA), single mutant (SGKAA), and double mutant (AGKAA and SGEAA) dhps alleles, whereas those from the west and south predominantly had SGKGA double mutant and SGEGA, SGNGA and AGEAA triple mutants. These findings are in accordance with a previous finding that the parasites from eastern Cambodia are generally less resistant to SP (Cambodia National Malaria Control Program, unpublished data). It is important to note that although SP officially has not been in use in Cambodia for more than a decade, sulfadoxine-resistant dhps alleles are rampant in the population. This may also be explained by the easy availability of SP as a cheap over-the-counter antimalarial in Cambodia, particularly in rural areas. Interestingly, almost all mutant dhps alleles (188/189) had the 437G mutation, consistent with its critical role in sulfadoxine resistance and its wide occurrence globally [24],[58],[59]. The first mutation to occur and persist in populations in response to sulfadoxine pressure is 437G followed by mutations at additional codons, which lead to progressive increases in the level of resistance [15]. The 437G (SGKAA) or 437G/540E (SGEAA) dhps alleles in conjunction with double (51I//59R) or triple (51I/59R/108N) mutant dhfr alleles have been found to be associated with SP therapeutic failure in studies from Africa and Asia [58],[59],[60],[61],[62],[63],[64],[65],[66],[67],[68]. Similarly, the 437G/540E/581G (SGEGA) allele with the dhfr 51I//108N/164L allele has been correlated with in vivo SP resistance in South America [19].
Approximately 20% of the isolates, mainly from the west and south Cambodia, harbored a 540N mutation, always in association with 437G/581G (SGNGA). The Pailin area had ∼45% isolates with this novel triple mutant dhps allele (Table 1). To our knowledge, this is the first time that this allele has been observed in any malaria-endemic region of the world. However, it is not known how and when this new mutation emerged and what role this mutation plays in the response of the sulfadoxine. Nonetheless, its mere occurrence on the Thailand-Cambodia border and in southern Cambodia and strong selective signature around this allele (Fig. 2), suggest that this mutation may be playing an important role in sulfadoxine resistance. It is also likely that cross resistance to related drugs like cotrimoxazole (trimethoprim plus sulfamethoxazole), may have a role in selection of this allele. Cotrimoxazole is another antifolate combination widely used to treat bacterial infections. In one study, dhps mutations were analyzed in 53 Cambodian isolates, however the 540N mutation was not found, possibly due to the fact that most of these samples were from the eastern part of the country [16]. Similarly, we found only one isolate with the 540N mutation in eastern Cambodia (Table 1). However, a recent study has reported 540N dhps mutation in 19 of the 55 P. falciparum isolates from the Car Nicobar Islands of India, collected in October 2005 almost 9 months after the December 2004 tsunami, though; it was seen in association with 436A/437G (AGNAA) [69], unlike what we found in Cambodia. The 540N mutation was not found in samples (n = 50) collected before the tsunami. Lumb et al. [69] argue that after the tsunami many patients were simultaneously treated with two antifolate combinations: SP for malaria and trimethoprim-sulfamethoxazole for bacterial infections. This strong antifolate pressure may have allowed the selection of 540N mutation in the population. Interestingly, this mutation was not seen in samples collected at four other time points after the tsunami (n = 103) [69]. Like SGNGA, the role of the AGNAA dhps allele in sulfadoxine resistance is yet to be determined.
As expected under a model of a selective sweep, the dhps alleles with three mutations had the highest reduction in diversity at their flanking loci, followed by double and single mutant alleles (Fig. 2A). We also noted that the pattern of selective sweep at each allele was broadly correlated with the level of in vitro resistance to sulfadoxine. The triple mutant allele AGEAA had greater reduction in heterozygosity (inhibitory constant for sulfadoxine, Ki = 98.3±7.5 µM) followed by SGKGA (Ki = 16.2±2.7 µM), SGEAA (Ki = 27.7±6.1 µM), AGKAA (Ki = 19.9±4.0 µM), SGKAA (Ki = 1.39±0.23 µM) and SAKAA (wild type, Ki = 0.14±0.01 µM) [13]. The reduction in heterozygosity due to a selective sweep was greatest for the SGEGA and SGNGA triple mutant alleles indicating stronger selection for these alleles (Fig. 2C). However, the Ki's of these alleles for sulfadoxine are not known.
Underlying population structure may influence the amount of gene flow between resistant parasites, clouding estimates of selection. However, we have shown there are few barriers to gene flow either based on mutation or geography using the neutral loci (Fig. 3A and Table 2). Our estimates of global FST using the neutral microsatellite loci are below 0.05 using either geography or mutation to partition variation in Cambodia, indicating little genetic structuring in this population of P. falciparum. In contrast, quite high estimates of FST (greater than 0.12) are obtained using the microsatellite loci surrounding dhps (Fig. 3A) clearly supporting a role for strong selective pressure on this gene. Further, the FST outlier analysis also suggest that all 10 dhps loci have statistically higher FST values than expected under neutrality (Fig. 3B; Table S4), supporting that these loci are subject to directional selection.
The present data suggest three major independent origins of double mutant dhps alleles in Cambodia. The 437G mutation (SGKAA) was found to occur on multiple genetic backgrounds with subsequent mutations at 540, 436 and 581 producing the double mutants SGEAA, AGKAA and SGKGA, respectively (Fig. 4B). In accordance, the microsatellite haplotype backgrounds of these three double mutants were distinct from each other (Panel C, D and E in Table S5). Further, the SGKGA double mutant after acquiring 540E or 540N gave rise to SGEGA or SGNGA triple mutants, respectively, which is strongly supported by their common microsatellite backgrounds (Fig. 4B, Panel C of Table S5). Thus, SGEGA and SGNGA alleles have a common origin from the double mutant SGKGA. The SGKGA and SGEGA alleles in South America also have been found to share single and common haplotype backgrounds [21] yet different from Cambodia. At this point, we do not know whether 540E and 540N mutation in Cambodia simultaneously emerged and rose in frequency under sulfadoxine pressure or if they emerged at different time points. Analysis of the retrospective samples from this region may provide some clue on this aspect. However, because the 540E mutation is so widespread, it is likely that this would have been the first mutation to occur followed by 540N. Similarly, the widely spread triple mutant dhps allele AGEAA in Asia [17],[18] likely resulted from the AGKAA double mutant acquiring the mutation at codon 540. As shown in Fig. 4A (also in panel D of Table S5), the double and triple mutants share identical haplotype backgrounds, reaffirming their shared origin. It is likely that the novel AGNAA triple mutant allele observed on the Car Nicobar Islands [69] may have also emerged on the AGKAA background, similar to AGEAA (Fig. 4B). Thus, our data suggest that there are three major independent origins for the double mutant dhps alleles and two major independent origins for the triple mutant alleles in Cambodia (Fig. 4).
Gene flow plays an important role in the spread of drug resistant mutations as has been seen in the case of CQ and pyrimethamine resistance. Genetic evidence suggests that CQ and pyrimethamine resistance originated on the Thailand-Cambodia border and spread to Africa in the late 1970s and mid 1980s, respectively [26],[31],[70]. Thus, we also attempted to understand the evolutionary relationships between the Southeast Asian, African and South American dhps alleles. Results suggest that the SGEGA alleles both in South America [20],[21] and Cambodia evolved independently (Fig. 5). In a recent study, Pearce et al. [24] attempted to understand the origin and dispersal of sulfadoxine resistance in Africa by analyzing three microsatellite loci flanking the dhps gene. Based on the data from several African countries it has been concluded that the sulfadoxine resistance-conferring dhps alleles in east Africa (predominantly SGEAA) are different from those in west Africa (SGKAA and AGKAA) and these alleles are associated with multiple genetic backgrounds consisting of five major lineages [24]. This corroborates with our previous data [25] as well as the present study, which confirms multiple, independent origins of the above dhps alleles in east and west Africa (Fig. 5). None of the triple mutant dhps alleles reported in other parts of the world has appeared in Africa. It remains to be seen whether a triple mutant dhps allele will independently evolve in Africa or expand from migration of parasites from other parts of the world, as in the case of triple mutant dhfr. It is surprising though that the major triple mutant dhfr allele found in Africa is believed to have been imported from Southeast Asia and spread across the continent while none of the triple mutant dhps alleles have been established in Africa. One possibility may be that the introduction of dhfr mutant allele may have preceded the development of triple mutant dhps allele.
In conclusion, results from this study reveal some very interesting findings: i) presence of a novel triple mutant dhps SGNGA as a predominant allele in Cambodia along with two previously reported triple mutant alleles SGEGA and AGEAA; ii) the triple mutant dhps alleles were present at high frequencies in the western and southern parts of Cambodia while they were rarely seen in the east, indicating that sulfadoxine resistance may be a lesser problem in the east; iii) the triple mutant dhps alleles have not declined over several years after SP was officially removed as the drug of choice for the primary treatment of malaria in this region; iv) at least three independent origins of the double mutants and two independent origins of triple mutant dhps alleles are evident in Cambodia; v) the origin of triple mutant dhps allele SGEGA in South America is completely distinct from the Cambodian SGEGA allele; vi) the double mutant SGEAA allele in Kenya and AGKAA allele in Cameroon are evolutionary distinct from their counterparts in Cambodia, indicating multiple origins for these alleles. Finally, it is also evident that unlike pfcrt and dhfr resistant alleles, which have single origin in Thailand-Cambodia region, the dhps resistant alleles have multiple origins in this region. Thus, we have provided comprehensive new data illustrating multiple global origins for sulfadoxine resistant dhps alleles.
Supporting Information
{kind=link}
Zdroje
1. EylesDE
HooCC
WarrenM
SandoshamAA
1963 Plasmodium falciparum resistant to chloroquine in Cambodia. Am J Trop Med Hyg 12 840 843
2. VerdragerJ
1986 Epidemiology of the emergence and spread of drug-resistant falciparum malaria in South-East Asia and Australasia. J Trop Med Hyg 89 277 289
3. HurwitzES
JohnsonD
CampbellCC
1981 Resistance of Plasmodium falciparum malaria to sulfadoxine-pyrimethamine (‘Fansidar’) in a refugee camp in Thailand. Lancet 1 1068 1070
4. WongsrichanalaiC
MeshnickSR
2008 Declining artesunate-mefloquine efficacy against falciparum malaria on the Cambodia-Thailand border. Emerg Infect Dis 14 716 719
5. WHO 2002 Development of South-Asia surveillance network for malaria drug resistance. Report on informal consultative meeting. World Health Organization, Geneva, Switzerland
6. HurlyMG
1959 Potentiation of pyrimethamine by sulphadiazine in human malaria. Trans R Soc Trop Med Hyg 53 412 413
7. ChulayJD
WatkinsWM
SixsmithDG
1984 Synergistic antimalarial activity of pyrimethamine and sulfadoxine against Plasmodium falciparum in vitro. Am J Trop Med Hyg 33 325 330
8. LaingAB
1970 Studies on the chemotherapy of malaria. I. The treatment of overt falciparum malaria with potentiating combinations of pyrimethamine and sulphormethoxine or dapsone in The Gambia. Trans R Soc Trop Med Hyg 64 562 568
9. WHO 2008 World Malaria Report 2008, World Health Organization, Geneva, Switzerland
10. BrownGM
1962 The biosynthesis of folic acid. II. Inhibition by sulfonamides. J Biol Chem 237 536 540
11. BrooksDR
WangP
ReadM
WatkinsWM
SimsPF
1994 Sequence variation of the hydroxymethyldihydropterin pyrophosphokinase: dihydropteroate synthase gene in lines of the human malaria parasite, Plasmodium falciparum, with differing resistance to sulfadoxine. Eur J Biochem 224 397 405
12. TrigliaT
CowmanAF
1994 Primary structure and expression of the dihydropteroate synthetase gene of Plasmodium falciparum. Proc Natl Acad Sci U S A 91 7149 7153
13. TrigliaT
MentingJG
WilsonC
CowmanAF
1997 Mutations in dihydropteroate synthase are responsible for sulfone and sulfonamide resistance in Plasmodium falciparum. Proc Natl Acad Sci U S A 94 13944 13949
14. WangP
ReadM
SimsPF
HydeJE
1997 Sulfadoxine resistance in the human malaria parasite Plasmodium falciparum is determined by mutations in dihydropteroate synthetase and an additional factor associated with folate utilization. Mol Microbiol 23 979 986
15. TrigliaT
WangP
SimsPF
HydeJE
CowmanAF
1998 Allelic exchange at the endogenous genomic locus in Plasmodium falciparum proves the role of dihydropteroate synthase in sulfadoxine-resistant malaria. EMBO J 17 3807 3815
16. KhimN
BouchierC
EkalaMT
IncardonaS
LimP
2005 Countrywide survey shows very high prevalence of Plasmodium falciparum multilocus resistance genotypes in Cambodia. Antimicrob Agents Chemother 49 3147 3152
17. van den BroekIV
van der WardtS
TalukderL
ChakmaS
BrockmanA
2004 Drug resistance in Plasmodium falciparum from the Chittagong Hill Tracts, Bangladesh. Trop Med Int Health 9 680 687
18. AhmedA
LumbV
DasMK
DevV
Wajihullah
2006 Prevalence of mutations associated with higher levels of sulfadoxine-pyrimethamine resistance in Plasmodium falciparum isolates from Car Nicobar Island and Assam, India. Antimicrob Agents Chemother 50 3934 3938
19. KublinJG
WitzigRS
ShankarAH
ZuritaJQ
GilmanRH
1998 Molecular assays for surveillance of antifolate-resistant malaria. Lancet 351 1629 1630
20. CorteseJF
CaraballoA
ContrerasCE
PloweCV
2002 Origin and dissemination of Plasmodium falciparum drug-resistance mutations in South America. J Infect Dis 186 999 1006
21. McCollumAM
MuellerK
VillegasL
UdhayakumarV
EscalanteAA
2007 Common origin and fixation of Plasmodium falciparum dhfr and dhps mutations associated with sulfadoxine-pyrimethamine resistance in a low-transmission area in South America. Antimicrob Agents Chemother 51 2085 2091
22. BaconDJ
McCollumAM
GriffingSM
SalasC
SoberonV
2009 Dynamics of malaria drug resistance patterns in the Amazon basin region following changes in Peruvian national treatment policy for uncomplicated malaria. Antimicrob Agents Chemother 53 2042 2051
23. ZhouZ
GriffingSM
de OliveiraAM
McCollumAM
QuezadaWM
2008 Decline in sulfadoxine-pyrimethamine-resistant alleles after change in drug policy in the Amazon region of Peru. Antimicrob Agents Chemother 52 739 741
24. PearceRJ
PotaH
EveheMS
Ba elH
Mombo-NgomaG
2009 Multiple origins and regional dispersal of resistant dhps in African Plasmodium falciparum malaria. PLoS Med 6 e1000055 doi:10.1371/journal.pmed.1000055
25. McCollumAM
BascoLK
TaharR
UdhayakumarV
EscalanteAA
2008 Hitchhiking and selective sweeps of Plasmodium falciparum sulfadoxine and pyrimethamine resistance alleles in a population from central Africa. Antimicrob Agents Chemother 52 4089 4097
26. NairS
WilliamsJT
BrockmanA
PaiphunL
MayxayM
2003 A selective sweep driven by pyrimethamine treatment in southeast asian malaria parasites. Mol Biol Evol 20 1526 1536
27. McCollumAM
PoeAC
HamelM
HuberC
ZhouZ
2006 Antifolate resistance in Plasmodium falciparum: multiple origins and identification of novel dhfr alleles. J Infect Dis 194 189 197
28. MitaT
TanabeK
TakahashiN
CulletonR
NdoungaM
2009 Indigenous evolution of Plasmodium falciparum pyrimethamine resistance multiple times in Africa. J Antimicrob Chemother 63 252 255
29. CertainLK
BricenoM
KiaraSM
NzilaAM
WatkinsWM
2008 Characteristics of Plasmodium falciparum dhfr haplotypes that confer pyrimethamine resistance, Kilifi, Kenya, 1987–2006. J Infect Dis 197 1743 1751
30. RoperC
PearceR
BredenkampB
GumedeJ
DrakeleyC
2003 Antifolate antimalarial resistance in southeast Africa: a population-based analysis. Lancet 361 1174 1181
31. RoperC
PearceR
NairS
SharpB
NostenF
2004 Intercontinental spread of pyrimethamine-resistant malaria. Science 305 1124
32. MaigaO
DjimdeAA
HubertV
RenardE
AubouyA
2007 A shared Asian origin of the triple-mutant dhfr allele in Plasmodium falciparum from sites across Africa. J Infect Dis 196 165 172
33. ArieyF
FandeurT
DurandR
RandrianarivelojosiaM
JambouR
2006 Invasion of Africa by a single pfcrt allele of South East Asian type. Malar J 5 34
34. NdiayeD
DailyJP
SarrO
NdirO
GayeO
2006 Defining the origin of Plasmodium falciparum resistant dhfr isolates in Senegal. Acta Trop 99 106 111
35. LynchC
PearceR
PotaH
CoxJ
AbekuTA
2008 Emergence of a dhfr mutation conferring high-level drug resistance in Plasmodium falciparum populations from southwest Uganda. J Infect Dis 197 1598 1604
36. MitaT
TanabeK
TakahashiN
TsukaharaT
EtoH
2007 Independent evolution of pyrimethamine resistance in Plasmodium falciparum isolates in Melanesia. Antimicrob Agents Chemother 51 1071 1077
37. McCollumAM
2007 Drug resistant Plasmodium falciparum: Genetic hitchhiking, selective sweeps, and multiple origins [Ph.D Thesis], Emory University, Atlanta, USA
38. SuX
FerdigMT
HuangY
HuynhCQ
LiuA
1999 A genetic map and recombination parameters of the human malaria parasite Plasmodium falciparum. Science 286 1351 1353
39. PeakallR
SmousePE
2006 GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Notes 6 288 295
40. WeirBS
CockerhamCC
1984 Estimating F-statistics for the analysis of population structure. Evolution 38 1358 1370
41. LewisPO
ZaykinD
2001 Genetic Data Analysis: Computer program for the analysis of allelic data. Version 1.0 (d16c). Free program distributed by the authors over the internet from http://lewis.eeb.uconn.edu/lewishome/software.html
42. LewontinRC
KrakauerJ
1973 Distribution of gene frequency as a test of the theory of the selective neutrality of polymorphisms. Genetics 74 175 195
43. BeaumontMA
NicholsRA
1996 Evaluating loci for use in the genetic analysis of population structure. Proc R Soc Lond B: Biol Sci 263 1619 1626
44. WildingCS
ButlinRK
GrahameJ
2001 Differential gene exchange between parapatric morphs of Littorina saxatilis detected using AFLP markers. J Evol Biol 14 611 619
45. AndersonTJ
NairS
SudimackD
WilliamsJT
MayxayM
2005 Geographical distribution of selected and putatively neutral SNPs in Southeast Asian malaria parasites. Mol Biol Evol 22 2362 2374
46. StorzJF
2005 Using genome scans of DNA polymorphism to infer adaptive population divergence. Mol Ecol 14 671 688
47. BeaumontMA
2005 Adaptation and speciation: what can F(st) tell us? Trends Ecol Evol 20 435 440
48. DemontisD
PertoldiC
LoeschckeV
MikkelsenK
AxelssonT
2009 Efficiency of selection, as measured by single nucleotide polymorphism variation, is dependent on inbreeding rate in Drosophila melanogaster. Mol Ecol 18 4551 4563
49. AntaoT
LopesA
LopesRJ
Beja-PereiraA
LuikartG
2008 LOSITAN: a workbench to detect molecular adaptation based on a Fst-outlier method. BMC Bioinformatics 9 323
50. GoudetJ
2001 FSTAT, a program to estimate and test gene diversities and fixation indices Version 2.9.3.2. www2.unil.ch/popgen/softwares/fstat.htm
51. SmithJM
SmithNH
O'RourkeM
SprattBG
1993 How clonal are bacteria? Proc Natl Acad Sci U S A 90 4384 4388
52. HudsonRR
1994 Analytical results concerning linkage disequilibrium in models with genetic transformation and conjugation. J Evol Biol 7 535 548
53. HauboldB
HudsonRR
2000 LIAN 3.0: detecting linkage disequilibrium in multilocus data. Linkage Analysis. Bioinformatics 16 847 848
54. BandeltHJ
ForsterP
RohlA
1999 Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol 16 37 48
55. FeilEJ
LiBC
AanensenDM
HanageWP
SprattBG
2004 eBURST: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J Bacteriol 186 1518 1530
56. NashD
NairS
MayxayM
NewtonPN
GuthmannJP
2005 Selection strength and hitchhiking around two anti-malarial resistance genes. Proc Biol Sci 272 1153 1161
57. HarrisEE
HeyJ
1999 X chromosome evidence for ancient human histories. Proc Natl Acad Sci U S A 96 3320 3324
58. GregsonA
PloweCV
2005 Mechanisms of resistance of malaria parasites to antifolates. Pharmacol Rev 57 117 145
59. PicotS
OlliaroP
de MonbrisonF
BienvenuAL
PriceRN
2009 A systematic review and meta-analysis of evidence for correlation between molecular markers of parasite resistance and treatment outcome in falciparum malaria. Malar J 8 89
60. CurtisJ
DuraisinghMT
WarhurstDC
1998 In vivo selection for a specific genotype of dihydropteroate synthetase of Plasmodium falciparum by pyrimethamine-sulfadoxine but not chlorproguanil-dapsone treatment. J Infect Dis 177 1429 1433
61. WangP
LeeCS
BayoumiR
DjimdeA
DoumboO
1997 Resistance to antifolates in Plasmodium falciparum monitored by sequence analysis of dihydropteroate synthetase and dihydrofolate reductase alleles in a large number of field samples of diverse origins. Mol Biochem Parasitol 89 161 177
62. NzilaAM
MberuEK
SuloJ
DayoH
WinstanleyPA
2000 Towards an understanding of the mechanism of pyrimethamine-sulfadoxine resistance in Plasmodium falciparum: genotyping of dihydrofolate reductase and dihydropteroate synthase of Kenyan parasites. Antimicrob Agents Chemother 44 991 996
63. OmarSA
AdaguIS
GumpDW
NdaruNP
WarhurstDC
2001 Plasmodium falciparum in Kenya: high prevalence of drug-resistance-associated polymorphisms in hospital admissions with severe malaria in an epidemic area. Ann Trop Med Parasitol 95 661 669
64. StaedkeSG
SendagireH
LamolaS
KamyaMR
DorseyG
2004 Relationship between age, molecular markers, and response to sulphadoxine-pyrimethamine treatment in Kampala, Uganda. Trop Med Int Health 9 624 629
65. KunJF
LehmanLG
LellB
Schmidt-OttR
KremsnerPG
1999 Low-dose treatment with sulfadoxine-pyrimethamine combinations selects for drug-resistant Plasmodium falciparum strains. Antimicrob Agents Chemother 43 2205 2208
66. KublinJG
DzinjalamalaFK
KamwendoDD
MalkinEM
CorteseJF
2002 Molecular markers for failure of sulfadoxine-pyrimethamine and chlorproguanil-dapsone treatment of Plasmodium falciparum malaria. J Infect Dis 185 380 388
67. KyabayinzeD
CattamanchiA
KamyaMR
RosenthalPJ
DorseyG
2003 Validation of a simplified method for using molecular markers to predict sulfadoxine-pyrimethamine treatment failure in African children with falciparum malaria. Am J Trop Med Hyg 69 247 252
68. NageshaHS
DinS
CaseyGJ
SusantiAI
FryauffDJ
2001 Mutations in the pfmdr1, dhfr and dhps genes of Plasmodium falciparum are associated with in-vivo drug resistance in West Papua, Indonesia. Trans R Soc Trop Med Hyg 95 43 49
69. LumbV
DasMK
MittraP
AhmedA
KumarM
2009 Emergence of an unusual sulfadoxine-pyrimethamine resistance pattern and a novel K540N mutation in dihydropteroate synthetase in Plasmodium falciparum isolates obtained from Car Nicobar Island, India, after the 2004 Tsunami. J Infect Dis 199 1064 1073
70. WoottonJC
FengX
FerdigMT
CooperRA
MuJ
2002 Genetic diversity and chloroquine selective sweeps in Plasmodium falciparum. Nature 418 320 323
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2010 Číslo 3
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- Kaposi's Sarcoma-Associated Herpesvirus ORF57 Protein Binds and Protects a Nuclear Noncoding RNA from Cellular RNA Decay Pathways
- Endocytosis of the Anthrax Toxin Is Mediated by Clathrin, Actin and Unconventional Adaptors
- Perforin and IL-2 Upregulation Define Qualitative Differences among Highly Functional Virus-Specific Human CD8 T Cells
- Exoerythrocytic Parasites Secrete a Cysteine Protease Inhibitor Involved in Sporozoite Invasion and Capable of Blocking Cell Death of Host Hepatocytes
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy