
DAF-12 Regulates a Connected Network of Genes to Ensure Robust Developmental Decisions
The nuclear receptor DAF-12 has roles in normal development, the decision to pursue dauer development in unfavorable conditions, and the modulation of adult aging. Despite the biologic importance of DAF-12, target genes for this receptor are largely unknown. To identify DAF-12 targets, we performed chromatin immunoprecipitation followed by hybridization to whole-genome tiling arrays. We identified 1,175 genomic regions to be bound in vivo by DAF-12, and these regions are enriched in known DAF-12 binding motifs and act as DAF-12 response elements in transfected cells and in transgenic worms. The DAF-12 target genes near these binding sites include an extensive network of interconnected heterochronic and microRNA genes. We also identify the genes encoding components of the miRISC, which is required for the control of target genes by microRNA, as a target of DAF-12 regulation. During reproductive development, many of these target genes are misregulated in daf-12(0) mutants, but this only infrequently results in developmental phenotypes. In contrast, we and others have found that null daf-12 mutations enhance the phenotypes of many miRISC and heterochronic target genes. We also find that environmental fluctuations significantly strengthen the weak heterochronic phenotypes of null daf-12 alleles. During diapause, DAF-12 represses the expression of many heterochronic and miRISC target genes, and prior work has demonstrated that dauer formation can suppress the heterochronic phenotypes of many of these target genes in post-dauer development. Together these data are consistent with daf-12 acting to ensure developmental robustness by committing the animal to adult or dauer developmental programs despite variable internal or external conditions.
Published in the journal:
. PLoS Genet 7(7): e32767. doi:10.1371/journal.pgen.1002179
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1002179
Summary
The nuclear receptor DAF-12 has roles in normal development, the decision to pursue dauer development in unfavorable conditions, and the modulation of adult aging. Despite the biologic importance of DAF-12, target genes for this receptor are largely unknown. To identify DAF-12 targets, we performed chromatin immunoprecipitation followed by hybridization to whole-genome tiling arrays. We identified 1,175 genomic regions to be bound in vivo by DAF-12, and these regions are enriched in known DAF-12 binding motifs and act as DAF-12 response elements in transfected cells and in transgenic worms. The DAF-12 target genes near these binding sites include an extensive network of interconnected heterochronic and microRNA genes. We also identify the genes encoding components of the miRISC, which is required for the control of target genes by microRNA, as a target of DAF-12 regulation. During reproductive development, many of these target genes are misregulated in daf-12(0) mutants, but this only infrequently results in developmental phenotypes. In contrast, we and others have found that null daf-12 mutations enhance the phenotypes of many miRISC and heterochronic target genes. We also find that environmental fluctuations significantly strengthen the weak heterochronic phenotypes of null daf-12 alleles. During diapause, DAF-12 represses the expression of many heterochronic and miRISC target genes, and prior work has demonstrated that dauer formation can suppress the heterochronic phenotypes of many of these target genes in post-dauer development. Together these data are consistent with daf-12 acting to ensure developmental robustness by committing the animal to adult or dauer developmental programs despite variable internal or external conditions.
Introduction
From bacteria to humans, organisms sense environmental cues and reprogram life history by altering gene expression patterns. In the nematode C. elegans, the nuclear receptor DAF-12 alters life history in response to the environment. During development, when conditions are unfavorable, due to starvation, crowding or high temperature, daf-12 promotes diapause and formation of dauer larvae [1]–[3]. In favorable conditions, the DAF-12 receptor is activated by a group of steroidal carboxylic acids, termed dafachronic acids (DA) [4]–[6], produced from cholesterol via a multi-step pathway involving the daf-36 Rieske-like oxygenase and the daf-9 cytochrome P450 enzyme [7]–[9]. The activation of DAF-12 by DA leads to bypass of dauer arrest and expression of later larval developmental and reproductive programs [10]. On the other hand, in unfavorable conditions, DA is not produced, and DAF-12 binds to the co-repressor DIN-1 promoting dauer programs [11]. DAF-12 is also required for the normal lifespan of worms [12]–[14] and for the increased longevity of germline ablated animals [7], [15], [16].
DAF-12 also alters life history by regulation of the heterochronic circuit. The heterochronic pathway in C. elegans determines specific cell fate programs for each larval stage [17]. Several daf-12 alleles, especially those in phenotypic class 1, have heterochronic phenotypes [10]. These mutants repeat L2 programs instead of progressing to L3 programs, resulting in excessive hypodermal seam cell division in the L3 stage and defects in distal tip cell migration in the L4 stage [1], [10]. The heterochronic phenotypes depend on the activity of the co-repressor din-1, as daf-12(rh61);din-1 mutants develop normally [11]. In part, DAF-12 regulates the heterochronic circuit by controlling expression of let-7 family miRNAs [18], [19], which are necessary to promote L3 and adult programs through the down-regulation of the heterochronic genes hbl-1 and lin-41 respectively [20], [21]. DAF-12 might also affect the heterochronic circuit via dauer formation, as many heterochronic phenotypes are suppressed in post dauer development [22]–[25].
Previously, Shostak et al identified DAF-12 binding sites in vitro. However, a small number of genes were described and the in vivo relevance of these target genes was not always clear. To identify novel genes regulated by DAF-12, we developed transgenic animals expressing an epitope-tagged daf-12-transgene. These worms were used to perform chromatin immunoprecipitation followed by hybridization of the precipitated DNA to whole genome tiling arrays. From the arrays, we identified 1175 genomic regions bound to DAF-12 and these regions are within 5 Kb of 3179 genes. Importantly, we detected binding sites in multiple heterochronic genes, miRISC (miRNA-induced silencing complex) genes, and a total of 40 miRNAs, suggesting that DAF-12 controls developmental progression through the direct regulation of these targets.
Biological systems are subject to mutations and environmental variation during development, but still are able to ensure a stereotyped developmental process. This is possible because organisms can buffer many perturbations and produce an invariant output. This capacity is called ‘robustness’ [26]. Since many heterochronic and miRISC genes found in our array were mis-regulated in a daf-12(0) null allele daf-12(rh61rh411), but did not result in abnormal development [10], we hypothesized that DAF-12 could have a role in developmental robustness. Consistent with this idea, we found that daf-12(0) worms can enhance multiple heterochronic phenotypes, indicating that daf-12 null worms are sensitized to genetic mutations. Further, we found that daf-12(0) worms are sensitized to environmental variability and show stronger heterochronic phenotypes in variable instead of constant conditions. In addition, multiple DAF-12 target genes were repressed in dauer, suggesting that in diapause DAF-12 can ‘reset’ the heterochronic circuit to ensure a stereotyped developmental program. These results illustrate a role for DAF-12 in the commitment to reproductive versus dauer development and suggest a novel function for this nuclear receptor to ensure a stereotyped phenotype despite internal and external variability.
Results
Use of a daf-12 transgenic worm for chromatin immunoprecipitation
To identify target genes for DAF-12, we developed transgenic animals expressing a TAP-tagged transgene in a daf-12(rh61rh411) (daf-12(0)) background (Figure 1A). The tandem affinity purification, or TAP-tag, was previously used successfully to identify nicotinic receptor-associated proteins in C. elegans [27], [28]. The TAP-tag was inserted in-frame in daf-12 exon 1 via homologous recombination [29] as insertion of a GFP-tag at this site produced a functional GFP:DAF-12 fusion protein [10]. By biolistic bombardment of ALF3 (daf-12(rh61rh411); unc-119(ed3)), we obtained two integrated lines, ALF4 and ALF9. In each of these strains, DAF-12 is produced solely from the transgene. During the generation of our transgenic lines, a new commercially available DAF-12 antibody was released and this antibody was able to detect the fusion protein (Figure 1B). However, likely due to low DAF-12 expression in N2, endogenous DAF-12 was not detected. This suggests that the daf-12 transgene is over-expressed relative to the endogenous levels. The transgene also rescues the daf-12(0) null allele, as starved transgenic worms form SDS resistant dauers (Figure 1B), suggesting that both transgenic lines express a functional daf-12 transgene.

We performed pilot ChIP-chip experiments with ALF9 using the TAP tag and precipitation with IgG sepharose. In two replicas, we identified 52 genes as potential DAF-12 targets (data not shown). To identify additional target genes, we then optimized the assay by comparing ALF9 to ALF4 using IgG sepharose that recognized the TAP tag or the anti-DAF-12 antibody. Regions of no binding (cyp-15) and high affinity binding (F35G12.2 or the DAF-12 co-repressor din-1) were quantified in both ALF9 and ALF4. Both the TAP and DAF-12 antibody precipitate the same regions in two separate transgenic strains (Figure 1C). Importantly, the strongest binding to F35G12.2 and din-1 was obtained using ALF4 and the DAF-12 specific antibody. Based on these findings, this combination was used to perform the ChIP-chip studies. We did not use the DAF-12 antibody for ChIP-chip studies with N2 worms, as these seemed less likely to be successful due to the very low level of endogenous DAF-12 expression.
To identify binding regions, arrays from three independent worm collections were analyzed and compared with daf-12(0). We chose L2 and adult stages because daf-12 is known to perform important and separate functions at these stages. Its activity at L2 is important for the dauer decision and developmental progression, whereas in the adult it is known to be required for influencing longevity [30]. Chromatin from ALF4 was collected at middle L2 and adult stages and mixed together for further analysis. As shown in Figure 1D, DAF-12 binding to din-1 and F35G12.2 is highly reproducible, as each of the replicas shows similar binding regions and intensity (ChIP enrichment ratio of 16 and 21 respectively).
Global analysis of DAF-12 binding sites
From the resulting tiling array data, peaks representing DAF-12 binding sites were identified using TAS software (Affymetrix) (see Materials and Methods for a complete description). The data reported in this publication have been deposited in the NCBI's Gene Expression Omnibus accession number GSE28350 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE28350) [31]. To be considered a legitimate DAF-12 target, we required the peak to be present in all three replicates and not in a daf-12(0) negative control. This resulted in 1175 genomic regions bound to DAF-12. 1155 were within 5 Kb of 3179 genes and just 20 genomic regions (1.7%) were not associated with any known gene (Table S1). We also repeated the analysis looking for genes within 1 Kb and within 2 Kb of the binding sites (Table S1). Also, a table for binding regions when triplicates were positives and daf-12(0) was also positive is found in Table S2. The number of candidate target genes is lower but comparable in magnitude to the 6018 target genes recently identified for PHA-4, another developmental regulator in C. elegans, via chromatin immunoprecipitation followed by sequencing (ChIP-seq) [32]. Furthermore, approximately 20% of previously identified DAF-12 targets by Shostak et al [33] were also found in our ChIP-chip analysis (not shown), including the lit-1 gene, which was extensively validated in their study.
These DAF-12 binding sites tended to cluster in proximal upstream regions with a smaller number inside or downstream of genes (Figure 2A). The identified DAF-12 target genes are involved in diverse biological functions based on gene ontology associations, including development, metabolism, growth and gene expression (Figure 2B). Examples of these targets include heterochronic genes (Representation Factor = 6.9 p<1.64×10−10), all identified components of the miRISC complex (RF = 9.4 p<1.46×10−6), including regulatory factors such as nhl-2 and xrn-2 [34], [35], genes required for autophagy (RF = 7.1 p<3.014e−04) and a total of 51 ribosome subunits (rpl and rps) (RF = 1.7 p<2.268e−04). Genes involved in development, ribosomal function, mitochondrial function, and aging were found to be particularly over-represented (Table S3) via analysis of the targets with the DAVID program [36].
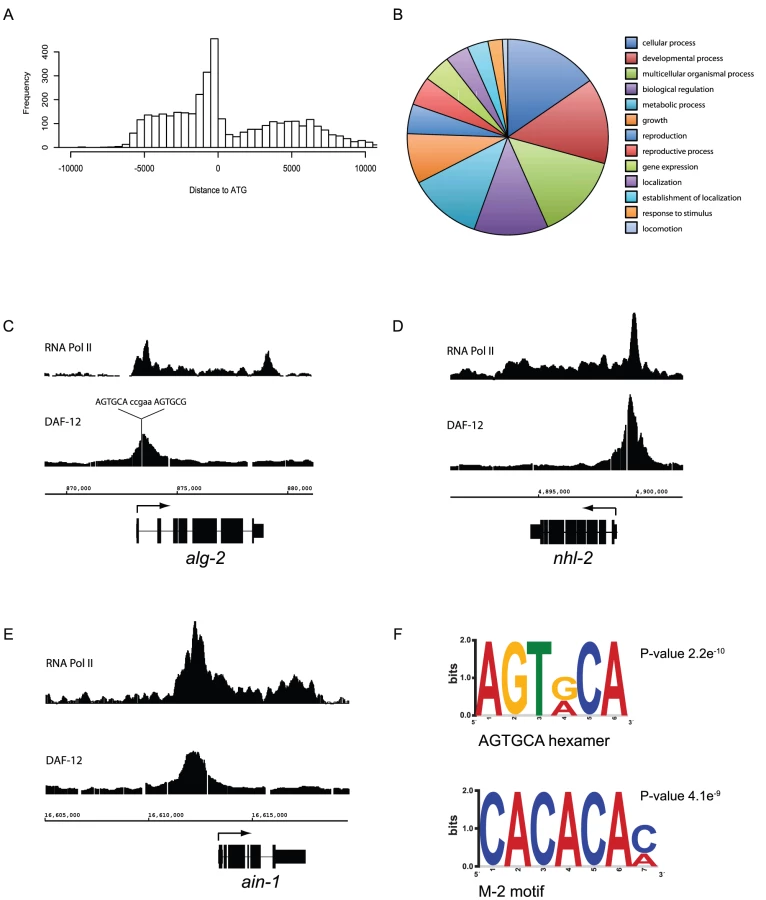
To examine whether DAF-12 binding sites are gene regulatory regions, we compared DAF-12 binding sites in the miRISC genes alg-2, nhl-2 and ain-1, with RNA Pol II binding sites from L2 stage larva as a proof of principle [32]. As expected, DAF-12 binding is localized to upstream promoter regions and associated with the recruitment of RNA Pol II binding, suggesting that DAF-12 is associated with regions of active transcription (Figure 2C–2E). The peak within the alg-2 promoter contains the DR5 repeat, sequence 5′-AGTGCAccgaaAGTGCG-3′, a very high affinity DAF-12 binding site [33] comprised of two hexameric DAF-12 binding sites separated by 5 bp (Figure 2C). The nhl-2 promoter does not contain DR repeats but a total of 15 hexameric sequences were found (Figure 2D). The ain-1 promoter contains just a single hexameric sequence, but has multiple M-2 motifs (CACACA), a previously identified DAF-12 binding site (Figure 2E) [37]. When analyzed globally, approximately 30% of DAF-12 target genes are actively transcribed in the L2 stage (not shown), which may reflect target genes being active at stages other than L2 or transcriptional repression instead of activation of a subset of target genes.
We then analyzed DAF-12 binding regions to find de novo motifs by MEME and DREME, which gives a minimal MEME-formatted motif [38]. Known DAF-12 binding sites, such as the hexameric sequences identified by Shostak et al [33] and the M-2 motif identified by Ao et al [37] were over-represented (Figure 2F). Together these results confirm that the binding sites found in our array contain known DAF-12 binding sites, which is consistent with them being bound in vivo by DAF-12.
The consensus sequence GAGAGA (Figure S1) was also over-represented among binding regions. Interestingly, this GAGA element was also identified as being enriched in PHA-4 binding sites identified by ChIP-seq [32], [37]. DAF-12 and PHA-4 have been shown to co-regulate genes involved in pharynx development and we found that this interaction may be more widespread as they share a significant number of target genes (Figure S2).
DAF-12–dependent regulation of target genes
To test whether binding sites in our array represent DAF-12 response elements, we transfected HEK cells with luciferase reporter constructs along with a myc-tagged DAF-12 expression plasmid [18]. This cell-based assay has the advantage of directly measuring agonist-dependent activation with DA, as opposed to functional assays in S. cerevisiae not involving agonist treatment [33] or worm transgenes, which could be regulated by transcription factors other than DAF-12. Among the targets we identified is the miRISC gene alg-2 whose 2 Kb promoter region is sufficient for agonist-dependent activation (Figure 3A). The alg-2 promoter contains a DR5 sequence that is required for activation (Figure 3A, box). Interestingly, both the WT promoter and the mutant alg-2 promoter that lacks the DR5 domain exhibit weak DAF-12-dependent activation in the absence of DA. This transcriptional activation, which does not require the DR5 domain, could be due to DAF-12 tethering. This activation mechanism was observed for other nuclear receptors, such as ER or GR, where adaptor molecules, like SP1 or AP-1 can recruit the nuclear receptor to the DNA and induce transcriptional activation independent of the agonist [39]. Alternatively, over-expressed DAF-12 could bind to low affinity sites, such as incomplete M-2 motifs or GAGA motifs present in the promoter (Figure S1). Regardless, our findings demonstrate that these play a minor role compared to DAF-12 actions through the DR5 domain.

Another putative target is the promoter region of the CEOP4404 operon, which contains the atg-7 and let-70 genes. As shown in Figure 3B, a 2 Kb promoter region for atg-7 and let-70 is strongly regulated by DAF-12 in cells. To address if this activation is due to DAF-12 recruiting a co-activator, we introduced a point mutation (P687S) within the DAF-12 AF-2 domain, which for other nuclear receptors is involved in recruiting co-activators after ligand binding [40]. This mutation is present in the daf-12(rh284) allele and causes a weak Daf-c phenotype at 25°C, perhaps due to impaired DAF-12 transcriptional activity [10]. As seen in Figure 3B, DAF-12 (P687S) is unable to promote transcriptional activity, even though expression of both proteins is similar (Figure 3B, inset).
We then analyzed DAF-12 target genes previously reported to be involved in dauer formation (Figure 3C). 2 Kb promoter regions of these genes were subcloned upstream of a luciferase gene reporter and transfected in HEK cells. As seen in Figure 3D, all promoter regions show agonist dependent activation, including the DAF-16 CBP/p300 co-activator CBP-1 [41]. These results suggest that DAF-12 could regulate dauer formation in part by regulation of daf-3, din-1 and daf-16 expression, but also by regulation of daf-16 activity through controlling CBP-1 expression. Furthermore, DAF-12 could also regulate daf-16 activity by controlling the expression of akt-1, akt-2 and pdk-1 [42] (Table S1). When considered together, these results demonstrate that the regions identified by ChIP-chip respond in vivo to DAF-12.
DAF-12 regulates post-embryonic development
Proper regulation of the heterochronic circuit is essential for normal development [43]. Mutation in these heterochronic genes can cause either a precocious (skipping stage-specific events) or retarded (reiterating stage-specific events in later stages) developmental phenotype. In addition, miRISC components, required for mRNA degradation or translational repression are also essential for proper development as miRNAs regulate the heterochronic circuit [20]. As shown in Figure 4A, multiple heterochronic genes and all known miRISC genes contain DAF-12 binding sites, suggesting that DAF - 12 can directly regulate their expression.

Regulatory regions of multiple heterochronic genes, such as the F box gene dre-1 [24], the RBCC gene lin-41 [44], the RNA binding gene lin-28 [45] and the circadian clock like genes lin-42A and kin-20 (Figure S3) [46], [47] were subcloned upstream of a luciferase reporter. As shown in Figure 4B, all promoter regions tested, except the RNA binding gene lin-28, were activated by DAF-12 in an agonist-dependent fashion. The same analysis was performed for members of the miRISC complex. As shown in Figure 4B, the decapping gene cgh-1 [48] was activated in a DAF-12-dependent agonist-independent fashion, representing perhaps another example of nuclear receptor tethering (Figure 3A). Also, the argonaute interacting protein, ain-1, shows a modest but reproducible activation (Figure 4B). In addition, alg-2 seems to be a DAF-12 target (Figure 3A), as well as NHL-2, a protein that binds to ALG-1 and ALG-2 to modulate miRNA potency [35] (Figure 4B).
We then examined the in vivo effects of daf-12 on heterochronic gene expression. Wild type (N2) and daf-12(0) worms were collected at different developmental stages, and heterochronic gene expression was quantified by RT-qPCR. As seen in Figure 4C, daf-12(0) worms show significant differences in expression when compared to N2. Importantly, DAF-12 can either repress or activate the same gene depending on the developmental stage (i.e. lin-41 at L4 or adult stage). Interestingly, the circadian clock gene lin-42, which has been suggested to be important for the molting process [47], is normally repressed by DAF-12 at the adult stage. This could be necessary to prevent extra molting cycles once development is completed.
As many heterochronic genes were mis-regulated in L3, we focused on this stage. We compared mRNA levels of different heterochronic and miRISC genes from N2 or daf-12(rh284), a DAF-12 mutant unable to promote transcriptional activity (Figure 3B), using NanoString analysis [49]. As shown in Figure 4D, although all observed differences were not statistically significant, in most cases DAF-12 is necessary for full expression of these targets. However, DAF-12 might be involved in repression of lin-28 as its expression is up-regulated in both daf-12 mutants at the L3 stage (Figure 4C and 4D). Consistent with this, lin-28 mRNA is also up-regulated in din-1(dh149) worms (data not shown). Our findings point to a role for DAF-12 in regulation of heterochronic and miRISC gene expression during post-embryonic development.
Although all known miRISC genes contain DAF-12 binding sites, most of them showed a weak mis-regulation (Figure 4D). To test whether this occurs because daf-12(rh284) still retains some aspect of DAF-12 activity, or because the mis-regulation of these genes is tissue-specific, we created transgenic animals carrying alg-2 and ain-1 promoter regions upstream of GFP in WT or daf-12(0) mutants. As seen in Figure 5A, daf-12(0) worms at L2 stage show impaired alg-2 expression most notably in the pharynx when compared to WT. Further, expression of ain-1 at L3 stage was also impaired in daf-12(0) worms (Figure 5B). Taken together, these results suggest that DAF-12 controls the expression of miRISC components during reproductive development.

daf-12(0) worms are sensitized to genetic mutations
Although many genes involved in development were found in our array, in daf-12(0) mutants development usually occurs normally [10]. Based on this observation, we hypothesized that DAF-12 could have a role in developmental robustness. To address this hypothesis, we performed perturbation experiments [50] using the rrf-3 mutant, which is hypersensitive to RNAi [51]. Both rrf-3 and rrf-3;daf-12 were exposed to miRISC gene RNAi, to see whether loss of daf-12 could unmask developmental defects. When rrf-3 worms were exposed to ain-1 RNAi, only 3–4% developed protruding (Pvl) or burst vulva (Bvl) phenotypes. In contrast, approximately 30% of rrf-3;daf-12 animals showed a penetrant vulva phenotype (Figure 6A). No vulva phenotype was observed for rrf-3 or rrf-3;daf-12 animals when exposed to control RNAi (not shown).

Treatment of rrf-3 worms with ain-1, ain-2, alg-2 or nhl-2 RNAi produced an increase in body length (Figure 6B and data not shown). This result suggests that the known increased body length of daf-12(0) worms [13] could be due to the down-regulation of miRISC genes. However, when daf-12(0) worms were fed nhl-2 or ain-2 RNAi, a synergistic response was observed (Figure 6B).
During development, most hypodermal seam cells undergo a single asymmetric cell division. One daughter cell fuses to the hypodermal syncytium, while the other remains undifferentiated as a seam cell. However, in L2 stage the seam cells V1–4, V6 and H1 undergo a symmetric cell division before the asymmetric division, increasing the number of seam cells from 10 to 16 per side at the L/A switch [52]. Some daf-12 class 1 alleles repeat L2 programs resulting in excessive hypodermal seam cells, but this phenotype is weaker in daf-12(0) animals [10]. As a result, we tested whether miRISC genes interacted with DAF-12 in the control of seam cell division. As shown in Figure 6C, while nhl-2 knock-down induced a modest increase in seam cell number, a synergistic phenotype was revealed in daf-12(0), as visualized with a nuclear localized SCM::GFP transgene. Furthermore, this synergistic response was also seen when other miRISC genes were down-regulated (Figure 6D).
C. elegans larvae hatch with 20 intestinal nuclei, and after 10–14 nuclear divisions in the L1 stage, reach 30–32 nuclei in the adult [53]. We studied postembryonic divisions in intestinal cells as visualized with a nuclear localized elt-2::GFP reporter. As shown in Figure 6E, ain-1 knock-down induced a modest increase in number of intestinal nuclei cells. In contrast, daf-12(0) worms exhibited a synergistic phenotype upon treatment with ain-1 RNAi (Figure 6E) or with RNAi for other miRISC genes (Figure 6F).
These results show that daf-12(+) worms can compensate for the loss of miRISC function to ensure normal development. However, daf-12(0) worms are unable to buffer these mutations. Further, previous observations have shown that daf-12(0) worms demonstrate enhanced developmental defects in response to mutations to multiple heterochronic genes that are also DAF-12 targets (Figure S4). Perhaps the combined mis-regulation of heterochronic genes and impaired miRNA activity, due to the control of miRNA [18] and miRISC expression by DAF-12, inhibits the ability of daf-12(0) worms to buffer genetic mutations during development. These findings point to a role for DAF-12 in ensuring developmental robustness during reproductive development, which we further test below.
DAF-12 represses heterochronic and miRISC genes during dauer formation
The results presented above show that DAF-12 plays a key role in post-embryonic development and directly regulates the heterochronic circuit, miRISC genes and miRNA expression. We reasoned that these target genes might be repressed in diapause, as DAF-12 binds to the co-repressor DIN-1 to promote dauer formation [11]. To test if DAF-12 target genes are repressed in diapause, synchronized daf-2(e1371) or daf-2(e1371);daf-12(rh61rh411) worms were grown at 25°C. At this temperature, daf-2 form dauers while daf-2;daf-12 double mutants continue reproductive development. Worms at L3 stage or dauers were collected for mRNA quantification using NanoString analysis. As seen in Figure 7A, both miRISC and heterochronic genes are strongly repressed in dauer larvae relative to L3, suggesting that during reproductive development DAF-12 induces expression of these genes, whereas in dauer formation these genes are instead silenced.

To determine if DAF-12 is involved in repression of these genes, we created transgenic worms carrying a 2 Kb promoter region from lin-42A upstream of a GFP reporter. The same promoter region was subjected to mutagenesis to eliminate DAF-12 binding sites, such as DR and M-2 domains (Figure S5) [33], [37]. We have chosen to study lin-42A for multiple reasons. DAF-12 regulates lin-42A expression in every developmental stage (Figure 4C), lin-42 is strongly repressed in diapause (Figure 7A) and down-regulation is necessary for dauer formation [54]. As shown in Figure 7B, in L3 stage both WT and mutant promoters drove GFP expression mainly in the pharynx and the tail, and GFP expression was occasionally observed in the intestine. GFP expression was strongly reduced in dauers when compared with the L3 stage for both reporter constructs (Figure 7B). Consistent with this, endogenous lin-42 was repressed approximately 1000× when compared N2 worms at the L3 or dauer stages (Figure 7D). When endogenous lin-42 and lin-42p::GFP were analyzed by RT-qPCR, GFP and lin-42 transcripts were similarly repressed when they form dauers (Figure 7B, upper graph). However, when endogenous lin-42 and the lin-42p(mut)::GFP were compared, the GFP transcript retained 25-fold more expression than the endogenous lin-42 transcript (Figure 7B, lower graph), suggesting that DAF-12 binding sites are strictly required for a full repression of lin-42 in diapause. This differential GFP expression in dauers was observed at a higher exposure and magnification of the pharynx (Figure 7C).
These results show that DAF-12 represses the heterochronic circuit to commit to dauer formation. Interestingly, many heterochronic phenotypes, such as those due to daf-12, dre-1, hbl-1, lin-4, lin-14, lin-28, lin-42, and lin-58 are suppressed after dauer formation [1], [22]–[25] (Figure S6). Together these results suggest that inhibition of the heterochronic circuit by DAF-12 could be necessary to interrupt development and enter diapause. In addition, this could be necessary to reset the heterochronic circuit to progress through development after worms exit dauer, adding another level of regulation to ensure developmental robustness.
DAF-12 acts as a developmental capacitor
As daf-12(0) worms are not able to buffer genetic mutations among key developmental genes such as miRISC genes, we hypothesized that this nuclear receptor could have a role as a phenotypic capacitor. Phenotypic capacitors act to minimize the effects of either environmental or internal variations of a stereotyped developmental process. To test whether daf-12 behaves this manner, we exposed WT or daf-12(0) worms carrying the SCM::GFP transgene to environmental fluctuations during development. Developing worms were grown for 72 hours at 20°C (control) or in a variable environment consisting of shifts between 15°C and 25°C every 4 hours. As seen in Figure 8A, while WT worms occasionally show an extra hypodermal cell division, the shifts in temperature during development significantly enhance the heterochronic defects exhibited in daf-12(0) worms. Interestingly, the number of seam cells in both sides is different, suggesting that environmental fluctuation leads to stochastic errors in seam cell division. When compared daf-12(0), stress conditions almost double the number of worms with strong heterochronic phenotypes (Figure 8A and 8B). Together these results suggest that daf-12(0) worms are sensitized to environmental conditions and confirm that DAF-12 has phenotypic capacitor properties.

Discussion
Identification of DAF-12 target genes
While DAF-12 is necessary for the decision between reproductive development and dauer formation, target genes necessary for these biological functions are still unidentified. In the present work, we identified putative in vivo target genes for DAF-12 in the L2 and adult stages. Several lines of evidence support the specificity and relevance of the binding sites found in our array. First, the highest density of binding sites is found within a 1.5 Kb region upstream of target genes, consistent with DAF-12 often acting by binding to proximal promoter regions. We also found binding sites within genes corresponding to promoter regions of alternative isoforms of the same transcript, such as din-1S and lin-42A. Second, DAF-12 binding sites overlap with RNA Pol II binding sites annotated in an independent ChIP-seq analysis, suggesting that DAF-12 binding sites correspond to regions of active transcription. Third, two independent strains (ALF4 and ALF9) carrying a TAP-tagged daf-12, precipitated either with IgG sepharose or with an anti-DAF-12 antibody, show the same binding regions (Figure 1C and not shown). Fourth, many targets, such as the heterochronic genes dre-1, hbl-1, lin-14, lin-28 and lin-66 have been shown to have genetic interactions with daf-12 [1], [11], [18], [20], [24], [55]. Moreover, known DAF-12 target genes, such myo-2, ceh-22, lit-1, mir-84 and mir-241 were identified by our ChIP-chip experiments [18], [33], [37]. Together these results suggest that we have successfully identified in vivo targets of DAF-12.
DAF-12 controls development by a multi-level regulation of gene expression
Our finding that multiple heterochronic genes contain DAF-12 binding sites, suggests that this receptor directly regulates the heterochronic circuit and is consistent with previous observations that the gain of function allele daf-12(rh61) shows heterochronic phenotypes [1], [10]. DAF-12 also regulates expression of additional genes with heterochronic phenotypes, such as miRNAs (Figure S7) [18], [19] and components of the miRISC complex [56]. Our results show that among miRISC genes, the nuclear receptor DAF-12 directly regulates at least the alg-2 and ain-1 promoters, suggesting that DAF-12 could regulate both miRNA expression and activity.
Beyond effects on miRNA expression and activity, DAF-12 regulates other genes that may control miRNA maturation. Recent evidence in mammals points to a role for LIN-28 in inhibiting let-7 processing from pri and pre-let-7 [57]. We show here that the RNA-binding gene lin-28 is repressed by DAF-12 from the L2 to L4 stages. Assuming a conserved function in C. elegans, lin-28 repression by DAF-12 could be required for let-7 activity to promote execution of L3 programs. We also found the F box gene dre-1 to be regulated by DAF-12. While no target proteins have been found so far for this ubiquitin ligase, its down-regulation produces heterochronic defects [24], suggesting a key function for DRE-1 in development. Control of dre-1 by DAF-12 could result in effects on protein stability or degradation. DAF-12 can also regulate its own expression, as well as genes that modulate its activity, such as the co-repressor DIN-1. Our results suggest that DAF-12 regulates gene expression by mechanisms including direct regulation of target promoters, miRNA biogenesis, miRNA activity via miRISC expression and perhaps control of miRNA maturation via lin-28 (Figure 9).

DAF-12 acts as a phenotypic capacitor
Fluctuations in environmental conditions and genetic mutations during development can have an impact on the phenotypic outcome of a given organism. How biological systems can reduce the impact of these stochastic perturbations to ensure a uniform phenotype is termed canalization or robustness [58]. Genes who function in part to reduce the effect of environment or genetic variability are called phenotypic capacitors and they contribute to robustness [26]. During reproductive development, our data shows that DAF-12 is required for proper expression of multiple heterochronic genes. While loss of daf-12 rarely produces penetrant heterochronic phenotypes during development, the phenotypes of many heterochronic genes are augmented by loss of daf-12 (Figure S4). The lack of a strong null phenotype and enhancement of other genotypes suggests that DAF-12 may act to minimize the developmental consequence of mutations in multiple developmental genes. This action is consistent with a role as a phenotypic capacitor. When conditions become inappropriate for reproductive development, DAF-12 binds to the co-repressor DIN-1 and together this complex represses the heterochronic circuit to enter diapause. Entry into dauer results in many heterochronic phenotypes being suppressed upon dauer recovery (Figure S6). The ability of dauer arrest to suppress the effects of heterochronic mutations is also consistent with a role for DAF-12 as a capacitor.
Besides dauer formation, DAF-12 has been implicated in other phenotypic decisions, such as the development of parasitic larvae [59] or formation of a tooth-like structure in the nematode P. pacificus [60]. Phenotypic capacitors such as DAF-12 can play important roles in evolution by buffering new mutations that could otherwise disrupt development and have allow the expression of new phenotypes in adult life [26].
DAF-12 could ensure robustness during reproductive development because it may act as a ‘hub’, interacting with a large number of genes to create a network of mutually regulating genes. Such networks have been shown to be important for robustness [61]. Also, DAF-12 could secure robustness by positive regulation of dauer loci, such as daf-3, daf-16 or hbl-1 and by repression of genes that inhibit dauer formation, such as lin-42 [54], [62]. In addition, the system can buffer perturbations because gene products, such as miRNAs or DIN-1, modulated by DAF-12 can also regulate DAF-12 activity (Figure 9) [19].
Materials and Methods
Strains
ALF3 strain (daf-12(rh61rh411), unc-119(ed3)) was generated by crossing DP38 (unc-119(ed3)) and AA86 (daf-12(rh61rh411)). The unc-119 phenotype was verified by behavior and daf-12 null allele was verified by PCR. AA86, AA82 (daf-12(rh284)), AA34 (daf-12(rh61)), JR667 (unc-119(e2498::Tc1)III; wIs51) and DR1568 (daf-2(e1371)) were obtained from the CGC which is supported by NIH funding. MM5438 (elt-2::GFP, rol-6) was kindly provided by Dr Morris F. Maduro (UC Riverside).
Generation of a TAP tagged daf-12 transgene
The daf-12:TAP transgene was generated from the fosmid WRM0611cB06 (Source Bioscience, Nottingham, UK) by homologous recombination in the SW106 E. coli strain as previously described [29]. Briefly, in the first step the galK gene was inserted into the fosmid between amino acids 26 and 27 in exon 1 using a PCR product generated from pMOD4 galK-GT using oligos containing 50 bp regions of fosmid homology (Dataset S1). Then, the galK gene was replaced with the TAP tag using a PCR product generated from pBS1761 with the same oligos [63]. The unc-119 marker was then added to this fosmid via cre-loxP recombination with pLoxP unc-119 [29]. The EPI300 strain (Epicentre Biotechnologies) was then used to amplify the fosmid for purification with the FosmidMAX DNA purification kit (Epicentre). The modified fosmid was then sequenced to verify the tag sequence and location.
Transgenic worms
ALF4 and ALF9 strains were obtained by biolistic bombardment of ALF3 with the TAP-tagged daf-12 fosmid as previously described [64]. ALF62 and ALF63 strains were generated by biolistic bombardment of the ALF3 strain. ALF62 (unc-119 (ed3), bafIs62 (lin-42p::GFP)) was generated by integration of an extra chromosomal array by TMP/UV irradiation [65]. ALF63 (unc-119 (ed3), bafIs63 (lin-42pmut::GFP)) was directly obtained from bombardment. ALF62 and ALF63 were then crossed with HT1593 (unc-119(ed3)) to generate daf-12(+) strains. ALF70 (unc-119 (ed3), bafIs70 (alg-2p::GFP)) and ALF72 (unc-119 (ed3), bafIs70 (ain-1p::GFP)) were generated by biolistic bombardment of the DP38 strain (unc-119(ed3)). These were crossed with ALF3 to place the reporters in a daf-12(0) background (ALF73 and ALF74 respectively).
Sample preparation and ChIP-chip
Synchronized worms at mid-L2 stage and young adults were collected by floatation, rinsed with milliQ water (DEPC treated) and frozen at −80°C. Whole C. elegans were simultaneously broken up and fixed by brief sonication in PBS containing 1% formaldehyde. Total fixation time was 30 minutes at room temperature, after which the reaction was quenched by the addition of glycine to a final concentration of 0.125 M. Preparation of chromatin and ChIP was carried out essentially as described [66]. In brief, lysates were sonicated and the DNA sheared to an average length of 300–500 bp. Genomic DNA (Input) was prepared by treating aliquots of chromatin with RNase, proteinase K and heat for de-crosslinking, followed by phenol/chloroform extraction and ethanol precipitation. Pellets were resuspended and the resulting DNA was quantified on a Nanodrop spectrophotometer. Extrapolation to the original chromatin volume allowed quantification of the total chromatin yield.
Equal amounts of L2 and adult chromatin (20 µg each) were mixed together and samples were pre-cleared with protein A agarose beads (Invitrogen). Genomic DNA regions of interest were isolated using an antibody against DAF-12 (Santa Cruz, ce-267). After incubation at 4°C overnight, protein A agarose beads were used to isolate the immune complexes. Complexes were washed, eluted from the beads with SDS buffer, and subjected to RNase and proteinase K treatment. Crosslinks were reversed by incubation overnight at 65°C, and ChIP DNA was purified by phenol-chloroform extraction and ethanol precipitation.
Quantitative PCR (QPCR) reactions were carried out in triplicate on specific genomic regions using SYBR Green Supermix (Bio-Rad), site-specific primer pairs, and an aliquot of ChIP DNA. The resulting signals were normalized for primer efficiency by carrying out QPCR for each primer pair using Input DNA.
ChIP and Input DNAs were amplified by whole-genome amplification (WGA) using the GenomePlex WGA Kit (Sigma). The resulting amplified DNAs were purified, quantified, and tested by QPCR at the same specific genomic regions as the original ChIP DNA to assess quality of the amplification reactions. Amplified DNAs were fragmented and labeled using the DNA Terminal Labeling Kit from Affymetrix, and then hybridized to Affymetrix GeneChip C. elegans Tiling 1.0R arrays at 45°C overnight. Arrays were washed and scanned, and the resulting CEL files were analyzed using Affymetrix TAS software that computed for each probe estimates of fold enrichment (in linear scale) over hybridization with input DNA. At the same time, TAS calculated for each probe a p-value by applying a Wilcoxon signed rank test. A threshold of 2.5 was selected, which corresponds to probe intensities approximately 2.5× stronger on the ChIP array than on the Input array. Additional TAS threshold parameters were MinRun = 180 bp, MaxGap = 300 bp. TAS analysis showed that the selected threshold of 2.5 corresponds approximately to a p-value of 0.01. In addition, the false discovery rate (FDR) was estimated as previously described [67]. At a threshold of 2.5, the FDRs for the 4 samples were estimated to be: AA86 = 3.17%, ALF4(1) = 0.25%, ALF4(2) = 0.34% and ALF4(3) = 2.33%. The TAS-generated BED files (containing the peak intervals) were analyzed using Genpathway (now Active Motif) proprietary software that provides comprehensive information on genomic annotation, peak metrics and sample comparisons for all peaks. Graphs of genomic regions with peak enrichment values were generated using the Affymetrix Integrated Genome Browser. The data discussed in this publication have been deposited in NCBI's Gene Expression Omnibus [31] and are accessible through GEO Series accession number GSE28350 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE28350).
Western blotting
Protein was extracted from N2, ALF3, AA86 or transgenic worms by boiling in LDS loading buffer (Invitrogen). After sonication, samples were run on a 10% polyacrylamide gel and transferred to PVDF. Membranes were probed with 1∶750 α-DAF-12 (ce-267, Santa Cruz), and 1∶3000 actin (sc-1616R, Santa Cruz) followed by IRDye 680 goat anti-rabbit (1∶10000) (Li-Cor Biosciences). For detection of DAF-12 expressed in HEK cells, α-myc (9E10, Covance) was used at a dilution of 1∶1000.
Computational analysis
Using the Affymetrix TAS software (Affymetrix), we identified all genes associated with probes that were positive in transgenic Chromatin IPs, but negative in the control and plotted the distance between the bound probe and the ATG proximal to each gene. We next downloaded available GO annotations for all genes associated with bound probes (WormMart, 04/09/2010) and classified them using the GO.db library (Version 2.2.0) in R (http://www.r-project.org/). To find de novo DAF-12 binding sites, we identified all sequence intervals associated with genes that are active in experimental samples, but not in the control. We submitted 600 unique sequences of 100 nt each, centered on the strongest DAF-12 binding signals to MEME (http://meme.mbcr.net) after masking repeat sequences using RepeatMasker (http://www.repeatmasker.org). As a negative control, we also likewise analyzed 600 unique sequences of 100 nt each from intervals with the weakest DAF-12 binding signals associated with genes that did not score positive in the control or experimental samples 1 and 2.
RNA preparation, RT-qPCR, and NanoString analysis
Eggs from N2 and mutant strains were obtained by hypochlorite treatment and hatched overnight in S-Basal with shaking. The arrested L1 larvae were put on NGA plates to resume development. Samples were collected by floatation at the indicated developmental stages, rinsed in milliQ water and snap-frozen. Dauer larvae were isolated by a 15% Ficoll 400, 0.1 M NaCl gradient. RNA extraction and qPCR was performed as previously described [68]. For Nanostring analysis, RNA samples from 6 worm collections were analyzed by Expression Analysis (Durham, NC).
Plasmid DNAs
The DAF-12 A1 cDNA was PCR amplified from C. elegans cDNA and subcloned as a SfiI-BglI fragment into pCMV myc (Clonetech). The DAF-12 rh284 mutant was made with the Quick-Change mutagenesis kit (Stratagene). For luciferase reporter genes, fragments between 2 Kb and 3.5 Kb upstream the ATG were PCR amplified and subcloned into pGL3 Luc Promoter (Promega). For GFP reporter strains, promoter regions were subcloned in pPD95.75 vector (A gift of Dr. Andy Fire). Oligos sequences are available in Figure S8. The unc-119 rescue fragment was introduced by recombination as previously described [69]. The lin-42::GFP promoter mutant was synthesized and subcloned into pPD95.75 vector (Celtek Bioscience).
Cell culture and reporter assay
HEK 293T cells were cultured and transfected (Fugene, Roche) in 24well plates, with DMEM and 10% charcoal FBS treated (Hyclone). Briefly, 225 ng of either pCMV myc DAF-12 A1 or pCMV myc vector were co-transfected with 12.5 ng of pRL-TK (Renilla luciferase) and 25 ng pGL3 Luc promoter with the promoter of interest (Promega). The following day, cells were stimulated with 0.5 µM (25R)-Δ7-DA [70] or DMSO for 16 hr. At this concentration, no difference in activity was observed between the (25R) - Δ7-DA and (25S) - Δ7-DA [71] (not shown). Luciferase was measured using Dual-Luciferase Reporter Assay System (Promega). Each experiment was repeated three times and fold activation is relative to the non stimulated control transfected with pCMV vector, unless indicated otherwise.
Digital imaging
Transgenic worms were mounted for digital photography using an Olympus BX51 upright microscope. All photos within a panel were taken on the same day with identical camera settings to allow direct comparison. Quantification of the GFP reporters was performed with image J software and analyzed with an unpaired t-test.
RNAi experiments
L3 worms were transferred to RNAi plates at 16°C for approximately 72 hr. 10 adults were transferred to new RNAi plates and left for 3 hr to lay eggs before being removed. Analysis was performed with the adult F1 progeny.
Environmental stress
Eggs from WT or daf-12(0) worms carrying the SCM::GFP transgene were obtained by hypochlorite treatment and incubated for 72 hours at 20°C or 15°C–25°C cycles of 4 hours each with a programmable incubator (Innova 4230, New Brunswick Scientific, New Brunswick, NJ).
Supporting Information
Zdroje
1. AntebiACulottiJGHedgecockEM 1998 daf-12 regulates developmental age and the dauer alternative in Caenorhabditis elegans. Development 125 1191 1205
2. CassadaRCRussellRL 1975 The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Dev Biol 46 326 342
3. RiddleDLSwansonMMAlbertPS 1981 Interacting genes in nematode dauer larva formation. Nature 290 668 671
4. GillMSHeldJMFisherALGibsonBWLithgowGJ 2004 Lipophilic regulator of a developmental switch in Caenorhabditis elegans. Aging Cell 3 413 421
5. HeldJMWhiteMPFisherALGibsonBWLithgowGJ 2006 DAF-12-dependent rescue of dauer formation in Caenorhabditis elegans by (25S)-cholestenoic acid. Aging Cell 5 283 291
6. MotolaDLCumminsCLRottiersVSharmaKKLiT 2006 Identification of ligands for DAF-12 that govern dauer formation and reproduction in C. elegans. Cell 124 1209 1223
7. GerischBWeitzelCKober-EisermannCRottiersVAntebiA 2001 A hormonal signaling pathway influencing C. elegans metabolism, reproductive development, and life span. Dev Cell 1 841 851
8. JiaKAlbertPSRiddleDL 2002 DAF-9, a cytochrome P450 regulating C. elegans larval development and adult longevity. Development 129 221 231
9. RottiersVMotolaDLGerischBCumminsCLNishiwakiK 2006 Hormonal control of C. elegans dauer formation and life span by a Rieske-like oxygenase. Dev Cell 10 473 482
10. AntebiAYehWHTaitDHedgecockEMRiddleDL 2000 daf-12 encodes a nuclear receptor that regulates the dauer diapause and developmental age in C. elegans. Genes Dev 14 1512 1527
11. LudewigAHKober-EisermannCWeitzelCBethkeANeubertK 2004 A novel nuclear receptor/coregulator complex controls C. elegans lipid metabolism, larval development, and aging. Genes Dev 18 2120 2133
12. FisherALLithgowGJ 2006 The nuclear hormone receptor DAF-12 has opposing effects on Caenorhabditis elegans lifespan and regulates genes repressed in multiple long-lived worms. Aging Cell 5 127 138
13. GemsDSuttonAJSundermeyerMLAlbertPSKingKV 1998 Two pleiotropic classes of daf-2 mutation affect larval arrest, adult behavior, reproduction and longevity in Caenorhabditis elegans. Genetics 150 129 155
14. LarsenPLAlbertPSRiddleDL 1995 Genes that regulate both development and longevity in Caenorhabditis elegans. Genetics 139 1567 1583
15. GerischBRottiersVLiDMotolaDLCumminsCL 2007 A bile acid-like steroid modulates Caenorhabditis elegans lifespan through nuclear receptor signaling. Proc Natl Acad Sci U S A 104 5014 5019
16. HsinHKenyonC 1999 Signals from the reproductive system regulate the lifespan of C. elegans. Nature 399 362 366
17. AmbrosV 1989 A hierarchy of regulatory genes controls a larva-to-adult developmental switch in C. elegans. Cell 57 49 57
18. BethkeAFielenbachNWangZMangelsdorfDJAntebiA 2009 Nuclear hormone receptor regulation of microRNAs controls developmental progression. Science 324 95 98
19. HammellCMKarpXAmbrosV 2009 A feedback circuit involving let-7-family miRNAs and DAF-12 integrates environmental signals and developmental timing in Caenorhabditis elegans. Proc Natl Acad Sci U S A 106 18668 18673
20. AbbottALAlvarez-SaavedraEMiskaEALauNCBartelDP 2005 The let-7 MicroRNA family members mir-48, mir-84, and mir-241 function together to regulate developmental timing in Caenorhabditis elegans. Dev Cell 9 403 414
21. ReinhartBJSlackFJBassonMPasquinelliAEBettingerJC 2000 The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 403 901 906
22. AbrahanteJEDaulALLiMVolkMLTennessenJM 2003 The Caenorhabditis elegans hunchback-like gene lin-57/hbl-1 controls developmental time and is regulated by microRNAs. Dev Cell 4 625 637
23. AbrahanteJEMillerEARougvieAE 1998 Identification of heterochronic mutants in Caenorhabditis elegans. Temporal misexpression of a collagen::green fluorescent protein fusion gene. Genetics 149 1335 1351
24. FielenbachNGuardavaccaroDNeubertKChanTLiD 2007 DRE-1: an evolutionarily conserved F box protein that regulates C. elegans developmental age. Dev Cell 12 443 455
25. LiuZAmbrosV 1991 Alternative temporal control systems for hypodermal cell differentiation in Caenorhabditis elegans. Nature 350
26. MaselJSiegalML 2009 Robustness: mechanisms and consequences. Trends Genet 25 395 403
27. RigautGShevchenkoARutzBWilmMMannM 1999 A generic protein purification method for protein complex characterization and proteome exploration. Nat Biotechnol 17 1030 1032
28. GottschalkAAlmedomRBSchedletzkyTAndersonSDYatesJR3rd 2005 Identification and characterization of novel nicotinic receptor-associated proteins in Caenorhabditis elegans. EMBO J 24 2566 2578
29. ZhangYNashLFisherAL 2008 A simplified, robust, and streamlined procedure for the production of C. elegans transgenes via recombineering. BMC Dev Biol 8 119
30. RottiersVAntebiA 2006 Control of Caenorhabditis elegans life history by nuclear receptor signal transduction. Exp Gerontol 41 904 909
31. EdgarRDomrachevMLashAE 2002 Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res 30 207 210
32. ZhongMNiuWLuZJSarovMMurrayJI 2010 Genome-wide identification of binding sites defines distinct functions for Caenorhabditis elegans PHA-4/FOXA in development and environmental response. PLoS Genet 6 e1000848 doi:10.1371/journal.pgen.1000848
33. ShostakYVan GilstMRAntebiAYamamotoKR 2004 Identification of C. elegans DAF-12-binding sites, response elements, and target genes. Genes Dev 18 2529 2544
34. ChatterjeeSGrosshansH 2009 Active turnover modulates mature microRNA activity in Caenorhabditis elegans. Nature 461 546 549
35. HammellCMLubinIBoagPRBlackwellTKAmbrosV 2009 nhl-2 Modulates microRNA activity in Caenorhabditis elegans. Cell 136 926 938
36. Huang daWShermanBTLempickiRA 2009 Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc 4 44 57
37. AoWGaudetJKentWJMuttumuSMangoSE 2004 Environmentally induced foregut remodeling by PHA-4/FoxA and DAF-12/NHR. Science 305 1743 1746
38. BaileyTLWilliamsNMislehCLiWW 2006 MEME: discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res 34 W369 373
39. MusgroveEASutherlandRL 2009 Biological determinants of endocrine resistance in breast cancer. Nat Rev Cancer 9 631 643
40. MasuyamaHBrownfieldCMSt-ArnaudRMacDonaldPN 1997 Evidence for ligand-dependent intramolecular folding of the AF-2 domain in vitamin D receptor-activated transcription and coactivator interaction. Mol Endocrinol 11 1507 1517
41. NasrinNOggSCahillCMBiggsWNuiS 2000 DAF-16 recruits the CREB-binding protein coactivator complex to the insulin-like growth factor binding protein 1 promoter in HepG2 cells. Proc Natl Acad Sci U S A 97 10412 10417
42. ParadisSAilionMTokerAThomasJHRuvkunG 1999 A PDK1 homolog is necessary and sufficient to transduce AGE-1 PI3 kinase signals that regulate diapause in Caenorhabditis elegans. Genes Dev 13 1438 1452
43. MossEG 2007 Heterochronic genes and the nature of developmental time. Curr Biol 17 R425 434
44. SlackFJBassonMLiuZAmbrosVHorvitzHR 2000 The lin-41 RBCC gene acts in the C. elegans heterochronic pathway between the let-7 regulatory RNA and the LIN-29 transcription factor. Mol Cell 5 659 669
45. MossEGLeeRCAmbrosV 1997 The cold shock domain protein LIN-28 controls developmental timing in C. elegans and is regulated by the lin-4 RNA. Cell 88 637 646
46. BanerjeeDKwokALinSYSlackFJ 2005 Developmental timing in C. elegans is regulated by kin-20 and tim-1, homologs of core circadian clock genes. Dev Cell 8 287 295
47. JeonMGardnerHFMillerEADeshlerJRougvieAE 1999 Similarity of the C. elegans developmental timing protein LIN-42 to circadian rhythm proteins. Science 286 1141 1146
48. CollerJParkerR 2005 General translational repression by activators of mRNA decapping. Cell 122 875 886
49. GeissGKBumgarnerREBirdittBDahlTDowidarN 2008 Direct multiplexed measurement of gene expression with color-coded probe pairs. Nat Biotechnol 26 317 325
50. RutherfordSLLindquistS 1998 Hsp90 as a capacitor for morphological evolution. Nature 396 336 342
51. SimmerFTijstermanMParrishSKoushikaSPNonetML 2002 Loss of the putative RNA-directed RNA polymerase RRF-3 makes C. elegans hypersensitive to RNAi. Curr Biol 12 1317 1319
52. RougvieAE 2001 Control of developmental timing in animals. Nat Rev Genet 2 690 701
53. LambieEJ 2002 Cell proliferation and growth in C. elegans. Bioessays 24 38 53
54. TennessenJMOppermanKJRougvieAE 2010 The C. elegans developmental timing protein LIN-42 regulates diapause in response to environmental cues. Development
55. MoritaKHanM 2006 Multiple mechanisms are involved in regulating the expression of the developmental timing regulator lin-28 in Caenorhabditis elegans. EMBO J 25 5794 5804
56. GrishokAPasquinelliAEConteDLiNParrishS 2001 Genes and mechanisms related to RNA interference regulate expression of the small temporal RNAs that control C. elegans developmental timing. Cell 106 23 34
57. ViswanathanSRDaleyGQGregoryRI 2008 Selective blockade of microRNA processing by Lin28. Science 320 97 100
58. WaddingtonCH 1959 Canalization of development and genetic assimilation of acquired characters. Nature 183 1654 1655
59. WangZZhouXEMotolaDLGaoXSuino-PowellK 2009 Identification of the nuclear receptor DAF-12 as a therapeutic target in parasitic nematodes. Proc Natl Acad Sci U S A 106 9138 9143
60. BentoGOgawaASommerRJ 2010 Co-option of the hormone-signalling module dafachronic acid-DAF-12 in nematode evolution. Nature 466 494 497
61. LehnerBCrombieCTischlerJFortunatoAFraserAG 2006 Systematic mapping of genetic interactions in Caenorhabditis elegans identifies common modifiers of diverse signaling pathways. Nat Genet 38 896 903
62. KarpXAmbrosV 2011 The Developmental Timing Regulator hbl-1 Modulates the Dauer Formation Decision in Caenorhabditis elegans. Genetics 187 345 353
63. PuigOCasparyFRigautGRutzBBouveretE 2001 The tandem affinity purification (TAP) method: a general procedure of protein complex purification. Methods 24 218 229
64. HochbaumDFergusonAAFisherAL 2010 Generation of transgenic C. elegans by biolistic transformation. J Vis Exp
65. Gengyo-AndoKMitaniS 2000 Characterization of mutations induced by ethyl methanesulfonate, UV, and trimethylpsoralen in the nematode Caenorhabditis elegans. Biochem Biophys Res Commun 269 64 69
66. SoutoglouETalianidisI 2002 Coordination of PIC assembly and chromatin remodeling during differentiation-induced gene activation. Science 295 1901 1904
67. JohnsonWELiWMeyerCAGottardoRCarrollJS 2006 Model-based analysis of tiling-arrays for ChIP-chip. Proc Natl Acad Sci U S A 103 12457 12462
68. FergusonAASpringerMGFisherAL 2010 skn-1-Dependent and -independent regulation of aip-1 expression following metabolic stress in Caenorhabditis elegans. Mol Cell Biol 30 2651 2667
69. FergusonAAFisherAL 2009 Retrofitting ampicillin resistant vectors by recombination for use in generating C. elegans transgenic animals by bombardment. Plasmid 62 140 145
70. MartinRDäbritzFEntchevEVKurzchaliaTVKnölkerH-J 2008 Stereoselective synthesis of the hormonally active (25S)-delta7-dafachronic acid, (25S)-delta4-dafachronic acid, (25S)-dafachronic acid, and (25S)-cholestenoic acid. Org Biomol Chem 6 4293 4295
71. MartinRSchmidtAWTheumerGKrauseTEntchevEV 2009 Synthesis and biological activity of the (25R)-cholesten-26-oic acids–ligands for the hormonal receptor DAF-12 in Caenorhabditis elegans. Org Biomol Chem 7 909 920
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2011 Číslo 7
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- A Rice Plastidial Nucleotide Sugar Epimerase Is Involved in Galactolipid Biosynthesis and Improves Photosynthetic Efficiency
- Genome-Wide Association Study Identifies Novel Restless Legs Syndrome Susceptibility Loci on 2p14 and 16q12.1
- Loss of the BMP Antagonist, SMOC-1, Causes Ophthalmo-Acromelic (Waardenburg Anophthalmia) Syndrome in Humans and Mice
- Gene-Based Tests of Association
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy