
Missense Mutation in Exon 2 of SLC36A1 Responsible for Champagne Dilution in Horses
Champagne coat color in horses is controlled by a single, autosomal-dominant gene (CH). The phenotype produced by this gene is valued by many horse breeders, but can be difficult to distinguish from the effect produced by the Cream coat color dilution gene (CR). Three sires and their families segregating for CH were tested by genome scanning with microsatellite markers. The CH gene was mapped within a 6 cM region on horse chromosome 14 (LOD = 11.74 for θ = 0.00). Four candidate genes were identified within the region, namely SPARC [Secreted protein, acidic, cysteine-rich (osteonectin)], SLC36A1 (Solute Carrier 36 family A1), SLC36A2 (Solute Carrier 36 family A2), and SLC36A3 (Solute Carrier 36 family A3). SLC36A3 was not expressed in skin tissue and therefore not considered further. The other three genes were sequenced in homozygotes for CH and homozygotes for the absence of the dilution allele (ch). SLC36A1 had a nucleotide substitution in exon 2 for horses with the champagne phenotype, which resulted in a transition from a threonine amino acid to an arginine amino acid (T63R). The association of the single nucleotide polymorphism (SNP) with the champagne dilution phenotype was complete, as determined by the presence of the nucleotide variant among all 85 horses with the champagne dilution phenotype and its absence among all 97 horses without the champagne phenotype. This is the first description of a phenotype associated with the SLC36A1 gene.
Published in the journal:
. PLoS Genet 4(9): e32767. doi:10.1371/journal.pgen.1000195
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1000195
Summary
Champagne coat color in horses is controlled by a single, autosomal-dominant gene (CH). The phenotype produced by this gene is valued by many horse breeders, but can be difficult to distinguish from the effect produced by the Cream coat color dilution gene (CR). Three sires and their families segregating for CH were tested by genome scanning with microsatellite markers. The CH gene was mapped within a 6 cM region on horse chromosome 14 (LOD = 11.74 for θ = 0.00). Four candidate genes were identified within the region, namely SPARC [Secreted protein, acidic, cysteine-rich (osteonectin)], SLC36A1 (Solute Carrier 36 family A1), SLC36A2 (Solute Carrier 36 family A2), and SLC36A3 (Solute Carrier 36 family A3). SLC36A3 was not expressed in skin tissue and therefore not considered further. The other three genes were sequenced in homozygotes for CH and homozygotes for the absence of the dilution allele (ch). SLC36A1 had a nucleotide substitution in exon 2 for horses with the champagne phenotype, which resulted in a transition from a threonine amino acid to an arginine amino acid (T63R). The association of the single nucleotide polymorphism (SNP) with the champagne dilution phenotype was complete, as determined by the presence of the nucleotide variant among all 85 horses with the champagne dilution phenotype and its absence among all 97 horses without the champagne phenotype. This is the first description of a phenotype associated with the SLC36A1 gene.
Introduction
Many horse breeders value animals with variation in coat color. Several genes are known which diminish the intensity of the coloration and are phenotypically described as “dilutions”. Two of these are a result of the Cream (CR) locus and Silver (Z) locus. The molecular basis for Cream is the result of a single base change in exon 2 of the SLC45A2 (Solute Carrier 45 family A2, aka MATP for membrane associated transport protein) on ECA21 [1],[3]. This change results in the replacement of a polar acidic aspartate with a polar neutral asparagine in a putative transmembrane region of the protein coded for by this gene [3],[2]. CR has an incompletely dominant mode of expression. Heterozygosity for CR dilutes only pheomelanin (red pigment) whereas homozygosity for CR results in extreme dilution of both pheomelanin and eumelanin (black pigment) [4].
The Silver dilution is the result of a missense mutation of PMEL17 (Premelanosomal Protein) on ECA6. The base change causes replacement of a cytosolic polar neutral arginine with non-polar neutral cysteine in PMEL17 [2]. In contrast to CR, the Z locus is fully dominant and affects only eumelanin causing little to no visible change in the amount of pheomelanin regardless of zygosity. The change in eumelanin is most apparent in the mane and tail where the black base color is diluted to white and gray [5].
The coat color produced by the CH locus is similar to that of the CR locus in that both can cause dilution phenotypes affecting pheomelanin and eumelanin. However, the effect of CH differs from CR in that; 1) CH dilutes both pheomelanin and eumelanin in its heterozygous form and 2) heterozygotes and homozygotes for CH are phenotypically difficult to distinguish. The homozygote may differ by having less mottling or a slightly lighter hair color than the heterozygote. Figure 1 displays images of horses with the three base coat colors chestnut, bay and black and the effect of CH upon each. Figure 2 shows that champagne foals are born with blue eyes, which change color to amber, green, or light brown and pink “pumpkin skin which acquires a darker mottled complexion around the eyes, muzzle, and genitalia as the animal matures [6]. Foals with one copy of CR also have pink skin at birth but their skin is slightly darker and becomes black/near black with age. The champagne phenotype is found among horses of several breeds, including Tennessee Walking Horses and Quarter Horses. Here we describe family studies that led to mapping the gene and subsequent investigations leading to the identification of a genetic variant that appears to be responsible for the champagne dilution phenotype.


Results
Linkage Analyses
Table 1 summarizes the evidence for linkage of the CH gene to a region of ECA14. The linkage phase for each family was apparent based on the number of informative offspring in each family. Recombination rates (θ) were based on the combined recombination rate from all families. Four microsatellites showed significant linkage to the CH locus: VHL209 (LOD = 6.03 for θ = 0.14), TKY329 (LOD = 3.64 for θ = 0.10), UM010 (LOD = 5.41 for θ = 0.04) and COOK007 (LOD = 11.74 for θ = 0.00).

Figure 3 identifies the haplotypes for offspring of a single sire showing recombination between the genetic markers and the CH locus. Pedigrees of the three sire families and haplotype information are provided in Figure S1 and Table S1 respectively. The CH locus maps to an interval between UM010 and TKY329 with microsatellite. No recombinants were detected among 39 informative offspring between the CH and COOK007 locus.

Candidate Genes
Candidate genes were selected on the basis of proximity to the marker COOK007 and as genes previously characterized in other species as influential in the production or migration of pigment cells.
SPARC was located closest at ∼90 kb downstream from COOK007 and is coded for on the plus strand of DNA. It has been implicated in migration of retinal pigment epithelial cells in mice [7].
SLC36A family members are solute carriers and other solute carrier families have been found to play a role in coat color. SLC36A1 is located ∼250 kb downstream from COOK007. It is the first and most proximal to COOK007 of three genes in this family and is coded for on the minus strand of DNA.
SLC36A2 and SLC36A3 are coded for on the plus strand of DNA and are approximately 350 k and 380 k downstream from COOK007 respectively. A2 and A3 have been found to be expressed in a limited range of tissues in humans and mice [8].
RT-PCR
RT-PCR (reverse transcription-polymerase chain reaction) was used to determine if SLC36A1, SLC36A2 or SLC36A3 were expressed in skin. SLC36A1 and SLC36A2 were expressed in skin and their genomic exons were sequenced. SLC36A3 was not detected in skin and therefore not investigated for detection of SNPs. Results for RT-PCR of these three genes are shown in Figure 4.

Sequencing
All 9 exons of SPARC were sequenced. Three SNPs were found in exons but none showed associations with the champagne phenotype and are shown in Table S2.
SLC36A2 was sequenced with discovery of 9 SNPs in exons. None of the SNPs showed associations with CH. These SNPs and all other variations detected are described in Table S2.
SLC36A1 was sequenced. Only one SNP was found, a missense mutation involving a single nucleotide change from a C to a G at base 76 of exon 2 (c.188C>G) (Figure 5). These SLC36A1 alleles were designated c.188[C/G], where c.188 designates the base pair location of the SNP from the first base of SLC36A1 cDNA, exon 1. Sequencing traces for the partial coding sequence of SLC36A1 exon 2 with part of the flanking intronic regions for one non-champagne horse and one champagne horse were deposited in GenBank with the following accession numbers respectively: EU432176 and EU432177. This single base change at c.188 was predicted to cause a transition from a threonine to arginine at amino acid 63 of the protein (T63R).

Protein Alignment
Figure 6 shows the alignment of the protein sequence for exons 1 and 2 of SLC36A1 for seven mammalian species with sequence information from Genbank (horse, cattle, chimpanzee, human, dog, rat and mouse). Alignment was performed using AllignX function of Vector NTI Advance 10 (Invitrogen Corp, Carlsbad, California). The alignment demonstrates that this region is highly conserved among all species. At position 63, the amino acid sequence is completely conserved among these species, with the exception of horses possessing the champagne phenotype. This replacement of threonine with arginine occurs in a putative transmembrane domain of the protein [9].

Population Data
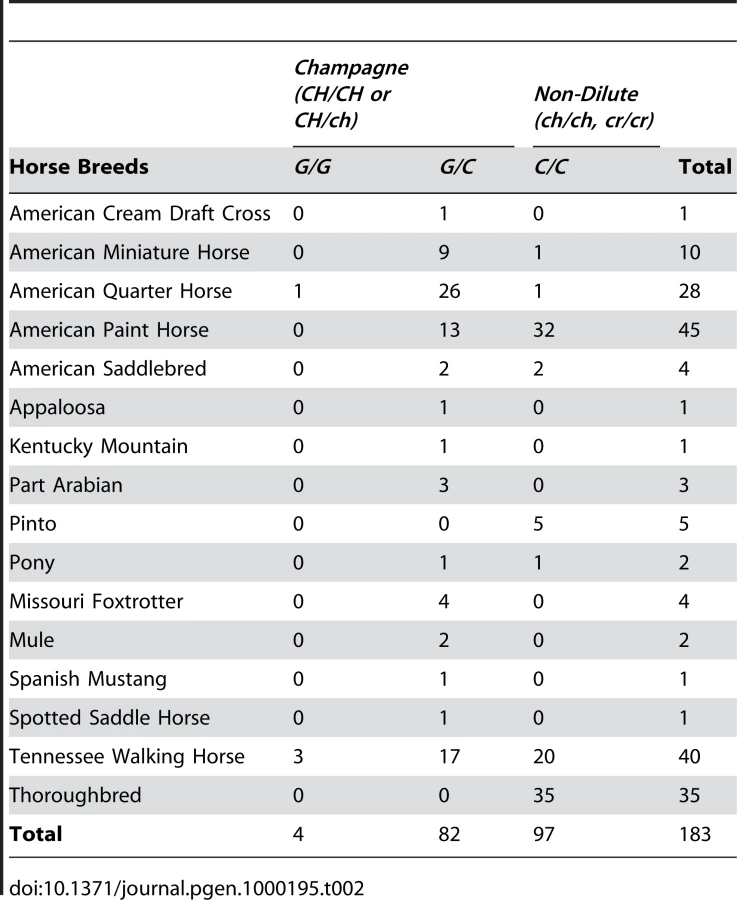
The distribution of c.188G allele among different horse breeds and among horses with and without the champagne phenotype was investigated. Table 2 is a compilation of the population data collected via the genotyping assay. All dilute horses (85) which did not have the CR gene, tested positive for the c.188G allele with genotypes c.188C/G or c.188G/G. No horses in the non-dilute control group (97) possessed the c.188G allele. The horses used for the population study were selected for coat color and not by random selection; therefore measures of Hardy-Weinberg equilibrium are not applicable and were not calculated.
Discussion
Family studies clearly showed linkage of the gene for the champagne dilution phenotype within a 6 cM region on ECA14 [10] (Table 1). Based on the Equine Genome Assembly V2 as viewed in ENSEMBL genome browser (http://www.ensembl.org/Equus_caballus/index.html) this region spans approximately 2.86 Mbp [11]. Within that region, four candidate genes were investigated; one based on known effects on melanocytes (eg. SPARC) and three for their similarity to other genes previously shown to influence pigmentation (eg, SLC36A1, A2, and A3). While SNPs were found within the exons of SPARC, none were associated with CH. Of the other 3 candidate genes, only SLC36A1 and SLC36A2 were found to be expressed in skin cells. Therefore, the exons of those two genes were sequenced. A missense mutation in the second exon of SLC36A1 showed complete association with the champagne phenotype across several breeds. While SNPs were found for SLC36A2, none showed associations at the population level for the champagne dilution phenotype.
This observation is the first demonstration for a role of SLC36A1 in pigmentation. Orthologous genes in other species are known to affect pigmentation. For example, the gene responsible for the cream dilution phenotypes in horses, SLC45A2 (MATP), belongs to a similar solute carrier family. In humans, variants in SLC45A2 have been associated with skin color variation [12] and a similar missense mutation (p.Ala111Thr) in SLC24A5 (a member of potassium-dependent sodium-calcium exchanger family) is implicated in dilute skin colors caused from decreased melanin content among people of European ancestry [13]. The same gene, SLC24A5 is responsible for the Golden (gol) dilution as mentioned in the review of mouse pigment research by Hoekstra (2006) [14]
It is proposed, here, that the missense mutation in exon 2 of SLC36A1 is the molecular basis for champagne dilution phenotype. While this study provides evidence that this is the mutation responsible for the champagne phenotype, the proof is of a statistical nature and a non-coding causative mutation can not be ruled out at this point. SLC36A1, previously referred to by the name PAT1 (proton/amino acid transporter 1) in human and mouse [15], is a proton coupled small amino acid transporter located and most active in the brush border membranes of intestinal epithelial cells. This protein has also been characterized in rats under the name LYAAT1 (lysosomal amino acid transporter 1). LYAAT1 is localized in the membrane of lysozomes in association with LAMP1 (lysosomal associated protein 1) and in the cell membrane of post-synaptic junctions. In lysozomes it allows outward transport of protons and amino acids from the lysozome to the cytosol [16]. During purification and separation of early-stage melanosomes LAMP1 is found in high concentrations in the fraction containing stage II melanosomes [17],. Perhaps SLC36A1 plays a role in transitions from lysozome-like precursor to melanosome. Since organellular pH affects tyrosine processing and sorting [18], an amino acid substitution in this protein may affect pH of the early stage melanosome and the ability to process tyrosine properly. There must be an increase in pH, before the tyrosinase can be activated. The cytosolic pH gradient must also be maintained for proper sorting and delivery of the other proteins required for melanosome development [19]. Thus, the pH gradient of the cell may be altered by this mutation.
This variant, discovered in association with a coat dilution in the horse, is the first reported for the SLC36A1 gene. The phenotype resulting from this mutation, a reduction of pigmentation in the eyes, skin and hair, illustrates previously unknown functions of the protein product of SLC36A1. Furthermore, now that a molecular test for champagne dilution is established, the genotyping assay can be used in concert with available tests for cream dilution and silver dilution to clarify the genetic basis of a horse's dilution phenotype. This will give breeders a new tool to use in developing their breeding programs whether they desire to breed for these dilutions or to select against them.
Materials and Methods
Horses
Three half-sibling families, designated 1, 2 and 3, were used for mapping studies. Family 1 consisted of a Tennessee Walking Horse (TWH) stallion, known heterozygous at the Champagne locus (CH/ch), and his 17 offspring out of non-dilute mares (ch/ch). Family 2 consisted of an American Paint Horse stallion (CH/ch) and his 10 offspring out of non-dilute (ch/ch) mares. Family 3 consisted of a TWH stallion (CH/ch), 23 offspring and their 12 non-dilute dams (ch/ch) and 1 dilute (buckskin) dam (ch/ch, CR/cr).
To investigate the distribution of the gene among dilute and non-dilute horses of different horse breeds, 97 non-champagne horses were chosen from stocks previously collected and archived at the MH Gluck Equine Research Center. These horses were from the following breeds: TWH (20), Thoroughbreds (TB, 35), American Paint Horses (APHA, 32), Pintos (5), American Saddlebreds (ASB, 2), one American Quarter Horse (AQHA), one pony, and one American Miniature (AMH) Horse.
Hair and blood samples from horses with the champagne dilution phenotype were submitted by owners along with pedigree information and photographs showing the champagne color and characteristics of each horse. Samples were collected from the following breeds (85 total): American Miniature Horse (9), American Cream Draft cross (1), American Quarter Horse (27), American Paint Horse (13, in addition to the family), American Saddlebred (2), Appaloosa (1), ASB/Friesian cross (1), Arabian crossed with APHA or AQHA horses (3), Missouri Foxtrotter(4), Mule (2), Pony (1), Spanish Mustang 1), Spotted Saddle Horse (1), Tennessee Walking Horse (20, in addition to the families).
Color Determination
To be characterized as possessing the champagne phenotype, horses exhibited a diminished intensity of color (dilution) in black or brown hair pigment and met at least two of the three following criteria: 1) mottled skin around eyes, muzzle and/or genitalia, 2) amber, green, or light brown eyes, or 3) blue eyes and pink skin at birth [6]. This was accomplished by viewing photo evidence of these traits or by personal inspection. Due to potential confusion between phenotypes of cream dilution and champagne dilution, all DNA samples from horses with the dilute phenotype were tested for the CR allele and data from those testing positive were not included in the population data.
DNA Extraction
DNA from blood samples was extracted using Puregene whole blood extraction kit (Gentra Systems Inc., Minneapolis, MN) according to its published protocol. Hair samples submitted by owners were processed using 5–7 hair bulbs according to the method described by Locke et al. (2002). The hair bulbs were placed in 100 µl lysis solution of 1× FastStart Taq Polymerase PCR buffer (Roche, Mannheim, Germany), 2.5 mM MgCl2 (Roche), 0.5% Tween 20 (JT Baker, Phillipsburg, NJ) and 0.01 mg proteinase K (Sigma-Aldrich, St Louis, MO) and incubated at 60°C for 45 minutes, followed by 95°C for 45 min to deactivate the proteinase K.
Microsatellite Genome Scan
The genome scan was done in polymerase chain reaction (PCR) multiplexes of 3 to 6 microsatellites per reaction. The 102 microsatellite markers used are listed in Table S3. Primers for these microsatellites were made available in connection with the USDA-NRSP8 project [20]. Two additional microsatellites were used; TKY329 [21] was selected based on its map location between two microsatellites used for genome scanning (UM010 and VHL209) and COOK007 was developed in connection with this study based on DNA sequence information from the horse genome sequence viewed in the UCSC genome browser [8] in order to investigate linkage within the identified interval. Primers for COOK007 were designed using Primer 3 software accessed online (Forward, 5′ - 6FAM-CATTCCAAACACCAACAACC - 3′), (Reverse, 5′ – GGACATTCCAGCAATACAGAG – 3′) [22]. The initial scan was conducted on a subset of samples from Family 3; including sire 3, five non-champagne offspring and five champagne offspring. When the microsatellite allele contribution from the sire was not informative, (e.g. the sire and offspring had the same genotype), dams from family 3 were typed to determine the precise contribution from the sire. When the inheritance of microsatellite markers in family 3 appeared to be correlated with the inheritance of the CH allele, then the complete families A, B and C were typed and the data analyzed for linkage by LOD score analysis [23].
Amplification for fragment analysis was done in 10 µl PCR reactions using 1× PCR buffer with 2.0 mM MgCl2, 200 µM of each dNTP, 1 µl genomic DNA from hair lysate, 0.1 U FastStart Taq DNA polymerase (Perkin Elmer, Waltham, MA) and the individual required molarity of each primer from the fluorescently labeled microsatellite parentage panel primer stocks at the MH Gluck Equine Research Center. Samples were run on a PTC-200 thermocycler (MJ research, Inc., Boston, MA) at a previously determined optimum annealing temperature for each multiplex. Capillary electrophoresis of product was run on an ABI 310 genetic analyzer (Applied Biosystems Inc. ABI, Foster City, CA). Results were then analyzed using the current version of STRand microsatellite analysis software (http://www.vgl.ucdavis.edu/informatics/STRand/).
Sequencing
PCR template for sequencing was amplified in 20 µl PCR reactions using 1× PCR buffer with 2.0 mM MgCl2, 200 µM of each dNTP, 1 µl genomic DNA from hair lysate, 0.2 U FastStart Taq DNA polymerase (Perkin Elmer) and 50 nM of each primer. Exon 2 of SLC36A1 was sequenced with the following primers: Forward (5′-CAG AGC CTA AGC CCA GTG TC-3′) and Reverse (5′-GGA GGA CTG TGT GGA AAT GG-3′) at an annealing temperature of 57°C. Primers used to sequence the other SLC36A1 exons and primers for sequencing genomic exons of SLC36A2 are provided in parts 1 and 2 respectively of Table S4. Template product was quantified on a 1% agarose gel, then amplified with BigDye Terminator v1.1 cycle sequencing kit according to manufacturer's instructions (Applied Biosystems), cleaned using Centri-Sep columns (Princeton Separations Inc., Adelphia, NJ), and run on and ABI 310 genetic analyzer (Applied Biosystems). Six samples were initially sequenced: 2 suspected homozygous champagnes (based on production of all champagne dilution offspring when bred to at least 10 non-dilute dams), 2 heterozygotes, and 2 non-dilute horses. The results were analyzed and compared by alignment using ContigExpress from the Vector NTI Advance 10.3 software package (Invitrogen Corporation, Carlsbad, California).
Reverse Transcription (RT-PCR)
RT-PCR was performed in 25 µl reactions a Titan One Tube RT-PCR Kit (Roche) according to enclosed protocol with the primers listed in part 3 of Table S4. RNA from different tissues of non-dilute horses was used to acquire partial cDNAs containing the first two exons for SLC36A1, first three exons SLC36A2 and first 4 exons of SLC36A3. The cDNA acquired was sequenced and the resulting sequences were verified for their respective genes with a BLAT search using the equine assembly v2 in ENSEMBL (http://www.ensembl.org/Equus_caballus/index.html) genome browser. RT-PCR was also performed utilizing RNA extracted from skin, kidney and testes of non-dilute animals currently in lab stocks. SLC36A1 cDNA was produced from the skin and blood using 50 ng RNA per reaction. SLC36A2 cDNA was produced from testes using 1 mRNA per RT-PCR reaction then following up with a nested PCR for shorter product. SLC36A2 cDNA was produced from skin using 50 ng mRNA per RT-PCR reaction. Nested PCR was not necessary. SLC36A3 cDNA was produced from testes using 1 ug mRNA per reaction. 9 µl of initial reaction was visualized on a 2% agarose gel to check for visible bands of product. When product was not initially detected an additional 20 µl PCR was performed in reactions as outlined above using 5 µl of RT product in the place of hair lysate per reaction. Detected product was then sequenced with the protocol listed above. Sequences were then used in a BLAST search using equine genome assembly 2 on ENSEMBL genome browser to verify the correct cDNA was amplified.
Custom TaqMan Probe Assay
A Custom TaqMan SNP Genotyping Assay (Applied Biosystems) was designed for c.188C/G SNP in filebuilder 3.1 software (Applied Biosystems) to test the population distribution of the SLC36A1 alleles. A similar assay was also designed to test for the cream SNP. These assays were run on a 7500HT Fast Real Time-PCR System (Applied Biosystems). All dilute horses tested for SLC36A1 variants were concurrently tested for SLC45A variants. Horses testing positive for CR alleles were not used in the dataset to avoid any confusion over the origin of their dilution phenotype.
Supporting Information
{kind=link}
Zdroje
1. MariatD
TaouritS
GuerinG
2002 A Mutation in the MATP gene causes the cream coat colour in the horse. Genet Sel Evol 35(1) 119 133
2. BrunbergE
AnderssonS
CothranG
SandbergK
MikkoS
2006 A missense mutation in PMEL17 is associated with the Silver coat color in the horse. BMC Genet doi: 10.1186/1471-2156-7-46
3. LockeM
RuthLS
MillonLV
PenedoMCT
MurrayJD
2001 The cream dilution gene, responsible for the palomino and buckskin coat colours, maps to horse chromosome 21. Anim Genet 32 340 343
4. AdalsteinssonS
1974 Inheritance of the palomino color in Icelandic horses. J Hered 65 15 20
5. BowlingA
2000 Genetics of Color variation.
BowlingAT
RuvinskyA
New York CABI Publishing 62 In the Genetics of the Horse Edited by:
6. SponenbergD
2003 Equine Color Genetics. 2nd Edition Ames, IA Iowa State University Press 46 49
7. SheridanCM
MageeRM
HiscottPS
HaganS
WongDH
McGalliardJN
Grierson
2002 The role of matricellular proteins thrombospondin-1 and osteonectin during RPE cell migration in proliferative vitreoretinopathy. Curr Eye Res 25(5) 279 85
8. BerminghamJ
PenningtonJ
2004 Organization and expression of the SLC36 cluster of amino acid transporter genes. Mamm Genome DOI:10.1007/s00335-003-2319-3
9. BollM
FoltzM
Rubio-AliagaI
DanielH
2003 A cluster of proton/amino acid transporter genes in the human and mouse genomes. Genomics 82 47 56
10. PenedoMCT
MillonLV
BernocoD
BaileyE
BinnsM
2005 International Equine Gene Mapping Workshop Report: A comprehensive linkage map constructed with data from new markers and by merging four mapping resources. Cytogenet and Genome Res 111 5 15
11. KentWJ
SugnetCW
FureyTS
RoskinKM
PringleTH
2002 The Human Genome Browser at UCSC. Genome Res 12(6) 996 1006
12. GrafJ
HodgsonR
van DaalA
2005 Single nucleotide polymorphisms in the MATP gene are associated with normal human pigmentation variation. Hum Mutat 25(3) 278 84 2005 Mar;
13. LamasonRL
MohideenMA
MestJR
WongAC
NortonHL
2005 SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans. Science 310 1782 1786
14. HoekstraHE
2006 Genetice, development and evolution of adaptive pigmentation in vertebrates. Heredity 97 222 234
15. ChenZ
FeiYJ
AndersonCM
WakeKA
MiyauchiS
2003 Structure, function and immunolocalization of a proton-coupled amino acid transporter (hPAT1) in the human intestinal cell line Caco-2. J Physiol 15;546(Pt 2) 349 61
16. WredenCC
JohnsonJ
TranC
SealRP
CopenhagenDR
2003 The H+-coupled electrogenic lysosomal amino acid transporter LYAAT1 localizes to the axon and plasma membrane of hippocampal neurons. J Neurosci 15;23(4) 1265 75
17. KushimotoT
BasrurV
ValenciaJ
MatsunagaJ
VieiraW
2001 A model for melanosome biogenesis based on the purification and analysis of early melanosomes. Proc Natl Acad Sci Vol. 98 no. 19 10698 10703
18. WatabeH
ValenciaJC
YasumotoK
KushimotoT
AndoH
2003 Regulation of Tyrosinase Processing and Trafficking by Organellar pH and by Proteasome Activity. J Biol Chem 279 7971 7981
19. WatabeH
ValenciaJC
LePapeE
YamaguchiY
NakamuraM
2008 Involvement of Dynein and Spectrin with Early Melanosome Transport and Melanosomal Protein Trafficking. J Invest Dermatol DOI: 10.1038/sj.jid.5701019
20. GuérinG
BaileyE
BernocoD
AndersonI
AntczakDF
1999 Report of the international equine gene mapping workshop: male linkage map. Anim Genet 30 341 54
21. TozakiT
MashimaS
HirotaK
MiuraN
Choi-MiuraNH
TomitaM
2001 Characterization of equine microsatellites and microsatellite-linked repetitive elements (eMLREs) by efficient cloning and genotyping methods. DNA Res 8(1) 33 45
22. RozenS
SkaletskyHJ
1998 Primer3. Code available at http://www-genome.wi.mit.edu/genome_software/other/primer3.html. Accessed at: http://biotools.umassmed.edu/bioapps/primer3_www.cgi
23. MortonNE
1956 Sequential tests for the detection of linkage. Amer. J. Hum. Genet 7 277 318
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2008 Číslo 9
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Missense Mutation in Exon 2 of SLC36A1 Responsible for Champagne Dilution in Horses
- Novel Mutations in (TDP-43) in Patients with Familial Amyotrophic Lateral Sclerosis
- Sex-Specific Genetic Structure and Social Organization in Central Asia: Insights from a Multi-Locus Study
- Chromosome Mechanics and Meiotic Engine Maintenance
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy