
Sequencing of Loci from the Elephant Shark Reveals a Family of Genes in Vertebrate Genomes, Forged by Ancient Duplications and Divergences
Pax6 is a developmental control gene essential for eye development throughout the animal kingdom. In addition, Pax6 plays key roles in other parts of the CNS, olfactory system, and pancreas. In mammals a single Pax6 gene encoding multiple isoforms delivers these pleiotropic functions. Here we provide evidence that the genomes of many other vertebrate species contain multiple Pax6 loci. We sequenced Pax6-containing BACs from the cartilaginous elephant shark (Callorhinchus milii) and found two distinct Pax6 loci. Pax6.1 is highly similar to mammalian Pax6, while Pax6.2 encodes a paired-less Pax6. Using synteny relationships, we identify homologs of this novel paired-less Pax6.2 gene in lizard and in frog, as well as in zebrafish and in other teleosts. In zebrafish two full-length Pax6 duplicates were known previously, originating from the fish-specific genome duplication (FSGD) and expressed in divergent patterns due to paralog-specific loss of cis-elements. We show that teleosts other than zebrafish also maintain duplicate full-length Pax6 loci, but differences in gene and regulatory domain structure suggest that these Pax6 paralogs originate from a more ancient duplication event and are hence renamed as Pax6.3. Sequence comparisons between mammalian and elephant shark Pax6.1 loci highlight the presence of short - and long-range conserved noncoding elements (CNEs). Functional analysis demonstrates the ancient role of long-range enhancers for Pax6 transcription. We show that the paired-less Pax6.2 ortholog in zebrafish is expressed specifically in the developing retina. Transgenic analysis of elephant shark and zebrafish Pax6.2 CNEs with homology to the mouse NRE/Pα internal promoter revealed highly specific retinal expression. Finally, morpholino depletion of zebrafish Pax6.2 resulted in a “small eye” phenotype, supporting a role in retinal development. In summary, our study reveals that the pleiotropic functions of Pax6 in vertebrates are served by a divergent family of Pax6 genes, forged by ancient duplication events and by independent, lineage-specific gene losses.
Published in the journal:
. PLoS Genet 9(1): e32767. doi:10.1371/journal.pgen.1003177
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1003177
Summary
Pax6 is a developmental control gene essential for eye development throughout the animal kingdom. In addition, Pax6 plays key roles in other parts of the CNS, olfactory system, and pancreas. In mammals a single Pax6 gene encoding multiple isoforms delivers these pleiotropic functions. Here we provide evidence that the genomes of many other vertebrate species contain multiple Pax6 loci. We sequenced Pax6-containing BACs from the cartilaginous elephant shark (Callorhinchus milii) and found two distinct Pax6 loci. Pax6.1 is highly similar to mammalian Pax6, while Pax6.2 encodes a paired-less Pax6. Using synteny relationships, we identify homologs of this novel paired-less Pax6.2 gene in lizard and in frog, as well as in zebrafish and in other teleosts. In zebrafish two full-length Pax6 duplicates were known previously, originating from the fish-specific genome duplication (FSGD) and expressed in divergent patterns due to paralog-specific loss of cis-elements. We show that teleosts other than zebrafish also maintain duplicate full-length Pax6 loci, but differences in gene and regulatory domain structure suggest that these Pax6 paralogs originate from a more ancient duplication event and are hence renamed as Pax6.3. Sequence comparisons between mammalian and elephant shark Pax6.1 loci highlight the presence of short - and long-range conserved noncoding elements (CNEs). Functional analysis demonstrates the ancient role of long-range enhancers for Pax6 transcription. We show that the paired-less Pax6.2 ortholog in zebrafish is expressed specifically in the developing retina. Transgenic analysis of elephant shark and zebrafish Pax6.2 CNEs with homology to the mouse NRE/Pα internal promoter revealed highly specific retinal expression. Finally, morpholino depletion of zebrafish Pax6.2 resulted in a “small eye” phenotype, supporting a role in retinal development. In summary, our study reveals that the pleiotropic functions of Pax6 in vertebrates are served by a divergent family of Pax6 genes, forged by ancient duplication events and by independent, lineage-specific gene losses.
Introduction
Development is critically dependent on a core set of developmental regulator genes, most of which are highly conserved across metazoans and carry out pleiotropic functions as part of multiple gene regulatory networks. Variation in the functional output from these genes is an important factor in evolutionary divergence between species, yet their essential and pleiotropic role precludes dramatic changes in their primary sequence. Instead it is believed that duplication of key developmental control gene loci, in combination with variation in their spatio-temporal domains and expression levels are major components of this process [1]–[4]. Duplication of gene loci provides an initial freedom from selective pressure to allow divergence between gene loci.
Pax6 is a developmental control gene with an essential function in the development of eyes throughout the animal kingdom [5], [6]. Its ability to induce the full program for eye formation from ocular and non-ocular imaginal discs in Drosophila embryos has revealed it as the first of a small set of master regulators for eye development [7]–[9]. In vertebrates the key role of Pax6 in eye formation is equally well established. In addition Pax6 plays important roles in development and maintenance of the endocrine pancreas, the olfactory system and the central nervous system (CNS) where it is required for multiple cellular processes including maintenance of the neuronal progenitor pool at early developmental stages, and neurogenesis at later stages [10]. It is also required for cell migration and axon guidance in parts of the brain (reviewed in [11], [12]). In humans heterozygous disruption of the gene gives rise to the congenital eye malformation aniridia through haploinsufficiency, in some cases accompanied by additional phenotypes such as epilepsy, defective interhemispheric auditory transfer, anosmia or diabetes [13]–[16], while homozygous loss of gene function is incompatible with life [17]. In mice and rats heterozygous mutants have small eyes and exhibit many of the same features as found in aniridia patients [18], [19]. Homozygous mutants die at birth with severe brain malformation and complete lack of eyes and nasal structures [19]. Overexpression of the gene also causes eye malformations [20], [21], indicating that Pax6 dosage is critical for correct eye development.
In mammals a single Pax6 gene carries out the wide variety of developmental regulatory functions. This is achieved by strict control of its expression through a complex, extended cis-regulatory domain containing a large number of tissue-specific enhancers [22]–[29]. While some cis-elements are found upstream or within introns of the gene, most of the characterized long-range control elements are found in the downstream region. The importance of distant cis-regulatory elements for Pax6 gene expression has been highlighted by the existence of aniridia patients with chromosomal abnormalities (deletions/translocations) that separate these elements from the body of the gene [30]–[32]. Investigation of the locus beyond the patient breakpoints by DNaseI hypersensitivity mapping led to the identification of a region containing several cis-regulatory elements, embedded within the introns of an adjacent gene, Elp4, forcing synteny conservation between the genes [25], [33], [34].
In addition to a large array of distal enhancers, the single mammalian Pax6 gene uses at least three different promoters, P0, P1 and Pα, and the resulting transcripts produce three protein isoforms, Pax6, Pax6(+5a) and Pax6ΔPD [35]–[37]. The canonical Pax6 isoform encodes a paired and homeodomain containing transcription factor with a PST-rich transactivation domain at the C-terminus. Inclusion of an alternative exon, exon 5a, results in a protein with an interrupted paired domain that recognizes a different DNA binding sequence [35]. The paired-less ΔPD isoform is produced from a transcript initiating at an internal promoter Pα located in intron 4 of the gene [27]. The resulting protein contains the paired-type homeodomain and transactivation domain, but lacks the N-terminal paired domain. The function of this isoform is unknown, but overexpression has been shown to cause a microphthalmia phenotype in mice [24], [38], [36]. Apart from its DNA-binding capacity the homeodomain is also suggested to function in protein-protein interactions and could be involved in dimerisation [39].
In zebrafish the role of Pax6 is fulfilled by duplicate Pax6 genes, Pax6a and Pax6b, thought to have arisen by the fish-specific whole genome duplication event (FSGD). The FSGD is estimated to have taken place around 320 million years ago in the teleost fish lineage [40], and is often cited as the main contributing factor in the emergence of the large diversity of teleost fish, which make up nearly half of all vertebrate species [41]–[43]. The FSGD is a third genome wide duplication event (3R) that follows two earlier rounds (1R, 2R) of whole genome duplications (WGD) that occurred very early in the evolution of vertebrates [44], though debate about the nature and timing of these WGD events still continues.
As proposed nearly 40 years ago, genome duplication events are powerful drivers of evolution [45]. Genome or gene duplication events provide an initial freedom from selective pressure, and thus create an opportunity for modification or mutation of gene duplicates, as well as for alteration of the cis-regulatory landscape around the duplicate gene loci, while critical functions are maintained by the other copy under selective pressure. The various ways that can lead to retention of both gene copies following a duplication event are explained by the Duplication-Degeneration-Complementation (DDC) model [46]. Commonly, particularly in genes with a single function, one of the duplicates will start to accumulate mutations and degenerate beyond recognition over time, leaving the other copy to fulfill its single function (non-functionalization) [47]; Retention of both duplicates is more likely in pleiotropic genes, a category that includes many developmental regulatory genes. A number of scenarios can lead to such an outcome: one of the duplicates may acquire a novel function (neo-functionalization), the functions of the ancestral gene may become divided between the duplicates (sub-functionalization), or a combination of these (neo-subfunctionalization) [46]–[48]. The relative contribution of each of these possibilities is likely to be different for each gene locus and between species.
In many cases the functional divergence of gene duplicates is driven by changes in their cis-regulatory domains. In recent years identification of cis-regulatory elements has been greatly facilitated by comparative analysis of sequence conservation between distantly related species [49]. Compared to surrounding neutral sequences, sequences with important regulatory function are maintained under selective pressure and stand out as conserved non-coding elements (CNEs). However, functionally conserved regulatory regions with little or no sequence conservation have also been identified [28], [50], while sequence-similar enhancers can drive dissimilar expression patterns [51], [52] fuelling the debate on whether sequence conservation is necessary for functional conservation.
It is now thought that a massive appearance of novel CNEs has occurred early in the evolution of jawed vertebrates [53]. Comparisons with available sequence from the jawless vertebrate lamprey suggests it contains far fewer CNEs that are also shorter and less well conserved, while very few conserved non-coding elements can be identified in amphioxus [54], or ascidians (Ciona). In addition to the large-scale acquisition of ancient enhancers during the early gnathostome period [55], additional novel CNEs have been recruited at later stages in the evolution of specific lineages [56]. On the other hand, some ancient CNEs have been lost independently in different bony vertebrate lineages [57]. Thus absence of sequence conservation in teleosts at the position of a conserved element in the tetrapod locus could either indicate the loss of that element in fish species, or gain of the element in the tetrapod lineage.
Two Pax6 genes were previously identified in the zebrafish genome [58], and comparative analysis of their genomic loci suggested their evolution has largely followed the DDC model of divergence and complementation [59]. This led us to ask whether duplicate copies of Pax6 could also be identified in other teleost fish species and how the duplicate copies may have diverged in those species. Here we show that duplicate Pax6 genes are present in several other teleost species such as medaka, stickleback, fugu and Tetraodon (acanthopterygians). Examining the patterns of non-coding sequence conservation around the duplicate loci reveals a strong difference in the evolutionary divergence of the Pax6 loci in the acanthopterygians when compared to the zebrafish Pax6a/b divergence. While in zebrafish both loci have retained a large and overlapping portion of the cis-regulatory repertoire, the difference in the cis-regulatory domains of the duplicates in other teleosts is much more dramatic. Comparison with the mammalian locus indicates conservation of the ancient cis-regulatory landscape at the Pax6a loci, and a complete absence of conserved cis-elements at the Pax6b loci. In combination with a loss of the potential to encode the alternative exon5a this suggested a different evolutionary origin for the Pax6b loci in the acanthopterygians.
To relate our observations to a more ancestrally diverged species, we proceeded to obtain genomic sequence for the Pax6 locus of the elephant shark (Callorhinchus milii), a cartilaginous fish. Cartilaginous fishes are the most basal group of living jawed vertebrates and hence constitute a critical outgroup for bony vertebrates. The elephant shark is particularly attractive for comparative studies into the evolution of cis-regulatory landscapes in different jawed vertebrate lineages. Initial low coverage (∼1.4×) sequencing of its relatively small genome (910 Mb) revealed a greater complement of conserved non-coding sequences between the elephant shark and human genome than between human and zebrafish [60]. Surprisingly, screening of our elephant shark BAC library indicated the existence of a second Pax6 locus in this species. Sequence analysis of the BAC containing this locus revealed that this gene is a paired-less Pax6 gene, which we named Pax6.2. To determine whether the existence of this gene indicates a shark-specific duplication, or is the result of an ancient duplication event predating the split between cartilaginous fishes and bony vertebrates, we searched the genomes of other species for the presence of additional Pax6 genes using synteny relations from the elephant shark Pax6.2 locus. This approach revealed the presence of Pax6.2 orthologs in Xenopus, lizard, zebrafish and other teleosts. Thus the zebrafish genome contains 3 Pax6 genes. Further evidence for the shared ontogeny of these orthologs comes from identification and analysis of conserved cis-elements located near the genes. We show that these elements share both sequence and functional conservation, and the study reveals them as ancient enhancers linked to Pax6 regulation from the earliest stages of vertebrate evolution. Based on our phylogenetic analyses we present a model for the evolutionary history of the Pax6 gene family in vertebrates. In this model the two rounds of WGD at the base of the vertebrate lineage gave rise to four Pax6 genes. One of these was lost early on, while a variable subset of the remaining three has been retained in different species as a result of independent divergences and gene losses in different lineages. In the teleost lineage the FSGD produced further duplicates of the three Pax6 genes of which variable subsets remain in contemporary species.
Results
Different origins of the Pax6 duplicates of zebrafish versus acanthopterygian teleosts
To identify duplicate Pax6 genes in teleost fishes other than zebrafish, we searched the genomes of four other teleost fish species for which genomic sequences were available in the Ensembl database: medaka (Oryzias latipes), stickleback (Gasterosteus aculeatus), green spotted pufferfish (Tetraodon nigroviridis) and fugu (Takifugu rubripes). We found evidence for the presence of duplicated Pax6 gene loci in these four species of acanthopterygians. We compared the genes from these five teleost fish species with the Pax6 gene from a selection of other vertebrate species. Alignment of the amino acid sequences of the encoded proteins demonstrates a high level of similarity between the duplicated fish proteins and tetrapod Pax6 in all protein domains, including the paired domain, homeodomain and PST-rich transactivating regions (Figure 1A). The two major isoforms of full length Pax6 in tetrapods, Pax6 and Pax6(+5a), differ by a 14 amino acid insertion into the paired domain, encoded by an alternative exon 5a [35]. This alternative exon is present in the Pax6a gene of all five teleost fishes, and it is also found in the zebrafish Pax6b gene. Comparisons of the alternative exon between human and the fish species indicate significant variation between the peptide encoded by the exon 5a, both with respect to the amino acid sequence and length of the peptide (Figure 1A). However, while exon 5a is found in both zebrafish Pax6 duplicates, the exon is notably absent from the second Pax6 gene in the other four teleost species (Figure 1A). The region between exons 5 and 6 of the Pax6b genes in these fishes lacks any sequence homology to the exon, and manual searching of translated sequence in all reading frames failed to detect evidence for the presence of this exon. This indicates that the Pax6b genes of medaka, stickleback, fugu and Tetraodon do not have the ability to encode a Pax6(+5a) isoform, while both zebrafish Pax6a and Pax6b have retained this ability.
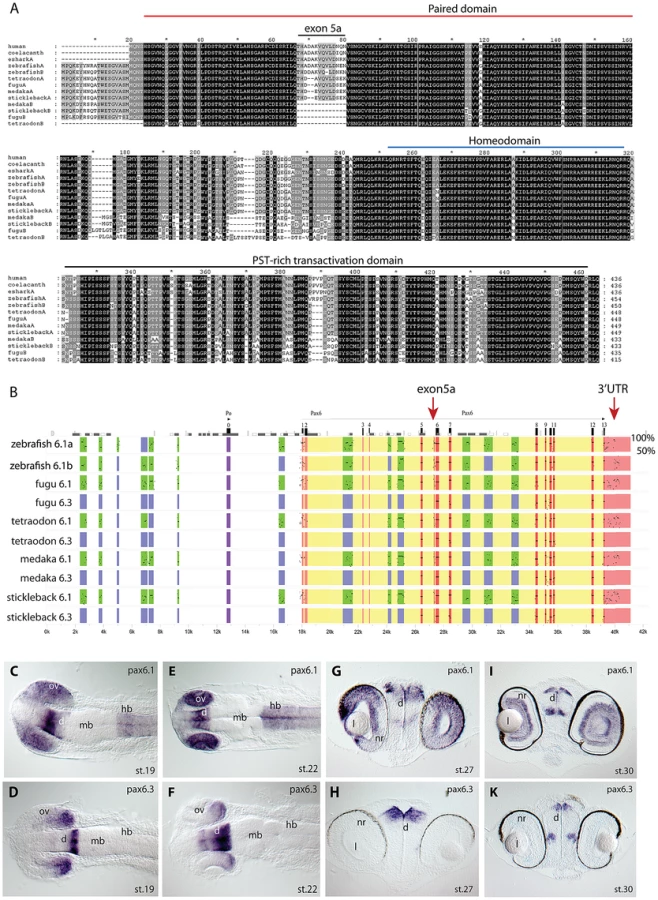
In tetrapods translation of full length Pax6 strictly starts at the ATG initiator codon located in exon 4. It has been noted that in teleost fish translation can also start in exon 2, creating an N-terminal extension to the encoded Pax6 protein [37], [58], [61]. Alignment of available ESTs and Genewise protein predictions shows most teleost Pax6a and Pax6b genes have the ability to code for this N-terminal protein extension, suggesting this teleost specific feature was acquired early in evolution and has been retained throughout the divergence of teleost Pax6 genes (Figure 1A).
We next downloaded the sequence scaffolds around the genes, and performed sequence alignments using PipMaker [62] using the human genomic region as reference sequence, to examine divergence of the paralogous genomic loci in the teleost species (Figure 1B). Examining the patterns of sequence conservation around the gene duplicates reveals a much more dramatic divergence between the paralogous loci in the four teleost fish, when compared to the relatively balanced sub-partitioning of conserved elements around the Pax6 duplicates in zebrafish. While in zebrafish both loci have retained a large and overlapping portion of the cis-regulatory repertoire, the divergence in the other teleosts has led to conservation of most of the ancient cis-regulatory landscape only at the Pax6a loci. The acanthopterygian Pax6b loci on the other hand have experienced a dramatic divergence or loss of cis-elements, such that, with one exception (see below), no recognizably conserved non-coding sequences remain in the locus in comparison with the mammalian or zebrafish Pax6 loci (Figure 1B). Furthermore, the blocks of sequence conservation in the 3′UTR are also absent from the acanthopterygian Pax6b, while found sub-partitioned between the zebrafish Pax6a and Pax6b loci. The lack of alternative exon 5a, the absence of CNEs and the differences in synteny conservation around the genes (see below) strongly suggest that the Pax6 duplicates of medaka, stickleback, fugu and Tetraodon originate from a separate duplication event compared to the zebrafish duplicates. To distinguish between these separate sets of Pax6 genes we will refer to the canonical Pax6 gene as Pax6.1 in species with multiple Pax6 loci. Thus all fish Pax6a genes will be termed Pax6.1, while the duplicate zebrafish Pax6a and Pax6b genes will be redefined as Pax6.1a and Pax6.1b. The second full-length Pax6 gene found in acanthopterygians will hereafter be referred to as Pax6.3.
The early expression pattern of medaka Pax6.3
As the near complete lack of CNEs between the Pax6.1 and Pax6.3 loci could suggest non-functionalization of the acanthopterygian Pax6.3 genes, we carefully inspected the coding sequences of the genes. All Pax6.3 genes contain open reading frames that encode clear Pax6 homologs. The conservation of ORFs in all four acanthopterygian species examined suggests that the gene is functional, which is further supported by the existence of ESTs for the Pax6.3 gene in medaka (GenBank BJ013007 and AM298948). To look for the expression pattern of Pax6.3, a probe was made from one of the medaka ESTs (clone MF01SSA182E11) and used for RNA in situ hybridizations in early medaka embryos (Figure 1C–1K). At stage19, weak expression of Pax6.3 was observed in the anterior diencephalon, whereas the posterior diencephalon and posterior part of the optic vesicle showed stronger expression (Figure 1D). Broader expression was seen for medaka Pax6.1 in the diencephalon, hindbrain and entire optic vesicle at this stage (Figure 1C, and [63]). At stage 22, the Pax6.3 expression domain in the diencephalon appears widened and persists in the posterior optic cup (Figure 1F), while Pax6.1 expression is maintained throughout the optic cup, diencephalon and hindbrain (Figure 1E). Expression of Pax6.3 persists in the diencephalon at stage 27 and 30, but eye expression was no longer observed (Figure 1H, 1K). In contrast Pax6.1 expression is maintained in the diencephalon, hindbrain and eyes at stage 27 (Figure 1G), and is restricted to the ganglion cell layer and inner nuclear layer of the neural retina at stage 30 (Figure 1I). This restricted expression pattern of medaka Pax6.3 mRNA in combination with the presence of an intact ORF suggests that the gene is functional, and is subject to regulated control of expression. We therefore generated a sequence alignment of the teleost Pax6 gene loci, using the stickleback Pax6.3 locus as baseline sequence (Figure S1). The alignment indicated the presence of a small number of CNEs that are conserved specifically between the Pax6.3 loci. As expected no non-coding conservation was found with the Pax6.1 loci, with exception of one small region of conserved sequence (E-200, see below).
Comparison of cis-regulatory activity in intron 7 of Pax6
To assess how the presence or absence of particular CNEs correlates with functional cis-regulatory activity we focused on intron 7 of the Pax6 gene. We have previously studied the murine intron 7 in transgenic mice and shown it to contain a number of tetrapod conserved enhancers [23]. These elements show a variable pattern of conservation in the fish loci (Figure 2A). As it contains a divergent complement of CNEs and, importantly, is flanked by the clearly recognizable landmarks of exons 7 and 8 to demarcate the region, we considered intron 7 particularly well-suited to study the correlation between sequence conservation patterns and functional enhancer activity. We cloned the full intron 7 sequences from the zebrafish Pax6.1a and Pax6.1b and medaka Pax6.1 and Pax6.3 genes and made reporter constructs for analysis in transgenic zebrafish. We also made reporter constructs for individual intron 7 CNEs from the zebrafish Pax6.1a locus to assess their contributions to the full intron 7 expression pattern. Analysis of the individual 7CE1, 2 and 3 elements indicated that expression in the diencephalon is contributed by the 7CE2 element (Figure 2D, 2E) and expression in the hindbrain by 7CE3 (Figure 2F, 2G). Reporter analysis of the 7CE1 element did not reveal any consistent expression pattern in transient transgenic fish. The 7CE2 and 7CE3 expression patterns in diencephalon and hindbrain are in agreement with the expression sites observed with the murine elements in reporter transgenic mice. However, we found no evidence for 7CE1 driven expression in the eye, or for consistent ectopic activity in the heart for 7CE2 as found in transgenic mice previously [23]. Next we examined reporter expression driven by the full intron 7 sequences from the Pax6 gene duplicates of zebrafish and medaka. We found that zebrafish Pax6.1a intron 7 (Dr6.1a-int7) drives strong reporter expression in the hindbrain and diencephalon of transgenic fish, but again no consistent expression was observed in the eyes (Figure 2H, 2J, 2K, 2L, 2N, 2O). Reporter expression driven by Pax6.1b full intron 7 (Dr6.1b-int7) is seen in the hindbrain only (Figure 2I, 2J, 2K, 2M, 2N, 2O). The fluorescence in the fin buds seen in Figure 2M, 2N, 2O is due to the site of integration and was not observed in other transgenic fish. The use of a dual fluorescence reporter system allows direct comparison of the expression patterns of two constructs in the same transgenic fish. Close examination of the reporter fluorescence patterns in the hindbrain of zebrafish transgenic for both the Pax6.1a full intron 7 and Pax6.1b full intron 7 constructs indicates that while the intron 7 sequences from the zebrafish Pax6.1a and Pax6.1b loci both drive expression in the hindbrain, the patterns are not fully overlapping. The zebrafish Pax6.1a region drives strong expression throughout the width of the hindbrain neural tube, while expression driven by the Pax6.1b region is more strongly concentrated along the midline. Furthermore, Pax6.1b intron 7 driven expression extends along the length of the neural tube from the hindbrain towards the caudal end, while Pax6.1a intron 7 driven expression is strong in the hindbrain segment but diminishes in level more caudally (Figure 2J, 2N, 2O).

Alignment of the full zebrafish and medaka intron 7 sequences indicates conservation of the 7CE2 and 7CE3 elements in the medaka Pax6.1 locus and a lack of conservation in the medaka Pax6.3 locus (Figure 2A). We analyzed reporter expression patterns for the medaka intron 7 constructs from both loci in transgenic zebrafish. Reporter fluorescence driven by medaka Pax6.1 intron 7 (Ol6.1-int7) was seen in hindbrain and diencephalon in a pattern that closely overlapped with zebrafish Pax6.1 intron 7 (Figure 2P, 2R). Next we assayed reporter expression driven by medaka Pax6.3 intron 7. Surprisingly, while having no conservation to the intron 7 sequences of the tetrapod or teleost Pax6.1 genes, Ol6.3-int7 consistently elicited reporter expression in the eyes of transgenic zebrafish. The retinal expression driven by Ol6.3-int7 correlates with the Pax6.3 RNA in situ signal in the eye of early medaka embryos, though it extends more anteriorly as well (Figure 2Q, 2R, 2S). In later stage embryos reporter fluorescence was also seen in the lens. As sequence alignment had shown the presence of a Pax6.3 loci specific conserved element in intron 7 (Figure 2B), we made transgenic zebrafish with the medaka Pax6.3 7CE element on its own, and again observed reporter expression in the eye (Figure 2S), suggesting this Pax6.3 locus-specific CNE drives Pax6.3 expression in the eye.
The elephant shark genome contains two Pax6 loci
The regulated expression of Pax6 in mammals depends on a wide genomic domain containing a large number of cis-regulatory elements [22]–[28]. Several of these are located at large distances from the promoters and coding region of the gene [24], [25], [26]. The distant downstream region in particular has been shown to be essential for Pax6 expression as its heterozygous removal through deletion or translocation leads to the eye malformation aniridia in human patients [30], [31], [25], [64]. Alignments of the wider Pax6 locus between mammalian and fish Pax6.1 loci show conservation of most proximal CNEs, but absence of many of the distal ones in fish. We considered whether the absence of clear sequence conservation in teleost fish at the relative position of a conserved element in tetrapods would indicate the loss of that element in fish, or the gain of that element in the tetrapod lineage. To help distinguish between these possibilities we set out to obtain an outgroup for comparative studies. The elephant shark is a cartilaginous fish with a relatively small genome, situated at the base of the jawed vertebrate lineage, having split off from the bony vertebrate lineage before the split between the ray-finned fish and tetrapods [65]. We searched the 1.4× coverage sequence of the elephant shark genome [66] for the Pax6 gene by BLAST. To our surprise the search identified two scaffolds each containing a different Pax6 fragment. PCR primers were designed for these fragments and used to identify positive BAC clones (23H6 and 37E6) from a genomic BAC library. After complete sequencing the BAC clones were found to encode two different Pax6 genes belonging to separate gene loci (Figure 3A). The first elephant shark Pax6 (hereafter referred to as Pax6.1) gene is highly homologous to the mammalian Pax6 gene, both in terms of gene structure and protein conservation (Figure 3B, 3C). The second elephant shark Pax6 gene lacks the N-terminal exons of the canonical Pax6 gene and encodes a Pax6 protein lacking the paired-box DNA-binding domain (Figure 3B, Figure S2). We have termed this novel paired-less Pax6 gene as Pax6.2. We next used the genomic sequence to design probes to rescreen our BAC library and walk outward from the Pax6-containing BACs. In total we obtained 477 kb of genomic sequence for the Pax6.1 locus and 156 kb of sequence for the Pax6.2 locus. A number of flanking genes were present in the BAC contigs (Figure 3A) and allowed analysis of synteny around the genes. Synteny around the elephant shark Pax6.1 gene is fully conserved to the human PAX6 locus, with all genes present in our BAC contig found in equivalent positions to their order in the mammalian locus (Figure 3A, Figure 7B), clearly indicating a common ancestry of elephant shark Pax6.1 and mammalian Pax6. The elephant shark Pax6.2 gene is adjacent to a reticulocalbin gene (Rcn3), but none of the other genes in the Pax6.2 locus is syntenic with the mammalian Pax6 locus (Figure 3A, Figure 7D). Next we looked for tissue-specific expression of elephant shark Pax6.1 and Pax6.2. rtPCR analysis using RNA from 13 different adult elephant shark tissues revealed expression of Pax6.1 in brain, eye and pancreas, with a weaker signal in intestine, in accordance with the expression of Pax6.1 in other organisms. Elephant shark Pax6.2 has a more limited expression in the eye and kidney (Figure 3D). In mammals a paired-less Pax6 protein isoform, equivalent to the elephant shark Pax6.2-encoded protein, is produced from an internal promoter (Pα)in intron 4 of the single Pax6 gene [38]. To determine whether the existence of the Pax6.2 gene in elephant shark would be to act as substitute for the internal transcript of the mammalian gene (or vice versa) we investigated whether a similar α-transcript is also produced from the elephant shark Pax6.1 gene. We found an elephant shark Pax6.1α transcript in the brain and eye, but not in pancreas (Figure 3E). While the expression of Pax6.2 in the eye is unsurprising for a Pax6 homolog, it is intriguing to see expression in the kidney, suggesting a novel function has been acquired by elephant shark Pax6.2 in this tissue.


An ancient conserved element identifies a novel long-range enhancer
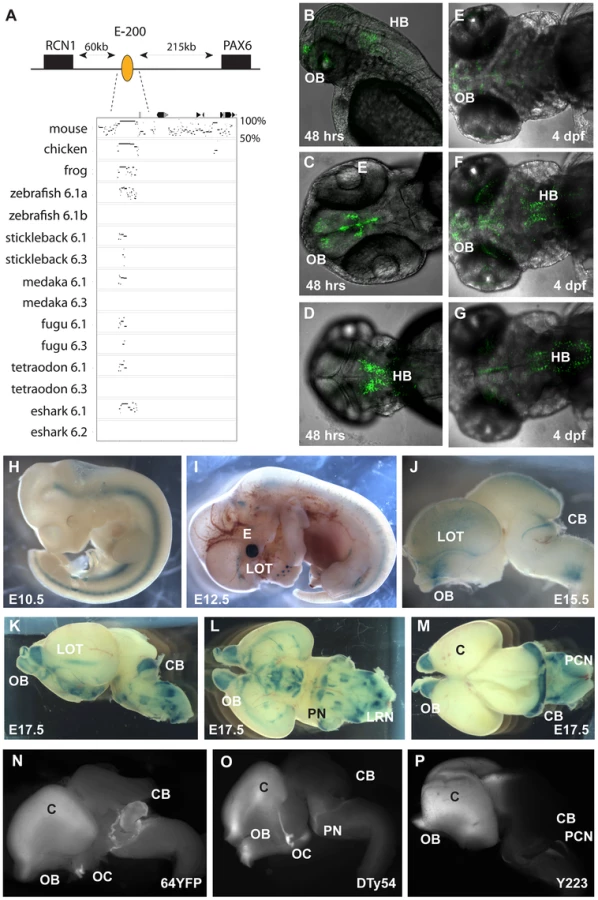
Multispecies sequence alignments with the elephant shark Pax6.1 genomic sequence reveal a large number of highly conserved non-coding elements in the locus. In accordance with a general observation [57] the number and conservation level of non-coding elements around Pax6 is significantly higher when comparing the elephant shark Pax6.1 and mammalian Pax6 locus, than when either is compared to the teleost Pax6.1 loci. In particular, clear conservation with elephant shark is observed for several mammalian long-range CNEs that appear absent in teleosts, both in the distal upstream and downstream regions, indicating these are ancient non-coding elements that have been secondarily lost in teleosts (Figure 3F).
We focussed on the region upstream of Pax6. As all extragenic aniridia patient breakpoints identified so far disrupt the Pax6 downstream region [32], [67], regulatory activity in the upstream region has remained poorly investigated. Comparative sequence analysis between mammals and the elephant shark Pax6.1 locus reveals the presence of a number of long-range CNEs in this region. Many of these CNEs are not found in teleosts, and are therefore candidate Pax6 enhancers that have been lost in fish. Analysis of these CNEs will be described elsewhere. Here we characterize one element of particularly prominent sequence conservation, located approximately 200 kb upstream of human Pax6, which has been named E-200. The E-200 element displays strong sequence conservation between elephant shark and human, but is also detected in the Pax6.1 loci of several teleosts. Moreover, in fugu and stickleback a low level of sequence conservation is also seen in the Pax6.3 loci (Figure 4A, Figure S3). The location of the element upstream of Pax6, between Pax6 and Rcn1, suggests Pax6 as its most likely target gene, but synteny conservation around the Pax6.1 locus leaves the possibility that other genes in the region, notably Wt1 and Rcn1, might be the actual targets. The conservation to several fish duplicate loci provides an opportunity to check linkage between the element and these genes. We found that in all gene loci containing the E-200 element, it was linked to Pax6, while linkage to Wt1 or Rcn1 was only found where Pax6 and Wt1 or Rcn1 are both present (not shown). To determine the function of this element we first made transgenic zebrafish, using the human element. We observed expression of the fluorescent reporter in the olfactory bulbs, olfactory tracts and the hindbrain regions of transgenic embryos (Figure 4B–4G). Next we generated transgenic mice with the human E-200 element. At early developmental stages (E9.5–E11.5) no consistent staining pattern was seen. From E12.5 expression appeared in the olfactory tract regions and the upper rhombic lip of transgenic mice (4 expressing/10 total transgenic). The staining in the eye in Figure 4I was not consistent between all expressing lines and is most likely due to site of integration. By E15.5 expression was found around the olfactory bulbs, in the lateral olfactory tracts (LOTs) and started to appear in the cerebellum (Figure 4J). At E17.5 strong staining was seen in the cerebellum, as well as around the olfactory bulbs and LOTs. Staining was also observed in the precerebellar neuroepithelium (PCN), migratory streams and precerebellar nuclei (Figure 4K–4M). The E-200 element is a long-range enhancer located around 200 kb upstream of the PAX6 P0 promoter. It is not present on the human YAC that was used in previous reporter transgenic studies on the long range regulation of PAX6 [24]. Reporter fluorescence in the cerebellum of all transgenic lines carrying the YAC (Figure 4O, 4P) is weak or absent compared to the expected level as demonstrated by the fluorescence from a targeted insertion of YFP into the endogenous Pax6 gene (Figure 4N; Kleinjan et al. in preparation). This suggests the presence of the long-range E-200 element is required to achieve appropriate expression of Pax6 in the cerebellum. In contrast, fluorescence in other sites of E-200 activity is not noticeably lower in the YAC transgenics, suggesting sufficient redundancy of cis-regulatory activity in these tissues (Figure 4N–4P). The importance of the E-200 enhancer for cerebellar expression is in accordance with the conservation of this ancient enhancer linked to Pax6 since early vertebrate evolution.
The second Pax6 locus in the elephant shark genome
The elephant shark Pax6.1 locus with its high level of sequence homology and synteny conservation to the mammalian Pax6 loci has provided insight into the ancient history of the Pax6 cis-regulatory domain. In addition to the Pax6.1 locus, our elephant shark BAC library screening yielded a BAC contig containing a second Pax6 locus. The BACs were fully sequenced to reveal a novel Pax6 homolog lacking the N-terminal exons that encode the paired box in canonical Pax6 isoforms. This second elephant shark Pax6 gene was named Pax6.2. We aligned genomic sequence from the Pax6.2 locus with other Pax6 loci to identify putative CNEs in the locus. This revealed one clear CNE with conservation to the Pax6.1 locus (Figure 5A). This element is located upstream of the Pax6.2 gene but has homology to the intronic neural retina enhancer (NRE) located in intron 4 of canonical Pax6.1 [27], [68]. Interestingly, in addition to retinal-specific enhancer activity the NRE element also serves as an internal promoter (Pα) in the mouse and zebrafish Pax6.1 genes, and transcripts initiated from this promoter give rise to paired-less Pax6 isoforms in these species [38]; [37]. To test whether the elephant shark Pax6.2 CNE (eshark6.2NRE) has enhancer activity we generated a number of mouse transgenic reporter lines with the element driving LacZ expression (4 expressing/8 total transgenics). Staining was found in the developing retina from E9.5 onwards in a pattern that closely resembles the expression of the murine NRE element [27]. No staining is seen at E9.5 (Figure 5B). Expression starts at E10.5 in two lateral domains on either side of the optic cup (Figure 5C), spreading wider from E11.5 (Figure 5D) to the full developing retina from E12.5 (Figure 5E), and continuing in the retina at E14.5 and E17.5 (Figure 5F, 5G). To assess the functional equivalence of the conserved NRE elements from the elephant shark Pax6.1 and Pax6.2 loci, we produced transgenic zebrafish with the CNEs driving GFP and mCherry fluorescent reporter genes. Both CNEs drove a clean and overlapping expression pattern in the retina (Figure 5H–5J). This confirms the elephant shark Pax6.1 and Pax6.2 NREs as neuroretinal enhancers that derive from an ancestral element that was present before the duplication event that created the elephant shark Pax6.1 and Pax6.2 loci.

Phylogenetic and synteny relationships between vertebrate Pax6 loci
The existence of a second, paired-less Pax6 gene in elephant shark suggested two possibilities: a cartilaginous fish-specific duplication event after the split from the bony vertebrate lineage; or an ancient duplication event that occurred in a gnathostome ancestor. We reasoned that if the duplication had occurred in a gnathostome ancestor, we might be able to find orthologs of this paired-less Pax6 gene in some species of bony vertebrates. We therefore searched the genomes of bony vertebrates for additional Pax6 loci with similarities to elephant shark Pax6.2 in gene structure, locus synteny and cis-element content. Remarkably we found novel paired-less Pax6 genes in the genomes of frog (Xenopus tropicalis), Anolis lizard and teleost fishes (zebrafish, stickleback, medaka, and fugu). Evidence for the existence of Pax6.2 orthologs was also found in cod, sea bass and Nile tilapia (data not shown). Despite extensive searches, no Pax6.2 gene was found in birds or mammals. Thus in many fish species three Pax6 genes exist, the canonical Pax6.1 as well as the newly identified Pax6.2 and re-defined Pax6.3 genes. The zebrafish genome also harbors three Pax6 genes, but in this case the trio consists of the fish specific duplicates of canonical Pax6.1 (Pax6.1a and Pax6.1b [58], [59]) and the novel paired-less Pax6.2 gene. To visualize the relation between the members of the newly defined Pax6 gene family we generated a phylogenetic tree using the Neighbor Joining method [69]. This phylogenetic tree supports the classification of gnathostome Pax6 genes into three clades, with zebrafish Pax6a and Pax6b both in the Pax6.1 clade. The paired-less Pax6.2 genes form their own clade and the acanthopterygian Pax6.3 genes make up the most distant clade (Figure 6A). Comparison of synteny relationships around the Pax6 family members of different species provides further insight into the divergence of the gene loci. The well known synteny block around the mammalian Pax6 gene is fully conserved in the elephant shark Pax6.1 contig. Synteny is also largely conserved in the Pax6.1 loci of acanthopterygian fishes, but a subpartitioning of genes is observed at the zebrafish Pax6.1a and Pax6.1b loci (Figure 6B). In contrast, synteny conservation around the Pax6.3 gene in acanthopterygians is limited and only encompasses the adjacent Dnajc24 gene in medaka, fugu and stickleback (Figure 6C). Closer examination of the Dnajc24 genes reveals that conservation between orthologs from the same locus is higher than across loci. In particular, zebrafish Dnajc24 is more related to mammalian Dnajc24 than to medaka or stickleback Dnajc24. Synteny around the elephant shark Pax6.2 locus and the Xenopus, lizard and zebrafish loci provides evidence that they are of common descent. In these species the Pax6.2 gene is found in close synteny with Rcn3 (reticulocalbin 3), Nosip (nitric oxide synthase interacting protein), Prrg2 (proline rich G-carboxyglutamic acid 2) and Cpt1 (Figure 6D). No Pax6.2 ortholog was found in the Nosip/Rcn3 locus in birds or mammals (Figure 6D). In acanthopterygian fish species the Pax6.2 gene is also absent from the Nosip/Rcn3 locus, but in these species a Pax6.2 ortholog is present adjacent to the Mapk7, Prss22 and Mmp25 genes. The locus also harbors Trpm4, Irf3 and Prr12 paralogs suggesting a link with the Nosip/Rcn3 locus. This locus is well conserved in zebrafish, except for a lack of Pax6.2. These observations strongly suggest that after the FSGD duplicate copies of the Pax6.2 gene (Pax6.2a and Pax6.2b) must have persisted for some time in the basal teleost genome until after the split between the zebrafish and acanthopterygians lineages. Subsequently, one copy of the gene was lost reciprocally from the two loci in zebrafish and the acanthopterygians. In zebrafish, Pax6.2a was retained alongside Rcn3 and Nosip, while in acanthopterygians the reciprocal duplicate (Pax6.2b) was retained in the more derived locus between the Prss22 and Mmp25 genes (Figure 6D, Figure S4).

Characterisation of the novel zebrafish Pax6.2 gene
Having identified a Pax6.2 homolog in zebrafish we performed rtPCR for this novel Pax6 gene to check for expression. cDNA was generated from zebrafish embryos covering the first five days post fertilization (1–5 dpf). rtPCR results show that Pax6.2 is expressed in zebrafish embryos at all stages examined. rtPCR for Islet-1, known to be expressed at these stages was used as control (Figure 7A). Alignment of Pax6.2 proteins from various species shows clear homology between the genes, but indicates a higher similarity between zebrafish Pax6.2 and the acanthopterygian Pax6.2 genes (Figure 7B). To assess the tissue-specific expression pattern of zebrafish Pax6.2 we performed RNA in situ hybridization analysis on fixed zebrafish embryos from 1 dpf to 5 dpf. The staining pattern reveals Pax6.2 to have a highly restricted expression. At 24 hpf staining is seen more widely in the head region of embryos (Figure 7C). The expression becomes limited to the developing retinae only from the next stages examined (48 hpf to 5 dpf) (Figure 7D, 7G–7I). We used Optical Projection Tomography (OPT, [70]) to visualize the in situ expression pattern at 2 dpf, which confirmed the restricted expression of Pax6.2 in the retina only (Figure 7E, 7F, Video S1), where it appears limited to the inner nuclear layer, potentially marking the amacrine cells. Finally we performed morpholino knock-down experiments to investigate the potential function of Pax6.2. Injections of a Pax6.2 morpholino into zebrafish oocytes resulted in zebrafish embryos with relatively smaller eyes compared to embryos injected with a control morpholino (Figure 7J). To quantify this observation we titrated the morpholino concentration and repeated the injections with the optimal dose of Pax6.2 and control morpholinos. Embryos were fixed at 2 dpf and tested for the presence of Pax6.2 transcript by in situ hybridization. Pax6.2 ISH signal was unaffected in control morpholino injected embryos. The majority of Pax6.2 morpholino injected embryos had completely lost Pax6.2 according to ISH signal, while the remainder showed partial ISH signal loss. We measured the eye diameter of Pax6.2 morphants relative to total body length of the embryos. A clear deficiency in eye size was seen in the Pax6.2 morphants in comparison with control morpholino injected embryos, indicating an essential role for the paired-less Pax6.2 protein in eye development (Figure 7K, Figure S5).
Finally, we performed sequence alignments using PipMaker to identify CNEs in the Pax6.2 gene loci. A distinct region of sequence homology outside the exons of the gene was found in the upstream regions of zebrafish, stickleback, medaka and fugu (Figure 8A). Conservation of the CNE between zebrafish and the other teleosts is seen despite their origin from reciprocal Pax6.2 duplicate loci. A smaller sub-region of the CNE also showed conservation to the elephant shark Pax6.2 upstream region. Intriguingly, this short conserved fragment maps to the edge of the elephant shark Pax6.2 NRE element, but has no distinguishable homology to the NRE sequence in intron 4 of Pax6.1 genes. To examine the putative functional activity of the Pax6.2 CNE region, a longer fragment covering the homology region of the teleost CNEs, and a shorter fragment centered around the zebrafish to elephant shark homology region, were cloned from the zebrafish Pax6.2 locus and inserted into fluorescence reporter constructs for the production of transgenic fish. Fluorescence was seen in the retinae of transgenic fish at 72 hpf with both the long (GFP) and short (mCherry) fragments (Figure 8B). In addition the long fragment showed some minor expression in the forebrain region in a subset of transgenics. Fluorescence signal was restricted to the inner nuclear layer (INL), in accordance with the RNA in situ pattern of Pax6.2 (Figure 7G). This suggests that sequence divergence at the NRE in separate branches of the Pax6 gene family has led to subtle differences in the spatial detail of its retinal enhancer activity. It confirms the ancient role of the NRE element as a retinal enhancer in the ancestral Pax6 locus. In summary, we have discovered a novel, paired-less Pax6 gene in the genomes of multiple species. In zebrafish Pax6.2a is expressed in the inner nuclear layer of the retina and we have identified a conserved enhancer driving this expression pattern.

Discussion
The genome is a remarkable repository of biological information. Within its sequence it contains not only a complete set of instructions for embryonic development of the organism, maintenance of adult homeostasis and the response to environmental interactions, but also a record of the evolutionary history of its genes and associated sequences. By comparison of genomic sequences from multiple contemporary species of divergent lineages attempts can be made to reconstruct the ontogeny/phylogeny of specific genes and their regulatory landscapes. It is well established that early vertebrate evolution was accompanied by two rounds of whole genome duplications (2R) [44], while a third round (3R) has occurred later specifically in the teleost lineage of bony fish around 320 million years ago [40]. Duplication events are recognized as powerful drivers of evolutionary change as they provide enhanced opportunity for the subsequent modification or mutation of gene duplicates, and/or the alteration of their cis-regulatory landscapes, while the other copy maintains critical functions under selective pressure [45], [48].
We have previously shown that divergence of the cis-regulatory landscapes around the duplicate genes of the developmental regulator Pax6 in the zebrafish has led to their subfunctionalization [59]. In this study we demonstrate that duplicate Pax6 loci also exist in other vertebrate species. Unexpectedly, we have uncovered that these duplicates form a diverse family of Pax6 genes that are derived from multiple independent duplication events. These duplications were followed by multiple independent gene losses in separate vertebrate lineages, such that a variable subset of family members has been retained in contemporary vertebrate species. We show that while the genomes of mammals and birds contain only a single Pax6 gene, other species have two (elephant shark, Xenopus tropicalis, Anolis lizard) or even three Pax6 genes (zebrafish, stickleback, medaka, other teleost species). The various Pax6 family members are characterized by differences in their protein structure and in the composition of their cis-regulatory landscapes.
In mammals a multitude of key functions in development and maintenance of the eye, brain and pancreas are carried out by a single Pax6 gene [10]. A number of variant protein isoforms encoded by this single mammalian gene are thought to implement different, complementary subsets of Pax6 activity [71]–[73]. The canonical Pax6 protein contains two DNA binding domains: an N-terminal paired domain followed by a paired-type homeodomain. A proline-serine-threonine rich transactivation domain is located at the C-terminus [13], [19]. An alternative isoform, Pax6(+5a) is made by inclusion of an alternatively spliced exon 5a resulting in a 14 amino acid insertion in the paired domain leading to recognition of a different binding sequence [35]. A third isoform, Pax6ΔPD, produced from a transcript initiated at an internal promoter (Pα) located in intron 4 of the gene lacks the entire paired domain [27], [38]. Reporter transgenic studies of a BAC engineered to express dsRed from the Pα promoter have shown that Pax6ΔPD is expressed in a highly restricted expression pattern [36], [37]. In mouse Pax6ΔPD is found in the retina and olfactory bulbs, while in the zebrafish it is only seen in the amacrine cells in the retina. The role of this isoform is currently unclear.
Inspection of the Pax6 genes of several ray-finned fish species indicated that the ability to encode the alternative exon 5a is present only in the Pax6.1 genes of the fish species examined. Sequence comparison between the 5a exons from multiple species shows that it is less conserved than the rest of the paired-box, both in amino acid composition and length (Figure 1A), suggesting that the main function of this peptide might be disruption of the paired box, putting less stringency on the actual sequence itself. In contrast, the other Pax6 copies of the teleost species examined completely lack the alternative exon 5a. The exception is zebrafish where both Pax6 duplicates have a well conserved exon 5a. Moreover, the conspicuous sub-partitioning of CNEs between the duplicate zebrafish Pax6a and Pax6b loci is not seen in the multiple gene loci of medaka, stickleback, Tetraodon and pufferfish, where instead the loci are devoid of any recognizable sequence conservation between them outside of the exons of the Pax6b loci, apart from a short conserved fragment at the E-200 cis-element. Thirdly, in zebrafish the ubiquitous Elp4 gene has been retained next to one of the Pax6 copies (Pax6.1b) while the long-range enhancers of the DRR have mostly been conserved in the other copy (Pax6.1a) [59]. In the acanthopterygians both the Elp4 gene and the long-range enhancers are found adjacent to the Pax6.1 gene, while the other Pax6 duplicate is located in a different synteny region. Taken together these observations strongly suggest that the Pax6 duplicates of zebrafish and those of the other teleosts derive from different duplication events. The near total lack of CNE conservation suggests that the acanthopterygian Pax6 duplicates have a more ancient evolutionary origin [57], [74]. Based on these observations we propose to refer to the zebrafish duplicates as Pax6.1a/b and refer to the acanthopterygian duplicates as Pax6.1 and Pax6.3.
To gain more insight into the evolutionary origin of the fish Pax6 loci we screened a BAC library from the elephant shark as an outgroup for comparative studies. Contrary to expectation we identified two separate Pax6 loci in this cartilaginous fish, which we designated as Pax6.1 and Pax6.2. The elephant shark Pax6.1 locus is highly similar to the tetrapod Pax6 locus and the Pax6.1 loci of ray-finned fish. In contrast, the second elephant shark Pax6 locus encodes a Pax6 gene lacking the N-terminal exons of the canonical Pax6 and is predicted to produce a Pax6 homolog without the paired domain. It is thus similar to the Pax6ΔPD isoform derived from the internal Pα promoter of mammalian Pax6. The presence of a separate paired-less Pax6 gene in the elephant shark genome suggested that it might be fulfilling the equivalent role of the Pα-derived mammalian paired-less Pax6 isoform. However, we show that a paired-less isoform is also produced from the elephant shark Pax6.1 gene. Nevertheless, persistence of the gene suggests it does serve a unique function and it is possible that its specific expression not only in the eye but also in the kidney accounts for its retention in the elephant shark genome.
The identification of two Pax6 gene loci in the elephant shark raised two possibilities: Either the duplication of the Pax6 locus occurred uniquely in the cartilaginous fish lineage after the split from the bony vertebrate lineage, or the duplicate loci had arisen before the split between cartilaginous and bony fish lineages. We resolved this question by looking for potential Pax6.2 homologs in non-cartilaginous species, using the genes in synteny with the elephant shark Pax6.2 gene in our searches. Sequence analysis of our elephant shark Pax6.2 BAC contig revealed the presence of an adjacent reticulocalbin gene, Rcn3, as well as the genes Nosip, Prrg2, Cpt1a. A search for loci containing these genes in other vertebrate genomes led us to the identification of novel Pax6.2 orthologs in several species, including frog (Xenopus tropicalis), lizard (Anolis lizard) and many fish species including medaka, stickleback and zebrafish. This clearly indicates that the duplication that gave rise to the Pax6.2 gene must have occurred before the split between cartilaginous and bony fish. Although the presence of a Pax6.2 in teleosts means that the gene was present during the FSGD, we could only find a single Pax6.2 gene in the fish genomes examined, indicating the second Pax6.2 duplicate has been lost. Under the DDC model [46], [48] non-functionalization of one copy of gene duplicates is often seen for genes with a single function, and accordingly we show by in situ hybridization that Pax6.2 has a highly specific expression restricted only to the neuroretina of developing fish embryos. Nevertheless we do find evidence for the original presence of duplicated Pax6.2 genes in early teleost fish by comparisons of synteny around the zebrafish Pax6.2 locus with synteny around the Pax6.2 loci of medaka, stickleback and fugu. While in zebrafish Pax6.2 is found in a synteny block with the Nosip and Rcn3 genes in common with the elephant shark, frog and lizard, the gene is located in a different synteny block in medaka, stickleback and fugu. This indicates that reciprocal duplicate copies of the gene were lost in zebrafish versus the acanthopterygian species. Despite being reciprocal duplicates (which we refer to as Pax6.2a for zebrafish and Pax6.2b for the acanthopterygians), both gene loci share a well conserved CNE in their upstream region that also shows some homology to the elephant shark Pax6.2 upstream region. In reporter transgenic zebrafish both the full-length fish CNE and a shorter fragment around the elephant shark-zebrafish conserved region drive highly specific expression in the inner nuclear layer of the retina in accordance with the RNA in situ pattern for Pax6.2. The retention of a Pax6.2 gene in multiple species suggests functional importance and our morpholino knock-down experiment in zebrafish embryos demonstrates a role for Pax6.2 (Pax6c) in eye development. Nevertheless the Pax6.2 gene has eventually been lost in the avian and mammalian lineages. It is currently unclear whether this suggests a change in the molecular networks for eye development between species, or redundancy in the availability of a paired-less form of the Pax6 transcription factor. Despite the absence of Pax6.2 a paired-less Pax6 isoform is produced in birds and mammals, generated from a transcript initiating at an internal promoter Pα in intron 4 of the canonical Pax6.1 gene [37], [38]. Expression of this paired-less Pax6.1 isoform (pax6ΔPD) has been shown to be specific to the eye in zebrafish and mouse [38]. However, these alternative ways of producing a paired-less isoform are not mutually exclusive as the Pax6.1 paired-less isoform is also produced in zebrafish [37] and elephant shark (this study) which have nonetheless retained the Pax6.2 gene.
The genomic region around mammalian Pax6 contains a large number of cis-regulatory elements [22]–[28]. Functional constraint on these elements imposes a strong demand on the conservation of their sequence, and consequently many cis-regulatory elements can be identified as CNEs. However, significant divergence of regulatory sequence following duplication can occur with or without concomitant changes in expression pattern. In extremis this can result in conservation of functional activity despite the disappearance of recognizable sequence conservation [75]. Our analyses of reporter expression driven by individual CNEs versus the full zebrafish intron 7 sequences show that the regulatory activity residing in the intron is contributed by the 7CE2 and 7CE3 elements. Loss of the conserved sequences of the 7CE2 element from zebrafish Pax6.1b intron 7 correlates with absence of expression in the diencephalon, suggesting that regulatory activity is not maintained in the absence of sequence conservation. Curiously no specific expression was observed at the stages examined for the 7CE1 element despite its conservation in both loci, suggesting the 7CE1 CNE must fulfill some other, unknown function. In contrast, analysis of reporter expression driven by intron 7 of medaka Pax6.3 in comparison with the patterns directed by the introns 7 of both zebrafish Pax6.1a and Pax6.1b loci and medaka Pax6.1, suggest that this region does show conservation of functional activity despite lack of sequence conservation. However, our new observations on the more ancient ontogeny of the Pax6.1 and Pax6.3 loci suggest that most enhancer elements have formed independently in the Pax6.1 and Pax6.3 loci, and that the presence of enhancers located in similar positions (e.g. intron 7) in the acanthopterygian Pax6.1 and Pax6.3 loci is most likely coincidental. The independent acquisition of cis-regulatory elements in the Pax6.1 and Pax6.3 loci, which would be predicted if the duplication of the loci occurred before the large-scale appearance of cis-regulatory elements during the early stages of gnathostome evolution [53]??[74], is supported by sequence alignments using stickleback Pax6.3 as baseline, which reveal a number of Pax6.3 loci-specific CNEs in addition to the 7CE(6.3) element as further candidate Pax6.3 enhancers that are not found in the Pax6.1 loci (Figure S1).
Sequence alignments between the three Pax6 sub-family members support the independent acquisition of most cis-regulatory elements after the WGD events. The large number of CNEs conserved between mammalian and elephant shark Pax6.1 loci indicates that the majority of Pax6.1 enhancers must have appeared in the timeframe between the WGD and divergence of cartilaginous and bony fish, with few additional cis-elements having appeared since. However, we also show that a limited number of CNEs are present in multiple branches of the Pax6 gene family. Comparisons of the elephant shark Pax6.2 locus with multiple Pax6.1 loci revealed a single conspicuous CNE. Intriguingly, the homologous element maps to the neuro-retina enhancer (NRE) in Pax6.1 intron 4 that coincides with the internal Pα promoter [27]. We show that the sequence conservation of this CNE extends to function as reporter expression driven by the elephant shark 6.2 NRE element in transgenic mice and zebrafish is very similar to the retinal expression driven by the mouse NRE element [27]. Using transgenic zebrafish we show that both elephant shark 6.1NRE and 6.2NRE elements drive an identical expression pattern in the neuroretina. This conservation of sequence and function of the NRE indicates the element was already present in the ancestral gene locus before the WGD events that led to separate Pax6.1 and Pax6.2 genes.
Another ancient enhancer that must have been present before the 1R/2R WGD events is located in the region upstream of Pax6, between the Pax6 and Rcn1 genes. This highly conserved element, E-200, is present in all Pax6.1 loci including the tetrapod, elephant shark Pax6.1 and teleost Pax6.1 loci. A small central fragment of this element is also conserved in two of the teleost Pax6.3 loci, medaka and Tetraodon (Figure S1), and was subsequently also found in both Pax6.1 and Pax6.3 loci of the Nile tilapia. It is the only non-coding sequence conserved between the acanthopterygian Pax6.3 loci and the fish/mammalian Pax6.1 loci. Both mouse and zebrafish reporter transgenesis revealed expression from the E-200 element in the hindbrain/cerebellar region, olfactory bulbs and lateral olfactory tracts. Reporter expression in the cerebellum was of particular interest as we had previously noted that a large YAC reporter transgene was deficient in driving expression in the cerebellum of transgenic mice. As the PAX6 genomic region contained in the YAC does not extend to the E-200 element we propose that the lack of cerebellar expression from the YAC is due to the absence of this element, though further long-range elements could also be involved. The presence of the E-200 CNE near Pax6.3 clearly links the element to Pax6 since the Pax6.3 loci do not contain Rcn1 or Wt1.
In summary, we demonstrate that the locus for the developmental regulator Pax6 has undergone multiple duplication events, followed by variable divergence or loss of the gene duplicates. Divergences between the loci have affected both the coding region as well as the cis-regulatory landscapes in lineage specific ways, resulting in a family of variant Pax6 genes in vertebrate genomes. The existence of multiple variant Pax6 genes and proteins also occurs in other parts of the animal kingdom. In C. elegans two different classes of mutation, vab-3 and mab-18, are known to affect different isoforms from the same Pax6 gene. One of these, the mab-18 isoform, encodes a paired-less isoform that causes a male fertility phenotype due to a sensory defect in the male sensory organ [76]. The Drosophila genome contains four Pax6 orthologs, occurring in two sets of adjacent gene pairs. The eyeless and twin-of-eyeless pair encode the canonical Pax6 homologs, whose absence causes the eyeless phenotype [7], [77]. A second pair of Pax6-like genes, the eyegone and twin-of-eyegone gene pair, have an incomplete paired box and are considered to be the functional equivalent in Drosophila of the mammalian Pax6(+5a) isoform [78].
Based on the phylogenetic relationships of the various Pax6 genes in vertebrates and their conservation of synteny and cis-regulatory domains, we propose the following model to explain the evolution of the family of Pax6 genes in jawed vertebrates (Figure 9, Figure S6). The 1R and 2R that occurred in the stem vertebrate lineage gave rise to four copies of Pax6 (Pax6.1 to Pax6.4), one of which (Pax6.4) was lost before the diversification of gnathostomes. Of the remaining three genes one evolved into the canonical Pax6 (Pax6.1), as represented by the well-studied contemporary mammalian Pax6 locus. The second gene lost its 5′ exons encoding the paired domain and evolved to encode a paired-less form of Pax6 (Pax6.2) expressed in a highly restricted pattern in the retina. This gene was subsequently lost independently in the mammalian and chicken lineages. The third copy (Pax6.3) was lost independently in the elephant shark lineage and in the common ancestor of tetrapods but retained in the ray-finned fish lineage. The origin of the Pax6 family at the 1R/2R is consistent with the paucity of CNEs between the Pax6 member loci as the WGDs occurred before the rapid and large-scale appearance of cis-regulatory sequences at the base of the jawed vertebrate lineage [53], [74].

The stem ray-finned fish lineage that diverged from the common ancestor of the tetrapods contained three Pax6 loci (Pax6.1, Pax6.2 and Pax6.3). The 3R that occurred in this lineage before the teleost radiation resulted in duplicate copies of each of these gene loci (Pax6.1a/Pax6.1b; Pax6.2a/Pax6.2b; and Pax6.3a/Pax6.3b). Among teleosts, zebrafish retained both copies of canonical Pax6.1 due to cis-regulatory subfunctionalization, while the acanthopterygian ancestor lost one of its Pax6.1 duplicates, resulting in a single copy of the canonical Pax6 (Pax6.1a) locus in medaka, stickleback and fugu. Of the duplicate copies of the paired-less form of Pax6, one copy (Pax6.2a) was lost in the acanthopterygian ancestor, while the reciprocal copy (Pax6.2b) was lost in the zebrafish lineage. This has resulted in zebrafish retaining the Pax6.2a copy while medaka, stickleback and fugu retaining Pax6.2b. In the case of the third Pax6 gene (Pax6.3), one of the two duplicate copies was lost before the divergence of the zebrafish and acanthopterygian lineages, whereas the second copy was lost in the zebrafish lineage after the split from the common ancestor of the acanthopterygians. Thus, no copies of the Pax6.3 locus remain in zebrafish, whereas medaka, stickleback and fugu contain only one copy of this gene.
At present this model only presents a rough schematic of the progression of the Pax6 gene family through vertebrate evolution, but future availability of whole genome sequences for additional vertebrate species will allow a more detailed model to emerge. The lamprey and hagfish genome sequences in particular could provide useful information on the Pax6 duplications at the 1R and 2R events. In light of the presumed presence of the NRE and E-200 elements in the ancestral, pre-WGD Pax6 locus we made alignments with the amphioxus (Branchiostoma floridae) and Ciona Pax6 loci, but no sequence conservation apart from the coding exons was found.
Our model highlights the fact that the many pleiotropic functions of the key developmental regulator Pax6 are shared between different members of a family of genes and isoforms in various species, and may be helpful in further dissection of species-specific differences in development and morphological variation. It therefore remains of great interest to continue the analysis of the cis-regulatory landscapes around Pax6 genes from multiple species to trace the time-frame of acquisition of their cis-regulatory hardwiring and link the presence and divergence of enhancer elements with differences in expression patterns and in anatomical and developmental features.
Materials and Methods
Ethics statement
All mouse experiments were approved by the University of Edinburgh ethical committee (TR-11-08) and performed under UK Home Office license number PPL 60/3785.
Sequence analysis
Genomic sequences were collected from Ensembl release 65, December 2011 [79] for these species: Human GRCh37 assembly (February 2009), Zebrafish Zv9 (Apr 2010), Medaka HdrR (Oct 2005), Tetraodon TETRAODON 8.0 (Mar 2007), Stickleback BROAD S1 (Feb 2006), Fugu FUGU 4.0 (Jun 2005). Sequences were manipulated using programs from the EMBOSS package [80]. Where necessary, missing regions of interest were incorporated following Blast searches [81] using human or fish Pax6 sequences. Ensembl gene annotation and protein predictions were used, in some cases supplemented with manual annotation using Genewise [82] for gene prediction from genomic sequence, with human or fish Pax6 protein as template. Other sources of information (proteins, ESTs) were used to further refine gene predictions and search for small exons, including exon 5a and 5′ non-coding exons. Protein sequences were aligned with the CLUSTALW program [83] and edited and displayed using the Genedoc software www.psc.edu/biomed/genedoc [84].
Genomic sequences were aligned, annotated and displayed using the PipMaker tools (http://bio.cse.psu.edu/pipmaker/) [62]. Evolutionary sequence comparison of the elephant shark Pax6 loci with other species was prepared using the ‘glocal’ alignment program SLAGAN [85] with a window size of 100 bp and a minimal sequence identity of 70% and visualized using VISTA [86].
A neighbour-joining (NJ) tree for the Pax6 protein alignment was generated using MEGA (version 5.0.5; http://www.megasoftware.net/) [87]. A Poisson substitution model was used for distance calculation and 1000 bootstrap replicates were used for node support. The tree was viewed and edited using TreeView ver. 1.6.6 [88].
Identification of elephant shark BACs
The DNA prepared from 92,160 clones of an elephant shark BAC library (IMCB_Eshark BAC library; unpublished) were pooled in three dimensions and used for identifying BAC clones by three-step PCR screening. The 1.4× coverage sequence of the elephant shark genome [66] was searched for the Pax6 gene by BLAST. The search identified two scaffolds each containing a different Pax6 fragment. PCR primers were designed for these fragments and used to identify positive BAC clones (23H6 and 37E6). These BACs were sequenced completely. BACs overlapping these seed-BACs were identified by PCR screening and sequenced completely. Altogether, four BACs were sequenced to obtain the sequence of Pax6.1 locus (200B18, 50G15, 23H6 and 108J23) (GenBank JX135563) while two BACs were sequenced for the Pax6.2 locus (87E22 and 37E6) (GenBank JX135564).
Sequencing and assembly of BACs
BAC clones were sequenced using the standard shotgun sequencing method and gaps were filled by PCR amplification and primer walking. Sequencing was done using the BigDye Terminator Cycle Sequencing Kit (Applied Biosystems, USA). Sequences were processed and assembled using Phred-Phrap [89] and Consed [90]. Repetitive sequences were identified and masked using CENSOR at default settings [91]. Protein-coding genes were predicted using a combination of ab initio (e.g. FGENESH) and homology-based methods. Sequences for the human, chicken, lizard, Xenopus and zebrafish Pax6 loci and genes were extracted from the UCSC Browser (http://genome.ucsc.edu/).
RT–PCR and 5′RACE analyses of elephant shark Pax6 genes
RT–PCR was performed using primers for Pax6.1 and Pax6.2 exons designed to encompass at least one intron. cDNA from 13 different tissues of elephant shark (brain, eye, gills, heart, kidney, liver, muscle, ovary, pancreas, spleen, intestine, testis and uterus) were used as template. Actin was amplified as a control to check for the quality of cDNA. The cycling conditions were 95°C for 2 mins, followed by 35 cycles of 95°C for 30 secs, 60°C for 30 secs and 72°C for 2 mins. 5′RACE was performed using the SMART cDNA amplification kit (BD Clontech, USA) and Advantage 2 PCR enzyme mix (BD Clontech, USA). The RACE products were sequenced either directly or after cloning into modified pBluescript.
Cloning of conserved non-coding elements and genomic regions
CNEs selected for analysis in transgenic reporter assays were cloned by PCR amplification of the fragment containing the CNE plus flanking sequence from genomic DNA using Phusion high-fidelity polymerase (NEB). attB4 and attB1r sequences were included in the PCR primers for use with the Gateway recombination cloning system (Invitrogen). For zebrafish transgenic studies the amplified fragment was first cloned into the Gateway pP4P1r entry vector and sequenced using M13 forward and reverse primers for verification. Test elements in the pP4P1r vector were combined with a pDONR221 construct containing either a gata2 promoter-eGFP-polyA or a gata2 promoter-mCherry-polyA cassette, and recombined into a destination vector with a Gateway R4-R2 cassette flanked by Tol2 recombination sites [59], [92], [93].
For mouse transgenic LacZ reporter studies the selected fragments were PCR amplified using primers containing NotI and SalI restriction sites and cloned into pGEM-T Easy vector (Promega). Fragments were then transferred into the hsp68-LacZ containing p610+ vector using the NotI and SalI sites. For the elephant shark 6.2 NRE fragment the selected regions were amplified using primers containing Gateway compatible attB4 and attB1r sites, and cloned directly into an hsp68-LacZ vector containing a P4-P1r entry cassette. The forward primer also included an AscI site to allow subsequent micro-injection fragment isolation.
The following primers were used:
Medaka pax6.1 intron 7 : 5′-GCCCAACCAAGGTGAGCCTC-3′ and 5′-TTGGCAGCCATCTGAAGGTG
Medaka pax6.3 intron 7 : 5′-GCTGGGACCACACTCTCCTCCA-3′ and 5′-TGGAGTTCACCGAGATGCCGT-3′
Zebrafish pax6.1a intron7 : 5′-CCAATCAAGGTATGGCTGT-3′ and 5′-TGACTGTTGGCAACCATCTGA-3′
Zebrafish pax6.1b intron7 : 5′-CCAAATCAAGGTGAGACAGCCA-3′ and 5′-TCCTGTTGCTGGCAACCGTCT-3′
Zebrafish pax6.1a 7CE1 : 5′-CCAATCAAGGTATGGCTGT-3′ and 5′-GTTATGCCCTAAATCAAAGGCGT-3′
Zebrafish pax6.1a 7CE2 : 5′-GTCCATAGGCTGTTAGTTTGGGT-3′ and 5′-ACTGCACGTATTTCCCCCTAG-3′
Zebrafish pax6.1a 7CE3 : 5′-GCCACTGGTGTCTAACAAC-3′ and 5′-TTACAGCACTTTTCAGGCC-3′
Medaka pax6.1 7CE2 : 5′-TGGCTCCAATCCCCGTTATAGGA-3′ and 5′-AAGGATGCCGCATGTGAGGGT-3′
Elephant shark pax6.1 7CE2 : 5′-CTTCGACCATCAGCTGACAG-3′ and 5′-TCGGTCACTTTATGCCCACA-3′
Medaka pax6.3 7CE: 5′-GAAATGTGACAGCTGACAGGGT-3′ and 5′-CTTCATTTGGGGACTGAACAG-3′
Cm6.2NRE into p610+ and Tol2:
Cm6.2NRE forw: 5′-attB4-gcctcgcttcaaggccgatct-3′
Cm6.2NRE rev: 5′-attB1r-gccgtgcgggataaggtaga
Cm6.1 NRE into Tol2:
Cm6.1NRE forw: 5′-attB4-gctagaaccttcgcatctg
Cm6.2NRE rev: 5′-attB1r-gcatggcaaggctgcatgc
Zebrafish pax6.2_CNE1 into Tol2:
Drpax6.2_CNE1 forw: 5′-attB4-CCCTCATCCTCCCTCACATCT
Drpax6.2_CNE1 rev_short:5′-attB1r-TGTGACGTTGCGAGTGCGT
Drpax6.2_CNE1 rev_long:5′-attB1r-GCTCATACTGCGGTCCCAGA
Transgenic reporter mice
Production of transgenic mice by micro-injection into mouse oocytes was performed according to standard procedures. For analysis, embryos were collected at the appropriate stages, washed in PBS and fixed for 1 hour in a solution of 1% formaldehyde; 0.2% glutaraldehyde; 2 mM MgCl2; 5 mM EGTA and 0.02% NP-40 in PBS. After fixation the embryos were washed in PBS containing 0.02% NP-40, before being stained for several hours at 37°C in the dark in a solution containing 5 mM K3Fe(CN)6; 5 mM K4Fe(CN)6.3H2O; 2 mM MgCl2; 0.01% sodium deoxycholate; 0.02% NP-40 and 0.1% 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-gal). Embryos were photographed on a Leica MZ FLIII Microscope fitted with a Hamamatsu Orca-ER digital camera and a CRI micro-color filter. Table S1 contains details of the number of independent transgenic lines analysed and the observed sites of reporter gene expression.
Comparison of Pax6 synteny regions
Mapping of adjacent genes onto Pax6 synteny regions was done using the Ensembl and UCSC genome browsers, with additional information obtained using the Genomicus website v66.01 [79] (http://www.dyogen.ens.fr/genomicus-66.01/cgi-bin/search.pl).
rtPCR and RNA in situ analysis
Zebrafish Pax6.2 in Ensembl: zgc193504; Ensembl identifier: ENSDARG00000053364; Location chromosome 3 : 32,694,591–32,702,244.
RNA was isolated from pooled wild type zebrafish embryos from 1 dpf to 5 dpf using Tri-Reagent (Sigma). rtPCR was performed using Superscript III. PCR primers generating a fragment spanning several introns were designed for Pax6.2 and Isl1.
rtPCR primers for zebrafish Pax6.2:
forw: 5′-CTCAATGCACAGTCGGAGTG-3′
rev: 5′-TGCTGATTGAAGCTCTGCTGGT-3′
rtPCR primers for zebrafish Isl1:
islet1_fw CGGCGCACATATTCACATAC
islet1_rv TAAGCTTTAATACGACTCACTATAGGGAGAACGGACACGAACACATGAAA
mRNA in situ hybridization
RNA in situ hybridization on fish embryos was performed as previously described [94].
The medaka Pax6.1 RNA in situ probe was generated from the cDNA clone AJ000938 [63]; The RNA in situ probe for medaka Pax6.3 was generated from EST clone MF01SSA182E11 (Genbank BJ013007 and BJ027294).
The RNA in situ probe for zebrafish Pax6.2 was generated from the cDNA with primers:
T7 : 5′-TAATACGACTCACTATAGGTGCTGATTGAAGCTCTGCTGGT-3′
5′-CTCAATGCACAGTCGGAGTG-3′
Morpholino knock-down of zebrafish Pax6.2
A zebrafish Pax6.2 antisense morpholino oligonucleotide was obtained from Gene Tools, LLC, with the following sequence: 5′ GTCATACATCAAATGTCACCTGTGA 3′, directed against the translation start site of the gene.
As control we used the Gene ToolsLLC standard negative control morpholino: 5′ CCTCTTACCTCAGTTACAATTTATA 3′. The morpholinos were injected into 1 to 2-cell stage of at least 100 embryos to deliver an approximate amount of 2.5 ng per embryo.
Generation of zebrafish transgenic lines
Zebrafish were maintained in a recirculating water system according to standard protocol [95]. Embryos were obtained by breeding adult fish of standard stains (AB and WIK) and raised at 28.5°C as described [95]. Embryos were staged by hours post fertilization (hpf) as described [96].
Reporter plasmids were isolated using Qiagen miniprep columns and were given extra purification via a Qiagen PCR purification column (Qiagen), and diluted to 50 ng/microliter with DNAse/RNAse free water. tol2 transposase RNA was synthesized from a NotI-linearized pCS2-TP plasmid [93] using the SP6 mMessage mMachine kit (Ambion), and similarly diluted to 50 ng/microliter. Equal volumes of the reporter construct(s) and the transposase RNA were mixed immediately prior to injections to give an injection solution containing 25 ng/microliter of DNA and 25 ng/microliter of transposase RNA. 1–2 nl of the solution was micro-injected per embryo into the cytoplasm of 200 embryos at the 1 - to 2-cell stage. Embryos were screened for mosaic fluorescence at 1–5 days post-fertilization (dpf) and raised to adulthood. Germline transmission was identified by mating of sexually mature adults to wild-type fish and examining their progeny for fluorescence. Positive embryos were raised to adulthood and lines were maintained by outcrossing. Fish showing the best representative expression pattern for each construct were selected for confocal imaging. A summary table detailing numbers of independent lines analysed for each construct and their expression sites is included in Table S1.
Imaging of zebrafish reporter transgenic embryos
Embryos for imaging were treated with 0.003% PTU (1-phenyl 2-thio-urea) from 24 hpf to prevent pigmentation. Embryos selected for imaging were anaesthetised with tricaine and mounted in 1% low-melting agarose. Images were taken on a Zeiss LSM510 confocal microscope and processed using Fiji software.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. CarrollSB (2008) Evo-devo and an expanding evolutionary synthesis: a genetic theory of morphological evolution. Cell 134 : 25–36.
2. NeiM (2007) The new mutation theory of phenotypic evolution. Proc Natl Acad Sci U S A 104 : 12235–12242.
3. Prud'hommeB, GompelN, CarrollSB (2007) Emerging principles of regulatory evolution. Proc Natl Acad Sci U S A 104 Suppl 18605–8612.
4. Wittkopp PJ, Kalay G Cis-regulatory elements: molecular mechanisms and evolutionary processes underlying divergence. Nat Rev Genet 13 : 59–69.
5. GehringWJ (2002) The genetic control of eye development and its implications for the evolution of the various eye-types. Int J Dev Biol 46 : 65–73.
6. GehringWJ (1996) The master control gene for morphogenesis and evolution of the eye. Genes Cells 1 : 11–15.
7. QuiringR, WalldorfU, KloterU, GehringWJ (1994) Homology of the eyeless gene of Drosophila to the Small eye gene in mice and Aniridia in humans. Science 265 : 785–789.
8. CallaertsP, HalderG, GehringWJ (1997) PAX-6 in development and evolution. Annu Rev Neurosci 20 : 483–532.
9. HalderG, CallaertsP, GehringWJ (1995) Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila. Science 267 : 1788–1792.
10. SimpsonTI, PriceDJ (2002) Pax6; a pleiotropic player in development. Bioessays 24 : 1041–1051.
11. Georgala PA, Carr CB, Price DJ The role of Pax6 in forebrain development. Dev Neurobiol 71 : 690–709.
12. OsumiN, ShinoharaH, Numayama-TsurutaK, MaekawaM (2008) Concise review: Pax6 transcription factor contributes to both embryonic and adult neurogenesis as a multifunctional regulator. Stem Cells 26 : 1663–1672.
13. TonCC, HirvonenH, MiwaH, WeilMM, MonaghanP, et al. (1991) Positional cloning and characterization of a paired box - and homeobox-containing gene from the aniridia region. Cell 67 : 1059–1074.
14. SisodiyaSM, FreeSL, WilliamsonKA, MitchellTN, WillisC, et al. (2001) PAX6 haploinsufficiency causes cerebral malformation and olfactory dysfunction in humans. Nat Genet 28 : 214–216.
15. BamiouDE, MusiekFE, SisodiyaSM, FreeSL, MitchellTN, et al. (2004) Defective auditory interhemispheric transfer in a patient with a PAX6 mutation. Neurology 62 : 489–490.
16. YasudaT, KajimotoY, FujitaniY, WatadaH, YamamotoS, et al. (2002) PAX6 mutation as a genetic factor common to aniridia and glucose intolerance. Diabetes 51 : 224–230.
17. GlaserT, JepealL, EdwardsJG, YoungSR, FavorJ, et al. (1994) PAX6 gene dosage effect in a family with congenital cataracts, aniridia, anophthalmia and central nervous system defects. Nat Genet 7 : 463–471.
18. MatsuoT, Osumi-YamashitaN, NojiS, OhuchiH, KoyamaE, et al. (1993) A mutation in the Pax-6 gene in rat small eye is associated with impaired migration of midbrain crest cells. Nat Genet 3(4): 299–304.
19. HillRE, FavorJ, HoganBL, TonCC, SaundersGF, et al. (1991) Mouse small eye results from mutations in a paired-like homeobox-containing gene. Nature 354 : 522–525.
20. SchedlA, RossA, LeeM, EngelkampD, RashbassP, et al. (1996) Influence of PAX6 gene dosage on development: overexpression causes severe eye abnormalities. Cell 86 : 71–82.
21. ChanasSA, CollinsonJM, RamaeshT, DoraN, KleinjanDA, et al. (2009) Effects of elevated Pax6 expression and genetic background on mouse eye development. Invest Ophthalmol Vis Sci 50 : 4045–4059.
22. GriffinC, KleinjanDA, DoeB, van HeyningenV (2002) New 3′ elements control Pax6 expression in the developing pretectum, neural retina and olfactory region. Mech Dev 112 : 89–100.
23. KleinjanDA, SeawrightA, ChildsAJ, van HeyningenV (2004) Conserved elements in Pax6 intron 7 involved in (auto)regulation and alternative transcription. Dev Biol 265 : 462–477.
24. KleinjanDA, SeawrightA, MellaS, CarrCB, TyasDA, et al. (2006) Long-range downstream enhancers are essential for Pax6 expression. Dev Biol 299 : 563–581.
25. KleinjanDA, SeawrightA, SchedlA, QuinlanRA, DanesS, et al. (2001) Aniridia-associated translocations, DNase hypersensitivity, sequence comparison and transgenic analysis redefine the functional domain of PAX6. Hum Mol Genet 10 : 2049–2059.
26. McBride DJ, Buckle A, van Heyningen V, Kleinjan DA DNaseI hypersensitivity and ultraconservation reveal novel, interdependent long-range enhancers at the complex Pax6 cis-regulatory region. PLoS ONE 6: e28616 doi:10.1371/journal.pone.0028616.
27. KammandelB, ChowdhuryK, StoykovaA, AparicioS, BrennerS, et al. (1999) Distinct cis-essential modules direct the time-space pattern of the Pax6 gene activity. Dev Biol 205 : 79–97.
28. XuPX, ZhangX, HeaneyS, YoonA, MichelsonAM, et al. (1999) Regulation of Pax6 expression is conserved between mice and flies. Development 126 : 383–395.
29. NavratilovaP, FredmanD, HawkinsTA, TurnerK, LenhardB, et al. (2009) Systematic human/zebrafish comparative identification of cis-regulatory activity around vertebrate developmental transcription factor genes. Dev Biol 327 : 526–540.
30. FantesJ, RedekerB, BreenM, BoyleS, BrownJ, et al. (1995) Aniridia-associated cytogenetic rearrangements suggest that a position effect may cause the mutant phenotype. Hum Mol Genet 4 : 415–422.
31. LauderdaleJD, WilenskyJS, OliverER, WaltonDS, GlaserT (2000) 3′ deletions cause aniridia by preventing PAX6 gene expression. Proc Natl Acad Sci U S A 97 : 13755–13759.
32. RobinsonDO, HowarthRJ, WilliamsonKA, van HeyningenV, BealSJ, et al. (2008) Genetic analysis of chromosome 11p13 and the PAX6 gene in a series of 125 cases referred with aniridia. Am J Med Genet A 146A: 558–569.
33. KleinjanDA, SeawrightA, ElgarG, van HeyningenV (2002) Characterization of a novel gene adjacent to PAX6, revealing synteny conservation with functional significance. Mamm Genome 13 : 102–107.
34. KikutaH, LaplanteM, NavratilovaP, KomisarczukAZ, EngstromPG, et al. (2007) Genomic regulatory blocks encompass multiple neighboring genes and maintain conserved synteny in vertebrates. Genome Res 17 : 545–555.
35. EpsteinJA, GlaserT, CaiJ, JepealL, WaltonDS, et al. (1994) Two independent and interactive DNA-binding subdomains of the Pax6 paired domain are regulated by alternative splicing. Genes Dev 8 : 2022–2034.
36. KimJ, LauderdaleJD (2008) Overexpression of pairedless Pax6 in the retina disrupts corneal development and affects lens cell survival. Dev Biol 313 : 434–454.
37. LakowskiJ, MajumderA, LauderdaleJD (2007) Mechanisms controlling Pax6 isoform expression in the retina have been conserved between teleosts and mammals. Dev Biol 307 : 498–520.
38. KimJ, LauderdaleJD (2006) Analysis of Pax6 expression using a BAC transgene reveals the presence of a paired-less isoform of Pax6 in the eye and olfactory bulb. Dev Biol 292 : 486–505.
39. MikkolaI, BruunJA, HolmT, JohansenT (2001) Superactivation of Pax6-mediated transactivation from paired domain-binding sites by dna-independent recruitment of different homeodomain proteins. J Biol Chem 276 : 4109–4118.
40. ChristoffelsA, KohEG, ChiaJM, BrennerS, AparicioS, et al. (2004) Fugu genome analysis provides evidence for a whole-genome duplication early during the evolution of ray-finned fishes. Mol Biol Evol 21 : 1146–1151.
41. HoeggS, BrinkmannH, TaylorJS, MeyerA (2004) Phylogenetic timing of the fish-specific genome duplication correlates with the diversification of teleost fish. J Mol Evol 59 : 190–203.
42. TaylorJS, BraaschI, FrickeyT, MeyerA, Van de PeerY (2003) Genome duplication, a trait shared by 22000 species of ray-finned fish. Genome Res 13 : 382–390.
43. PostlethwaitJ, AmoresA, CreskoW, SingerA, YanYL (2004) Subfunction partitioning, the teleost radiation and the annotation of the human genome. Trends Genet 20 : 481–490.
44. DehalP, BooreJL (2005) Two rounds of whole genome duplication in the ancestral vertebrate. PLoS Biol 3: e314 doi:10.1371/journal.pbio.0030314.
45. Ohno S (1970) Evolution by Gene Duplication. New York: Springer-Verlag.
46. ForceA, LynchM, PickettFB, AmoresA, YanYL, et al. (1999) Preservation of duplicate genes by complementary, degenerative mutations. Genetics 151 : 1531–1545.
47. LynchM, ConeryJS (2000) The evolutionary fate and consequences of duplicate genes. Science 290 : 1151–1155.
48. PrinceVE, PickettFB (2002) Splitting pairs: the diverging fates of duplicated genes. Nat Rev Genet 3 : 827–837.
49. ElgarG, SandfordR, AparicioS, MacraeA, VenkateshB, et al. (1996) Small is beautiful: comparative genomics with the pufferfish (Fugu rubripes). Trends Genet 12 : 145–150.
50. FisherS, GriceEA, VintonRM, BesslingSL, McCallionAS (2006) Conservation of RET regulatory function from human to zebrafish without sequence similarity. Science 312 : 276–279.
51. Navratilova P, Fredman D, Lenhard B, Becker TS Regulatory divergence of the duplicated chromosomal loci sox11a/b by subpartitioning and sequence evolution of enhancers in zebrafish. Mol Genet Genomics 283 : 171–184.
52. BladerP, LamCS, RastegarS, ScardigliR, NicodJC, et al. (2004) Conserved and acquired features of neurogenin1 regulation. Development 131 : 5627–5637.
53. McEwenGK, GoodeDK, ParkerHJ, WoolfeA, CallawayH, et al. (2009) Early evolution of conserved regulatory sequences associated with development in vertebrates. PLoS Genet 5: e1000762 doi:10.1371/journal.pgen.1000762.
54. HollandLZ, AlbalatR, AzumiK, Benito-GutierrezE, BlowMJ, et al. (2008) The amphioxus genome illuminates vertebrate origins and cephalochordate biology. Genome Res 18 : 1100–1111.
55. WangJ, LeeAP, KodziusR, BrennerS, VenkateshB (2009) Large number of ultraconserved elements were already present in the jawed vertebrate ancestor. Mol Biol Evol 26 : 487–490.
56. MikkelsenTS, WakefieldMJ, AkenB, AmemiyaCT, ChangJL, et al. (2007) Genome of the marsupial Monodelphis domestica reveals innovation in non-coding sequences. Nature 447 : 167–177.
57. Lee AP, Kerk SY, Tan YY, Brenner S, Venkatesh B Ancient vertebrate conserved noncoding elements have been evolving rapidly in teleost fishes. Mol Biol Evol 28 : 1205–1215.
58. NornesS, ClarksonM, MikkolaI, PedersenM, BardsleyA, et al. (1998) Zebrafish contains two pax6 genes involved in eye development. Mech Dev 77 : 185–196.
59. KleinjanDA, BancewiczRM, GautierP, DahmR, SchonthalerHB, et al. (2008) Subfunctionalization of duplicated zebrafish pax6 genes by cis-regulatory divergence. PLoS Genet 4: e29 doi:10.1371/journal.pgen.0040029.
60. VenkateshB, KirknessEF, LohYH, HalpernAL, LeeAP, et al. (2006) Ancient noncoding elements conserved in the human genome. Science 314 : 1892.
61. MilesC, ElgarG, ColesE, KleinjanDJ, van HeyningenV, et al. (1998) Complete sequencing of the Fugu WAGR region from WT1 to PAX6: dramatic compaction and conservation of synteny with human chromosome 11p13. Proc Natl Acad Sci U S A 95 : 13068–13072.
62. SchwartzS, ElnitskiL, LiM, WeirauchM, RiemerC, et al. (2003) MultiPipMaker and supporting tools: Alignments and analysis of multiple genomic DNA sequences. Nucleic Acids Res 31 : 3518–3524.
63. LoosliF, KosterRW, CarlM, KroneA, WittbrodtJ (1998) Six3, a medaka homologue of the Drosophila homeobox gene sine oculis is expressed in the anterior embryonic shield and the developing eye. Mech Dev 74 : 159–164.
64. TzoulakiI, WhiteIM, HansonIM (2005) PAX6 mutations: genotype-phenotype correlations. BMC Genet 6 : 27.
65. Inoue JG, Miya M, Lam K, Tay BH, Danks JA, et al. Evolutionary origin and phylogeny of the modern holocephalans (Chondrichthyes: Chimaeriformes): a mitogenomic perspective. Mol Biol Evol 27 : 2576–2586.
66. VenkateshB, KirknessEF, LohYH, HalpernAL, LeeAP, et al. (2007) Survey sequencing and comparative analysis of the elephant shark (Callorhinchus milii) genome. PLoS Biol 5: e101 doi:10.1371/journal.pbio.0050101.
67. CrollaJA, van HeyningenV (2002) Frequent chromosome aberrations revealed by molecular cytogenetic studies in patients with aniridia. Am J Hum Genet 71 : 1138–1149.
68. PlazaS, DozierC, LangloisMC, SauleS (1995) Identification and characterization of a neuroretina-specific enhancer element in the quail Pax-6 (Pax-QNR) gene. Mol Cell Biol 15 : 892–903.
69. SaitouN, NeiM (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4 : 406–425.
70. SharpeJ, AhlgrenU, PerryP, HillB, RossA, et al. (2002) Optical projection tomography as a tool for 3D microscopy and gene expression studies. Science 296 : 541–545.
71. HaubstN, BergerJ, RadjendiraneV, GrawJ, FavorJ, et al. (2004) Molecular dissection of Pax6 function: the specific roles of the paired domain and homeodomain in brain development. Development 131 : 6131–6140.
72. GrawJ, LosterJ, PukO, MunsterD, HaubstN, et al. (2005) Three novel Pax6 alleles in the mouse leading to the same small-eye phenotype caused by different consequences at target promoters. Invest Ophthalmol Vis Sci 46 : 4671–4683.
73. Dames P, Puff R, Weise M, Parhofer KG, Goke B, et al. Relative roles of the different Pax6 domains for pancreatic alpha cell development. BMC Dev Biol 10 : 39.
74. Lowe CB, Kellis M, Siepel A, Raney BJ, Clamp M, et al. Three periods of regulatory innovation during vertebrate evolution. Science 333 : 1019–1024.
75. McGaugheyDM, VintonRM, HuynhJ, Al-SaifA, BeerMA, et al. (2008) Metrics of sequence constraint overlook regulatory sequences in an exhaustive analysis at phox2b. Genome Res 18 : 252–260.
76. CinarHN, ChisholmAD (2004) Genetic analysis of the Caenorhabditis elegans pax-6 locus: roles of paired domain-containing and nonpaired domain-containing isoforms. Genetics 168 : 1307–1322.
77. CzernyT, HalderG, KloterU, SouabniA, GehringWJ, et al. (1999) twin of eyeless, a second Pax-6 gene of Drosophila, acts upstream of eyeless in the control of eye development. Mol Cell 3 : 297–307.
78. DominguezM, Ferres-MarcoD, Gutierrez-AvinoFJ, SpeicherSA, BeneytoM (2004) Growth and specification of the eye are controlled independently by Eyegone and Eyeless in Drosophila melanogaster. Nat Genet 36 : 31–39.
79. Flicek P, Amode MR, Barrell D, Beal K, Brent S, et al. Ensembl 2012. Nucleic Acids Res 40: D84–90.
80. RiceP, LongdenI, BleasbyA (2000) EMBOSS: the European Molecular Biology Open Software Suite. Trends Genet 16 : 276–277.
81. AltschulSF, MaddenTL, SchafferAA, ZhangJ, ZhangZ, et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25 : 3389–3402.
82. BirneyE, ClampM, DurbinR (2004) GeneWise and Genomewise. Genome Res 14 : 988–995.
83. LarkinMA, BlackshieldsG, BrownNP, ChennaR, McGettiganPA, et al. (2007) Clustal W and Clustal X version 2.0. Bioinformatics 23 : 2947–2948.
84. Nicholas KaN, HB (1997) GeneDoc: A tool for editing and annotating multiple sequence alignments.
85. BrudnoM, MaldeS, PoliakovA, DoCB, CouronneO, et al. (2003) Glocal alignment: finding rearrangements during alignment. Bioinformatics 19 Suppl 1i54–62.
86. FrazerKA, PachterL, PoliakovA, RubinEM, DubchakI (2004) VISTA: computational tools for comparative genomics. Nucleic Acids Res 32: W273–279.
87. TamuraK, PetersonD, PetersonN, StecherG, NeiM, et al. (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28 : 2731–2739.
88. PageRD (1996) TreeView: an application to display phylogenetic trees on personal computers. Comput Appl Biosci 12 : 357–358.
89. EwingB, HillierL, WendlMC, GreenP (1998) Base-calling of automated sequencer traces using phred. I. Accuracy assessment. Genome Res 8 : 175–185.
90. GordonD, AbajianC, GreenP (1998) Consed: a graphical tool for sequence finishing. Genome Res 8 : 195–202.
91. KohanyO, GentlesAJ, HankusL, JurkaJ (2006) Annotation, submission and screening of repetitive elements in Repbase: RepbaseSubmitter and Censor. BMC Bioinformatics 7 : 474.
92. FisherS, GriceEA, VintonRM, BesslingSL, UrasakiA, et al. (2006) Evaluating the biological relevance of putative enhancers using Tol2 transposon-mediated transgenesis in zebrafish. Nat Protoc 1 : 1297–1305.
93. KawakamiK (2004) Transgenesis and gene trap methods in zebrafish by using the Tol2 transposable element. Methods Cell Biol 77 : 201–222.
94. ThisseC, ThisseB (2008) High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat Protoc 3 : 59–69.
95. Westerfield M (2007) The Zebrafish Book: A guide for the laboratory use of zebrafish Danio rerio. Eugene, OR: University of Oregon Press.
96. KimmelCB, BallardWW, KimmelSR, UllmannB, SchillingTF (1995) Stages of embryonic development of the zebrafish. Dev Dyn 203 : 253–310.
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2013 Číslo 1
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Function and Regulation of , a Gene Implicated in Autism and Human Evolution
- An Insertion in 5′ Flanking Region of Causes Blue Eggshell in the Chicken
- Comprehensive Methylome Characterization of and at Single-Base Resolution
- Susceptibility Loci Associated with Specific and Shared Subtypes of Lymphoid Malignancies
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy