
Cryo Electron Tomography of Native HIV-1 Budding Sites
The structure of immature and mature HIV-1 particles has been analyzed in detail by cryo electron microscopy, while no such studies have been reported for cellular HIV-1 budding sites. Here, we established a system for studying HIV-1 virus-like particle assembly and release by cryo electron tomography of intact human cells. The lattice of the structural Gag protein in budding sites was indistinguishable from that of the released immature virion, suggesting that its organization is determined at the assembly site without major subsequent rearrangements. Besides the immature lattice, a previously not described Gag lattice was detected in some budding sites and released particles; this lattice was found at high frequencies in a subset of infected T-cells. It displays the same hexagonal symmetry and spacing in the MA-CA layer as the immature lattice, but lacks density corresponding to NC-RNA-p6. Buds and released particles carrying this lattice consistently lacked the viral ribonucleoprotein complex, suggesting that they correspond to aberrant products due to premature proteolytic activation. We hypothesize that cellular and/or viral factors normally control the onset of proteolytic maturation during assembly and release, and that this control has been lost in a subset of infected T-cells leading to formation of aberrant particles.
Published in the journal:
. PLoS Pathog 6(11): e32767. doi:10.1371/journal.ppat.1001173
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1001173
Summary
The structure of immature and mature HIV-1 particles has been analyzed in detail by cryo electron microscopy, while no such studies have been reported for cellular HIV-1 budding sites. Here, we established a system for studying HIV-1 virus-like particle assembly and release by cryo electron tomography of intact human cells. The lattice of the structural Gag protein in budding sites was indistinguishable from that of the released immature virion, suggesting that its organization is determined at the assembly site without major subsequent rearrangements. Besides the immature lattice, a previously not described Gag lattice was detected in some budding sites and released particles; this lattice was found at high frequencies in a subset of infected T-cells. It displays the same hexagonal symmetry and spacing in the MA-CA layer as the immature lattice, but lacks density corresponding to NC-RNA-p6. Buds and released particles carrying this lattice consistently lacked the viral ribonucleoprotein complex, suggesting that they correspond to aberrant products due to premature proteolytic activation. We hypothesize that cellular and/or viral factors normally control the onset of proteolytic maturation during assembly and release, and that this control has been lost in a subset of infected T-cells leading to formation of aberrant particles.
Introduction
HIV-1 particles are assembled at the cell membrane, as the 55 kDa viral polyprotein Gag multimerizes on its inner face [1]. Gag recruits other viral components such as the RNA genome and the surface spike proteins, as well as cellular proteins of the ESCRT machinery required for virus release [2], [3], [4]. The viral protease (PR) is essential to convert the immature form of the virion into an infectious mature particle. Both forms of the virion are pleiomorphic structures, with the repetitive structural elements of the virus arranged non-symmetrically and variably from one particle to the other.
In the immature virion, uncleaved Gag is anchored to the plasma membrane via a charged surface and a myristoyl tail in its N-terminal matrix (MA) domain [1]. As shown by cryo electron microscopy (cEM), Gag arranges in a regular manner, with its internal capsid (CA) domain forming a hexameric lattice with a spacing of 8.0 nm [5]. C-terminally of CA, the nucleocapsid (NC) domain binds the RNA genome, and the p6 domain recruits the ESCRT machinery to facilitate particle release [6], [7]. CA and NC, as well as NC and p6, are separated by short spacer peptides (SP1 and SP2, respectively) which are processed during maturation.
Proteolytic maturation of HIV-1 has been proposed to initiate at or shortly after assembly and release [8]. The active dimeric form of the viral PR cleaves Gag and GagPol at multiple sites, leading to the structural transition from the immature particle with its Gag shell forming a truncated sphere to the mature particle with its cone-shaped CA core encasing the condensed nucleoprotein complex in the interior of the virion. In an in vitro study Pettit et al. detected large differences in processing kinetics at the five cleavage sites in Gag, the cleavage at the site between SP1 and NC being an order of magnitude faster than the second fastest cleavage event [9]. These results suggested ordered processing during virion maturation, and this was supported by mutagenesis studies of individual cleavage sites [10]. Processing at all sites except NC-SP1 [11] appears to be important for infectivity and small amounts of partially processed Gag products have been shown to exhibit a strong trans-dominant negative effect on viral infectivity [12], [13], [14]. Furthermore, premature processing [15] as well as low concentrations of protease inhibitors, insufficient to significantly affect proteolytic maturation [14], [16], [17], were both shown to efficiently block viral infectivity, indicating an intricate interplay between proteolytic activation and virus formation. Morphological maturation is believed to occur rapidly following release of the immature virion as no intermediates of maturation have been observed so far and all cellular budding sites appear to carry an immature Gag shell.
The last years have seen increasingly detailed structural studies of released HIV-1 particles in their immature and mature forms [18], [19], [20], [21], [22], [23], [24], [25]. Analysis of immature virions by cryo electron tomography (cET) revealed that the immature lattice covers only part of the viral membrane [22], arranging as an incomplete “∼2/3” sphere [26]. These studies suggested that ESCRT involvement in HIV-1 release occurs earlier than previously thought and that so-called late budding sites carrying an almost complete Gag lattice are likely to be dead-end products rather than intermediates in the release pathway [26]. A later analysis by cET and subtomogram averaging provided a 17 Å structure of the immature Gag lattice and showed how this incomplete hexagonal lattice attains its curvature through inclusion of symmetry defects of irregular shape and size [19]. In the mature capsid, CA also forms a hexameric lattice, but with a different lattice constant and arrangement than in the immature lattice [27]. Curvature of the mature capsid is achieved by asymmetric incorporation of pentameric defects at the narrow and wide end of the cone [28]. While cET studies of released virions have significantly advanced our understanding of HIV-1 morphogenesis, we are currently lacking any three-dimensional information regarding native intracellular particle assembly and budding. These processes have hitherto been inaccessible for detailed structural studies due to the lack of an experimental system enabling their visualization by cET. Here, we present a structural study of HIV-1 assembly sites by cET of intact, plunge-frozen human cells. These snapshots allow detailed structural interpretations of the Gag lattice structure at its assembly site in situ, as well as the arrangement of the cortical actin in its immediate vicinity. We further determined the structure of a previously not reported Gag lattice type lacking the NC-RNA layer which we propose to be associated with loss of control of PR activation.
Results
Visualization of HIV-1 assembly by cryo electron tomography
To enable structural analysis of HIV-1 assembly in its cellular context, we set out to establish a system for studying this process by cET on intact human cells. Since cET is limited to samples thinner than ∼500 nm, and thus to thin peripheral areas of adherent cells, the natural HIV-1 host cell types (T-cells and macrophages) cannot be used for cET. This led us to use two human glioblastoma cell lines, U-87 MG and U-373 MG. These cells grow adherently, have extended, thin peripheral areas, and are amenable to transfection or transduction with adenoviral vectors. Transfection efficiency proved insufficient for structural analyses, however, and we therefore constructed adenoviral vectors expressing HIV-1 Gag (AdGag) and HIV-1 Gag-Pol (AdGagPol), respectively. Both adenoviral vectors were based on Rev-independent versions of the respective genes [29], allowing expression of either Gag or Gag-Pol without co-transduction of Rev. U-87 MG or U-373 MG cells were seeded on EM sample grids concomitant with transduction using the respective adenoviral vector. Immunofluorescence staining for HIV-1 CA at 24–48 h post transduction revealed that almost 100% of the cells expressed HIV-1 Gag, while retaining normal adherent morphology (Fig. 1A–B). Furthermore, when U-373 MG cells were transduced with AdGagPol, specific cleavage products of HIV-1 Gag were found in the cell lysate and in the particle fraction purified from the culture medium (Fig. 1C), while particle release but no processing was observed for AdGag (data not shown). Cells transduced with AdGag or AdGagPol were vitrified by plunge-freezing and subjected to cEM. Upon inspection of the vitrified grids, HIV-1 assembly sites and released virions were present at low but reproducible frequencies at the periphery of the cells (Fig. 1D) in regions accessible to cET. A total of 40 tilt series were recorded on such positions of AdGag and AdGagPol infected cells, respectively (Table 1).

In cryo electron tomograms of AdGag or AdGagPol transduced cells (Fig. 2) structural features of the cytoplasm were preserved at the high level previously reported for cellular cET [30], with single ribosomes, actin filaments and microtubules being clearly resolved. Both viral budding sites (Fig. 2A, C–F) and released virions (Fig. 2B–C, E) were present in the recorded tomograms. The budding sites were found at the plasma membrane on lamellipodia–like areas (Fig. 2C, E), occasionally in membrane invaginations (Fig. 2B, D) or at the tips (Fig. 2F, 3B) or sides (Fig. 2A, 3A) of filopodial structures. In the immature Gag lattice of these budding sites and released particles, the two density layers corresponding to CA and NC-RNA could be resolved, and occasionally the hexagonal symmetry in the CA layer was apparent by eye in more tangential tomographic slices. Released immature particles (ip) were found adjacent to cells expressing Gag (Fig. 2B, D) or GagPol (Fig. 2E), whereas released particles with mature morphology (mp) were only found adjacent to cells expressing GagPol (Fig. 2E). No density attributable to components of the ESCRT complex was detected at budding sites at the current resolution.

The preservation of cytoplasmic structure in cET allowed for three-dimensional snapshots of filamentous actin associated with HIV-1 budding sites. The presence of actin filaments was generally high at budding sites (Fig. 2A, C, F and Fig.3A–D), as judged by visual inspection where F-actin is clearly resolvable from e.g. intermediate filaments. Furthermore, we often observed what appeared to be a direct interaction of actin filaments with the budding site (Fig. 2A, F; Fig. 3D). To categorize the budding sites with respect to their actin context, they were sorted into 5 classes according to the type of actin structures they were associated with (Fig.3, top panel). This classification revealed that 34 of the 39 budding sites analyzed were found adjacent to filamentous actin (Fig. 3A–D), with half of the buds (20 of 39) appearing on the sides or tips of filopodia-like structures characterized by a parallel actin organization (Fig. 3A–B).

In situ organization of the assembling Gag lattice
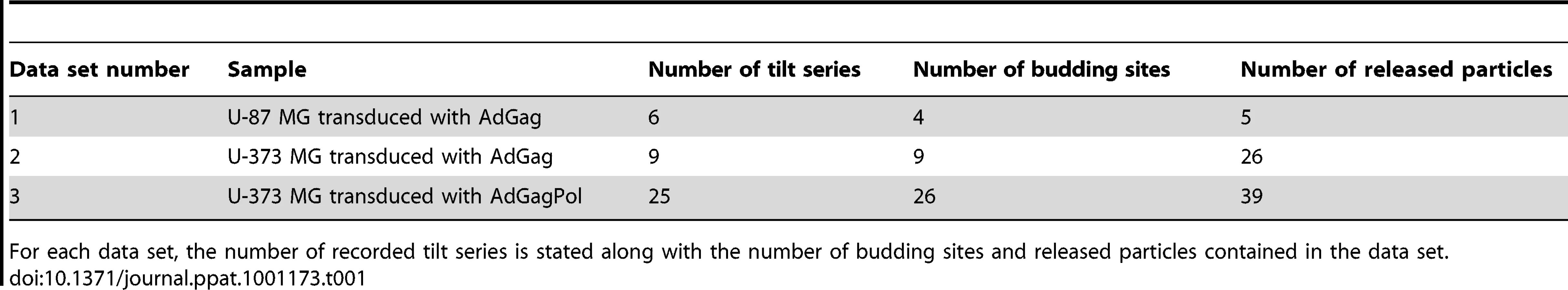
Cryo electron tomograms containing budding sites on intact cells with apparently high signal-to-noise ratio were selected for further analysis by sub-tomogram averaging as previously performed for isolated immature HIV-1 [19]. This analysis provides two kinds of information about the Gag protein lattice: the global arrangement of the repetitive elements into an ordered lattice structure (“lattice map”) and the local structure of these repetitive elements (“unit cell structure”).
The immature Gag lattice parameters in the budding sites were the same as previously determined for released immature virions [5], [19]: a hexagonal lattice with a lattice constant of 8.0 nm. The lattice maps derived from the budding sites (Fig. 4A) revealed several sites of symmetry breakage, similarly heterogeneous in size and shape as those described for the released virions [19].

The unit cell structure derived from the cellular tomograms contained density corresponding to the N-terminal and C-terminal domains of CA, respectively, as well as density for the membrane and the NC-RNA layer (Fig. 5A). At the present resolution (roughly 40 Å by the 0.5 Fourier shell correlation criterion), this structure was indistinguishable from that previously reported for released particles [19].
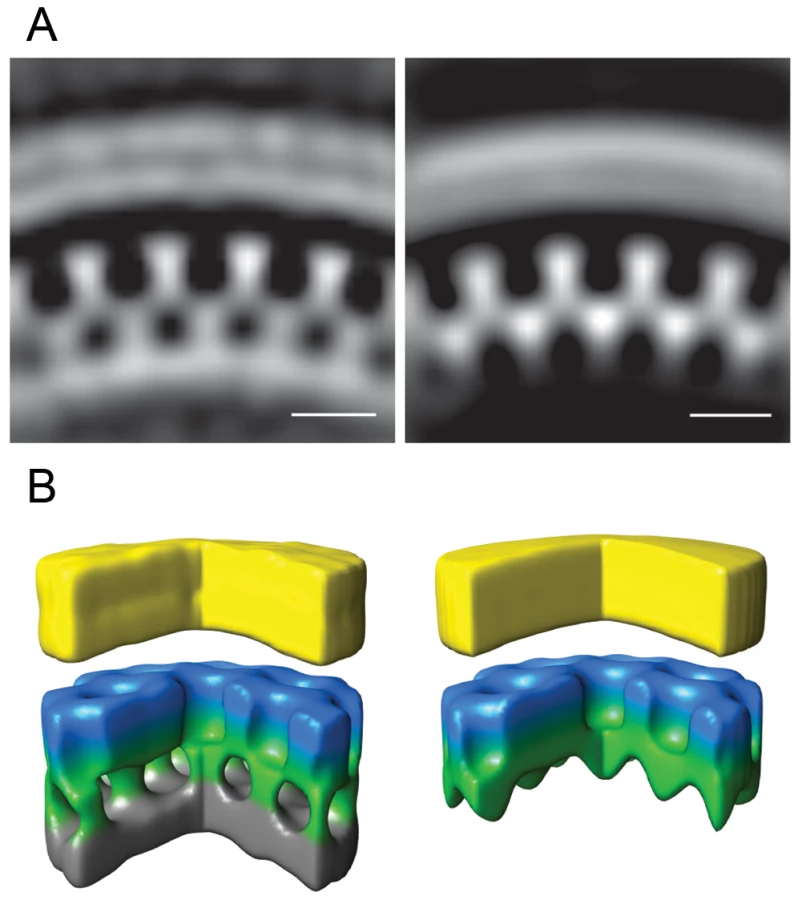
A previously undescribed Gag lattice form is present in tomograms of GagPol-expressing cells
Immunoblot analysis of AdGagPol-transduced cells and the particle fraction recovered from the culture medium showed proteolytic processing of Gag (Fig. 1C). Furthermore, particles containing mature-looking cores were found in cryo electron tomograms in immediate proximity of cellular budding sites and next to immature particles (Fig. 2E). Of note, structures resembling the mature capsid were never found in budding sites still connected to the plasma membrane. Besides the well-described mature and immature particles, we observed a third, previously not described form of particles in the tomograms of AdGagPol-transduced cells (Fig. 6A). This apparently “intermediate morphology” exhibiting a thinner Gag lattice than the immature structure was observed both in extracellular particles (Fig. 6A, Fig. S2 in Supporting Information S1) and in HIV-1 budding sites connected to the cell (Fig. 6A–B). Budding sites and extracellular particles with the previously unrecognized Gag lattice were characterized by a single density layer bound to the inner face of the membrane (Figure 6B), as opposed to the two layers (CA and NC-RNA) seen in the immature lattice (Fig. 6C). Density plots orthogonal to the cell membrane confirmed that the density layer found in these novel structures was located at the same position relative to the plasma membrane (peak at −12 nm) as the CA layer of the complete immature lattice (Fig. S1 in Supporting Information S1).

Strikingly, budding sites and released particles with this novel Gag lattice displayed the same hexagonal symmetry and lattice spacing as the immature lattice (Fig. 4B). The apparently intermediate type lattices were typically more complete with smaller lattice defects than the immature lattices, but the lattice defects were similarly heterogeneous. Furthermore, the unit cell structure of the new lattice revealed an arrangement identical to the CA region of the immature lattice, with density corresponding to the N - and C-terminal domains of CA (Fig. 5B), but lacking any density corresponding to the NC-RNA layer. The cellular budding sites and extracellular particles containing this lattice also lacked the internal density corresponding to a condensed NC-RNA complex. Such densities were readily observed within mature capsids and in virions carrying partially processed Gag proteins (de Marco et al., accompanying paper).
Budding sites with the novel Gag lattice are also observed in infected T-cell lines
To determine whether the newly described lattice also occurs in other HIV-1 producing cells including T-cell lines, we recorded electron tomograms on sections of resin-embedded cells producing HIV-1. Although resin-embedded material is not preserved at the molecular level, it might still be possible to distinguish the two lattice types in these data, and this would allow recording sufficiently large data sets for statistical analyses. Visual inspection of resin-embedded sections from infected MT4 cells suggested the presence of a thinner density layer in some budding sites (Fig. 7A, B). This interpretation was validated based on the following criteria: (i) different types of membrane-bound densities in budding sites in resin embeddings could be classified by an unbiased scheme, and (ii) the presence of structures corresponding to the novel Gag lattice was dependent on an active viral PR.

Classification of budding sites and particles was based on linear density plots of the membrane-bound Gag density. Fig. 7C shows a typical example, demonstrating that enough structure is retained to resolve the plasma membrane and distinct regions in the Gag layer. The classification was based on the parts of the plots containing the density bound to the inner face of the membrane (shaded area in Fig. 7C). Data were recorded on resin embeddings of HIV-1 infected MT4 cells, HeLa cells transfected with a proviral HIV-1 plasmid as well as positive and negative controls for the presence of the two lattice types (Table 2). In order to avoid overrepresentation of single cells, two tilt series were recorded per cell, and they were recorded as far apart as possible in the section plane (usually >5 µm). The same cell was never studied in consecutive sections. Our published data on infected MT4 cells [26] were also included in the analysis (Table 2, data set 8).

Principal component analysis of the variance in the data set revealed the presence of one major cluster with residual variance and a smaller, more homogeneous one (Fig. S3A–D in Supporting Information S1). Hierarchical clustering of the cropped line plots identified the two main classes as the two clusters observed in the PCA representation (Fig. S3E,F in Supporting Information S1). The less populated class (class 1, dashed line in Fig. 7D) lacked the innermost density peak corresponding to NC-RNA, whereas this peak was present in the more populated class (class 2, solid line in Fig. 7D). Further division of class 2 into subclasses mainly revealed a slight variation in the position and relative intensity of the NC-RNA peak (Fig. S4 in Supporting Information S1).
Table 3 shows the distribution of budding sites into the two classes for each sample. Class 1 contained 18% of the budding sites on HIV-1 infected MT4 cells and 2.9% of the budding sites on transfected HeLa cells. For infected MT4 cells treated with the PR inhibitor lopinavir, no class 1 budding sites were observed. As a control for the thinner Gag lattice, released virions carrying uncleaved MA-SP1 were studied. In this case, Gag processing is stalled at the MACASP1 fragment due to inactivating mutations at cleavage sites between MA and CA and CA and SP1, respectively (see [10] and accompanying paper by de Marco et al.). These particles generally fell into class 1. Taken together, these results suggest that class 1 corresponds to the newly described lattice detected by cET, while class 2 corresponds to the immature lattice.

Radius and closure of the Gag layer was calculated separately for the two classes (Table 4, Fig. 7F). Whereas the average radii of the two classes were similar, the closure of the Gag layer was significantly higher for the class 1 buds (Wilcoxon rank sum test, p = 0.0021). Assuming the same lattice packing in both classes, class 1 budding sites in this data set contain 5,000±1,300 Gag molecules while class 2 budding sites in this data set contain 3,300±1,500 Gag molecules.

Finally, we asked whether individual cells carried only one class of budding sites or both. In the case of HIV-1 infected MT4 cells, 47 cells exhibited more than one budding site in the tomogram (Table 2, data set 4). Fig. 7E (black bars) shows the relative frequency of class 1 budding sites on these 47 cells, overlayed with the theoretical distribution (grey circles) that would be expected if all cells had the average of 18% class 1 budding sites. Clearly, most cells had either only class 1 or only class 2 budding sites, indicating that the budding phenotype is determined at the single cell level.
Discussion
Here, we established a system to visualize HIV-1 virus-like particle assembly and release in situ at macromolecular resolution in three dimensions using cET. This method permits structural interpretation of assembly sites and their cytoplasmic and membrane surroundings without the limitations imposed by the fixation, staining and dehydration methods commonly used in cellular electron microscopy. It allowed a detailed analysis of the Gag lattice in cellular budding sites, suggesting that the organization of the immature HIV-1 particle is already determined at the point of its intracellular assembly, and further identified a hitherto undescribed Gag lattice form. We show that this Gag lattice form is predominant in a subset of infected T-cells, and hypothesize that it is the result of premature PR activation in these cells.
The cET analysis of HIV-1 Gag and Gag-Pol expressing cells also provided unprecedented snapshots of cortical actin filaments in the vicinity of viral budding sites (Fig. 2, 3). It is well established that HIV-1 particles contain substantial amounts of actin [31], but the interplay between cortical actin and retrovirus assembly is still unknown. A recent study indicated a role for star-shaped actin filament arrangements emanating from the viral budding sites in HIV-1 assembly [32]. Our cET data provide 3D structures suggesting directed arrangement of actin filaments towards some budding sites, particularly in filopodia-associated buds. Strikingly, half of the budding sites in the current data set were present on actin-filled filopodia. To what extent this is a consequence of the particular region of the cell accessible to cET remains to be determined. Future cET studies using altered Gag constructs and selective manipulation of the actin dynamics will provide the necessary tools to untangle the molecular details of the involvement of actin at HIV-1 budding sites.
Extracellular immature HIV-1 particles are the direct products of viral budding at the plasma membrane, making it likely that their Gag protein lattice is determined during assembly. The observation that the Gag lattice is incomplete in the immature virus [22], [26] and contains defects [19], [22] raised the alternative hypothesis that Gag rearrangements occur following release. This could involve post-release association of smaller patches of Gag hexamers, or post-release symmetry breakage and dissociation of a hexagonal lattice containing evenly spaced pentameric defects [22]. Analysis of the lattice in extracellular immature virions revealed continuous hexameric symmetry over most of the lattice with curvature induced by defects of irregular size and shape [19]. These studies were performed on purified released particles, however, and the degree of order attained during assembly at the plasma membrane may have been higher (and broken after release) or lower (and subsequently assembled into a more ordered lattice). Our cET data of budding sites revealed structures of the immature Gag lattice in intact unperturbed cells prior to release, allowing us to draw conclusions on the early steps of the HIV-1 assembly process. There was no indication of separated “islands” of hexameric Gag in the in situ budding sites; upon visual inspection they consistently contained one continuous layer of Gag protein. A more detailed analysis of the lattice in the immature budding sites revealed that its unit cell structure was highly similar to that previously described for released immature virions [19], and included irregular lattice defects of similar size and distribution. This suggests that the organization of the immature HIV-1 particle is indeed determined at the point of its intracellular assembly, and not as a result of large scale post-release ordering or disordering.
Besides the described immature Gag lattice, we observed a previously undescribed lattice type in extracellular particles and in budding sites still connected to the cell. Formation of this lattice was dependent on an active HIV-1 PR. The newly discovered lattice exhibited hexagonal symmetry with a CA layer identical to the immature lattice, but lacked the second layer of density corresponding to the NC-RNA complex. The particles and budding sites also contained no visible NC-RNA condensate in the center of the particles or adjacent to the buds, suggesting that the viral genome is absent. A comparison with the analysis performed in the accompanying paper by de Marco et al. on isolated virions with specific Gag cleavage site mutations showed that the newly described lattice is most likely composed of the MA-SP1 region of HIV-1 Gag. Although the tomographic average at current resolution does not allow identification of the density corresponding to the SP1 peptide, de Marco et al. showed that this peptide is present, independent of whether the cleavage site between CA and SP1 is intact or not. Notably, the particles produced from the MA-CA and MA-SP1 cleavage site mutants which exhibit an MA-SP1 type lattice do contain a clearly distinguishable density corresponding to condensed NC-RNA (de Marco et al.), which distinguishes them from the particles with this lattice type in the current study.
As further described in the accompanying paper by de Marco et al., the MACASP1 lattice is likely to constitute a natural intermediate in the proteolytic maturation of HIV-1. Cleavage between SP1 and NC is the fastest processing step in vitro and abscission of the C-terminal NC-p6 regions from the immature lattice is required for condensation of the inner ribonucleoprotein complex of the virion prior to formation of the capsid. Thus, extracellular particles containing the MACASP1 lattice and inner density corresponding to NC-RNA are suggested to represent a “frozen” intermediate in HIV-1 maturation (de Marco et al.). Such particles were not observed for wild-type HIV-1 constructs, however, indicating that maturation intermediates are normally short-lived. An MACASP1 lattice without adjacent NC-RNA density was observed in 18% of HIV-1 budding sites on infected MT-4 cells, on the other hand, and its frequent presence suggests that it is a metastable structure with a significant lifetime in this case.
We previously reported that budding sites on infected MT4 cells have a broad distribution of Gag layer morphologies. Some resemble actual released immature virions, but some have a more complete Gag layer, containing more Gag molecules than the released virions and thus rather resemble arrested “late” budding sites not thought to be virus precursors [26]. When re-evaluating these data with respect to the type of Gag lattice, the immature lattice was predominantly found in budding sites resembling released virions, whereas the budding sites with the novel lattice had a significantly more closed Gag layer than released virions (Table 4, Fig. 7F). In the case of late-domain defective variants, formation of a closed layer is thought to be caused by a failure to recruit the ESCRT machinery and thus to drive the release process. Consequently, Gag assembly continues until reaching an equilibrium structure, but the resulting late budding sites are mostly dead-end products. It appears likely that a similar process is also relevant for the budding sites with the novel lattice. We hypothesize that premature proteolytic activation prior to confinement of virion constituents in the budding virion leads to removal of NC-p6 from Gag and of PR and downstream pol products from Gag-Pol. The cleaved C-terminal fragments are lost from the assembly site, while the MACASP1 lattice is stably retained. No ESCRT recruitment occurs since the p6 domain has been removed and no further processing occurs because PR, not being confined in a viral particle, has been lost. Accordingly, no budding structures with mature cores were observed. Further assembly of the truncated lattice may involve additional Gag molecules undergoing partial processing or addition of cytoplasmic MACASP1 molecules since a truncated MACASP1 construct has been shown to form budlike structures when overexpressed [33], [34]. The presence of extracellular particles with the MACASP1 lattice and lacking NC-RNA may indicate some ESCRT-independent release. Release of virions lacking most of the NC domain (but in this case carrying the p6 domain) has also been observed in previous studies [35], [36].
The results of the current study clearly show that morphological maturation can be initiated inside the producer cell, but indicate that it is largely non-productive if the virion constituents are not confined in a structure sequestered from the cytoplasm. Assembly, release and maturation would therefore require the sequential processes of Gag association, ESCRT recruitment (prior to closure of the Gag shell), and PR activation (after or concomitant with ESCRT-mediated sequestration of the bud). The observation of aberrant, apparently dead-end structures in a subset of cells in HIV-1 infected T-cell lines suggests that the control of these processes can be lost, however. Infected MT-4 cells generally contained only one type of budding sites indicating that this loss of control may be caused by the host cell environment. Conceivably, host cell factors may be involved in regulating the kinetics of the consecutive stages of viral assembly, budding and release, and the subset of T-cells may have lost this regulation, although a general cytopathic effect cannot be ruled out at present. Identifying viral or host factors involved in this regulation will be of great importance for our understanding of HIV-1 morphogenesis, and may also lead to new approaches to render HIV-1 infection non-productive.
Materials and Methods
Construction of adenoviral vectors for HIV-1 Gag and GagPol
Adenoviral vectors for HIV-1 Gag (AdGag) and GagPol (AdGagPol) were constructed from the Rev independent versions of these genes [29], using the BD Adeno-X Expression System 1 (BD Biosciences Clontech). Briefly, the 1539 nt gag-gene was amplified by PCR adding flanking NheI/XbaI sites and ligated into the transfer vector pShuttle2. The gag-pol gene was excised with NdeI/XbaI from pcDNA3.1syngagpol [29] and directly cloned into pShuttle2. After verification of the insert by sequence analysis, these plasmids were digested with PI-Sce/I-Ceu. The inserts were ligated into the pre-digested Adeno.X viral DNA. Resulting plasmids were verified by restriction digest and DNA sequencing. Recombinant adenoviruses were generated by transfection of the AdGag or Ad GagPol plasmids into HEK 293 cells and amplified according to the protocol supplied with the kit. Titers of adenoviral vectors were determined using the Adeno-X rapid Titer Kit (Clontech); titers of both AdGag and Ad GagPol typically reached 1–2×109/ml. Immunofluorescence staining of AdGag and AdGagPol transduced cells for HIV-1 Gag was performed using a rabbit polyclonal anti-CA antibody and a FITC-conjugated goat-anti-rabbit secondary antibody. Western blots of cell lysates and particles purified from the culture media of AdGag and AdGagPol transduced cells were performed using the same polyclonal anti-CA antibody, with an IRDye 800CW goat anti-rabbit secondary antibody (LI-COR Biosciences) as recommended by the manufacturer.
Cell culture and adenoviral vector transductions for cryo electron tomography
Human glioblastoma cell lines U-87 MG and U-373 MG were maintained in DMEM and MEM, respectively, supplemented with Penicillin-Streptomycin and 10% foetal bovine serum. When grown on EM grids (CF-2/1-2AU, Protochips, inc.), cells were seeded at low density and kept in medium with 0.5% serum which increased the extent of flat areas of the cells. Transduction with AdGag or AdGagPol was performed concomitant with or one day after seeding cells on EM grids. A multiplicity of ‘infection’ (MOI) of 10 was typically used, calculated based on cell culture dish area and assuming that added vector will adhere to any substrate surface. One to three days after seeding cells on the grids, they were plunge-frozen into liquid ethane after application of 10 nm colloidal gold.
Sample preparation for electron tomography of sections of resin-embedded cells
Cell culture and preparation for sections of resin embedded cells were performed as previously described [26]. For production of immature virus, cells were grown in the presence of 1 µM lopinavir. For production of particles containing an uncleaved MA-SP1 layer, HEK293T cells were transfected with a proviral HIV-1NL4-3 plasmid carrying mutations in the cleavage sites between MA and CA and CA and SP1, respectively ([14] and accompanying paper by de Marco et al.). Cells were fixed, stained, dehydrated and EPON-embedded, and 300 nm sections of the embeddings were cut and transferred to EM grids after deposition of colloidal gold onto the EM grid support film.
Electron tomography
Cryo electron tomography was performed with a Philips CM300 or a FEI Tecnai F30 Polara transmission electron microscope (FEI; Eindhoven, The Netherlands), both equipped with 300 kV field emission guns, Gatan GIF 2002 post-column energy filters and 2048×2048 Multiscan CCD cameras (Gatan; Pleasanton, CA). All data collection was performed at 300 kV, with the energy filter operated in zero loss mode. Tilt series of cryo specimens were typically recorded from −60° to +60° with an angular increment of 1.5°, a total electron dose of 80 electrons Å−2, and a defocus of −6.0 µm to −8.0 µm. The pixel sizes at the specimen level were 8.21 Å at the CM300, and 8.05 Å or 7.13 Å at the FEI Tecnai F30 Polara, respectively. Electron tomography of resin sections was performed at an FEI Tecnai F20 transmission electron microscope (FEI; Eindhoven, The Netherlands), equipped with 200 kV field emission gun and a 4096×4096 Eagle CCD camera (FEI; Eindhoven, The Netherlands). Tilt series on resin sections were recorded at room temperature from −60° to +60° with an angular increment of 1.0°, with a one time binned pixel size of 2*3.72 Å and a defocus of -4.0 µm. For both cryo and room temperature tilt series, the applied electron dose for a given tilt angle α was proportional to 1/cos(α) to compensate for the higher effective specimen thickness at higher tilts. At the Tecnai microscopes, the SerialEM acquisition software [37] was used for tilt series acquisition. Three-dimensional reconstructions from tilt series were calculated using the weighted back-projection method [38], as implemented in the TOM toolbox [39] for MATLAB (Mathworks, Natick, Massachusetts, United States) with the “exact weighting” option, or in IMOD [40].
Subtomogram averaging
Subtomogram averaging was performed using MATLAB. Sub-volumes of (38.6 nm)3 were extracted along the surface of the budding particles. The subtomograms were then iteratively aligned against the average in a reference free manner, while applying 6-fold symmetry as described in the accompanying article (de Marco et al). For individual particles, the distribution of the cross correlation values between the aligned subtomograms and the reference was bimodal. The threshold of the subtomograms that went into the reconstruction was set as minimum between the two peaks.
Classification of Gag lattice types in tomograms of resin-embedded sections
The classification of Gag densities in resin embeddings of HIV-1 budding sites was performed as described in the Supplementary Information. In brief, linear density profiles were calculated orthogonally to the plasma membrane in the budding sites. These one-dimensional density profiles were cropped to contain only the membrane-apposed density, and subjected to hierarchical clustering.
Supporting Information
Zdroje
1. FreedEO
1998 HIV-1 gag proteins: diverse functions in the virus life cycle. Virology 251 1 15
2. HurleyJH
2008 ESCRT complexes and the biogenesis of multivesicular bodies. Curr Opin Cell Biol 20 4 11
3. MoritaE
SundquistWI
2004 Retrovirus budding. Ann Rev Cell Dev Biol 20 395 425
4. WilliamsRL
UrbeS
2007 The emerging shape of the ESCRT machinery. Nat Rev Mol Cell Biol 8 355 368
5. BriggsJAG
SimonMN
GrossI
KräusslichHG
FullerSD
2004 The stoichiometry of Gag protein in HIV-1. Nat Struct Mol Biol 11 672 675
6. GarrusJE
von SchwedlerUK
PornillosOW
MorhamSG
ZavitzKH
2001 Tsg101 and the vacuolar protein sorting pathway are essential for HIV-1 budding. Cell 107 55 65
7. VerPlankL
BouamrF
LaGrassaTJ
AgrestaB
KikonyogoA
2001 Tsg101, a homologue of ubiquitin-conjugating (E2) enzymes, binds the L domain in HIV type 1 Pr55(Gag). Proc Natl Acad Sci U S A 98 7724 7729
8. CoffinJM
HughesSH
VarmusHE
1997 Retroviruses: Cold Spring Harbor Laboratory Press.
9. PettitSC
MoodyMD
WehbieRS
KaplanAH
NantermetPV
1994 The p2 domain of human immunodeficiency virus type 1 Gag regulates sequential proteolytic processing and is required to produce fully infectious virions. J Virol 68 8017 8027
10. WiegersK
RutterG
KottlerH
TessmerU
HohenbergH
1998 Sequential steps in human immunodeficiency virus particle maturation revealed by alterations of individual Gag polyprotein cleavage sites. J Virol 72 2846 2854
11. CorenLV
ThomasJA
ChertovaE
SowderRC
GagliardiTD
2007 Mutational analysis of the C-terminal Gag cleavage sites in human immunodeficiency virus type 1. J Virol 81 10047 10054
12. CheckleyMA
LuttgeBG
SoheilianF
NagashimaK
FreedEO
2010 The capsid-spacer peptide 1 Gag processing intermediate is a dominant-negative inhibitor of HIV-1 maturation. Virology 400 137 144
13. LeeSK
HarrisJ
SwanstromR
2009 A strongly transdominant mutation in the human immunodeficiency virus type 1 gag gene defines an Achilles heel in the virus life cycle. J Virol 83 8536 8543
14. MüllerB
AndersM
AkiyamaH
WelschS
GlassB
2009 HIV-1 Gag processing intermediates trans-dominantly interfere with HIV-1 infectivity. J Biol Chem 284 29692 29703
15. KräusslichHG
1991 Human immunodeficiency virus proteinase dimer as component of the viral polyprotein prevents particle assembly and viral infectivity. Proc Natl Acad Sci U S A 88 3213 3217
16. KaplanAH
ZackJA
KniggeM
PaulDA
KempfDJ
1993 Partial inhibition of the human immunodeficiency virus type 1 protease results in aberrant virus assembly and the formation of noninfectious particles. J Virol 67 4050 4055
17. MooreMD
FuW
SoheilianF
NagashimaK
PtakRG
2008 Suboptimal inhibition of protease activity in human immunodeficiency virus type 1: effects on virion morphogenesis and RNA maturation. Virology 379 152 160
18. BenjaminJ
Ganser-PornillosBK
TivolWF
SundquistWI
JensenGJ
2005 Three-dimensional structure of HIV-1 virus-like particles by electron cryotomography. J Mol Biol 346 577 588
19. BriggsJA
RichesJD
GlassB
BartonovaV
ZanettiG
2009 Structure and assembly of immature HIV. Proc Natl Acad Sci U S A 106 11090 11095
20. BriggsJAG
GrünewaldK
GlassB
FörsterF
KräusslichHG
2006 The mechanism of HIV-1 core assembly: Insights from three-dimensional reconstructions of authentic virions. Structure 14 15 20
21. LiuJ
BartesaghiA
BorgniaMJ
SapiroG
SubramaniamS
2008 Molecular architecture of native HIV-1 gp120 trimers. Nature 455 109 113
22. WrightER
SchoolerJB
DingHJ
KiefferC
FillmoreC
2007 Electron cryotomography of immature HIV-1 virions reveals the structure of the CA and SP1 Gag shells. EMBO J 26 2218 2226
23. ZhuP
LiuJ
BessJJr
ChertovaE
LifsonJD
2006 Distribution and three-dimensional structure of AIDS virus envelope spikes. Nature 441 847 852
24. ZhuP
WinklerH
ChertovaE
TaylorKA
RouxKH
2008 Cryoelectron tomography of HIV-1 envelope spikes: further evidence for tripod-like legs. PLoS Pathog 4 e1000203
25. BriggsJAG
WilkT
WelkerR
KräusslichHG
FullerSD
2003 Structural organization of authentic, mature HIV-1 virions and cores. EMBO J 22 1707 1715
26. CarlsonLA
BriggsJA
GlassB
RichesJD
SimonMN
2008 Three-dimensional analysis of budding sites and released virus suggests a revised model for HIV-1 morphogenesis. Cell Host Microbe 4 592 599
27. LiS
HillCP
SundquistWI
FinchJT
2000 Image reconstructions of helical assemblies of the HIV-1CA protein. Nature 407 409 413
28. GanserBK
LiS
KlishkoVY
FinchJT
SundquistWI
1999 Assembly and analysis of conical models for the HIV-1 core. Science 283 80 83
29. WagnerR
GrafM
BielerK
WolfH
GrunwaldT
2000 Rev-independent expression of synthetic gag-pol genes of human immunodeficiency virus type 1 and simian immunodeficiency virus: implications for the safety of lentiviral vectors. Hum Gene Ther 11 2403 2413
30. MedaliaO
WeberI
FrangakisAS
NicastroD
GerischG
2002 Macromolecular architecture in eukaryotic cells visualized by cryoelectron tomography. Science 298 1209 1213
31. OttDE
CorenLV
KaneBP
BuschLK
JohnsonDG
1996 Cytoskeletal proteins inside human immunodeficiency virus type 1 virions. J Virol 70 7734 7743
32. GladnikoffM
ShimoniE
GovNS
RoussoI
2009 Retroviral assembly and budding occur through an actin-driven mechanism. Biophys J 97 2419 2428
33. CristRM
DattaSA
StephenAG
SoheilianF
MirroJ
2009 Assembly properties of human immunodeficiency virus type 1 Gag-leucine zipper chimeras: implications for retrovirus assembly. J Virol 83 2216 2225
34. HockleyDJ
NermutMV
GriefC
JowettJB
JonesIM
1994 Comparative morphology of Gag protein structures produced by mutants of the gag gene of human immunodeficiency virus type 1. J Gen Virol 75 Pt 11 2985 2997
35. OttDE
CorenLV
ChertovaEN
GagliardiTD
NagashimaK
2003 Elimination of protease activity restores efficient virion production to a human immunodeficiency virus type 1 nucleocapsid deletion mutant. J Virol 77 5547 5556
36. PopovaE
PopovS
GottlingerHG
2010 Human Immunodeficiency Virus Type 1 Nucleocapsid p1 Confers ESCRT Pathway Dependence. J Virol 84 6590 6597
37. MastronardeDN
2005 Automated electron microscope tomography using robust prediction of specimen movements. J Struct Biol 152 36 51
38. FrankJ
2006 Electron Tomography: Methods for Three-Dimensional Visualization of Structures in the Cell Berlin: Springer
39. NickellS
FörsterF
LinaroudisA
Del NetW
BeekF
2005 TOM software toolbox: acquisition and analysis for electron tomography. Journal of Structural Biology 149 227 234
40. KremerJR
MastronardeDN
McIntoshJR
1996 Computer visualization of three-dimensional image data using IMOD. J Struct Biol 116 71 76
41. WestfallPH
YoungSS
1993 Resampling-based multiple testing: examples and methods for P-value adjustment: Wiley-Interscience 360
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2010 Číslo 11
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- Zn Inhibits Coronavirus and Arterivirus RNA Polymerase Activity and Zinc Ionophores Block the Replication of These Viruses in Cell Culture
- The Female Lower Genital Tract Is a Privileged Compartment with IL-10 Producing Dendritic Cells and Poor Th1 Immunity following Infection
- Crystal Structure and Size-Dependent Neutralization Properties of HK20, a Human Monoclonal Antibody Binding to the Highly Conserved Heptad Repeat 1 of gp41
- The Arabidopsis Resistance-Like Gene Is Activated by Mutations in and Contributes to Resistance to the Bacterial Effector AvrRps4
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy