
Mutations in the SLC2A9 Gene Cause Hyperuricosuria and Hyperuricemia in the Dog
Allantoin is the end product of purine catabolism in all mammals except humans, great apes, and one breed of dog, the Dalmatian. Humans and Dalmatian dogs produce uric acid during purine degradation, which leads to elevated levels of uric acid in blood and urine and can result in significant diseases in both species. The defect in Dalmatians results from inefficient transport of uric acid in both the liver and renal proximal tubules. Hyperuricosuria and hyperuricemia (huu) is a simple autosomal recessive trait for which all Dalmatian dogs are homozygous. Therefore, in order to map the locus, an interbreed backcross was used. Linkage mapping localized the huu trait to CFA03, which excluded the obvious urate transporter 1 gene, SLC22A12. Positional cloning placed the locus in a minimal interval of 2.5 Mb with a LOD score of 17.45. A critical interval of 333 kb containing only four genes was homozygous in all Dalmatians. Sequence and expression analyses of the SLC2A9 gene indicated three possible mutations, a missense mutation (G616T;C188F) and two promoter mutations that together appear to reduce the expression levels of one of the isoforms. The missense mutation is associated with hyperuricosuria in the Dalmatian, while the promoter SNPs occur in other unaffected breeds of dog. Verification of the causative nature of these changes was obtained when hyperuricosuric dogs from several other breeds were found to possess the same combination of mutations as found in the Dalmatian. The Dalmatian dog model of hyperuricosuria and hyperuricemia underscores the importance of SLC2A9 for uric acid transport in mammals.
Published in the journal:
. PLoS Genet 4(11): e32767. doi:10.1371/journal.pgen.1000246
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1000246
Summary
Allantoin is the end product of purine catabolism in all mammals except humans, great apes, and one breed of dog, the Dalmatian. Humans and Dalmatian dogs produce uric acid during purine degradation, which leads to elevated levels of uric acid in blood and urine and can result in significant diseases in both species. The defect in Dalmatians results from inefficient transport of uric acid in both the liver and renal proximal tubules. Hyperuricosuria and hyperuricemia (huu) is a simple autosomal recessive trait for which all Dalmatian dogs are homozygous. Therefore, in order to map the locus, an interbreed backcross was used. Linkage mapping localized the huu trait to CFA03, which excluded the obvious urate transporter 1 gene, SLC22A12. Positional cloning placed the locus in a minimal interval of 2.5 Mb with a LOD score of 17.45. A critical interval of 333 kb containing only four genes was homozygous in all Dalmatians. Sequence and expression analyses of the SLC2A9 gene indicated three possible mutations, a missense mutation (G616T;C188F) and two promoter mutations that together appear to reduce the expression levels of one of the isoforms. The missense mutation is associated with hyperuricosuria in the Dalmatian, while the promoter SNPs occur in other unaffected breeds of dog. Verification of the causative nature of these changes was obtained when hyperuricosuric dogs from several other breeds were found to possess the same combination of mutations as found in the Dalmatian. The Dalmatian dog model of hyperuricosuria and hyperuricemia underscores the importance of SLC2A9 for uric acid transport in mammals.
Introduction
Uric acid is the predominant product of purine metabolism in humans, great apes and one breed of dog, the Dalmatian; all other mammals excrete allantoin. During primate evolution, urate oxidase (UOX), which catalyzes the oxidation of uric acid into allantoin, accumulated several independent nonsense mutations that led to its silencing and resulted in high serum and urine uric acid levels in humans and great apes [1],[2]. Huu in the Dalmatian results from a different cause [3],[4]. Uric acid freely circulates in the form of urate, the salt of uric acid, in the plasma where it serves as a free-radical scavenger. Although uric acid has evolved in humans to be the main product of purine metabolism, this change has had some negative effects. High levels of urate predispose humans to gout [5],[6]. In addition, uric acid levels have been correlated with hypertension, vascular disease and metabolic syndrome although it is unclear whether hyperuricemia is primary or secondary in these cases [7]–[10].
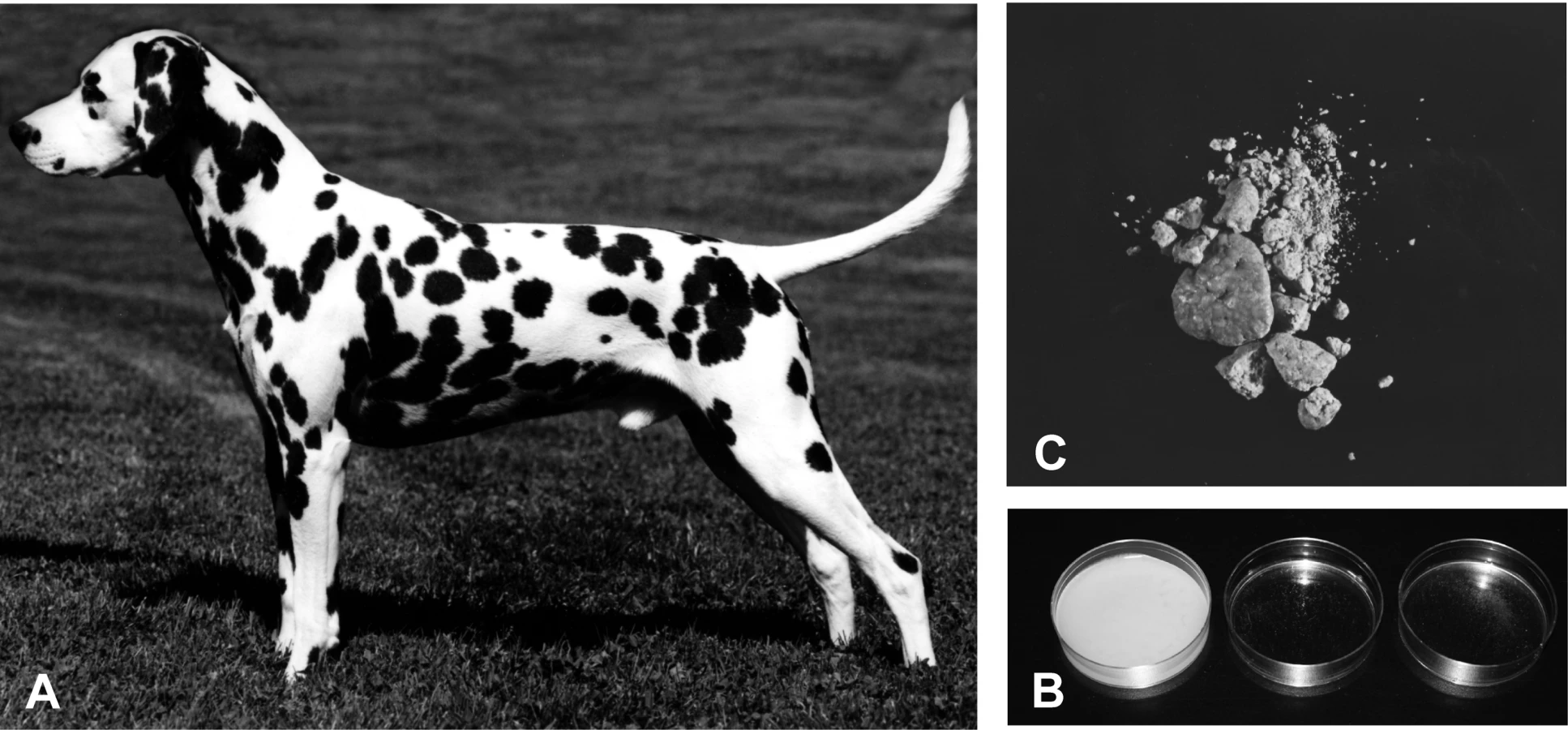
As in humans, all Dalmatian dogs have a defect in urinary metabolism that leads to excretion of uric acid rather than allantoin [11]. As a result, Dalmatians are predisposed to form urinary calculi composed of urate (Figure 1B). Hyperuricosuria in the Dalmatian is relatively easy to identify since Dalmatian urine forms a crystallized precipitate when cooled (Figure 1C). This trait was probably fixed in the breed through selection for a more distinctive spotting pattern [12],[13]. Dalmatian coat pattern involves mutations in at least three different spotting genes (Figure 1A). Dalmatians have a mutation for extreme white in the MITF gene [14] that leads to an all white coat. A dominant mutation, called T for ticking [15], is responsible for adding the pigmented spots to the white coat. Based on segregation analysis, the huu locus appears to be closely linked to a modifier of spot size [16].
In mammals that produce uric acid rather than allantoin, the level of uric acid in the blood is controlled by differences in production as well as differences in the amount that is excreted in the urine. In the kidney, uric acid is filtered by the glomerulus and then a portion is reabsorbed in the proximal tubules where it re-enters circulation. There are species-specific differences in the production of uric acid versus allantoin and the relative amounts of reabsorption and secretion in the proximal tubules, making the use of animal models in this area of research challenging [17].
Dogs and humans, unlike many other mammals, undergo bidirectional transport of urate along the nephron, which results in net reabsorption of urate from the glomerular filtrate. In Dalmatians, reabsorption is lost entirely and urate excretion equals or exceeds the glomerular filtration rate [18]. This change in uric acid excretion by the Dalmatian kidney is not secondary to hyperuricemia since non-Dalmatian dogs with artificially raised serum uric acid levels can only clear uric acid at ∼1/3 the rate of the Dalmatian [18]. Free-flow micropuncture experiments were used to demonstrate that in Dalmatian kidneys there is a deficiency of proximal tubular reabsorption of urate [19].
Although findings stated above implicate the kidney in Dalmatian huu, reciprocal liver and kidney transplant experiments between Dalmatian and non-Dalmatian dogs demonstrate that the liver is also important for the phenotype. Kidney transplants between normal dogs and Dalmatians only partially ameliorated the hyperuricosuria phenotype [20],[21]. However, Dalmatian hepatocyte transplants can correct the phenotype [20]. Therefore, a logical cause for the Dalmatian phenotype is a mutation in urate oxidase, similar to humans. However, Dalmatian liver homogenates are capable of oxidizing uric acid to allantoin and the urate oxidase gene was excluded genetically [3]. The Dalmatian phenotype could also be explained by a generalized defect of urate transport since liver slices were not capable of converting uric acid to allantoin [4]. Dalmatian dog erythrocytes have been shown to transport urate normally, demonstrating that Dalmatians do not have a generalized defect in urate transport [22]. Although Dalmatians have functional urate oxidase activity in their livers, they effectively have a similar phenotype to humans and great apes since they cannot transport urate into the liver for degradation. The Dalmatian phenotype can be summarized as a hepatic and renal urate transport defect which leads to hyperuricosuria and relative (compared to other dogs) hyperuricemia.
The discovery of various proteins that transport urate has shed some light on the control of serum and urine uric acid levels [23]. In the kidney, urate is transported across the apical membrane of the proximal tubules and then across the basolateral membrane before re-entering circulation. In humans, the transporter that reabsorbs urate across the apical membrane in the proximal tubule is Urat1, or SLC22A12 [24], which is expressed exclusively in the kidney. Mutations in SLC22A12 are thought to be the major cause of idiopathic renal hypouricemia in humans [24],[25], which is also called “Dalmatian hypouricemia” since people with this disorder spill uric acid into their urine resulting in a phenotype similar to the Dalmatian dogs [26]. In addition, recent work has shown an association of SLC2A9 with serum uric acid levels in several different populations [27],[28]. Variants in the non-coding region of SLC2A9 are associated with gout and uric acid levels in several human populations [29]–[31]. SLC2A9 has been shown by Xenopus oocyte experiments to transport uric acid [30]. In humans and mice it is expressed in liver and kidney [32],[33]. In particular, in humans SLC2A9 isoforms have been localized to both the apical and basolateral membrane of the proximal tubules, allowing the possibility that SLC2A9 influences serum uric acid levels by transport in the kidney [34].
All Dalmatians are homozygous for huu, which is inherited as a simple autosomal recessive trait as demonstrated by crosses performed between Dalmatians and other breeds of dogs [12],[13]. In order to identify the causative gene, an interbreed backcross (Dalmatian×Pointer) was developed which introduced the wildtype version of the huu gene into the Dalmatian breed while maintaining the breed characteristics of the Dalmatian. Based on linkage analysis using this cross (LOD 6.55), huu localizes to CFA03, excluding SLC22A12 as a candidate [35]. Using recombination breakpoints in the interbreed backcross and taking advantage of the homozygosity within the Dalmatian breed for huu, a critical interval containing four candidate genes was defined. Microsatellite markers within this interval gave LOD scores over 17 for linkage to huu. A candidate causal missense mutation (C188F) was identified within a highly conserved transmembrane (TM5) of the 12 transmembrane transporter protein, SLC2A9. The missense mutation is homozygous in all Dalmatians tested (247) as well as in hyperuricosuric dogs of other breeds.
Results
Linkage analysis using a Dalmatian×Pointer backcross family localized Dalmatian huu to CFA03. Haplotype analysis defined a 3.3 Mb critical interval (CFA03 72,063,073–75,355,028 Mb), estimated to contain 24 candidate genes [35]. Additional backcross dogs were genotyped with the microsatellites used for haplotype analysis and with new microsatellites mined from the critical region on CFA03. The full pedigree of all the dogs used for this analysis is shown in Figure S1. Urine uric acid/creatinine ratios were used to categorize the dogs' genotypes at huu. LOD scores were determined for a subset of these markers. Two markers within the critical interval defined by recombination breakpoints gave LOD scores over 17, further confirming the linkage to this region. Recombination events in two dogs narrowed the critical interval to a 2.5 Mb region containing 19 candidate genes (CFA03; 71,796,048–74,348,350 Mb) (Figure 2).

Since Dalmatians are fixed for huu, it was expected that an area of homozygosity around the huu locus would be identified. Blocks of linkage disequilibrium (LD) in purebred dogs extend between several megabases in rare breeds with small population sizes to several kilobases in popular breeds with larger population sizes [14],[36]. Dalmatians have a moderate population size, so the homozygous region surrounding huu should be smaller than 2.5 Mb. Microsatellites mined from the canine genome and located ∼100 Kb apart were genotyped in the backcross dogs. Markers that were homozygous in huu/huu individuals and heterozygous in huu/+ individuals were typed in 24 unrelated Dalmatians to verify that the region of homozygosity is not the result of familial linkage disequilibrium. Haplotypes were constructed to rule out the possibility of more than a single ancestral mutation. Regions of homozygosity were found between ms173 and ms9 (CFA03; 71,796,048–72,363,187) and ms187 and ms128 (CFA03; 74,183,928–74,245,743). Eighteen SNPs, extending across 2.5 Mb, were genotyped in 24 Dalmatians, a wild-type Labrador Retriever and a huu/+ backcross dog. The results excluded the area between ms187 and ms128 and a total of 13 SNPs spanning ∼333 Kb confirmed the homozygous region between ms173 and ms9 in the 24 Dalmatians (Figure 3). These SNPs are also heterozygous in the huu/+ backcross dog, polymorphic in two unaffected dogs (ND1 and ND2) and the Boxer genome assembly sequence.

Four candidate genes were identified in the region of homozygosity, LOC488823 (similar to mast cell immunoreceptor signal transducer – MIST), LOC479092 (zinc finger protein 518B, KIAA1729), LOC611070 (similar to WD-repeat protein 1 – WDR1) and LOC479093 (similar to solute carrier family 2, member 9 protein, isoform 1 – SLC2A9) (Figure 3). RT-PCR established that all four genes were expressed in canine liver and kidney. Since all the candidates were expressed in the appropriate organs, all four genes were sequenced from Dalmatian and non-Dalmatian liver cDNA samples and the untranslated regions (UTRs) were determined by 5′ and 3′ RACE (Genbank EU371511–EU371515).
The 5′UTR and exons 1–7 of MIST were outside of the LD region and were not pursued. A single silent mutation was identified in exon 16 of the MIST gene (T951C). Three silent mutations were found in the single exon of the canine KIAA1729 gene (T932C, C2480T, G3092A) and a 33 bp insertion/deletion was discovered in the 3′UTR. Both alleles of these three SNPs and the insertion/deletion were seen in unaffected non-Dalmatian dogs. WDR1 was sequenced in liver cDNA and genomic DNA from a Dalmatian and a non-Dalmatian. A single SNP was found in intron 9 that does not affect a splice site. Although SLC2A9 did not have an assigned function related to uric acid metabolism at the time this work was performed, it was considered to be the most promising candidate since it is a transporter protein.
A discrepancy in SLC2A9 exon annotation between NCBI and the UC Santa Cruz genome browser was addressed by 5′ RACE. Two SLC2A9 exon 1 variants were found in a Dalmatian and a wild-type Golden Retriever, variant O (CFA03 : 72,222,637–72,416,753) and variant N (CFA03 : 72,227,605–72,416,753). The difference between the two variants lies in the first 28 amino acids of the N terminus, similar to known mouse and human variants. Both transcripts were shown to be expressed in canine liver and kidney by RT-PCR. Expression differences were observed for variant O between Dalmatian and non-Dalmatian in both liver and kidney (equivalent to isoform 2 in human) but not for variant N (Figure 4A). Expression in the Dalmatian samples was ∼50% of non-Dalmatian levels in both tissues. SLC2A9 expression was further evaluated by RT-PCR using primers that amplify both transcripts in 11 different tissues from an unaffected Beagle. The highest levels of expression were observed in the kidney and liver (Figure 4B).

Sequencing of the SLC2A9 gene was performed on canine liver cDNA as well as genomic DNA so that intron/exon boundaries and promoter regions could be evaluated. SLC2A9 coding exons 2–12, as well as the exon-intron boundaries, were sequenced in a Dalmatian and a Labrador Retriever. Six SNPs were discovered in the SLC2A9 sequence. Two are exonic; exon 5 G563T;Cys188Phe (nucleotide and amino acid numbering are reported with reference to variant N) and exon 11 G1303A;Val435Ile (nucleotide and amino acid numbering are reported with reference to variant N), two are located 99 and 101 bp 5′ to the start codon of variant O. SNPs were also identified in introns 1 and 10. None of the intronic SNPs are within or near conserved splice-site elements. The SLC2A9 exon 11 G1303A SNP is polymorphic in the 24 unrelated Dalmatians tested, consistent with its genomic location outside of the region homozygous in Dalmatians (SNP17, Figure 3). The polymorphisms located 5′ to the start codon were tested in a panel of DNA samples from 15 unaffected dogs from various different breeds. Both SNPs are polymorphic in unaffected non-Dalmatian dogs, displaying both the Dalmatian haplotype (A–C) and other combinations. These SNPs are always fixed in affected Dalmatian dogs (SNP9-10, Figure 3). Primary sequence data from the exon 5 SNP is shown in Figure 5A.

The two non-synonymous SNPs found in SLC2A9 coding sequence were tested using the PANTHER program (http://www.pantherdb.org/tools/), which assigned the Cys188Phe substitution a score of −4.047 (scores range between 0 to −10, −10 indicating the most deleterious) with a probability that it is a deleterious substitution of 0.74 [37]. The Val435Ile variation was not given a prediction since the substitution is not conserved in other species and unlikely to be deleterious. The SIFT program also scored the Cys188Phe missense mutation as deleterious with a probability of 0.01, where any probability below 0.05 is considered deleterious [38]. In order to evaluate the degree of protein conservation in the region flanking the candidate SNP, SLC2A9 protein sequences were compared between chicken, mouse, rat, human, Boxer and Dalmatian (Figure 4B). This region of the protein has a high degree of identity across mammals. The variant O promoter SNPs (located 99 and 101 base pairs upstream of the initiator methionine) were analyzed for transcription factor binding site differences and the SNPs are predicted to disrupt binding sites for DeltaE, AML-1a, S8 and Cre-BP in the Dalmatian version.
The SLC2A9 exon 5 C188F amino acid substitution was further tested in 247 Dalmatians and 378 non-Dalmatian dogs from 58 different breeds. It was homozygous in all Dalmatians tested and only the wildtype allele was present in the non-Dalmatian dogs.
Individual dogs from non-Dalmatian breeds are known to form urate calculi and have been diagnosed with huu [39],[40]. Two Dogs from two of these breeds, Bulldogs and Black Russian Terriers, that had formed urinary urate calculi were tested for the SLC2A9 exon 5 G563T;Cys188Phe missense mutation (SNP 15 in Figure 3) and the variant O promoter SNPS (SNP 9, 10 in Figure 3) and found to be homozygous. These four dogs shared the Dalmatian haplotype across the homozygous interval defined during the positional cloning of the huu locus (Figure 3).
Discussion
The Dalmatian dog exhibits hyperuricosuria and relative hyperuricemia due to a defect of urate transport in the liver and kidney. Positional cloning of the huu locus using an interbreed backcross, as well as homozygosity within the Dalmatian breed, has identified SLC2A9 as the cause of the change in uric acid handling by this dog breed. Unrelated breeds of dog with hyperuricosuria or urate stone disease share the same haplotype as Dalmatian dogs, providing compelling evidence that this is the gene responsible. This result was somewhat unexpected because SLC2A9 was classified as a member of the large glucose transporter family and did not have an assigned function with respect to urate transport until recently.
SLC2A9 is classified as a part of the large glucose transporter family based on amino acid sequence identity of 44% and 38% to Glut5 and Glut1, respectively [32]. SLC2A9 has been localized to the cell surface in humans [34]. It contains 12 transmembrane domains and, based on homology to other glucose transporters, the central channel is essentially formed by helices 2, 4, 5, 7, 8, and 10 [41]. The Cys188Phe amino acid change occurs within a highly conserved residue located within the fifth transmembrane domain of the protein. Although a cysteine to phenylalanine amino acid change is not expected to disrupt the localization of the alpha helix to the transmembrane, this change could disrupt the proper function of the protein by altering the pore. Precedent for single amino acid changes altering substrate specificity has been shown for a number of the SLC2A transporters [42].
Expression differences between Dalmatian and non-Dalmatian samples were observed for one of the isoforms (O) of the gene (Figure 4). SNPs in the promoter region of this variant were identified and were fixed in the Dalmatian breed. These same SNPs were also identified in non-Dalmatians without hyperuricosuria. Therefore, these SNPs alone are not sufficient to confer the phenotype but along with the missense mutation they may contribute to the expression of the phenotype. It remains to be determined if there are variant O transcript differences in the general dog population and if the SNPs identified in this work are responsible for the differences in the level of variant O expression. In humans, the equivalent isoform to variant O is expressed on the apical surface of the proximal tubules [34]. Reduction in the amount of the protein combined with a deficiency in the protein itself may contribute to the decreased conversion to allantoin by the liver as well as the increased excretion of uric acid in these dogs. SLC2A9 tissue expression in dogs is similar to that observed in humans and mice. Expression in humans is highest in kidney, liver and placenta [32]. In humans, isoform 1 has been localized within polarized canine kidney cells to the basolateral membrane of the proximal tubules while isoform 2 is localized to the apical side [34]. The tissue and cellular localization of SLC2A9 is consistent with the Dalmatian huu phenotype and with SLC2A9 having a role in urate transport in both the liver and kidney.
Mutations in SLC2A9 will likely have important consequences for a number of different disorders of uric acid homeostasis in people. Significant alterations of SLC2A9 could cause primary renal hypouricemia in people similar to SLC22A12 mutations since genetic heterogeneity exists for this disorder [43]–[45]. Mutations in SLC2A9 could also cause cases of primary gout by significantly altering the serum uric acid concentration. In addition, more subtle changes could alter serum uric acid levels by changing the amount of uric acid excreted in the urine. There is evidence that polymorphisms in SLC22A12 are capable of increasing serum uric acid levels in Japanese populations [46]. A significant association of SLC2A9 to serum uric acid levels was recently reported among Caucasian individuals [28] and among Sardinian and Chianti cohorts [27]. Two other papers were recently published on the importance of the SLC2A9 gene to urate transport in humans. The first demonstrated that human SLC2A9 is a high capacity, low affinity uric acid transporter and that genetic variants are associated with gout; however causative mutations were not determined [30]. The second paper documented a strong association between SLC2A9 and uric acid levels in cohorts of German and Austrian nationalities as well as a correlation between RNA expression levels of this gene and serum uric acid levels [29]. Thus, strong evidence exists that SLC2A9 functions as a urate transporter in humans, and likely in dogs.
In addition to the potential influence with respect to human uric acid disorders, the present studies also impact our understanding of the history of this defect in dogs. The story of hyperuricosuria in dogs started in 1916 when Benedict first recognized the similarity of uric acid defects in the Dalmatian dog and people [11]. However, the origin of this defect may have preceded the development of the Dalmatian breed. The present studies show that the same genetic mutation is present in Bulldogs and Black Russian Terriers, breeds that are not known to be closely related to the Dalmatian. It appears that affected individuals from these breeds share the same haplotype as the Dalmatian, indicating that the mutation is identical by descent between these breeds. The mutation must be quite old since it would have to predate breed formation; however, additional evaluation of the extended haplotype in all three breeds is necessary to estimate an actual age. Alternatively, although unlikely, the mutation could have been introduced into these other breeds by crosses between breeds. Although Dalmatians are fixed for hyperuricosuria, this is not true of the other breeds. Therefore, genetic testing and selection in those breeds can eliminate the disease. Within the Dalmatian breed, the possibility exists for correction of this defect by the introduction of unaffected Dalmatian×Pointer backcross dogs into the purebred gene pool. These dogs are currently registered with the United Kennel Club in the United States. The disease allele probably became homozygous in modern Dalmatians through selection for more distinctive spots. However, most low uric acid excreting backcross dogs have acceptably sized spots (according to the breed standard), allowing breeders the unique opportunity to correct a fixed genetic defect while maintaining the breed characteristic that may ultimately be responsible for its fixation.
Two independent lines of evidence from different species point to the key role of SLC2A9 in urate transport; the Dalmatian uric acid phenotype itself and genome wide association studies linking SLC2A9 to uric acid levels in people along with direct uric acid transport data. There are many questions to be answered about the role of SLC2A9 in urate homeostasis and its other transport functions. In dogs and other mammals with endogenously low serum uric acid, SLC2A9 may normally play a different transport role than in people where it appears to have an important function in uric acid transport. Since degradation of uric acid to allantoin does not occur in humans and great apes, SLC2A9 may play a different role in transport in the liver as compared to those species that excrete allantoin. The positional cloning of the hyperuricosuria locus in the Dalmatian dog has provided a compelling new avenue of investigation toward a better understanding of urate transport in mammals and successfully completes a story started in 1916 when Benedict first recognized the similarity of the uric acid defect in the Dalmatian dog and people.
Materials and Methods
DNA samples from backcross dogs were acquired as previously described [35]. The full pedigree of the dogs used in this analysis is shown in Figure S1. Blood, buccal swab and DNA samples from Dalmatians and non-Dalmatian dogs were obtained from patients of the Veterinary Medical Teaching Hospital at UC Davis, the UC Davis Veterinary Genetics Laboratory, Dr. Gary Johnson at the University of Missouri, Columbia, and from private owners. The use of animals in this research was approved by the University of California, Davis animal care and use committee (protocol #11962). Urine uric acid and creatinine was measured in 3–7 week old puppies as previously described [13].
LOD scores were calculated as previously described [35]. Primers for microsatellites spaced on average 100 Kb apart in the 3.3 Mb candidate region of CFA03 were identified using the May 2005 CanFam2.0 sequence assembly on the UCSC genome browser and designed within sequence flanking each repeat using the Primer3 program (Table S1) [47].
Eight SNPs, which are part of the Affymetrix canine SNP array, were chosen because they are polymorphic in the general dog population. Additional SNPs were mined from the canine genome sequence or identified during sequencing of the candidate genes. Primers for SNPs (Table S2) were designed as described above. SNP and microsatellite sequences were PCR amplified and genotyped in the backcross dogs and only informative markers for this family were then genotyped in 24 unrelated purebred Dalmatians.
Human mRNA sequences were obtained from GenBank for each of the candidate genes (MIST NM_052964.1, KIAA1729 NM_053042, WDR1 NM_005112.4, SLC2A9 AF210317). These sequences were compared to the UCSC Genome Browser annotation of the canine genome using the BLAT function to obtain canine sequence for each gene. The Primer3 program was used to design primers for the canine sequences (Table S3). PCR reactions, genotyping and sequencing were done as previously described [35]. Sequences were visualized using Chromas2 (Technelysium, Tewantin, QLD, Australia) and analyzed with Vector NTI software (Informax, Frederick, MD, USA). TFsearch Version 1.3 was used to analyze the Variant O promoter SNPs affect on transcription factor binding sites.
5′ and 3′ RACE were performed for 3 of the 4 candidate genes. RACE primers (Table S3) were designed as described above and RACE products amplified with the SMART RACE cDNA Amplification Kit (Clontech, Mountain View, CA, USA) and cloned using the TOPO TA Cloning kit (pCR2.1-TOPO vector) with One Shot TOP10 Chemically Competent E. coli (Invitrogen, Carlsbad, CA, USA). Products were isolated with the Qiaprep Spin Miniprep kit (Qiagen, Valencia, CA, USA) and sequenced as described above. Genbank accession numbers for the transcripts are EU371511–EU371515. All genomic locations given in the text are based on the May 2005 CanFAm2.0 genome assembly and are viewed using the UCSC genome browser.
RNA was isolated from kidney and liver samples with the Micro-FastTrack 2.0 mRNA isolation kit (Invitrogen, Carlsbad, CA, USA). cDNA was synthesized with the SuperScript III First-Strand Synthesis System for RT-PCR (Invitrogen, Carlsbad, CA, USA). Expression was evaluated for LOC479092, LOC611070 and both variants of LOC479093 from liver and kidney from Dalmatians and unaffected Newfoundland×Border Collie crosses using primers SLC5′UTREx1VarNF and SLCR1 for variant N and primers IntF-Ex1 and SLCR1 for variant O. The expression of LOC479093 was also evaluated in an array of tissues from an unaffected Beagle (cerebellum, cerebral cortex, heart, kidney, liver, skeletal muscle, skin, spinal cord, spleen, testis and thymus) using primers IntF5 and SLCdnaR. RNAs for these tissues were acquired from Zyagen (San Diego, CA, USA) and cDNA synthesized as described. GAPDH was amplified (F primer-5′AAGATTGTCAGCAATGCCTCC3′, R primer -5′CCAGGAAATGAGCTTGACAAA3′) in these tissues to ensure that equivalent amounts of cDNA were added.
SLC2A9 genotypes were determined by a restriction fragment length polymorphism (RFLP) assay. PCR products were produced as described previously using an unlabeled forward primer, 5′-TGCTTCTCTGAAATTTACCTCCA – 3′ and a fluorescently labeled reverse primer, 5′-6FAM-CGAGAGGATGGTATACGGTGA -3′ (Applied Biosystems, Foster City, CA). Products were then digested with the enzyme HpyCH4V (New England Biolabs, Ipswitch, MA) for 1 hour at 37°C. Digestions were analyzed on an ABI 3100 Genetic Analyzer with GeneScan 400HD Rox size standard. A 79 base pair unlabeled product is generated from the 440 bp product by a control cut site. The A allele produces a labeled 361 base pair product and the G allele produces a labeled 106 base pair product.
Supporting Information
{kind=link}
Zdroje
1. FriedmanTB
PolancoGE
AppoldJC
MayleJE
1985 On the loss of uricolytic activity during primate evolution—I. Silencing of urate oxidase in a hominoid ancestor. Comp Biochem Physiol B 81 653 659
2. OdaM
SattaY
TakenakaO
TakahataN
2002 Loss of urate oxidase activity in hominoids and its evolutionary implications. Mol Biol Evol 19 640 653
3. SafraN
LingGV
SchaibleRH
BannaschDL
2005 Exclusion of urate oxidase as a candidate gene for hyperuricosuria in the Dalmatian dog using an interbreed backcross. J Hered 96 750 754
4. GieseckeD
TiemeyerW
1984 Defect of uric acid uptake in Dalmatian dog liver. Experientia 40 1415 1416
5. ChoiHK
MountDB
ReginatoAM
2005 Pathogenesis of gout. Ann Intern Med 143 499 516
6. LinKC
LinHY
ChouP
2000 The interaction between uric acid level and other risk factors on the development of gout among asymptomatic hyperuricemic men in a prospective study. J Rheumatol 27 1501 1505
7. ShortRA
TuttleKR
2005 Clinical evidence for the influence of uric acid on hypertension, cardiovascular disease, and kidney disease: a statistical modeling perspective. Semin Nephrol 25 25 31
8. HeinigM
JohnsonRJ
2006 Role of uric acid in hypertension, renal disease, and metabolic syndrome. Cleve Clin J Med 73 1059 1064
9. FeigDI
MazzaliM
KangDH
NakagawaT
PriceK
2006 Serum uric acid: a risk factor and a target for treatment? J Am Soc Nephrol 17 S69 S73
10. ChoiHK
FordES
2007 Prevalence of the metabolic syndrome in individuals with hyperuricemia. Am J Med 120 442 447
11. BenedictSR
1916 The Harvey Lectures. Journal of Laboratory and Clinical Medicine 1 346
12. KeelerCE
1940 The Inheritance of Predisposition to Renal Calculi in the Dalmatian. Journal of American Veterinary Medical Association 96 507 510
13. SchaibleRH
1986 Genetic predisposition to purine uroliths in Dalmatian dogs. Vet Clin North Am Small Anim Pract 16 127 131
14. KarlssonEK
BaranowskaI
WadeCM
Salmon HillbertzNH
ZodyMC
2007 Efficient mapping of mendelian traits in dogs through genome-wide association. Nat Genet 39 1321 1328
15. LittleC
1957 The Inheritance of coat color in dogs New York Howell Book House 194
16. SchaibleR
1976 Linkage of a pigmentary trait with a high level of uric acid excretion in the Dalmatian dog. Genetics 83 S68
17. Roch-RamelF
PetersG
1978 Urinary excretion of uric acid in nonhuman mammalian species.
WeinerKWIM
Uric Acid Berlin Springer-Verlag 211 255
18. FriedmanMaSOB
1948 Observations concerning the casues of the excess excretion of uric acid in the Dalmatian Dog. The Journal of Biological Chemistry 175 727 735
19. Roch-RamelF
WongNL
DirksJH
1976 Renal excretion of urate in mongrel and Dalmatian dogs: a micropuncture study. Am J Physiol 231 326 331
20. KusterG
ShorterR
DawsonB
HallenbeckG
1972 Uric acid metabolism in dalmatians and other dogs. Role of the liver. Archives of Internal Medicine 129 492 496
21. ApplemanR
HallenbeckG
ShorterRG
1966 Effect of reciprocal allogeneic renal transplantation between Dalmatian and non-dalmatian dogs on urinary excretion of uric acid. Proceedings of the Society of Experimental Biological Medicine 121 1094 1097
22. VinayP
GattereauA
MoulinB
GougouxA
LemieuxG
1983 Normal urate transport into erythrocytes in familial renal hypouricemia and in the Dalmatian dog. Can Med Assoc J 128 545 549
23. HedigerMA
JohnsonRJ
MiyazakiH
EndouH
2005 Molecular physiology of urate transport. Physiology (Bethesda) 20 125 133
24. EnomotoA
KimuraH
ChairoungduaA
ShigetaY
JutabhaP
2002 Molecular identification of a renal urate anion exchanger that regulates blood urate levels. Nature 417 447 452
25. TanakaM
ItohK
MatsushitaK
WakitaN
AdachiM
2003 Two male siblings with hereditary renal hypouricemia and exercise-induced ARF. Am J Kidney Dis 42 1287 1292
26. GreeneML
MarcusR
AurbachGD
KazamES
SeegmillerJE
1972 Hypouricemia due to isolated renal tubular defect. Dalmatian dog mutation in man. Am J Med 53 361 367
27. LiS
SannaS
MaschioA
BusoneroF
UsalaG
2007 The GLUT9 Gene Is Associated with Serum Uric Acid Levels in Sardinia and Chianti Cohorts. PLoS Genet 3 e194 doi:10.1371/journal.pgen.0030194
28. WallaceC
NewhouseSJ
BraundP
ZhangF
TobinM
2008 Genome-wide association study identifies genes for biomarkers of cardiovascular disease: serum urate and dyslipidemia. Am J Hum Genet 82 139 149
29. DoringA
GiegerC
MehtaD
GohlkeH
ProkischH
2008 SLC2A9 influences uric acid concentrations with pronounced sex-specific effects. Nat Genet 40 430 436
30. VitartV
RudanI
HaywardC
GrayNK
FloydJ
2008 SLC2A9 is a newly identified urate transporter influencing serum urate concentration, urate excretion and gout. Nat Genet 40 437 442
31. BrandstatterA
KiechlS
KolleritsB
HuntSC
HeidIM
2008 Sex-specific association of the putative fructose transporter SLC2A9 variants with uric acid levels is modified by BMI. Diabetes Care 31 1662 1667
32. PhayJE
HussainHB
MoleyJF
2000 Cloning and expression analysis of a novel member of the facilitative glucose transporter family, SLC2A9 (GLUT9). Genomics 66 217 220
33. KeembiyehettyC
AugustinR
CarayannopoulosMO
SteerS
ManolescuA
2006 Mouse glucose transporter 9 splice variants are expressed in adult liver and kidney and are up-regulated in diabetes. Mol Endocrinol 20 686 697
34. AugustinR
CarayannopoulosMO
DowdLO
PhayJE
MoleyJF
2004 Identification and characterization of human glucose transporter-like protein-9 (GLUT9): alternative splicing alters trafficking. J Biol Chem 279 16229 16236
35. SafraN
SchaibleRS
BannaschDL
2006 Linkage analysis with an interbreed backcross maps Dalmatian hyperuricosuria to CFA03. Mammalian Genome 17 340 345
36. Lindblad-TohK
WadeCM
MikkelsenTS
KarlssonEK
JaffeDB
2005 Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 438 803 819
37. ThomasPD
KejariwalA
CampbellMJ
MiH
DiemerK
2003 PANTHER: a browsable database of gene products organized by biological function, using curated protein family and subfamily classification. Nucleic Acids Res 31 334 341
38. NgPC
HenikoffS
2001 Predicting deleterious amino acid substitutions. Genome Res 11 863 874
39. BendeB
NemethT
2004 High prevalence of urate urolithiosis in the Russian black terrier. Vet Rec 155 239 240
40. LingGV
FrantiCE
RubyAL
JohnsonDL
1998 Urolithiasis in dogs. II: Breed prevalence, and interrelations of breed, sex, age, and mineral composition. Am J Vet Res 59 630 642
41. Salas-BurgosA
IserovichP
ZunigaF
VeraJC
FischbargJ
2004 Predicting the three-dimensional structure of the human facilitative glucose transporter glut1 by a novel evolutionary homology strategy: insights on the molecular mechanism of substrate migration, and binding sites for glucose and inhibitory molecules. Biophys J 87 2990 2999
42. ManolescuAR
WitkowskaK
KinnairdA
CessfordT
CheesemanC
2007 Facilitated hexose transporters: new perspectives on form and function. Physiology (Bethesda) 22 234 240
43. IchidaK
HosoyamadaM
HisatomeI
EnomotoA
HikitaM
2004 Clinical and molecular analysis of patients with renal hypouricemia in Japan—influence of URAT1 gene on urinary urate excretion. J Am Soc Nephrol 15 164 173
44. SperlingO
2006 Hereditary renal hypouricemia. Mol Genet Metab 89 14 18
45. TzovarasV
ChatzikyriakidouA
BairaktariE
LiberopoulosEN
GeorgiouI
2007 Absence of SLC22A12 gene mutations in Greek Caucasian patients with primary renal hypouricaemia. Scand J Clin Lab Invest 67 589 595
46. ShimaY
TeruyaK
OhtaH
2006 Association between intronic SNP in urate-anion exchanger gene, SLC22A12, and serum uric acid levels in Japanese. Life Sci 79 2234 2237
47. RozenS
SkaletskyH
2000 Primer3 on the WWW for general users and for biologist programmers. Methods Mol Biol 132 365 386
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2008 Číslo 11
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Mutations in the SLC2A9 Gene Cause Hyperuricosuria and Hyperuricemia in the Dog
- Spindle Positioning, Meiotic Nonreduction, and Polyploidy in Plants
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy