
Foxn1 Regulates Lineage Progression in Cortical and Medullary Thymic Epithelial Cells But Is Dispensable for Medullary Sublineage Divergence
The forkhead transcription factor Foxn1 is indispensable for thymus development, but the mechanisms by which it mediates thymic epithelial cell (TEC) development are poorly understood. To examine the cellular and molecular basis of Foxn1 function, we generated a novel and revertible hypomorphic allele of Foxn1. By varying levels of its expression, we identified a number of features of the Foxn1 system. Here we show that Foxn1 is a powerful regulator of TEC differentiation that is required at multiple intermediate stages of TE lineage development in the fetal and adult thymus. We find no evidence for a role for Foxn1 in TEC fate-choice. Rather, we show it is required for stable entry into both the cortical and medullary TEC differentiation programmes and subsequently is needed at increasing dosage for progression through successive differentiation states in both cortical and medullary TEC. We further demonstrate regulation by Foxn1 of a suite of genes with diverse roles in thymus development and/or function, suggesting it acts as a master regulator of the core thymic epithelial programme rather than regulating a particular aspect of TEC biology. Overall, our data establish a genetics-based model of cellular hierarchies in the TE lineage and provide mechanistic insight relating titration of a single transcription factor to control of lineage progression. Our novel revertible hypomorph system may be similarly applied to analyzing other regulators of development.
Published in the journal:
. PLoS Genet 7(11): e32767. doi:10.1371/journal.pgen.1002348
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002348
Summary
The forkhead transcription factor Foxn1 is indispensable for thymus development, but the mechanisms by which it mediates thymic epithelial cell (TEC) development are poorly understood. To examine the cellular and molecular basis of Foxn1 function, we generated a novel and revertible hypomorphic allele of Foxn1. By varying levels of its expression, we identified a number of features of the Foxn1 system. Here we show that Foxn1 is a powerful regulator of TEC differentiation that is required at multiple intermediate stages of TE lineage development in the fetal and adult thymus. We find no evidence for a role for Foxn1 in TEC fate-choice. Rather, we show it is required for stable entry into both the cortical and medullary TEC differentiation programmes and subsequently is needed at increasing dosage for progression through successive differentiation states in both cortical and medullary TEC. We further demonstrate regulation by Foxn1 of a suite of genes with diverse roles in thymus development and/or function, suggesting it acts as a master regulator of the core thymic epithelial programme rather than regulating a particular aspect of TEC biology. Overall, our data establish a genetics-based model of cellular hierarchies in the TE lineage and provide mechanistic insight relating titration of a single transcription factor to control of lineage progression. Our novel revertible hypomorph system may be similarly applied to analyzing other regulators of development.
Introduction
T cell development occurs in the thymus and depends on progressive interactions with the thymic stroma. Critical to this function is the thymic epithelium (TE), which comprises a diverse array of phenotypically and functionally distinct cell types broadly organized into cortical and medullary regions [1].
During development the thymus arises from the endoderm of the third pharyngeal pouches [2] which, from day 9 of embryonic development in the mouse (E9.0), contain cells specified to the TE lineage [2]. Until E11.5, this region uniformly expresses the cell surface determinant Plet-1 [3]-[5] and the earliest currently identified founder cells for the lineage are thus Plet-1+ third pharyngeal pouch cells. By E12.5 the thymic primordia, now colonized by haematopoietic progenitor cells and surrounded by a mesenchymal capsule, have separated from the pharyngeal endoderm and contain a predominant EpCam+Plet1+ epithelial population [3], [6], [7]. Recent evidence supports the existence within this population of a common thymic epithelial progenitor cell (TEPC) capable of generating both cortical and medullary TEC (cTEC and mTEC respectively) [7], [8]. Further studies indicate that a medullary sub-lineage specific progenitor cell is present at least as early as E13.5 [9], [10], and suggest the existence of a cortical sub-lineage specific progenitor [11]. However, the extent to which these progenitor activities persist in late organogenesis and the postnatal organ has not been determined and furthermore, the timing of emergence of the sub-lineage progenitors during organogenesis remains unclear.
The forkhead transcription factor Foxn1 [12], the gene mutated in the classical mouse mutant nude (nu) [13], is a pivotal regulator of TE lineage development. Nude mice are hairless and athymic [14], [15]. In Foxn1 null animals, the earliest stages of thymus development appear to occur normally, but development is arrested after initial formation of the organ primordium (around E12.0 in the mouse) and the primordium is not colonized by hematopoietic precursors [16]. Nonetheless, the Foxn1 null thymic primordium separates from the parathyroid component of the common primordium and migrates to the mid-line, where a small, alymphoid cystic thymic remnant persists postnatally [14]. Null alleles of Foxn1 in mouse, rat and human all exhibit essentially the same pleiotropic phenotype [17].
Forkhead family members play important roles in specification and progression of a number of cell lineages, and recent evidence reveals roles for some Forkhead proteins in chromatin modification/epigenetic regulation of cell fate [18]. Analysis of postnatal nude:wild-type chimeric mice demonstrated the cell autonomous requirement for Foxn1 in development of all major TEC sub-lineages and suggested that TE lineage cells unable to express functional Foxn1 might undergo maturational arrest at a common TEPC stage [19]. A revertible Foxn1 null model, Foxn1SA2/SA2 provided strong support for this hypothesis, as neonatal clonal reversion of this allele resulted in the generation of small units of functional thymus tissue containing both cortical and medullary compartments [8]. Further support for a role in differentiation comes from studies on keratinocytes, which implicate Foxn1 in regulating initiation of terminal differentiation [20], [21], and from analysis of a hypomorphic Foxn1 allele which generates a transcript lacking exon 3 and thus the N-terminal domain of Foxn1 [22]. In mice homozygous for this allele (Foxn1Δ), the postnatal Foxn1Δ/Δ thymus was highly cystic, contained no discernable cortical or medullary regions and could sustain only highly impaired thymocyte differentiation, suggesting that Foxn1 is actively required for TEC differentiation at stages beyond initiation of the TEC programme [22]. Evidence also supports roles for Foxn1 in TEC proliferation [23] and in regulating the balance between proliferation and differentiation in skin [24]. In addition, a requirement for Foxn1 for maintenance of the postnatal thymic microenvironment has recently been demonstrated [25]–[27], with evidence pointing to differential sensitivity of different TEC subsets to changes in Foxn1 dosage [26]. Collectively, these studies suggest that Foxn1 plays a complex role in regulating TEC lineage development. However, precisely how Foxn1 regulates the transit from the earliest fetal thymic epithelial progenitor cell to the fully functional postnatal thymic epithelium, and at which stages in this process it is required, remains undetermined. In addition, the molecular mechanisms regulated by this transcription factor in the thymus have not yet been addressed.
In this study, we have addressed the functions of Foxn1 throughout thymus ontogeny, via generation and analysis of a novel revertible hypomorphic allele of Foxn1, Foxn1R, which expresses only low levels of Foxn1 mRNA and protein. A particular advantage of our system is the revertible nature of the allele, which affords the capacity to test the relationship of cell states identified via analysis of mutant mice to states occurring in normal ontogeny. Our studies establish Foxn1 as a powerful regulator of differentiation in both the cTEC and mTEC sub-lineages. We find no evidence for a role for Foxn1 in regulating cell fate choice in the cortical or medullary TEC sub-lineages. Rather, we find that Foxn1 is required for progression of differentiation at multiple stages in cTEC and mTEC sub-lineage development in both the fetal and postnatal thymus, and show that different Foxn1 dosage is required to execute its function(s) at different differentiation stages. We further establish that Foxn1 regulates, either directly or indirectly, a suite of genes known to effect TEC function - including Dll4, CCL25, Cathepsin L, CD40, Pax1, and MHC Class II, and that these exhibit different response patterns to changes in Foxn1 dosage. Collectively, these findings significantly advance understanding of the role of Foxn1 in TEC, demonstrating for the first time its direct involvement in generating multiple TEC sub-types in both the cortical and medullary lineages in the fetal and postnatal thymus, and suggesting it acts effectively as a master regulator of the entire TEC programme.
Results
Generation of a revertible hypomorphic allele of Foxn1
We set out to generate a conditionally revertible null allele of Foxn1 as a tool for generating TEPC lines. Our rationale was that lack of Foxn1 expression would impose an early block on TEC lineage differentiation, effectively trapping TEC in an undifferentiated progenitor cell state, while reversion of the allele would remove this block and allow progression to terminal differentiation. Since the extent to which Foxn1 is required for TEC proliferation is unknown, SV40 T antigen was used to uncouple potential roles of Foxn1 in proliferation and differentiation. We thus generated the revertible Foxn1 allele, Foxn1R, by inserting a LoxP flanked cassette into intron 1b of the Foxn1 locus by homologous recombination in ES cells (Figure 1A, Figure S1) and used this ES line to generate the Foxn1R mouse strain. Initial characterization of postnatal Foxn1R/+ mice revealed thymus hyperplasia as expected, due to expression of SV40 Tag under the Foxn1 promoter. However, Foxn1R/R mice developed severely hypoplastic thymi rather than exhibiting the expected phenotype of complete thymic aplasia (Figure 1B). In keeping with this, immunoblotting for Foxn1 revealed low-level Foxn1 protein in Foxn1R/R thymi compared to wild-type (WT; Figure 1C) and RT-PCR analyses demonstrated that the transcripts produced from the Foxn1R allele contained either Exon1a-SV40Tag-IRES-eGFPneo elements or the full-length Foxn1 mRNA (Figure 1D). Thus some residual Foxn1 expression from Foxn1R occurred, and resulted from splicing around the targeted insertion subsequent to transcription proceeding beyond the transcriptional pause. No overt skin phenotypes were apparent in either Foxn1R/+ or Foxn1R/R mice. Collectively, these data establish that Foxn1R is a hypomorphic allele that expresses a low level of Foxn1 compared to WT and that expression of SV40Tag does not compensate for decreased levels of Foxn1.

The revertibility of the allele was demonstrated by crossing Foxn1R/+ mice with the ZP3Cre deletor strain, in which Cre recombinase is expressed in oocytes [28]. Foxn1R/+ZP3Cre/+ mice excised the LoxP flanked cassette in most if not all cells (Figure 1E), and showed normal thymus development and function.
SV40 T antigen does not affect TEC differentiation or function in Foxn1R/+ thymi
The Foxn1 null phenotype of severe athymia results from a complete early block in thymus development and has limited understanding of the role of Foxn1 later in ontogeny. We reasoned that the Foxn1R allele might be informative in this light. However, it was first necessary to test whether expression of SV40Tag under the Foxn1 promoter affected TEC differentiation or function. Therefore, we characterized the Foxn1R/+ thymus phenotype, in particular seeking evidence for impaired or perturbed TEC development and/or function. The two thymic lobes in Foxn1R/+ mice exhibited a greater than ten-fold increase in both TEC and thymocyte numbers compared to WT littermates (Figure 1), but were otherwise histologically normal (Figure 1B, 1F). In both fetal and postnatal Foxn1R/+ thymi, TEC and thymocyte subset distributions were indistinguishable from Foxn1nu/+ and WT littermates and no evidence of malignant transformation could be found (Figure 1G-1I). We therefore concluded that the thymic hyperplasia observed resulted from a uniform expansion of the TEC compartment due to expression of SV40Tag under control of the Foxn1 promoter, but that this did not affect TEC differentiation or function in either the fetal or postnatal thymus. This is consistent with the action of SV40TAg in a variety of other tissues [29]–[31] and its known function in regulating cell proliferation. Of note is that the Foxn1 promoter drives relatively low levels of gene expression in TEC, based on QRT-PCR analyses relative to a range of housekeeping genes. This demonstrated that Foxn1 is expressed in the same range as alpha-tubulin, which is recognized to be expressed at low levels [32] (Figure S2). Furthermore, SV40Tag was not detectable by immunohistochemical analysis of thymus sections from Foxn1R mutant mice (Figure S2).
Terminal differentiation of postnatal cortical and medullary TEC is blocked in Foxn1R/R mice
Next, we tested the effect of reduced Foxn1 dosage on postnatal TEC by analyzing the Foxn1R/R phenotype. Although small, Foxn1R/R thymi were grossly histologically normal, containing clear cortical and medullary regions (Figure 2A, 2B). Postnatal Foxn1R/R thymi contained all normal thymocyte subsets (Figure 2C, 2D). However, the CD4+8+ double positive (DP) subset was proportionally larger than in WT littermates while the CD4+ and CD8+ single positive (SP) subsets were proportionally reduced (DP proportions: Foxn1R/R, 91.2±0.85%; WT 82.9±0.21%, p = 0.0004) suggesting that the DP to SP transition and/or subsequent thymocyte maturation was perturbed in Foxn1R/R mice. The CD4-8- (Double negative, DN) cell subset distribution was also altered in postnatal Foxn1R/R thymi, with increased proportions of CD44+CD25- (DN1) and CD44-CD25+ (DN3) cells (Figure 2D). Most DN1 cells in postnatal Foxn1R/R thymi were B cells, as shown by B220 staining (Figure 2E), suggesting impaired commitment of hematopoietic progenitors to the T cell lineage [33], [34]. Furthermore, thymocyte development was delayed in Foxn1R/R mice early in ontogeny (Figure 2F). Thus, rather than simply resulting from reduced numbers of functionally normal TECs, the hypoplastic Foxn1R/R phenotype was characterized by impaired TEC functionality.

Interrogation of TEC subset distribution and marker expression in the Foxn1R/R thymus revealed apparently normal morphologies for medullary and cortical TEC (mTEC and cTEC respectively), and normal expression patterns for cytokeratin (K) 5, K8 and CD205 (Figure 2B, Figure 3A and 3Ai), indicating the presence of cortical and medullary compartments. Further analysis however revealed abnormal gene expression patterns in both compartments.
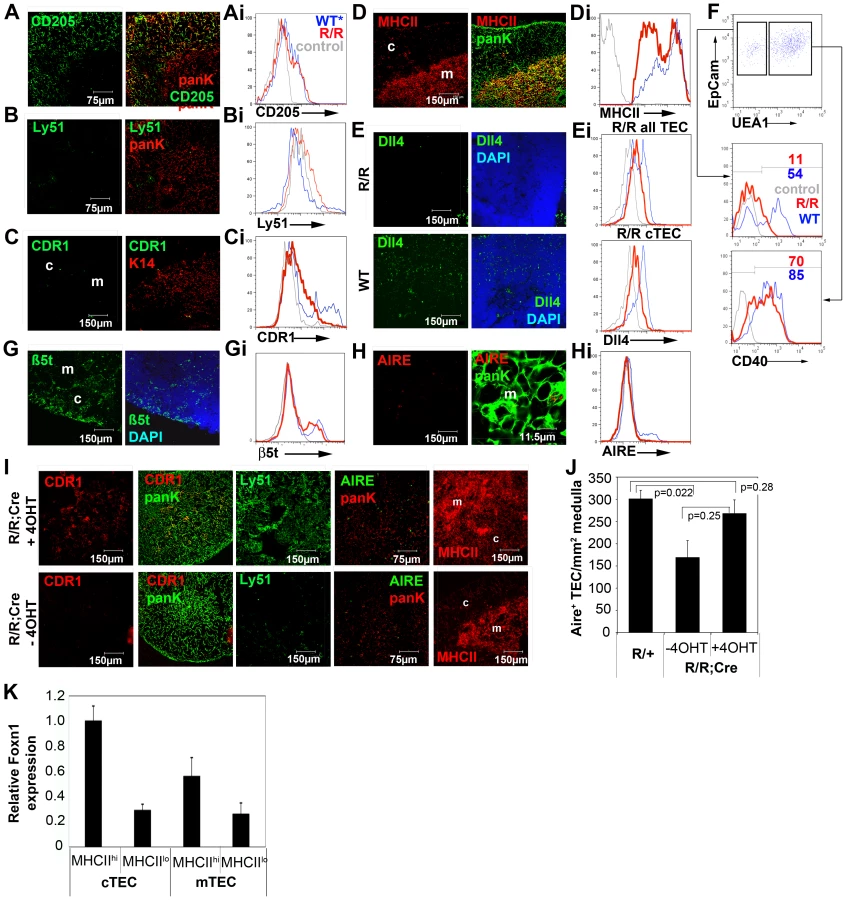
The cTEC marker CDR1 is acquired only postnatally in cTEC and thus represents a terminal differentiation marker for this TEC sub-lineage. Ly51 is another commonly used marker that is cTEC-specific within the thymus. Expression of both of these markers was severely down-regulated in Foxn1R/R thymi (Figure 3B-3Ci).
MHC class II, Cathepsin L and Dll4 are TEC markers with known functions in thymocyte development [11], [33]–[37]. MHC Class II staining on Foxn1R/R cTEC was dramatically reduced compared to Foxn1R/+ and WT littermates (Figure 3D and 3Di), and similarly, Cathepsin L expression was down-regulated in Foxn1R/R cTEC (Figure S3). Dll4 expression was dramatically reduced on both fetal and postnatal Foxn1R/R TEC compared to aged-matched WT controls (Figure 3E and 3Ei, see also Figure S3). These data provide a possible explanation for the altered thymocyte subset distributions present in Foxn1R/R mice.
CD40 is expressed on TEC and thymic dendritic cells and is required for medullary development [38], [39]. Expression of CD40 was also substantially down-regulated on postnatal Foxn1R/R cTEC but was largely unaffected on mTEC (Figure 3F). Expression of the recently identified cTEC-specific thymic proteasome subunit ß5t [40] was affected only slightly if at all (Figure 3G and 3Gi; proportion of TEC that are ß5t+: Foxn1R/R, twenty four percent; WT, twenty four percent).
Aire is a key regulator of thymus function, and within the thymus is specifically expressed in a subset of mTEC. Significantly fewer Aire+ mTEC were present in Foxn1R/R compared to WT thymi (Figure 3H and 3Hi).
Collectively, these data established that differentiation of both cortical and medullary TEC was impaired in the postnatal thymus of Foxn1R/R mutants. In particular, since cTEC morphology appeared normal but several cTEC-specific markers were severely down-regulated in Foxn1R/R thymi, they strongly suggested the hypothesis that terminal differentiation was blocked in the cTEC sub-lineage.
Reversion to wild-type Foxn1 expression levels in Foxn1R/R mice leads to rapid acquisition of terminal differentiation phenotypes in cTEC and mTEC
To test whether this hypothesis was correct, or alternatively whether the postnatal Foxn1R/R phenotype reflected the absence of a normal cTEC population and/or overgrowth of a normally minor TEC population, we bred a constitutively expressed tamoxifen inducible Cre recombinase allele, ROSA26CreERt2 [41], onto the Foxn1R/R background. We then tested the outcome of reverting the Foxn1R allele via Cre-mediated excision of the LoxP-flanked cassette, such that WT levels of Foxn1 mRNA expression were restored. Thus, adult Foxn1R/R; ROSA26CreERt2 mice were injected with 4-hydroxy tamoxifen (4OHT) to induce Cre recombinase activity. Two days after 4OHT injection, Foxn1 mRNA expression was restored to at least WT levels (Figure S3) and Ly51, CDR1 and MHC Class II staining was observed in a high proportion of cTEC (Figure 3I, see also Figure S3). Dll4 mRNA expression was also up-regulated by two days post-injection, as was expression of both CCL25 and Cathepsin L mRNA (Figure S3). Since the half-life of cTEC has been established as 10-14 days [42] these data cannot be explained by proliferation of a previously inhibited cell type followed by differentiation, but rather must reflect phenotypic conversion of existing cells. They are therefore consistent with release of a block in terminal differentiation. The proportions of Aire+ mTEC were also restored to WT levels in this experiment (Figure 3I, 3J).
Collectively, these data reveal two important and previously unreported roles of Foxn1. First, although the requirement for Foxn1 for postnatal TEC maintenance has recently been demonstrated [26], this is the first report of a role for Foxn1 in regulating the differentiation of postnatal TEC. Our data establish the requirement for this transcription factor for terminal differentiation of both cTEC and Aire+ mTEC in the postnatal thymus. Since the Foxn1R/R phenotype is most severe in the cortical compartment, they further suggest differential requirements for Foxn1 in the postnatal cortical and medullary TEC compartments. Consistent with this idea, QRT-PCR analysis indicated higher levels of Foxn1 mRNA in cortical than medullary TEC in the normal postnatal thymus (Figure 3K). Second, they also establish that Foxn1 regulates TEC-specific expression of MHC Class II, Dll4, CD40 and Cathepsin L, which are all critically required in TEC for execution of the T cell development programme. While this regulation may be either direct or indirect, it appears to be lymphocyte-independent, as all thymocyte subsets are present in Foxn1R/R thymi.
Generation of an allelic series expressing graded levels of Foxn1 mRNA
We next crossed Foxn1R/R mice with mice carrying the null allele, Foxn1LacZ [13]. Foxn1LacZ phenocopies the original nude mutation and is referred to as Foxn1- herein. Postnatal Foxn1R/- mice had small, alymphoid thymic rudiments that were devoid of any cortical or medullary organisation, and lacked peripheral T cells (Figure 4A); no hematopoietic colonization of the Foxn1R/- thymus was seen at any stage analyzed (Figure S4). Similar to Foxn1nu/nu TEC [19] postnatal Foxn1R/- TEC were predominantly Plet-1+ and co-expressed Keratin 5 and Keratin 8, with occasional cells expressing the cortical and medullary TEC markers CDR1 and UEA1 respectively (Figure 4B). However, unlike the hairless nu/nu or Foxn1-/- mice, Foxn1R/- mice produced an overtly normal coat of hair up to six weeks of age, after which they showed sporadic hair loss. Reactivation of Foxn1 by tamoxifen-mediated induction of Cre recombinase activity in postnatal Foxn1R/-; ROSA26CreERt2 mice up to at least four months old resulted in development of an organized, functional thymus containing all major cortical and medullary TEC sub-types (Figure 4C), indicating that thymic epithelial progenitor cells can persist for prolonged periods in vivo in the absence of wild-type levels of Foxn1, and demonstrating that expression of SV40Tag in these cells did not affect their differentiative capacity.

The differences observed between Foxn1R/- and Foxn1-/- mice suggested that further functions of Foxn1 might be revealed by analysis of an allelic series. We therefore established a series of six strains comprising all possible combinations of Foxn1-, Foxn1R and WT alleles. Precise determination of Foxn1 mRNA expression levels across this series relative to wild type was achieved by analysis of a defined TEC population, E12.5 Plet-1+ TEC (Figure 4D). In E12.5 Plet-1+ TEC, the Foxn1R allele expressed approximately twenty percent of WT Foxn1 mRNA levels (Figure 4D).
Foxn1 is not required for initial divergence of the medullary TEC sub-lineage
The postnatal Foxn1 null phenotype is characterized by the presence of linear aggregates of cells that express markers associated with the earliest thymus-restricted epithelial progenitor cells found in ontogeny [19], suggesting that, in the absence of Foxn1, TE lineage progression might be blocked at the earliest progenitor stage [19], [13]. This idea was supported by the recent demonstration of a persisting common TEPC in postnatal Foxn1-/- mice [8]. However, the possibility remains that more restricted precursors arise in the absence of Foxn1 but are not maintained later in ontogeny.
To determine the stages in early thymus ontogeny at which Foxn1 is required, we analyzed all strains of the allelic series (Figure 5) using marker combinations that identify early cortex (CD205 and K5lo/-) and early medulla (Claudin [Cldn] 4, UEA1, K5hi and MTS10) [10], [11], [43], [44]. Plet-1 [5] was also included as an indicator of founder/early progenitor cell status. We focussed on the E13.5 and E15.5 time points since at these stages the TE is undergoing active patterning into cortical and medullary compartments [45]. Notably, functional analyses have shown that the Cldn4hi cells present at E13.5 in the WT thymic primordium are medullary TE lineage-restricted precursors [10].

By E13.5 WT, Foxn1+/- and Foxn1R/+ thymi contained central K5hiCldn4hi clusters surrounded by K5lo/-K8+Cldn4lo/- regions (Figure 5A), as previously reported for WT thymi [3], [10], [45]; this central Cldn4hi region has previously been demonstrated to correspond to prospective medulla [10]. These data thus confirmed the normal progression of TEC lineage development in Foxn1R/+ mice.
K5hiCldn4hi and K5-Cldn4lo/- regions were also present in the thymic primordia of Foxn1-/- mice at both E13.5 and E15.5 (Figure 5A; Figure S5). These data establish that divergence of the mTEC lineage from the common TEPC occurs in the absence of Foxn1 and is thus Foxn1-independent, in contrast to the existing model of TE lineage development.
cTEC and mTEC differentiation was further probed using markers expressed at various intermediate stages of TEC development. Consistent with recent reports, neither CD205 [11] nor ß5t [40], [46] were detected in Foxn1-/- mice at any stage analyzed, confirming the dependence of the cortical sub-lineage on Foxn1 at least from the onset of expression of these markers. However, a CD205+ population was detectable by E15.5 in all other strains tested, including Foxn1R/- mice, demonstrating that only low levels of Foxn1 mRNA expression are required to promote the early stages of cTEC development (Figure 5B).
By E15.5, UEA1+ and MTS10+ mTEC were present in all strains in the allelic series that express ≥50% WT levels of Foxn1 mRNA (Foxn1+/-, Foxn1R/+ and WT thymi). However, while strains that expressed ≤20% WT Foxn1 mRNA all contained some UEA1+ TEC, MTS10 expression was not detected (shown for Foxn1-/-, Foxn1R/R and Foxn1+/-, Figure 5C). We note that UEA1 staining was present only on rare cells in the Foxn1-/-and Foxn1R/- thymic primordium, but that this staining was consistent and above background. Since MTS10 staining was detected as normal on postnatal mTEC in Foxn1R/R mutants, but was only observed on very rare cells in postnatal Foxn1R/- and Foxn1-/- thymi, we conclude that mTEC differentiation beyond the earliest mTEC sub-lineage progenitor state is blocked in the absence, and delayed at intermediate levels, of Foxn1 expression.
Collectively, the data described above indicate that Foxn1 is not required for divergence of the mTEC sub-lineage from the common TEPC but is required subsequently, in a dosage dependent manner, to promote TEC differentiation. Of note is that high level Plet-1 expression is maintained in Foxn1-/- TECs, supporting the conclusion that these cells remain blocked in the earliest differentiation state despite having undergone lineage divergence (Figure S5).
Low-level Foxn1 expression enables exit from the earliest sub-lineage TEPC states in the fetal thymus but is unable to sustain lineage progression
The above analysis suggested that specific intermediate states of TEC development are dependent on discrete levels of Foxn1 expression. To further test this notion, we used flow cytometry to provide quantitative evaluation of TEC differentiation status in the different allelic variants at defined time-points. For this we analyzed representative markers of the founder TE cell-state (Plet-1 [3], [5]) and functional maturation (MHC Class II [47]); TEC were positively identified using EpCam in all analyses.
In order to identify intermediate phenotypes that occur during normal TEC lineage progression we initially analyzed expression of these markers in WT mice at E12.5, E13.5 and E15.5 (Figure 5D). This demonstrated that normal TEC lineage progression is characterized by the down-regulation of Plet-1 coupled with acquisition of MHC Class II expression.
We then analyzed the allelic series. This revealed that TEC differentiation was delayed when Foxn1 expression was reduced, and that the severity of the delay was inversely proportional to Foxn1 mRNA level. In Foxn1R/R mice the TEC subset profiles at E13.5 and E15.5 were equivalent to those of WT mice at E12.5 and E13.5 respectively (Figure 5E, 5F). Thus, normal TEC differentiation occurred in the Foxn1R/R strain, but with delayed kinetics.
The two functionally athymic strains (Foxn1R/- and Foxn1-/-) each exhibited TEC subset profiles consistent with a severe early block in TEC differentiation. Unexpectedly, the severity of this block differed between these two strains. In Foxn1-/- thymi, the ratio of Plet-1+ to Plet-1lo/- TEC was maintained between E13.5 and E15.5 and was similar to that observed at E12.5 during normal development (determined based on comparison of the mean percentage of Plet-1+ and Plet-1lo/- cells as shown in Figure 5E, 5F legend). However, in Foxn1R/- thymi, the proportion of Plet-1lo/- TEC increased from twenty six percent at E13.5, comparable to E13.5 Foxn1-/- thymi, to forty one percent at E15.5 (see mean percentages provided in Figure 5E, 5F legend). This suggested that by E15.5, at least some Foxn1R/- TEC had been able to exit from the earliest progenitor state and enter the TEC differentiation programme, but that Foxn1-/- TEC could not make this transition. This conclusion was supported by analysis of the differentiation marker MHC Class II, which was expressed by a small population of Plet-1+ and Plet-1lo/- cells in Foxn1R/- mice at E15.5, but was virtually undetectable in Foxn1-/- mice (Figure 5F). Importantly, induction of MHC class II expression occurred in the absence of colonizing haematopoietic cells (Figure S4). Thus, while TECs in the Foxn1-/- thymus remained in an undifferentiated state apparently equivalent to the earliest progenitor stages of each TEC sub-lineage, Foxn1R/- TECs could initiate the very early events associated with TEC lineage differentiation but could not progress beyond this initial differentiation step.
Foxn1 controls allocation into or maintenance of the differentiated TEC compartment
Collectively, the above data establish that Foxn1 is required for progression through successive stages of both cTEC and mTEC differentiation. To test whether Foxn1 might also regulate TEC proliferation and/or survival, we also quantified both founder/early progenitor cells of the lineage (Plet-1+MHC Class II– TEC) and cells that were further into the TE differentiation programme (Plet-1- TEC) for each allelic variant.
At both E13.5 and E15.5, the absolute numbers of Plet-1+ cells correlated with Foxn1 dosage (Figure 6A), consistent with a role for Foxn1 in regulating proliferation of Plet-1+ TEC and thus potentially overall thymus size [48]. However, the most marked effect was on the numbers of Plet-1- TEC present at different levels of Foxn1. At intermediate and low Foxn1 levels (i.e. ≤20% of WT mRNA levels; Foxn1R/R, Foxn1R/- and Foxn1-/- mice) the numbers of Plet-1- TEC were decreased by four-fold compared to Foxn1+/- thymi at both E13.5 and E15.5. Notably, Foxn1 dosage was more important in determining Plet-1+ and Plet-1- cell numbers than the presence or absence of SV40Tag (Figure 6A). These data establish that a threshold level of Foxn1 mRNA between that found in Foxn1R/R and Foxn1+/- TEC is required for generation/expansion of the Plet-1- TEC population with normal kinetics, indicating a role for Foxn1 in regulating thymus size.

The above data could be consistent with roles for Foxn1 in the production, proliferation and/or maintenance of Plet-1- TEC. To begin to discriminate between these possibilities we performed cell cycle analysis on TEC in all of the allelic variants at E15.5. Both Foxn1R/- and Foxn1-/- TEC exhibited slower cell cycle times than WT TEC (Figure 6B; note the proportions of cells in the G1/G0 and S/G2/M fractions). However, no differences in cell cycle were observed between E15.5 WT, Foxn1R/+ and Foxn1R/R TEC (Figure 6B: note the proportions of cells in G0/G1, S and G2/M), establishing that the reduced Plet-1- cell numbers in fetal Foxn1R/R thymi could not be explained by differences in proliferation rate. We therefore conclude that in the fetal thymus, Foxn1 regulates allocation of cells into the differentiating Plet-1- TEC compartment and/or the maintenance of Plet-1- TEC. At present, we cannot distinguish between direct and indirect effects of Foxn1 in regulating fetal Plet-1- cell numbers, as we also observed a significant delay in maturation of the thymic mesenchyme and immigration of mesenchymal cells into the thymic epithelium in Foxn1R/R mice (Figure S6), raising the possibility that the deficit in Plet-1- TEC in these mutants at E15.5 might reflect impaired mesenchymal-epithelial cross talk.
Foxn1 regulates effector genes with diverse roles in thymic epithelial cell development and function
In order to relate our findings to molecular control of TEC differentiation and function, we investigated the response of a suite of genes with diverse functions in thymus biology – i.e. that are known to regulate particular aspects of thymus development or are required in TEC to support T cell development - to changes in Foxn1 dosage. In this panel, we included genes known to be down-regulated or absent from the Foxn1 null thymic primordium or identified in our analysis as being regulated by Foxn1 (Dll4 [34] and CCL25 [49]), or with defined roles in TEC development (Pax 1 [50]-[52]). We also included Fgf2, to date the only gene verified by chromatin immunoprecipitation in keratinocytes as a direct Foxn1 target [53]. Expression of each gene was analyzed at two time points. Initially, we analyzed expression in EpCam+ cells purified from the thymic primordium at E13.5. QRT-PCR analysis revealed that all of the genes tested except Fgf2 showed changes in expression in response to Foxn1 levels (Figure 6C-6E and 6C’-6D’; see also Figure S3). In this population, Dll4, CCL25 and Pax1 were each expressed in proportion to Foxn1 mRNA levels, though titrating to different threshold levels (Figure 6C, 6D). Our data indicate considerable heterogeneity among E13.5 TEC, raising the possibility that the observed expression patterns reflected altered TEC subset balances between the different allelic variants. Therefore, we also analyzed expression of these genes in a defined cell population, E12.5 Plet1+ TEC. The expression levels of all three genes were also proportional to Foxn1 expression in this population (Figure 6 C’ and D’), indicating that, in TEC, Foxn1 is genetically upstream of all three genes. In contrast, EpCam was expressed at comparable levels in the E12.5 Plet1+ TEC population in all of the allelic variants. Surprisingly Fgf2 expression was detected in thymic mesenchyme but not in TEC, indicating that Foxn1 regulates different targets in TEC and keratinocytes (Figure 6F). To test whether Foxn1 regulation of Dll4 and CCL25 was direct or indirect, we over-expressed either the full-length Foxn1 cDNA or GFP cDNA in a cell population that does not normally express Foxn1 or either Dll4 or CCL25. Both genes were significantly up-regulated in cells transfected with Foxn1 but not in control samples (Figure 6G). These data thus strongly suggest that Foxn1 directly regulates both Dll4 and CCL25.
In conjunction with the data presented above showing Foxn1 regulates multiple intermediate TEC states, these data suggest that Foxn1 is a master regulator of the overall ‘thymus programme’ rather than regulating a particular aspect of TEC biology.
Discussion
We have investigated the role of Foxn1 in the TE lineage via analysis of an allelic series that expresses defined levels of Foxn1 mRNA relative to WT. We have demonstrated that, in early ontogeny, Foxn1 is not required for divergence of the medullary TE sub-lineage from the common thymic epithelial progenitor cell; indeed we found no evidence for Foxn1 regulation of the choice between two alternative cell fates at any point in TE lineage progression. We have further demonstrated the requirement for Foxn1 for stable initiation of differentiation in the earliest TEC progenitors present in ontogeny, for subsequent progression through intermediate progenitor stages in both the cortical and medullary TEC sub-lineages, and for terminal differentiation of both cTEC and mTEC in the postnatal thymus. Taken together, these data establish that Foxn1 is required to execute the TE lineage programme at multiple stages in both the cortical and medullary TEC sub-lineages, but does not appear to regulate cell fate choice in TE lineage development. A model for Foxn1 regulation of cellular hierarchies during TE lineage development, based on the genetic analyses presented herein, is proposed in Figure 7.

In addition, we have established that Foxn1 is genetically upstream of a number of genes with known roles in TE lineage development or TEC function, specifically Pax1, Dll4, CD40, Cathepsin L and MHC Class II, demonstrating that Foxn1 regulates these genes either directly or indirectly. Finally, we have confirmed and extended observations that Foxn1 is required indirectly to regulate thymic mesenchymal development [54]. These findings significantly extend understanding of regulation of thymus development and function by this key transcription factor, and pave the way for molecular dissection of these processes.
Role of Foxn1 in TE lineage progression
With respect to TE lineage development during thymus organogenesis, our data (Figure 5) establish that Foxn1 is not required for divergence of the medullary TE sub-lineage from the common TEPC that is currently thought to exist at the basis of the TEC hierarchy [8]. This finding was unexpected, particularly given the recent demonstration of a persisting common TEPC in Foxn1 null mice [8]. Taken together with those of Bleul and colleagues, our data strongly suggest that only the common TEPC can persist in the postnatal Foxn1 null thymus and thus that Foxn1 is required directly or indirectly for TEC survival at stages subsequent to the common TEPC (including sub-lineage restricted progenitors). However, further work is required to explore this hypothesis, as an alternative possibility is that the activity of sub-lineage progenitors could not be detected in the clonal assay employed by Bleul.
Our data further establish that, subsequent to lineage divergence, Foxn1 is required in the fetal thymus for stable exit from the earliest progenitor cell state/entry into the differentiation programme leading to generation of functional TEC in both the cortical and medullary sub-lineages. Strikingly, low-level expression of Foxn1 (as found in Foxn1R/- TEC) is sufficient to initiate or stabilize entry into the TEC differentiation programme, as evidenced by acquisition of MHC Class II staining (see Figure 6), but is not sufficient to promote differentiation to a stage at which TEC in either sub-lineage are able to support T cell development. These data collectively establish that entry into the TE differentiation programme can be genetically separated from execution of the full programme (i.e. the programme leading to TEC that are functionally competent to support T cell development) based on different threshold expression levels of Foxn1. This indicates the requirement for Foxn1 for mediating both processes, and will enable dissection of the molecular role of Foxn1 at early and late differentiation stages.
We have also demonstrated, to our knowledge for the first time, that Foxn1 regulates differentiation of both cTEC and mTEC sub-lineages in the postnatal thymus. Specifically, we have shown that at intermediate levels of Foxn1 expression (as found in Foxn1R/R TEC) differentiation of cTEC is blocked at a CD205+CDR1- developmental stage and in addition, the number of Aire+ mTEC are significantly decreased. That the block in cTEC differentiation is relieved within two days of reversion to WT Foxn1 levels provides genetic evidence for the existence of the CD205+CDR1- intermediate stage in cortical TEC differentiation previously suggested by indirect analyses [11] and extends those findings from the fetal to the postnatal thymus. That the number of Aire+ mTEC is also rapidly restored to WT levels upon reversion of the Foxn1R allele suggests that differentiation of mTEC is also blocked at intermediate Foxn1 levels. From these data, we conclude that Foxn1 regulates similar mechanisms in each of the major TEC sub-lineages.
Our data further indicate MHC Class II as a direct or indirect target of Foxn1 in both fetal and postnatal TEC. Notably, TEC are unique outside the hematopoietic system in constitutively expressing MHC Class II, and the link between Foxn1 and MHC Class II expression is therefore significant.
Foxn1 regulation of genes required for TEC development or function
Finally, we have demonstrated that Foxn1 directly or indirectly regulates a panel of genes that mediate diverse aspects of thymus development or function. These include Pax1, an essential mediator of TEC differentiation/survival [52]. Pax1 is expressed in the third pharyngeal pouch from E9.5 and remains expressed throughout thymus ontogeny, being restricted to cTEC in the postnatal thymus [50]. Although regulation of this Pax gene is only poorly understood, from E11.0 sustained expression of Pax1 requires Hoxa3 [55]. Our data show, to our knowledge for the first time, that expression of Pax1 in the thymic primordium is Foxn1-dependent. They thus place Foxn1 and Hoxa3 together in a network or cascade that regulates Pax1 expression, providing the first demonstration of a genetic interaction between Hoxa3 and Foxn1 (Figure 6H). In this regard, it is striking that Hoxa3+/-Pax1-/-compound mutant mice share some phenotype characteristics with Foxn1R/R mice; both mutants have hypomorphic postnatal thymi and reduced levels of MHC class II expression on TEC [51]. As Hoxa3 itself is expressed in TEC in all of the Foxn1 mutants at E13.5 (data not shown), our data could support either of two alternative explanations: Hoxa3 may regulate Foxn1, which in turn regulates Pax1 expression in the thymic primordium, such that Hoxa3 regulation of Pax1 is Foxn1-dependent (Figure 6Gi), or Hoxa3 and Foxn1 may be required independently to maintain Pax1 expression in the third pharyngeal pouch/early thymus primordium (Figure 6Gii). We note that since Hoxa3 is expressed both in TEC and neural crest-derived mesenchymal cells in the thymic primordium, its regulation of Pax1 could be either direct or indirect.
We further show that Foxn1 regulates CCL25 and Dll4, which each play critical roles in thymocyte development – CCL25 regulates colonization of the fetal thymus [49] while Dll4 is the obligate Notch ligand controlling commitment of haematopoietic progenitors to the T cell lineage [34] respectively. It has previously been suggested, based on antibody staining, that although Dll4 and CCL25 are absent from the Foxn1 null thymus [56] their expression is Foxn1-independent in TEC [57]. However, this view has been challenged in a recent report, which placed Foxn1 upstream of dll4a and ccl25a expression in medaka fish [58]. Our data show that in both early fetal TEC and in the postnatal thymus Dll4 expression is proportional to Foxn1 expression, and further demonstrate that over-expression of Foxn1 in embryonic stem cells results in expression of both Dll4 and CCL25. They therefore extend the findings in medaka to demonstrate Foxn1 regulation of Dll4 and CCL25 in the mammalian thymus, and provide strong evidence that Foxn1 regulation of both of these target genes is direct. Reduced expression of Dll4 in Foxn1R/R mutants also correlates with a proportional increase in intrathymic B cells, indicating that Foxn1 expression within the normal range is necessary to sustain full TEC functionality. Taken together, these data provide mechanistic insight into our findings that Foxn1 regulates multiple aspects of TEC development and function. As they demonstrate Foxn1 regulation of genes required for many different aspects of TEC biology, they suggest that this transcription factor functions a master regulator of the core TEC lineage programme. Furthermore, the finding that different Foxn1-regulated genes show distinct response patterns to changes in Foxn1 dosage may explain why different levels of Foxn1 are required for different functions in TEC.
Collectively, the data presented herein establish Foxn1 as powerful regulator of TEC differentiation in the fetal and adult thymus. Furthermore, they demonstrate that different threshold levels of Foxn1 mRNA are required for different functions. Together with the recent finding that reduction of Foxn1 expression postnatally causes premature thymic involution characterized by loss of TEC [25]-[27], this suggests that Foxn1 may be required in TEC from initiation of differentiation throughout the cell's lifetime. Further work is required to determine whether it is a maintenance factor for differentiated TEC as well as having roles in proliferation and differentiation. Improved understanding of the regulation of Foxn1 in thymic development and homeostasis, and of Foxn1 regulation of its targets in different TEC sub-types, is thus likely to become a priority for strategies aimed at protecting or regenerating the thymus for therapeutic ends.
Materials and Methods
Ethics statement
All animal work was conducted according to UK Home Office guidelines, as established in the ANIMALS (SCIENTIFIC PROCEDURES) ACT 1986.
Mice
Rosa26CreERt2/+ [41] and ZP3-Cre [28] mice were maintained as homozygotes and crossed with Foxn1R/R mice as described. Foxn1-/- mice [59] were maintained as heterozygotes on a C57BL/6 background. Foxn1R mice were backcrossed onto the C57BL/6 background for at least 5 generations and subsequently maintained via intercrossing. For timed matings, noon of the day of the vaginal plug was taken as day 0.5.
Generation of Foxn1 targeting vector
A construct containing the targeting cassette shown in Figure S1 was generated by standard molecular biology techniques and verified by sequencing. Conventional subcloning was used for the majority of cloning steps. PCR cloning using a proof-reading Taq polymerase (Roche) and TOPO-TA cloning vector (Invitrogen) was employed to insert the SV40 T antigen cassette. RecET cloning [60] was used to isolate 4 kb of the mouse Foxn1 locus from PAC367b19.
Southern blotting
Genomic DNA was processed for Southern Blotting as described [61].
Gene targeting and blastocyst injection
Mouse sv129/ola ES cells (line E14tg2a) were electroporated with linearized targeting vector and grown under Blasticidin selection. Three correctly targeted clones, identified by Southern analysis (Figure S1), were expanded and transfected with a Cre recombinase expression plasmid to induce deletion of the BSD cassette. Clones in which this cassette had been deleted, but which retained the rest of the targeting construct including the two remaining LoxP sites, were identified by Southern analysis, verified by sequencing and injected into C57BL/6 blastocysts to generate chimeric mice. Germ-line transmission was confirmed by genotyping for two independently targeted ES cell clones.
Transient transfection
E14Tg2a cells were cultured in GMEM containing FCS and LIF. Transfection was carried out in 12 well plates, at a cell density of 90-95% confluence, using GeneJuice transfection reagent (Novagen). The ratio of transfection reagent to DNA was 5 : 1 and 1 µg of DNA was used for each well. The medium was changed 24 hrs after transfection and 1.5 µg/ml Puromycin was added to the new medium to select for transfected cells. Cells were harvested 48 hours after transfection.
Primers
The sequences of primers used for conventional PCR and QRT-PCR are shown in Table 1 and Table 2 respectively.


Genotyping
Foxn1F and Foxn1R were used to detect the wild-type Foxn1 allele. TF and TR were used to detect the SV40 T antigen cDNA.
Antibodies
MTS20 (IgM) and MTS24 (IgG2a), rat mAbs that recognise Plet-1 [5], and MTS10, a rat mAb that recognises medullary TEC, were kind gifts from R.L. Boyd; anti-EpCAM (G8.8, rat IgG2a, DSHB); anti-Cytokeratin 8 (Troma 1, rat IgG2a, DSHB); anti-Cytokeratin 14 (LL002, mouse IgG3 was a kind gift from E.B. Lane); anti-Cytokeratin 5 (AF138, rabbit IgG, Covance); anti-Claudin 4 (rabbit IgG was a kind gift from S. Tsukita); anti-Dll4 was a kind gift from F. Radtke; anti-CD3-FITC (145-2C11, hamster IgG1); anti-CD4-FITC or PE (H129.19, rat IgG2a); anti-CD8-FITC (53-6.7, rat IgG2a); anti-CD11b-FITC (M1/70, rat IgG2b); anti-CD11c-FITC (HL3, hamster IgG1); anti-α-CD19-FITC (1D3, rat IgG2a); anti-CD25-PE (3C7, rat IgG2b); anti-CD44-APC (1M7, rat IgG2b); anti-Ly76-FITC (Ter119, rat IgG2b); anti-CD31-FITC or purified (390, rat IgG2a); anti-CD45-APC (30-F11, rat IgG2b); anti-PDGFRalpha (APA-5, rat IgG2a); anti-SV40 T antigen (PAb100, mouse IgG1)(all Pharmingen); ERTR7 (rat IgG2a was a kind gift from W. van Ewijk); anti-Cytokeratin (rabbit IgG polyclonal, DAKO); CDR1 (Rat IgG2a, was a kind gift from B Kyewski); anti-Foxn1 C-terminal (IMG-3744, polyclonal, Imgenex); Ly51 (Clone 6C3, Rat IgG2a, Biolegend); anti-ß5t (Rabbit polyclonal, purified, IgG, MBL International); biotinlyated UEA-1 (Vector Laboratories). For detection of unconjugated primaries the following secondary antibodies were used; goat anti-rabbit IgG-alexa488; goat anti-rat IgG-alexa647; goat anti-rat IgG-alexa488; goat anti-mouse IgG-alexa647; donkey anti-rat IgG-alexa488; Streptavidin-alexa647 (all Molecular Probes); mouse anti-rat IgM-PE (G53-238, Pharmingen).
Flow cytometry
Analysis
Mouse fetal thymi were processed for flow cytometric analysis as described [3]. Adult TEC were enzymatically digested to single cell suspensions as described for fetal thymi, and processed for flow cytometric analysis without further enrichment. Adult thymocytes were isolated by mechanical disruption of 5–6 week old thymi and stained with the appropriate antibodies. All staining was for 20 minutes on ice in PBS/5%FCS/5U/ml DNAseI. Data were acquired using a FACS Cailibur (BD San Jose, Ca) or Cyan (Beckman Coulter, Miami, Fl) cytometers and analyzed using Flowjo version 6.4.6 (Tree Star, Inc) software.
Sorting
For isolation of fetal TEC, MTS20+CD45-Ter119-CD31-PDGFRalpha- and MTS20-CD45-Ter119-CD31-PDGFRalpha- populations were purified; anti-EpCam was also used to positively identify TEC in some sorts (as indicated). For isolation of adult TEC, TEC were enriched as described [62] and EpCAM+Ly51+MHC Class IIhi, EpCAM+Ly51+MHC Class IIlo, EpCAM+Ly51-MHC Class IIhi, and EpCAM+Ly51-MHC Class IIlo populations purified after gating against CD45-Ter119- cells. Cell sorting was performed using MoFlo (Dakocytomation) and FACSAria (BD) cell sorters, to a purity of >95% for each sample. For all samples, compensations were determined using antibody capture beads (made in house by S. Monard) and Fluorescence Minus One (FMO) staining was used to determine positivity for each antibody. 7AAD was used to identify dead cells in all samples.
Immunohistochemistry
Whole E13.5 – E15.5 embryos or adult thymi were processed for immunohistochemistry as described [2]. Isotype controls (not shown) were included in all experiments. Staining was analyzed using a Leica AOBS confocal microscope (Leica Microsystems GmbH). The images presented are either single optical sections or projected focus stacks of serial optical sections. The number of Aire+ TEC per mm medullary area was established by analyzing three medullary areas on each of two non-sequential sections from each of three individual mice (i.e. the number of Aire+ cells was counted for six different medullary areas for each of three individual mice per condition). The size of each medullary area was calculated using Adobe Photoshop and the number of Aire+ cells per square mm was then calculated. Statistical analysis was on the average number of Aire+ cells per condition.
Tamoxifen injection
Mice were treated with a single intraperitoneal injection of 1.5 mg 4-hydroxy tamoxifen (4OHT) prepared in ethanol and diluted appropriately in Cremophor (Sigma)/PBS.
RNA isolation
RNA was prepared using Tri-reagent and RNAeasy (both Qiagen) according to manufacturer's instructions. All samples were DNase treated.
QRT-PCR
cDNA was prepared using the Superscript II first strand synthesis kit (Invitrogen) with Oligo-dT primers, according to manufacturer's instructions. For data shown in Figure 4, the IQ SYBR Green Supermix (Bio-Rad) was used for quantification and the relative expression level of the target genes was normalized to the geometric mean of three control genes (Hprt, Ywhaz, Hmbs). For data shown in Figure 3, Figure 6C, 6D, 6F and 6G, Figure S2 and Figure S3, relative expression levels were determined using the Roche Universal Probe Library (Foxn1, Probe 68; SV40Tag, Probe 32; alpha-tubulin, probe 58; Dll4, probe 106; CCL25, probe 9; Pax1, probe 105; Fgf2, probe 4; EpCam, probe 52; EVA, probe 100) with the Roche Lightcycler 480. Relative expression levels are shown after normalization to alpha-tubulin expression using Roche LC480 Relative Quantification software. Technical duplicates or triplicates were run for all samples and no RT and no template controls were included in all experiments. For data shown in Figure 6C’, 6D’ and 6E, relative expression levels were determined using the Roche Universal Probe Library (as above) using microfluidic QRT-PCR (Fluidigm). Pre-amplification was carried out using target specific primer pairs according to the manufacturer's protocol. Samples were loaded onto a BioMark 48.48 Dynamic Array (Fluidigm) and thermal cycling was performed using a BioMark instrument (Fluidigm) according to the manufacturer's protocol. Data analysis was carried out using BioMark Real-Time PCR Analysis Software v2.0 (Fluidigm) and the ΔCt method [63]. Expression levels are shown relative to WT after normalization to Epithelial V-like Antigen (EVA)[64].
Cell cycle analysis
Pregnant female mice were injected with 1mg BrdU at E15.5 and embryos were collected 1 hour after injection. Thymi were immediately microdissected, dissociated and stained with G8.8 (Pharmingen), anti-CD45-PE, anti-BrdU-APC and 7-AAD using the Pharmingen APC BrdU Flow Kit according to manufacturer's instructions.
Western blotting
Nuclear protein fractions were prepared using the Active Motif Nuclear Extract Kit according to manufacturer's instructions, separated by electrophoresis on a SDS-PAGE gel (Novex, Invitrogen), and processed for Western Blotting as described [5].
Statistical analysis
Statistical analysis was performed using the one-way ANOVA test (two tailed), as appropriate for normally distributed data (normal distribution was tested using Chi2 goodness of fit). The alpha level is taken as 0.05. Errors shown are standard deviations throughout. Sample sizes of at least n = 3 were used for statistical analyses.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. GutierrezJCPalaciosR 1991 Heterogeneity of thymic epithelial cells in promoting T-lymphocyte differentiation in vivo. Proc Natl Acad Sci U S A 88 642 646
2. GordonJWilsonVABlairNFSheridanJFarleyA 2004 Functional evidence for a single endodermal origin for the thymic epithelium. Nat Immunol 5 546 553
3. BennettARFarleyABlairNFGordonJSharpL 2002 Identification and characterization of thymic epithelial progenitor cells. Immunity 16 803 814
4. Moore-ScottBAOpokaRLinSCKordichJJWellsJM 2007 Identification of molecular markers that are expressed in discrete anterior-posterior domains of the endoderm from the gastrula stage to mid-gestation. Dev Dyn 236 1997 2003
5. DepreterMGBlairNFGaskellTLNowellCSDavernK 2008 Identification of Plet-1 as a specific marker of early thymic epithelial progenitor cells. Proc Natl Acad Sci U S A 105 961 966
6. GillJMalinMHollanderGABoydR 2002 Generation of a complete thymic microenvironment by MTS24(+) thymic epithelial cells. Nat Immunol 3 635 642
7. RossiSWJenkinsonWEAndersonGJenkinsonEJ 2006 Clonal analysis reveals a common progenitor for thymic cortical and medullary epithelium. Nature 441 988 991
8. BleulCCCorbeauxTReuterAFischPMontingJS 2006 Formation of a functional thymus initiated by a postnatal epithelial progenitor cell. Nature 441 992 996
9. RodewaldHRPaulSHallerCBluethmannHBlumC 2001 Thymus medulla consisting of epithelial islets each derived from a single progenitor. Nature 414 763 768
10. HamazakiYFujitaHKobayashiTChoiYScottHS 2007 Medullary thymic epithelial cells expressing Aire represent a unique lineage derived from cells expressing claudin. Nat Immunol 8 304 311
11. ShakibSDesantiGEJenkinsonWEParnellSMJenkinsonEJ 2009 Checkpoints in the development of thymic cortical epithelial cells. J Immunol 182 130 137
12. NehlsMPfeiferDSchorppMHedrichHBoehmT 1994 New member of the winged-helix protein family disrupted in mouse and rat nude mutations. Nature 372 103 106
13. NehlsMKyewskiBMesserleMWaldschutzRSchuddekopfK 1996 Two genetically separable steps in the differentiation of thymic epithelium. Science 272 : 886 - 889
14. FlanaganSP 1966 ‘Nude’, a new hairless gene with pleiotropic effects in the mouse. Genet Res 8 295
15. PantelourisEM 1968 Absence of thymus in a mouse mutant. Nature 217 126 370 371
16. CordierACHeremansJF 1975 Nude mouse embryo:ectodermal nature of the primordial thymic defect. Scand J Immunol 4 193 196
17. FrankJPignataCPanteleyevAAProwseDMBadenH 1999 Exposing the human nude phenotype [letter]. Nature 398 473 474
18. MyattSSLamEW 2007 The emerging roles of forkhead box (Fox) proteins in cancer. Nat Rev Cancer 7 847 859
19. BlackburnCCAugustineCLLiRHarveyRPMalinMA 1996 The nu gene acts cell-autonomously and is required for differentiation of thymic epithelial progenitors. Proc Natl Acad Sci USA 93 5742 5746
20. BaxterRMBrissetteJL 2002 Role of the nude gene in epithelial terminal differentiation. J Invest Dermatol 118 303 309
21. JanesSMOfstadTACampbellDHWattFMProwseDM 2004 Transient activation of FOXN1 in keratinocytes induces a transcriptional programme that promotes terminal differentiation: contrasting roles of FOXN1 and Akt. J Cell Sci 117 4157 4168
22. SuDMNavarreSOhWJCondieBGManleyNR 2003 A domain of Foxn1 required for crosstalk-dependent thymic epithelial cell differentiation. Nat Immunol 4 1128 1135
23. ItoiMKawamotoHKatsuraYAmagaiT 2001 Two distinct steps of immigration of haematopoietic progenitors into the early thymus anlage. Int Immunol 13 1203 1211
24. BrissetteJLLiJKamimuraJLeeDDottoGP 1996 The product of the mouse nude locus, whn, regulates the balance between epithelial cell growth and differentiation. Genes and Development 10 2212 2221
25. ChengLGuoJSunLFuJBarnesPF Postnatal tissue-specific disruption of transcription factor FoxN1 triggers acute thymic atrophy. J Biol Chem 285 5836 5847
26. ChenLXiaoSManleyNR 2009 Foxn1 is required to maintain the postnatal thymic microenvironment in a dosage-sensitive manner. Blood 113 567 574
27. CorbeauxTHessISwannJBKanzlerBHaas-AssenbaumA 2010 Thymopoiesis in mice depends on a Foxn1-positive thymic epithelial cell lineage. Proc Natl Acad Sci U S A
28. LewandoskiMWassarmanKMMartinGR 1997 Zp3-cre, a transgenic mouse line for the activation or inactivation of loxP-flanked target genes specifically in the female germ line. Curr Biol 7 148 151
29. HauftSMKimSHSchmidtGHPeaseSReesS 1992 Expression of SV-40 T antigen in the small intestinal epithelium of transgenic mice results in proliferative changes in the crypt and reentry of villus-associated enterocytes into the cell cycle but has no apparent effect on cellular differentiation programs and does not cause neoplastic transformation. J Cell Biol 117 825 839
30. KimSHRothKAMoserARGordonJI 1993 Transgenic mouse models that explore the multistep hypothesis of intestinal neoplasia. J Cell Biol 123 877 893
31. HerringBPHoggattAMSmithAFGallagherPJ 1999 Targeted expression of SV40 large T-antigen to visceral smooth muscle induces proliferation of contractile smooth muscle cells and results in megacolon. J Biol Chem 274 17725 17732
32. ThellinOZorziWLakayeBDe BormanBCoumans B etal 1999 Housekeeping genes as internal standards: use and limits. J Biotechnol 75 291 295
33. HozumiKMailhosCNegishiNHiranoKYahataT 2008 Delta-like 4 is indispensable in thymic environment specific for T cell development. J Exp Med 205 2507 2513
34. KochUFioriniEBeneditoRBesseyriasVSchuster-GosslerK 2008 Delta-like 4 is the essential, nonredundant ligand for Notch1 during thymic T cell lineage commitment. J Exp Med 205 2515 2523
35. NakagawaTRothWWongPNelsonAFarrA 1998 Cathepsin L: critical role in Ii degradation and CD4 T cell selection in the thymus. Science 280 450 453
36. HoneyKNakagawaTPetersCRudenskyA 2002 Cathepsin L regulates CD4+ T cell selection independently of its effect on invariant chain: a role in the generation of positively selecting peptide ligands. J Exp Med 195 1349 1358
37. GrayDHSeachNUenoTMiltonMKListonA 2006 Developmental kinetics, turnover, and stimulatory capacity of thymic epithelial cells. Blood 108 3777 3785
38. Van Den BergTKHasboldJRenardel De LavaletteCDoppEADijkstraCD 1996 Properties of mouse CD40: differential expression of CD40 epitopes on dendritic cells and epithelial cells. Immunology 88 294 300
39. AkiyamaTShimoYYanaiHQinJOhshimaD 2008 The tumor necrosis factor family receptors RANK and CD40 cooperatively establish the thymic medullary microenvironment and self-tolerance. Immunity 29 423 437
40. MurataSSasakiKKishimotoTNiwaSHayashiH 2007 Regulation of CD8+ T cell development by thymus-specific proteasomes. Science 316 1349 1353
41. HameyerDLoonstraAEshkindLSchmittSAntunesC 2007 Toxicity of ligand-dependent Cre recombinases and generation of a conditional Cre deleter mouse allowing mosaic recombination in peripheral tissues. Physiol Genomics 31 32 41
42. GrayDHFletcherALHammettMSeachNUenoT 2008 Unbiased analysis, enrichment and purification of thymic stromal cells. J Immunol Methods 329 56 66
43. GodfreyDIIzonDJTucekCLWilsonTJBoydRL 1990 The phenotypic heterogeneity of mouse thymic stromal cells. Immunol 70 66 74
44. KlugDBCarterCCrouchERoopDContiCJ 1998 Interdependence of cortical thymic epithelial cell differentiation and T-lineage commitment. Proc Natl Acad Sci USA 95 11822 11827
45. KlugDBCarterCGimenez-ContiIBRichieER 2002 Cutting edge: thymocyte-independent and thymocyte-dependent phases of epithelial patterning in the fetal thymus. J Immunol 169 2842 2845
46. RipenAMNittaTMurataSTanakaKTakahamaY Ontogeny of thymic cortical epithelial cells expressing the thymoproteasome subunit beta5t. Eur J Immunol 41 1278 1287
47. GrayDAbramsonJBenoistCMathisD 2007 Proliferative arrest and rapid turnover of thymic epithelial cells expressing Aire. J Exp Med 204 2521 2528
48. JenkinsonWEBaconAWhiteAJAndersonGJenkinsonEJ 2008 An epithelial progenitor pool regulates thymus growth. J Immunol 181 6101 6108
49. LiuCSaitoFLiuZLeiYUeharaS 2006 Coordination between CCR7 - and CCR9-mediated chemokine signals in prevascular fetal thymus colonization. Blood 108 2531 2539
50. WallinJEibelHNeubuserAWiltingJKosekiH 1996 Pax1 is expressed during development of the thymus epithelium and is required for normal T-cell maturation. Development 122 23 30
51. SuDMManleyNR 2000 Hoxa3 and pax1 transcription factors regulate the ability of fetal thymic epithelial cells to promote thymocyte development. J Immunol 164 5753 5760
52. SuDEllisSNapierALeeKManleyNR 2001 Hoxa3 and Pax1 regulate epithelial cell death and proliferation during thymus and parathyroid organogenesis. Dev Biol 236 316 329
53. WeinerLHanRScicchitanoBMLiJHasegawaK 2007 Dedicated epithelial recipient cells determine pigmentation patterns. Cell 130 932 942
54. MoriKItoiMTsukamotoNAmagaiT Foxn1 is essential for vascularization of the murine thymus anlage. Cell Immunol 260 66 69
55. ManleyNRCapecchiMR 1995 The role of Hoxa-3 in mouse thymus and thyroid development. Development 121 1989 2003
56. TsukamotoNItoiMNishikawaMAmagaiT 2005 Lack of Delta like 1 and 4 expressions in nude thymus anlages. Cell Immunol 234 77 80
57. ItoiMTsukamotoNAmagaiT 2007 Expression of Dll4 and CCL25 in Foxn1-negative epithelial cells in the post-natal thymus. Int Immunol 19 127 132
58. BajoghliBAghaallaeiNHessIRodeINetuschilN 2009 Evolution of genetic networks underlying the emergence of thymopoiesis in vertebrates. Cell 138 186 197
59. GordonJBennettARBlackburnCCManleyNR 2001 Gcm2 and Foxn1 mark early parathyroid - and thymus-specific domains in the developing third pharyngeal pouch. Mech Dev 103 141 143
60. ZhangYMuyrersJPTestaGStewartAF 2000 DNA cloning by homologous recombination in Escherichia coli. Nat Biotechnol 18 1314 1317
61. MorrisLGordonJBlackburnCC 2006 Identification of a tandem duplicated array in the Rhox alpha locus on mouse chromosome X. Mamm Genome 17 178 187
62. DerbinskiJPintoSRoschSHexelKKyewskiB 2008 Promiscuous gene expression patterns in single medullary thymic epithelial cells argue for a stochastic mechanism. Proc Natl Acad Sci U S A 105 657 662
63. LivakKJSchmittgenTD 2001 Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25 402 408
64. DeMonteLPorcelliniSTafiESheridanJGordonJ 2007 EVA regulates thymic stromal organisation and early thymocyte development. Biochem Biophys Res Commun 356 334 340
65. AshfieldRPatelAJBossoneSABrownHCampbellRD 1994 MAZ-dependent termination between closely spaced human complement genes. EMBO J 13 5656 5667
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2011 Číslo 11
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Evidence-Based Annotation of Gene Function in MR-1 Using Genome-Wide Fitness Profiling across 121 Conditions
- De Novo Origins of Human Genes
- Capture of MicroRNA–Bound mRNAs Identifies the Tumor Suppressor miR-34a as a Regulator of Growth Factor Signaling
- TRY-5 Is a Sperm-Activating Protease in Seminal Fluid
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy