
Dopamine Signalling in Mushroom Bodies Regulates Temperature-Preference Behaviour in
The ability to respond to environmental temperature variation is essential for survival in animals. Flies show robust temperature-preference behaviour (TPB) to find optimal temperatures. Recently, we have shown that Drosophila mushroom body (MB) functions as a center controlling TPB. However, neuromodulators that control the TPB in MB remain unknown. To identify the functions of dopamine in TPB, we have conducted various genetic studies in Drosophila. Inhibition of dopamine biosynthesis by genetic mutations or treatment with chemical inhibitors caused flies to prefer temperatures colder than normal. We also found that dopaminergic neurons are involved in TPB regulation, as the targeted inactivation of dopaminergic neurons by expression of a potassium channel (Kir2.1) induced flies with the loss of cold avoidance. Consistently, the mutant flies for dopamine receptor gene (DopR) also showed a cold temperature preference, which was rescued by MB–specific expression of DopR. Based on these results, we concluded that dopamine in MB is a key component in the homeostatic temperature control of Drosophila. The current findings will provide important bases to understand the logic of thermosensation and temperature preference decision in Drosophila.
Published in the journal:
. PLoS Genet 7(3): e32767. doi:10.1371/journal.pgen.1001346
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1001346
Summary
The ability to respond to environmental temperature variation is essential for survival in animals. Flies show robust temperature-preference behaviour (TPB) to find optimal temperatures. Recently, we have shown that Drosophila mushroom body (MB) functions as a center controlling TPB. However, neuromodulators that control the TPB in MB remain unknown. To identify the functions of dopamine in TPB, we have conducted various genetic studies in Drosophila. Inhibition of dopamine biosynthesis by genetic mutations or treatment with chemical inhibitors caused flies to prefer temperatures colder than normal. We also found that dopaminergic neurons are involved in TPB regulation, as the targeted inactivation of dopaminergic neurons by expression of a potassium channel (Kir2.1) induced flies with the loss of cold avoidance. Consistently, the mutant flies for dopamine receptor gene (DopR) also showed a cold temperature preference, which was rescued by MB–specific expression of DopR. Based on these results, we concluded that dopamine in MB is a key component in the homeostatic temperature control of Drosophila. The current findings will provide important bases to understand the logic of thermosensation and temperature preference decision in Drosophila.
Introduction
To counteract the harmful effects of environmental temperature fluctuation, animals use homeostatic thermoregulatory mechanisms and/or behavioral responses to maintain optimal body temperature. Mammals regulate their body temperatures with various physiological responses such as change of metabolic rates, vasomotor control, and sweating. Due to the large surface area-to-volume ratio of its body, Drosophila maintains body temperature by heat exchange with the surrounding environment. In a genetically controlled process, flies track to an environmental temperature as close as possible to their desired body temperature [1]–[3]. The behavioural responses in Drosophila can be recorded easily by examining TPB on a thermal gradient plate [1], [4], [5]. Wild type fruit flies show a strong response to external temperature and robust TPB peaking at 24°C on the temperature gradient [4].
To identify the molecular components and neural circuits regulating thermosensation and body temperature regulation, various studies such as calcium imaging, genetic screens, and electrophysiology have been done in fruit flies, C. elegans, mice, and cultured cells [6]–[12]. From these, various thermo-transient receptor potential (TRP) channels and cyclic nucleotide-gated channels have been found to be molecular sensors of temperature. The perceived temperature information generated in neurons bearing these sensor molecules is transmitted to a higher order brain center. Here, the sensed temperature is compared to a set point temperature to determine whether it is too low or too high [2]. In vertebrate, the hypothalamic preoptic area is known to be the higher order brain center interpreting the temperature information [13].
In Drosophila, the temperature that is instinctively preferred plays a role equivalent to the set point temperature in mammals. Three thermo-TRP channels have been reported as molecular sensors of hot or warm temperature in Drosophila. Painless, a thermo-TRP channel, is important for avoidance of hazardous heat above 40°C [14]. Pyrexia, another thermo-TRP channel, is activated around 40°C and acts to prevent paralysis during high temperature stress [15]. dTRPA1 is activated above 27°C and essential for avoidance of warm temperature, and the loss-of-function mutants of dTRPA1 prefer warm temperature [16]. In addition to the discovery of these thermo-TRP channels, various genetic studies to understand the brain center, and the responsible neuromodulators and neural circuits of TPB in Drosophila have been actively pursued [1], [5], [16].
The adult Drosophila brain includes about 100,000 neurons whose projections cluster in neuropil structures to make two symmetric hemispheres [17]. The central complex and the mushroom body (MB) are two prominent structures in the central brain. The central complex is located centrally between the two hemispheres and consists of four interconnected neuropils: the ellipsoid body, the fan-shaped body, the protocerebral-bridge, and the nodule [18], [19]. The central complex is involved in behaviours such as locomotion, visual flight, courtship, and olfactory learning task [20], [21].
However, MB is involved in behaviours like sleep [22], decision making [23], and learinig [24]. Recently we showed that MB acts as a higher order brain center regulating TPB in Drosophila [1]. MB is made up of approximately 2,500 Kenyon cells in each hemisphere, which send out dendrites to the calyx [24]. Kenyon cells are grouped into one of three neuron classes, whose axons form morphologically and functionally distinct lobes of the αβ, α′β′ and γ lobes. For TPB, the αβ lobes are necessary while the γ lobe is dispensable [1]. We also showed that the cAMP-PKA signalling pathway is the key regulator working inside of MB to determine the temperature that Drosophila prefers [1].
Fifteen clusters of six-hundred dopaminergic neurons are distributed in the Drosophila brain [25], [26]. The dopaminergic neurons are broadly and stereotypically localized, and show particularly dense projections into MB [27]. For example, protocerebral posterior lateral 1 cluster which innervates MB is reported as routes for reinforcement and retrieval in memory formation [28], [29]. Similar to mammals, dopamine receptors are divided into two major subfamilies in Drosophila. Two D1-like receptors, DopR and DpoR2, and a D2-like receptor, D2R, have been identified [30]. Interestingly, DopR is highly expressed in MB and the central complex [31], and mediates learning [32], [33], caffeine-induced wakefulness [34], arousal [35], and ethanol-stimulated locomotion [36]. DopR2 is expressed in MB and is involved in larval aversive olfactory learning [37], [38]. D2R is expressed in some discrete cells in central nervous system and regulates locomotor activity [30]. However, it has remained unknown that dopamine is involved in TPB regulation in Drosophila.
To uncover the relationship between the dopamine system and TPB, we performed various genetic studies in Drosophila. Dopamine synthesis and dopaminergic neurons are critical in TPB regulation. In addition, we showed that DopR is essential to control TPB in the αβ lobes of MB. These results strongly suggested that dopamine is an important neuromodulator for TPB in Drosophila.
Results
Dopamine synthesis is required for TPB regulation
Our previous results have shown that cAMP signalling in MB modulates TPB [1]. Since dopamine is an important modulator of cAMP [39], [40] and dopaminergic neurons have a strong innervation profile in MB [27], we suspected that the dopamine system can modulate TPB in Drosophila.
The enzymatic reaction of tyrosine hydroxylase (TH), which converts tyrosine to L-DOPA (Figure 1A), is the rate limiting step in dopamine biosynthesis [25]. TH is encoded by pale locus in Drosophila. It is also known that loss of one copy of the TH gene can cause severe reduction of dopamine levels in the animal brain [25]. This led us to test a pale mutant for TPB as the first step to explore the involvement of dopamine signalling in the regulation of TPB. In the TPB assay, the experimental procedure was described previously [1]. The Avoidance Index against Low Temperatures (AILow) and Avoidance Index against High Temperatures (AIHigh) were calculated using the formulas shown in Figure 1B [1], [6]. The pale4/+ heterozygote showed abnormal TPB and the animals spread over a wide range of temperature colder than 24°C (Figure 2A). Their AILow was less than 34% of the wild type control w1118, while AIHigh was normal (Figure 2A and Table S1).

To reduce the biosynthesis of dopamine further, we fed pale4/+ flies with a TH inhibitor, α-methyl-p-tyrosine methyl ester (AMPT), for 4 days. In the TPB assay, these flies showed a more severe defect in their ability to avoid low temperature (Figure 2A). The flies spread almost randomly over all temperature ranges below 24°C (Figure 2A). The AILow of the flies was below 0 (Figure 2A and Table S1).

On the other hand, the reduced AILow of pale4/+ flies was ameliorated by introducing a genomic DNA fragment of the pale locus into the pale4/+ flies (Figure 2B). The genomic DNA fragment of the pale locus was reported to rescue pale flies from lethality [41]. The AILow of pale4/+ flies with the pale genomic DNA was 91% of wild type (Figure 2B and Table S1). This confirmed that the abnormal TPB of the pale4/+ flies was indeed resulted from a defect in the TH gene. These results strongly suggested that dopamine is a key neurotransmitter controlling fly temperature preference.
To convince ourselves further that dopamine is involved in TPB regulation, we examined a dopa decarboxylase (DDC) mutant. DDC converts L-DOPA to dopamine (Figure 1A). As shown in Figure 2C, flies homozygous for a DDC hypomorphic allele DdcDE1 showed reduced cold avoidance. The AILow of DdcDE1/DdcDE1 flies was 26% of wild type (Figure 2C and Table S1).
To examine whether further reduction of DDC activity worsens the ability of flies to avoid cold temperature, we fed the DdcDE1/ DdcDE1 flies with 3-hydroxy benzyl hydrazine (HBH). HBH is a potent DDC inhibitor [42]. As expected, the DDC mutant flies fed HBH showed almost complete loss of cold avoidance in the TPB assay (Figure 2C). Some flies even moved to the coldest region of the temperature gradient plate (Figure 2C). The AILow of these flies was −0.23 (Figure 2C and Table S1).
These data suggested that dopamine biosynthesis is critical for flies to avoid cold temperature. This was supported further by over expression experiments. When TH or DDC was over expressed separately in dopaminergic neurons, the flies showed normal TPB (Figure 3A, 3B and Table S2). However, when TH and DDC were over expressed together in dopaminergic neurons, the flies became thermophilic or hypersensitive to cold temperature (Figure 3A and 3C). Their AILow was higher than that of wild type while their AIHigh was only 39% of wild type (Figure 3C and Table S2).

Dopaminergic neurons are required for TPB regulation
To gain more evidence of TPB control by dopamine, we examined whether the normal function of dopaminergic neurons is essential for normal TPB. We over expressed tetanus toxin (TNT) and potassium channel Kir2.1 in dopaminergic neurons with TH-Gal4 and dopamine transporter (DAT)-Gal4. TNT only blocks the neural activity requiring n-Syb-dependent synaptic vesicle fusion [43], while Kir2.1 hyperpolarizes all neurons and suppresses action potential firing [43], [44]. Unfortunately, Kir2.1 when combined with TH-Gal4 or DAT-Gal4 killed flies before they hatched. In contrast, TH-Gal4>TNT and DAT-Gal4>TNT flies were viable and both spread over temperature ranges broader than wild type, mostly over the areas colder than 25°C (Figure 4A, 4B and Table S3).
To exclude the possibility of developmental defects induced by TNT and Kir2.1, we combined UAS-TNT and UAS-Kir2.1 with the Temporal and Regional Gene Expression Targeting system (TARGET) [45], which employs temperature-sensitive Gal80 protein (Gal80ts) under the control of tubulin promoter (tub-Gal80ts). In this system, the transcriptional activator of Gal4 is suppressed at 18°C but active at 32°C.

As expected, TH-Gal4>TNT flies with tub-Gal80ts showed normal TPB when they were raised continually at 18°C (Figure 5A and 5C). However, they unexpectedly showed normal TPB even after exposure to 32°C for 16 hr to induce TNT before the TPB assay (Figure 5B and 5D). We obtained similar results when we used DAT-Gal4 instead of TH-Gal4 (data not shown). It might be possible that temporal induction of TNT for 16 hr in the adult stage does not accumulate enough TNT to inactivate dopaminergic neurons sufficiently, while life-time induction of TNT with TH-Gal4 or DAT-Gal4 without Gal80ts does accumulate sufficient TNT. It is also known that TNT might not block synaptic vesicle release strongly enough to block all neuronal activities [43], [46]. On the other hand, it cannot be excluded that the abnormal TPB of TH-Gal4>TNT and DAT-Gal4>TNT flies shown in Figure 4A and 4B was resulted from developmental defects induced by TNT.

Because we obtained unexpected data from TH-Gal4>TNT;tub-Gal80ts/+ or DAT-Gal4>TNT;tub-Gal80ts/+ flies, we used UAS-Kir2.1 instead of TNT in combination with TH-Gal4;tub-Gal80ts. Kir2.1 is a stronger neuronal inactivator than TNT [43]. When TH-Gal4>Kir2.1;tub-Gal80ts/+ flies were raised continually at 18°C, they showed expected normal TPB (Figure 5A and 5E). However, when the flies were raised at 18°C until 3-day-old adult and exposed to 32°C for 16 hr just before the TPB assay, they lost cold avoidance severely. Most of the flies moved towards the 15°C area, which is the low temperature limit of the assay apparatus and their AILow was −0.2 (Figure 5B, 5F and Table S4). Similar TPB was also observed when DAT-Gal4 was used instead of TH-Gal4 (Figure 5G and 5H). DAT-Gal4>Kir2.1;tub-Gal80ts/+ flies spread over an area of cold temperature when they were exposed to 32°C for 16 hr before the TPB assay (Figure 5B and 5H).
From the above results, we concluded that defective dopamine biosynthesis or inactivation of dopaminergic neurons causes the fly has abnormal TPB. These results strongly suggested that dopamine signalling controls TPB in Drosophila.
DopR is important for TPB
We next examined whether the activities of dopamine receptors are required for TPB. Drosophila has three subtypes of dopamine receptor; DopR, DopR2, and D2R [30]. dumb3 is a hypomorphic allele of DopR, in which most MB expression of DopR was removed by a P element insertion [33]. dumb3/dumb3 homozygote flies exhibited reduced ability to avoid cold temperature (Figure 6A). Their AILow was lower than 50% of wild type (Figure 6A and Table S5). Consistently, flies having one copy of a deficiency chromosome Df(3R)red-P52, in which DopR is deleted, preferred temperatures colder than normal (Figure 6B). The axis of distribution profile of flies over the temperature gradient was shifted from 24°C to 21°C (Figure 6B). More convincingly, when this deficiency chromosome was combined with dumb3 chromosome, resulting in the genotype of dumb3/Df(3R)red-P52, the flies lost cold avoidance furthermore (Figure 6C). Their AILow was 23% of wild type (Figure 6C and Table S5).

There are no available mutant alleles for DopR2 and D2R. Therefore, we employed dsRNAi technology with and without UAS-Dcr2, which is known to enhance the effect of dsRNAi in neurons [47]. However, none of the RNAi transgenes reduced the transcript levels of these two receptors by more than 40% and the flies had normal TPB. Accordingly, we could not ascertain whether DopR2 or D2R is required for TPB control.
DopR regulates TPB in MB
Taken together, our results shown above demonstrated that dopamine signalling is a key factor controlling TPB in Drosophila. This finding raised the question as to which brain parts or neurons are the targets of dopamine signalling in TPB control. A clue was obtained from the results that we reported recently, which showed that MB is the brain center controlling TPB [1]. Importantly, DopR and DopR2 are expressed in MB [31]. However, they are also expressed in other Drosophila nerves, including the central complex and several neuro-secretory cells in the adult brain [31]. We confirmed that DopR is highly enriched in MB and DopR is also expressed in the central complex ellipsoid body, fan-shaped body, and nodules as previously reported (Figure 7A and [31]). In dumb3/dumb3 homozygote flies, DopR gene expression was strongly reduced in overall MB and the central complex (Figure 7B).

To examine if MB is the location where DopR regulates TPB, we generated flies in which DopR expression is maintained in MB. This was achieved by ectopic expression of the UAS-DopR transgene with c309 or MB247 in the DopR mutant background of dumb3/dumb3 (Figure 7C and 7D). c309 and MB247 drive gene expression specifically in MB.
As shown in Figure 7C, dumb3/dumb3 flies spread widely over the temperature gradient lower than 25°C. In contrast, the artificial restoration of DopR levels with c309>UAS-DopR transgene enabled dumb3/dumb3 to gather sharply around the 25°C area (Figure 7C). In other words, the flies displayed almost completely restored normal TPB. The AILow of the flies was 99% of wild type (Figure 7C and Table S6). Consistently, c309>DopR;dumb3/dumb3 flies also showed rescued TPB compared with c309/+;dumb3/dumb3 and UAS-DopR/+;dumb3/dumb3 flies (Figure S1A). This means that the abnormal TPB of dumb3/dumb3 homozygote flies was caused by reduced expression of DopR in the region of c309 expression and that MB may be the location where DopR regulate TPB. However, we could not completely exclude the possibility that the central complex participated in the TPB regulation of DopR as the expression of c309 was observed in other regions of the brain including the central complex (Figure S2A and [48]).
To confirm the specific role of MB, we next tested MB247 which strongly induces gene expression in the αβ and γ lobes with very low background expression (Figure S2B and [49]). When we used MB247 instead of c309 (Figure 7D, Figure S1B, and Table S6), MB247> DopR;dumb3/dumb3 flies also regained preference for 25°C, which is the normal preferred body temperature (Figure 7D and Figure S1B). They showed the AILow and AIHigh values comparable to those of wild type flies (Figure 7D and Table S6). This means that the restoration of DopR levels only in MB enabled dumb3/dumb3 to behave like wild type flies. These data indicated that dopamine signalling in MB is critical for TPB.
In the same way, we generated flies to rescue the cryophilic TPB of Df(3R)red-P52/+ by ectopic expression of UAS-DopR with MB-specific Gal4. Df(3R)red-P52/+ flies preferred temperature colder than normal and AILow was severely lower and AIHigh was higher than wild type (Figure 8A). In contrast, c309>DopR;Df(3R)red-P52/+ flies showed almost normal TPB (Figure 8A). The shifted axis of their distribution profile returned to 24°C, and their AILow and AIHigh showed no difference with the avoidance indices of wild type control w1118 (Figure 8A and Table S7). Consistently, c309>DopR;Df(3R)red-P52/+ flies also showed rescued TPB compared with c309/+;Df(3R)red-P52/+ and UAS-DopR/+;Df(3R)red-P52/+ flies (Figure S1C). Similarly, MB247>DopR;Df(3R)red-P52/+ flies also displayed normal TPB (Figure 8B, Figure S1D, and Table S7). These data suggested that the TPB phenotype of Df(3R)red-P52/+ flies is caused by reduced expression of DopR in MB. These results demonstrated again that DopR regulates TPB in MB.

DopR expression in the αβ lobes of MB is critical in TPB
As described above, both c309 and MB247could restore the TPB of dumb3/dumb3 with UAS-DopR (Figure 7C and 7D). c309 induces gene expression strongly in the αβ and γ lobes, but weakly in the α′β′ lobes (Figure S2A and [49]). MB247 induces gene expression in the αβ and γ lobes but not in the α′β′ lobes (Figure S2B and [49]). To investigate which lobe of MB is relevant to the DopR-mediated regulation of TPB, we used two more MB-specific Gal4 lines. c739>DopR;dumb3/dumb3 showed restored TPB and avoidance index (Figure 9A and Table S8). c739 is expressed in the αβ lobes (Figure S2C and [50]). However, 1471> DopR did not rescue the phenotype of dumb3/dumb3. The flies spread widely over the low and intermediate regions like dumb3/dumb3 and AILow was not restored to the level of wild type control (Figure 9B and Table S8). This 1471 induces gene expression in the γ lobe (Figure S2D and [51]). Taken together, the results suggested that the αβ lobes are relevant to DopR-mediated regulation of TPB and the γ lobe is dispensable.

DopR in the central complex is not involved in TPB
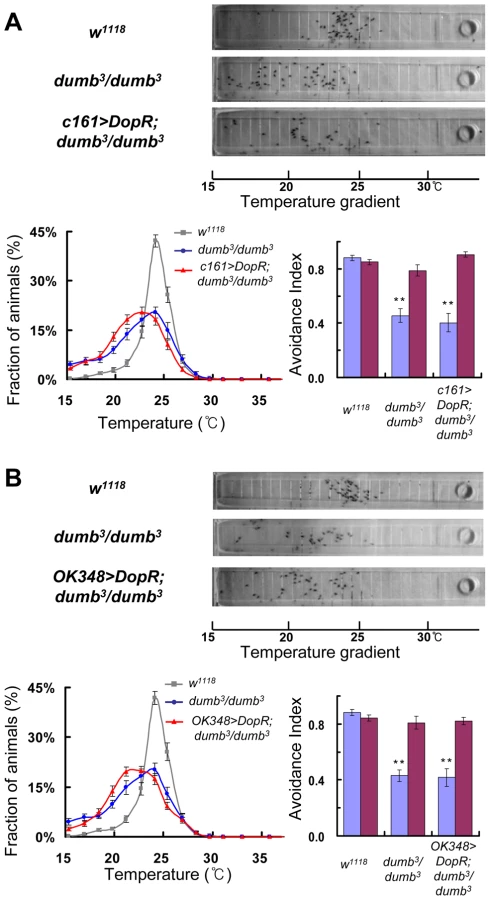
Finally, we examined whether the dopamine signalling in the central complex is required for normal TPB using two more Gal4 lines. c161 is expressed in the ellipsoid body of the central complex (Figure S2E and [18]). The ectopic expression of UAS-DopR by c161 in dumb3/dumb3 did not rescue the defective TPB. c161>DopR;dumb3/dumb3 flies spread over the areas colder than 25°C like dumb3/dumb3 and the lowered AILow of dumb3/dumb3 was not restored (Figure 10A and Table S9). OK348 is expressed in the fan-shaped body (Figure S2F and [52]). OK348>DopR;dumb3/dumb3 flies also showed loss of cold avoidance in the TPB assay and AILow was low like dumb3/dumb3 (Figure 10B and Table S9). These implicated that the ellipsoid body and the fan-shaped body are not relevant to DopR-mediated regulation of TPB.
Discussion
Monoamine neurotransmitters such as dopamine, norepinephrine, and serotonin have been identified as important factors in thermoregulation in mammals [53]–[56]. However, the exact functions of these monoamines in body temperature control are not clearly understood due to limited experimental methods in mammalian systems. Fortunately, our current study clearly revealed that dopamine plays a critical role in regulating TPB of Drosophila. Dopamine-deficient mutants and dopaminergic neuron-inactivated flies lost cold avoidance significantly (Figure 2, Figure 4, and Figure 5). However, the flies overexpressing TH and DDC together in dopaminergic neurons became thermophilic or hypersensitive to cold temperature (Figure 3). This suggests that dopamine levels are critical to regulate TPB; high dopamine levels lead to warm temperature preference and low dopamine levels lead to cool temperature preference.
In this study, we have shown evidence to support that the dopamine signaling in MB regulates TPB. We also demonstrated previously that cAMP signaling in MB modulates TPB; flies with low levels of cAMP prefer cold and the flies with high levels of cAMP prefer warm temperature [1]. In addition, there have been reports that dopamine regulates cAMP levels [39] and that dopamine stimulation of Drosophila brain leads to activation of PKA specifically in the α lobe [40]. Therefore, we propose that when a distinctive subset of dopaminergic neurons which innervate the TPB center in MB is activated, cAMP levels and PKA activity can be increased in the center. Activated PKA may phosphorylate and regulate various targets including ion channels and also induce gene expression of unidentified TPB regulatory genes with CREB binding sites. These PKA-dependent gene expression and posttranslational regulation would change the activities of the TPB center in MB, and fly can determine specific temperature preference or avoidance behaviours. Consistently, our unpublished data strongly suggest that a specific gene group induced by PKA-CREB signalling in the brain is critical for Drosophila TPB.
Previously, it was reported that MB is important for both cold and warm temperature avoidance and preference [1]. Interestingly, all the flies with defective dopamine signalling showed a significant loss in cold avoidance, but some of them also showed loss of warm avoidance (Figure 2, Figure 4, Figure 5, Figure 6). However, the loss of warm avoidance was not severe as MB-defective flies and dTRPA1-deficient flies [1], [16]. We do not think that only dopamine regulates TPB in MB sufficiently. It is possible that other neuromodulators and/or multiple innervated neurons may be involved in modulation of the MB process to regulate TPB. The additive or synergistic effect of dopamine and these modulators may regulate cAMP levels in MB and TPB with broader temperature ranges as shown in MB-defective mutants or cAMP signaling-disrupted flies [1].
It is also possible that dopaminergic neurons are involved in transmitting temperature information to MB. It was reported that TRP and TRPL are required for avoidance of cold temperature in larvae [57], and their cold avoidance phenotypes resemble the TPB of dopamine signalling-deficient flies. Understanding whether the neuronal pathway for the cold avoidance of TRP and TRPL is connected to the dopaminergic pathway of MB will be helpful to understand the regulatory mechanism of TPB. In another report, the flies with surgically removed third antennal segment and aristae showed loss in cold avoidance [16]. Therefore, it is also possible that dopaminergic neurons are involved in transmitting the sensory signals to MB from sensory tissues like the third antennal segment and aristae.
Although three subtypes of dopamine receptors are known in Drosophila, we found that DopR in MB is involved in TPB regulation while DopR in the ellipsoid body and the fan-shaped body is not essential for TPB control. This is consistent with the previous study that these substructures of the central complex are not involved in TPB control [1]. According to the previous results from others and our current data, DopR regulates multiple Drosophila behaviours via distinct neural circuits. DopR in the ellipsoid body is required to promote ethanol-stimulated locomotion and to regulate exogenous arousal negatively, while DopR in PDF-expressing circadian pacemaker cells is needed to regulate endogenous arousal positively [35], [36]. Moreover, DopR in MB is required in learning and memory [32]. In addition, all parts of MB may not work for TPB regulation; DopR in the αβ lobes of MB is relevant but this in the γ lobe is dispensable, which is also consistent with the previous report [1].
In conclusion, our results strongly suggest that dopamine controls TPB and body temperature in Drosophila. We believe that our findings provide important clues to understand the molecular mechanism of Drosophila TPB. Although further studies are required, we cautiously suggest that our fruit fly system can provide a highly useful model to further understand the physiological roles of dopamine in animal body temperature regulation.
Materials and Methods
Fly strains and transgenic lines
Fly stocks were raised on standard cornmeal food at 25°C and 40%–50% relative humidity. Canton-S and w1118 flies all showed a strong temperature preference for 24–25°C regardless of the temperatures at which they were reared in every TPB tests [1], [4]. w1118 was shown as the wild type Drosophila strain in the figures. Fly lines were provided as follows: pale4 (Bloomington, 3279), DdcDE1 (Bloomington, 3168), TH-Gal4 (Serge Birman), UAS-TH, UAS-DDC (Sean B. Carroll), UAS-TNT (Cahir J. O'Kane), UAS-Kir2.1 (Amita Sehgal), UAS-Kir2.1/CyO; tub-Gal80ts (Hiromu Tanimoto), dumb3 (Bloomington, 19491), Df(3R)red-P52 (Bloomington, 3484), ELAV-Gal4 (Bloomington, 458), MB247 (Troy Zars), UAS-DopR2 RNAi (VDRC, 3392), UAS-D2R RNAi (VDRC, 11471), c309, c739, OK348 (Leslie C. Griffith), 1471 (Bloomington, 9465), c161 (Bloomington, 27893). Genomic rescue transgenic fly of pale locus (P{ple+8}) was obtained from Kalpana White [41]. DAT-Gal4 was generated as reported previously [58]. The UAS-DopR construct was generated as follows: a 1.8 Kb fragment including DopR coding sequences was obtained from RT-PCR of fly adult head mRNA with the primers tcgctgaaaagagggaagcaa and cagtaggtagagggctggg; the fragment was cloned into pGEM-T Easy vector (Promega) and sequenced, and the fragment was transferred into NotI and XbaI sites of pUAST vector. We produced the UAS-DopR transgenic flies with standard microinjection methods.
TPB assay
TPB was performed as described previously [1]. Files were incubated under 12 hr∶12 hr dark:light cycle for 4 days before TPB assays. A linear temperature gradient from 15 to 45°C was established along the aluminium block. All TPB assays were done in a room with 35±5% relative humidity. The AILow and AIHigh were calculated as shown in Figure 1B. A mixed population of both sexes was tested for TPB. The number of flies was recorded and processed using Microsoft Excel. The AILow or AIHigh of each tested fly line was compared with the AILow or AIHigh of the control fly lines and then t-tests were performed. *, p<0.05; **, p<0.01.
Drug treatments
Two to three days old flies were raised on drug-containing food for 4 additional days and tested. Melted pre-made standard dextrose media were blended with drugs or the same amount of distilled water, their solvent. As the final drug concentrations, 20 mM α-methyl-p-tyrosine methyl ester (AMPT) (Sigma) and 20 mM 3-hydroxy benzyl hydrazine (HBH) (Sigma) were used. The standard dextrose media contain dextrose 70 g, yeast flake 50 g, cornmeal 35.3 g, agar 5 g, tegosept 7.3 ml and propionic acid 4.7 ml in 1 L total volume.
Conditional induction of TNT and Kir2.1 transgene in dopaminergic neurons
To induce TNT and Kir2.1 expression conditionally in dopaminergic neurons, we crossed UAS-TNT; tub-Gal80ts or UAS-Kir2.1/CyO; tub-Gal80ts flies with TH-Gal4 and DAT-Gal4 flies. These flies were incubated and transferred to new culture bottles periodically at 18°C. Eclosed F1 progenies of TH-Gal4>TNT;tub-Gal80ts/+, TH-Gal4>Kir2.1;tub-Gal80ts/+, or DAT-Gal4>Kir2.1;tub-Gal80ts/+ were collected and aged for 3 days at 18°C before the TPB assay. For TNT or Kir2.1 induction, the flies were transferred from 18°C to 32°C and incubated for 16 hr before the TPB assays. Heterozygous Gal4 flies and UAS-TNT/+;tub-Gal80ts/+ or UAS-Kir2.1/+;tub-Gal80ts/+ files were used as controls.
Immunohistochemistry
To confirm various brain-specific Gal4 fly lines (c309, MB247, c739, 1471, c161 and OK348), they were combined with UAS-LacZ. Adult brains were removed from the head capsules and fixed in 4% paraformaldehyde in phosphate buffered saline (PBS) for 30 min, and rinsed in PBST (PBS containing 0.5% triton X-100). Brains were incubated with anti-β-galactosidase (1∶1000; Promega) and then incubated with the appropriate red fluorescent labelled secondary antibody (Jackson Laboratories). To verify the DopR expression in w1118 and dumb3/dumb3 fly brains, rabbit anti-DopR antibody (1∶1250, From Wolf FW [36]) was used. Confocal analysis was performed on a Zeiss LSM5 microscope.
Supporting Information
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Zdroje
1. HongST
BangS
HyunS
KangJ
JeongK
2008 cAMP signalling in mushroom bodies modulates temperature preference behaviour in Drosophila. Nature 454 771 775
2. BearMF
ConnorsBW
ParadisoMA
2001 Neuroscience: exploring the brain. Lippincott Williams & Wilkins, Baltimore, Md 888
3. ZarsT
2001 Two thermosensors in Drosophila have different behavioral functions. J Comp Physiol A 187 235 242
4. SayeedO
BenzerS
1996 Behavioral genetics of thermosensation and hygrosensation in Drosophila. Proc Natl Acad Sci USA 93 6079 6084
5. HongST
BangS
PaikD
KangJ
HwangS
2006 Histamine and its receptors modulate temperature-preference behaviors in Drosophila. J Neurosci 26 7245 7256
6. PatapoutianA
PeierAM
StoryGM
ViswanathV
2003 ThermoTRP channels and beyond: mechanisms of temperature sensation. Nat Rev Neurosci 4 529 539
7. MoriI
1999 Genetics of chemotaxis and thermotaxis in the nematode Caenorhabditis elegans. Annu Rev Genet 33 399 422
8. ZarsT
2003 Hot and cold in Drosophila larvae. Trends Neurosci 26 575 577
9. TalaveraK
YasumatsuK
VoetsT
DroogmansG
ShigemuraN
2005 Heat activation of TRPM5 underlies thermal sensitivity of sweet taste. Nature 438 1022 1025
10. RosenzweigM
BrennanKM
TaylerTD
PhelpsPO
PatapoutianA
2005 The Drosophila ortholog of vertebrate TRPA1 regulates thermotaxis. Genes Dev 19 419 424
11. BautistaDM
SiemensJ
GlazerJM
TsurudaPR
BasbaumAI
2007 The menthol receptor TRPM8 is the principal detector of environmental cold. Nature 448 204 208
12. ClaphamDE
2007 SnapShot: mammalian TRP channels. Cell 129 220
13. BoulantJA
2000 Role of the preoptic-anterior hypothalamus in thermoregulation and fever. Clin Infect Dis 31 Suppl 5 S157 161
14. TraceyWD
WilsonRI
LaurentG
BenzerS
2003 painless, a Drosophila gene essential for nociception. Cell 113 261 273
15. LeeY
LeeY
LeeJ
BangS
HyunS
2005 Pyrexia is a new thermal transient receptor potential channel endowing tolerance to high temperatures in Drosophila melanogaster. Nat Genet 37 305 310
16. HamadaFN
RosenzweigM
KangK
PulverSR
GhezziA
2008 An internal thermal sensor controlling temperature preference in Drosophila. Nature 454 217 220
17. ChiangAS
LinCY
ChuangCC
ChangHM
HsiehCH
2011 Three-Dimensional Reconstruction of Brain-wide Wiring Networks in Drosophila at Single-Cell Resolution. Curr Biol 21 1 11
18. RennSCP
ArmstrongJD
YangM
WangZ
AnX
1999 Genetic analysis of the Drosophila ellipsoid body neuropil: Organization and development of the central complex. J Neurobiol 41 189 207
19. IzerginaN
BalmerJ
BelloB
ReichertH
2009 Postembryonic development of transit amplifying neuroblast lineages in the Drosophila brain. Neural Dev 4 44
20. LiuG
SeilerH
WenA
ZarsT
ItoK
2006 Distinct memory traces for two visual features in the Drosophila brain. Nature 2006 439 551 556
21. SakaiT
KitamotoT
2006 Differential roles of two major brain structures, mushroom bodies and central complex, for Drosophila male courtship behavior. J Neurobiol 66 821 834
22. JoinerWJ
CrockerA
WhiteBH
SehgalA
2006 Sleep in Drosophila is regulated by adult mushroom bodies. Nature 441 757 760
23. ZhangK
GuoJZ
PengY
XiW
GuoA
2007 Dopamine-mushroom body circuit regulates saliency-based decision-making in Drosophila. Science 316 1901 1904
24. BerryJ
KrauseWC
DavisRL
2008 Olfactory memory traces in Drosophila. Prog Brain Res 169 293 304
25. BudnikV
WhiteK
1987 Genetic dissection of dopamine and serotonin synthesis in the nervous system of Drosophila melanogaster. J Neurogenet 4 309 331
26. NässelDR
ElekesK
1992 Aminergic neurons in the brain of blowflies and Drosophila: dopamine - and tyrosine hydroxylase-immunoreactive neurons and their relationship with putative histaminergic neurons. Cell Tissue Res 267 147 167
27. MaoZ
DavisRL
2009 Eight different types of dopaminergic neurons innervate the Drosophila mushroom body neuropil: anatomical and physiological heterogeneity. Front Neural Circuits 3 5 Epub
28. Claridge-ChangA
RoordaRD
VrontouE
SjulsonL
LiH
2009 Writing memories with light-addressable reinforcement circuitry. Cell 139 405 415
29. KrashesMJ
DasGuptaS
VreedeA
WhiteB
ArmstrongJD
2009 A neural circuit mechanism integrating motivational state with memory expression in Drosophila. Cell 139 416 427
30. DraperI
KurshanPT
McBrideE
JacksonAS
KopinAS
2007 Locomotor activity is regulated by D2-like receptors in Drosophila: an anatomic and functional analysis. Dev Neurobiol 67 378 393
31. KimYC
LeeHG
SeongCS
HanKA
2003 Expression of D1 dopamine receptor dDA1/DmDop1 in the central nervous system of Drosophila melanogaster. Gene Expr Patterns 3 237 245
32. KimYC
LeeHG
HanKA
2007 D1 dopamine receptor dDA1 is required in the mushroom body neurons for aversive and appetitive learning in Drosophila. J Neurosci 27 7640 7647
33. SeugnetL
SuzukiY
VineL
GottschalkL
ShawPJ
2008 D1 receptor activation in the mushroom bodies rescues sleep-loss-induced learning impairments in Drosophila. Curr Biol 18 1110 1117
34. AndreticR
KimYC
JonesFS
HanKA
GreenspanRJ
2008 Drosophila D1 dopamine receptor mediates caffeine-induced arousal. Proc Natl Acad Sci USA 105 20392 20397
35. LebestkyT
ChangJS
DankertH
ZelnikL
KimYC
2009 Two different forms of arousal in Drosophila are oppositely regulated by the dopamine D1 receptor ortholog DopR via distinct neural circuits. Neuron 64 522 536
36. KongEC
WooK
LiH
LebestkyT
MayerN
2010 A pair of dopamine neurons target the D1-like dopamine receptor DopR in the central complex to promote ethanol-stimulated locomotion in Drosophila. PLoS ONE 5 e9954 doi:10.1371/journal.pone.0009954
37. HanKA
MillarNS
GrotewielMS
DavisRL
1996 DAMB, a novel dopamine receptor expressed specifically in Drosophila mushroom bodies. Neuron 16 1127 1135
38. SelchoM
PaulsD
HanKA
StockerRF
ThumAS
2009 The role of dopamine in Drosophila larval classical olfactory conditioning. PLoS ONE 4 e5897 doi:10.1371/journal.pone.0005897
39. MissaleC
NashSR
RobinsonSW
JaberM
CaronMG
1998 Dopamine receptors: from structure to function. Physiol Rev 78 189 225
40. GervasiN
TchénioP
PreatT
2010 PKA dynamics in a Drosophila learning center: coincidence detection by rutabaga adenylyl cyclase and spatial regulation by dunce phosphodiesterase. Neuron 65 516 529
41. NeckameyerWS
WhiteK
1993 Drosophila tyrosine hydroxylase is encoded by the pale locus. J Neurogenet 8 189 199
42. GoodaleDB
MooreKE
1976 A comparison of the effects of decarboxylase inhibitors on L-dopa-induced circling behaviour and the conversion of dopa to dopamine in the brain. Life Sci 19 701 706
43. ThumAS
KnapekS
RisterJ
Dierichs-SchmittE
HeisenbergM
2006 Differential potencies of effector genes in adult Drosophila. J Comp Neurol 498 194 203
44. BainesRA
UhlerJP
ThompsonA
SweeneyST
BateM
2001 Altered electrical properties in Drosophila neurons developing without synaptic transmission. J Neurosci 21 1523 1531
45. McGuireSE
LePT
OsbornAJ
MatsumotoK
DavisRL
2003 Spatiotemporal rescue of memory dysfunction in Drosophila. Science 302 1765 1768
46. RisterJ
HeisenbergM
2006 Distinct functions of neuronal synaptobrevin in developing and mature fly photoreceptors. J Neurobiol 66 1271 1284
47. DietzlG
ChenD
SchnorrerF
SuKC
BarinovaY
2007 A genome-wide transgenic RNAi library for conditional gene inactivation in Drosophila. Nature 448 151 156
48. KitamotoT
2002 Conditional disruption of synaptic transmission induces male-male courtship behavior in Drosophila. Proc Natl Acad Sci USA 99 13232 13237
49. AsoY
GrübelK
BuschS
FriedrichAB
SiwanowiczI
2009 The mushroom body of adult Drosophila characterized by GAL4 drivers. J Neurogenet 23 156 172
50. YangMY
ArmstrongJD
VilinskyI
StrausfeldNJ
KaiserK
1995 Subdivision of the Drosophila mushroom bodies by enhancer-trap expression patterns. Neuron 15 45 54
51. IsabelG
PascualA
PreatT
2004 Exclusive consolidated memory phases in Drosophila. Science 304 1024 1027
52. ConnollyJB
RobertsIJ
ArmstrongJD
KaiserK
ForteM
1996 Associative learning disrupted by impaired Gs signaling in Drosophila mushroom bodies. Science 274 2104 2107
53. SimmondsMA
IversenLL
1969 Thermoregulation: Effects of environmental temperature on turnover of hypothalamic norepinephrine. Science 163 473 474
54. BechmanAL
EisenmanJS
1970 Microeletrophoresis of biogenic amines on hypothalamic thermosensitive cells. Science 170 334 335
55. KerwinBC
LeeTF
1978 Dopamine receptors in the central thermoregulatory pathways of the rat. J Physiol 292 471 483
56. BruckK
HinckelP
1980 Thermoregulatory noradrenergic and serotonergic pathways to hypothalamic units. J Physiol 304 193 202
57. RosenzweigM
KangK
GarrityPA
2008 Distinct TRP channels are required for warm and cool avoidance in Drosophila melanogaster. Proc Natl Acad Sci USA 105 14668 14673
58. LeeY
PaikD
BangS
KangJ
ChunB
2008 Loss of spastic paraplegia gene atlastin induces age-dependent death of dopaminergic neurons in Drosophila. Neurobiol Aging 29 84 94
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2011 Číslo 3
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- Whole-Exome Re-Sequencing in a Family Quartet Identifies Mutations As the Cause of a Novel Skeletal Dysplasia
- Origin-Dependent Inverted-Repeat Amplification: A Replication-Based Model for Generating Palindromic Amplicons
- FUS Transgenic Rats Develop the Phenotypes of Amyotrophic Lateral Sclerosis and Frontotemporal Lobar Degeneration
- Limited dCTP Availability Accounts for Mitochondrial DNA Depletion in Mitochondrial Neurogastrointestinal Encephalomyopathy (MNGIE)
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy