
Genetic Analysis of Tropism Using a Naturally Attenuated Cutaneous Strain
Visceral leishmaniasis is one of the most lethal parasitic diseases, and the mechanisms that govern its survival in visceral organs are not understood. Here, we obtained an atypical cutaneous Leishmania donovani clinical isolate from Sri Lanka and compared it to a typical visceral disease causing clinical isolate. Through whole genome sequencing, bioinformatics analysis, experimental infection in mice and functional genomic analysis, this study provides novel information on what differentiates a deadly visceral strain from a benign cutaneous strain. Results indicate that the ability of Leishmania parasites to cause visceral or cutaneous leishmaniasis may be determined by mutations or amplification of a few genes, or combinations of these factors. Overall, this work contributes to the understanding of parasite virulence and may help guide disease control efforts.
Published in the journal:
. PLoS Pathog 10(7): e32767. doi:10.1371/journal.ppat.1004244
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1004244
Summary
Visceral leishmaniasis is one of the most lethal parasitic diseases, and the mechanisms that govern its survival in visceral organs are not understood. Here, we obtained an atypical cutaneous Leishmania donovani clinical isolate from Sri Lanka and compared it to a typical visceral disease causing clinical isolate. Through whole genome sequencing, bioinformatics analysis, experimental infection in mice and functional genomic analysis, this study provides novel information on what differentiates a deadly visceral strain from a benign cutaneous strain. Results indicate that the ability of Leishmania parasites to cause visceral or cutaneous leishmaniasis may be determined by mutations or amplification of a few genes, or combinations of these factors. Overall, this work contributes to the understanding of parasite virulence and may help guide disease control efforts.
Introduction
Leishmaniasis is a neglected tropical disease present in 98 countries, with over 350 million people at risk of infection and is caused by Leishmania protozoan parasites transmitted by infected sand flies [1], [2]. Visceral leishmaniasis is the most serious form of this disease and it is among the most lethal parasitic infections after malaria. Cutaneous leishmaniasis in comparison causes skin lesions which usually self-heal. Over 20 Leishmania species can infect humans; however only the Leishmania donovani complex including L. infantum cause the vast majority of visceral leishmaniasis cases worldwide [2], [3].
In Sri Lanka, an atypical L. donovani (strain MON-37) has been responsible for thousands of cutaneous leishmaniasis cases in the past decade [4], [5], [6]. This is of considerable interest because L. donovani typically causes visceral leishmaniasis in Asia and Africa. Visceral leishmaniasis is rare in Sri Lanka, with the first recorded case only in 2007 [7] and only 4 cases of autochthonous visceral leishmaniasis reported so far. We have recently demonstrated that the visceral leishmaniasis-causing strain is also L. donovani MON-37 [8]. It is however unknown whether the same or different sub-strains of L. donovani MON-37 are responsible for visceral and cutaneous leishmaniasis in Sri Lanka.
Among the most important questions in Leishmania research is why some species remain at the site of the sandfly bite and cause cutaneous infections and others metastasize to the internal organs where they cause visceral disease. By comparing genomes of Leishmania species which cause different pathology, we previously identified several L. donovani genes including A2 and Ldbpk_280340 which are required for visceral organ tropism [9], [10]. However, comparing genomes of different Leishmania species is insufficient to fully define determinants of disease tropism and pathology because of their evolutionary distance, introducing genetic changes unrelated to human pathology [3]. A more effective strategy is to compare genomes of closely related Leishmania isolates of the same species that cause different human pathologies. We therefore undertook a phenotypic and genotypic analysis of L. donovani clinical isolates derived from cutaneous and visceral leishmaniasis patients in Sri Lanka. Characterization of these L. donovani clinical isolates provides invaluable insight into the etiology of visceral and cutaneous leishmaniasis in Sri Lanka and provides unique insight into the genetic basis of visceral leishmaniasis.
Results
Infection of BALB/c mice with the CL-SL and VL-SL L. donovani strains
The cutaneous leishmaniasis L. donovani isolate (CL-SL) from Sri Lanka was derived from a skin lesion as detailed in methods and the visceral leishmaniasis L. donovani strain (VL-SL) has recently been reported [8]. We initially re-sequenced the 6-phosphogluconate dehydrogenase (6PGDH) isoenzyme gene from the CL-SL and VL-SL isolates to confirm they were both L. donovani MON-37 (Fig. S1).
These isolates were then compared with respect to their ability to cause disease when experimentally introduced into mice. BALB/c mice were injected in the tail vein with CL-SL or VL-SL to compare their ability to cause visceral infections and subcutaneously with CL-SL or VL-SL in the rear footpad to assess cutaneous infection. Liver and spleen parasite burdens were determined 4 weeks after visceral infection, and footpad swelling was monitored for 11 weeks following cutaneous infections. As shown in Fig. 1 A–C, mice injected with the VL-SL isolate had high levels of visceral infection in the liver and spleen. Mice injected with the CL-SL isolate had very little detectable visceral organ infection, demonstrating that the CL-SL parasite has lost the ability to survive in visceral organs (Fig. 1B, C). With respect to cutaneous infections, both the CL-SL and VL-SL isolates demonstrated low virulence; however, the CL-SL isolate was able to induce transient footpad swelling while the VL-SL isolate was unable to do so (Fig. 1D). Although the CL-SL isolate induced more footpad swelling than the VL-SL isolate, this was not associated with a significant increase in parasite number (Fig. S2) and therefore was likely due to a stronger inflammatory response.

Overall, these results conform to Koch's postulates demonstrating that cutaneous and visceral leishmaniasis are caused by distinct but closely related strains of L. donovani. These include 1) isolates were obtained directly from cutaneous or visceral sites of human infection; 2) isolates could be cultured and identified by isoenzyme gene sequencing; 3) isolates induced similar pathologies in experimental animals as in humans; and 4) the same strains could be re-isolated from the experimental infection site. Since the mouse infection phenotypes were similar to the human infections, this would argue that these isolates are representative of the cutaneous and visceral disease causing strains circulating in Sri Lanka.
Genome sequencing
The above experimental infections demonstrated that the CL-SL and VL-SL L. donovani isolates were phenotypically distinct and therefore provided strong justification for comparing their genomes. Whole genome sequencing was performed using the Illumina GAIIx next generation sequencer >200× coverage as detailed in supplementary methods (Text S1). The sequences of the CL-SL and VL-SL genomes were aligned and compared against the L. donovani reference strain BPK282A1 originating from Nepal [11], [12].
There were no gene deletions detected in either genome. However, coverage analysis of gDNA-seq reads in 10 kb segments from the CL-SL and VL-SL isolates indicated that there were nine regions with copy number variations some of which contained several genes (Table 1). Those higher in the VL-SL isolate included Ldbpk_111220.1 (ABC transporter) repeat cluster, LdbpK_161030.1 thru LdbpK_161110.1 hypothetical gene cluster, and LdbpK_200120.1 (phosphoglycerate kinase B, cytosolic and an rRNA locus in chromosome 27. The species-specific A2 multi-gene family coding regions and the flanking 5′ and 3′ A2rel genes [13] were assembled and examined manually because they are incorrectly assembled in the reference genome BPK282A1. Aligning genomic DNA reads against the reconstructed A2 region revealed the presence of more copies of the A2 genes in the VL-SL strain than the CL-SL strain (Table 1). The CL-SL isolate has higher copy number in a region of chromosome 23 which contains ABC thiol transporters (MRPA) H-region (LdbpK_230290.1), terbinafine resistance locus protein (yip1; LdbpK_230280.1) and regions of chromosome 1 (eukaryotic initiation factor 4a, putative), chromosome 19 (glycerol uptake proteins) and chromosome 29 (hypotheticals).

Single nucleotide polymorphism (SNP) and indel analysis
Differences identified between the CL-SL, VL-SL isolates and the reference L. donovani (BPK282A1) genome are summarized in Table 2. More than 80% of the differences are common to VL-SL and CL-SL revealing that the Sri Lanka isolates are more closely related to each other than to the reference L. donovani strain from Nepal. Furthermore, although the majority of variants are homozygous, about 20% of variants are heterozygous. As expected, SNPs accounts for the vast majority of variants which occur mostly in the intergenic regions, while about 20% of the SNPs are located in coding regions. Interestingly, there are over 70 pseudogenes resulting from frame shifts or stop codon variations which are in common to the Sri Lanka CL-SL and VL-SL isolates yet are functional in the reference L. donovani strain (Table 2). These genes therefore appear not to be essential or are not involved in virulence.
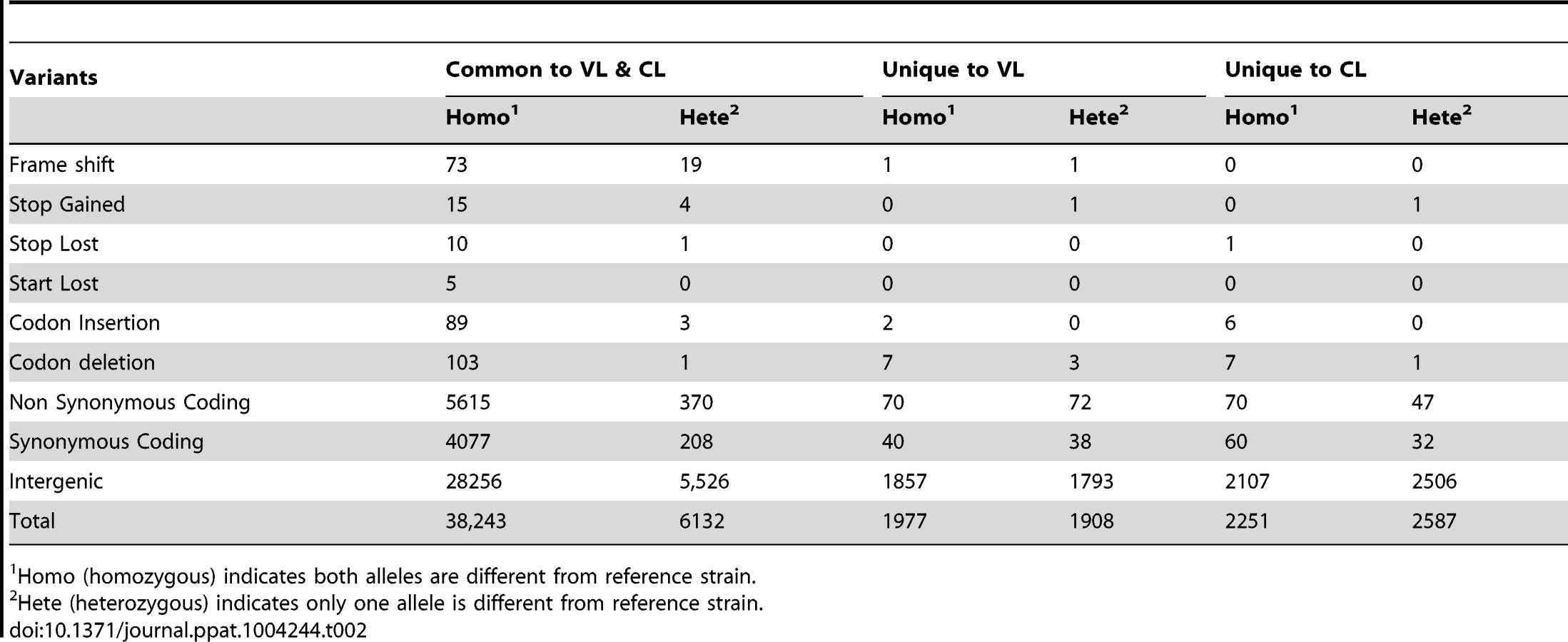
Variations between the VL-SL or CL-SL isolates are among the most interesting, and those specific to the CL-SL isolate would be expected to cause loss of the ability to survive in visceral organs. Surprisingly, as summarized in Table 3, there are only 5 genes unequally affected in the CL-SL and VL-SL isolates by frame shift or altered stop site that could be considered to be pseudogenes. Two genes with frame shifts (LdBPK_312060.1 and LdBPK_320030.1) are only present as homozygous or heterozygous in the VL-SL isolate. There are no homozygous pseudogenes specific to the CL-SL isolate except LdBPK_311390.1 which only differs by 3 amino acids at the C-terminus when compared to its homolog from the VL-SL isolate.

SNPs resulted in 117 (70 homo+47 hetero) non-synonymous and 92 (60 homo+32 hetero) synonymous changes specific to the CL-SL isolate (Tables 2, S1). Although the majority of these non-synonymous SNPs involve hypothetical proteins, 15 of the genes with homozygous mutations in the CL-SL isolate have putative functions (Table S1). Some of the non-synonymous SNPs in these 15 genes are located in functionally important regions of the genes described below. For instance, the non-synonymous change (G226D) in LdBPK_220120.1 phosphoinositide phosphatase is in a highly conserved region. Likewise, the non-synonymous change P147Q in LdBPK_171200.1 is in a highly conserved region of a putative 3-oxo-5-alphasteroid 4-dehydrogenase, responsible for the formation of dihydrotestosterone in higher eukaryotes [14]. LdBPK_252290.1 encodes a putative highly conserved DnaJ family protein that plays a role in protein translation, folding, translocation, and degradation [15]. LdBPK_322160.1 is a putative Rab GTPase family 2 (Rab2) protein involved in membrane trafficking [16]. LdBPK_341910.1 encodes a putative NAD dependent deacetylase which catalyzes NAD+-dependent protein/histone deacetylation and has been shown to regulate gene silencing, DNA repair, metabolic enzymes, and cellular life span [17]. LdBPK_366140.1 encodes a ras-like small GTPase-RagC which may be involved with mTOR regulated translation and cell growth [18]. SNPs in the non-coding region could also have an impact on protein levels since protein expression in Leishmania is regulated at the post-transcriptional level, including via sequences in the mRNA 3′UTR [19].
Functional analysis of SNPs in conserved genes
We considered non-synonymous SNPs in the CL-SL isolate of potential interest if the sequence differed from both the VL-SL isolate and the reference L. donovani strain and if the SNP appeared to be in a functionally important part of the gene as described above. To determine the potential impact of CL-SL specific SNPs on visceral infection and to complement potential functional defects, we cloned 7 corresponding genes from the VL-SL isolate described above, transfected them into the CL-SL isolate and measured the survival of the transgenic parasites in the spleen 4 weeks after infection. All of the CL-SL specific SNPs were verified by Sanger sequencing prior to transfection and their expression in the transfected CL-SL isolate verified by RT-PCR (Fig. 2A). As shown in Fig. 2B, only transfection of the VL-SL Rag C gene into the CL-SL isolate significantly increased infection levels in the spleen. It is noteworthy that the increased infection levels in separate experiments ranged from a 5 to 40 fold and were statistically significant revealing that expression of the VL-SL Rag C gene in CL-SL increased survival in the visceral organs.

The Rag C gene (LdBPK_366140.1) in the CL-SL isolate has a homozygous non-conservative amino acid substitution (R231C). As shown in Fig. 2C, the R231C polymorphism is in a highly conserved amino acid across trypanosomatids. RagC is a small ras-like GTPase involved in the activation of the target of the mammalian rapamycin (mTOR) protein in higher eukaryotes. The mTOR protein is a conserved serine-threonine protein kinase involved in multiple cellular processes including cell stress and proliferation [18]. There are 3 TOR genes (TOR 1–3) identified in the Leishmania genome and TOR 1–2 are essential whereas TOR3 is not essential for survival in the promastigote stage but is required for survival in macrophages and infected mice [20].
Chromosome somy and transcriptional analysis
Chromosome somy can vary in different Leishmania species [11], [12], [21] and therefore it was necessary to determine if there were differences in chromosome somy that could account for the differences in pathology. As shown in Figure 3A, most of the chromosomes were disomic in both isolates except chromosome 31 which was tetrasomic as previously shown for L. donovani [11] and chromosome 23 which appears to be trisomic. With respect to differences between the isolates, chromosomes 13 and 20 appeared to be trisomic in VL-SL and diploid in CL-SL.

We next examined whether the differences in chromosome somy was reflected in differences at the transcript level. cDNA libraries were prepared using mRNA derived from axenic promastigotes (Pro), axenic amastigotes (Ax) and macrophage-derived amastigotes (Am) and entire chromosome transcript levels were compared between these two isolates (Fig. 3B). Several variations between chromosomes were apparent when the median mRNA levels (normalized to the entire genome) were compared; most appear to be unrelated to somy. For example, the mRNA levels are lower than average for many of the small chromosomes (except chromosome 5); chromosome 31 has average mRNA levels despite being tetrasomic; and the mRNA levels for chromosome 13 are high for both isolates even though chromosome 13 is trisomic in the VL-SL isolate. Despite the fact that chromosomes 9 and 26 are disomic in both isolates, the mRNA level for chromosome 9 is about 40% higher in the VL-SL isolate than in the CL-SL isolate; conversely, the mRNA level for chromosome 26 has at least a 30% increase in the CL-SL isolate. However, consistent with the increased somy, the median mRNA level for chromosome 20 has been increased nearly 1.5 times in the VL-SL isolate. Overall these data suggest that chromosome somy may not be a major factor in the different pathologies caused by these isolates since ploidy is not closely mirrored by transcript level.
When comparing the individual gene transcript levels in different life cycle stages, the vast majority of genes were expressed at similar levels. A list of the most differentially expressed genes with at least a 4 fold difference in mRNA levels in the amastigote infected macrophage stage is shown in Table 4. Genes with at least a 2 fold difference are shown in Table S2 for genes more highly expressed in the VL-SL isolate and Table S3 for genes more highly expressed in the CL-SL isolate. Two gene families identified in Table 4, the ABCA3 and A2 gene families, also have higher gene copy numbers as shown in Table 1. One of the rate limiting enzymes in the glycolysis pathway, glycosomal phosphoglycerate kinase (Ldbpk_200110.1) and two hypothetical proteins were also highly up-regulated in the VL-SL strain. The CL-SL isolate had higher mRNA levels of a ribosomal subunit protein and a putative adaptin protein. Overall, these mRNA level differences could impact energy production, protein synthesis and, with respect to A2, visceral organ survival.

Functional analysis of the A2 proteins in the CL-SL and VL-SL L. donovani strains
One of the genes with an increased copy number (Table 1) and higher transcript levels (Table 4) in the VL-SL isolate was the A2 gene family. A2 proteins are stress-inducible factors necessary for visceral infection [9], [22]–[24]. We therefore compared sequence reads of the region of chromosome 22 containing several A2 gene family members (see Text S1), and this revealed an amplification of the A2 coding regions in the VL-SL isolate compared to the CL-SL isolate (Fig. 4A). We also performed Western blot analysis to determine whether the increased number of A2 gene sequences resulted in an increase in A2 protein levels. As shown in Figure 4B, there were higher levels of A2 and more A2 protein species in the VL-SL isolate compared to the CL-SL isolate. This observation would be consistent with previous studies showing that A2 genes with greater number of repeat sequences results in larger A2 proteins and are all inducible with heat stress [22]–[24].

Because of the difference in A2 proteins between the VL-SL and CL-SL isolates, we investigated whether this was functionally relevant for the different pathologies displayed by these strains. A2 expression was therefore experimentally increased in the CL-SL strain by transfection with the A2 gene-containing pKSneo plasmid expression vector. This construct was generated from a L. donovani genomic library and previously used to characterize the role of the A2 locus in visceralization [13]. Expression of the A2 transgene product (shown with arrow) was effectively up-regulated by temperature stress (40°C) similar to the endogenous chromosomal A2 genes (Fig. 4C). As shown in Fig. 4D, the CL+A2 transfected parasite expressing the additional A2 transgene displayed a higher level of survival in the spleen than the control-transfected CL-SL 4 weeks after infection. It is noteworthy that the increased infection levels in separate experiments ranged from a 5 to 50 fold and were statistically significant revealing that expression of the A2 transgene increased survival in the visceral organs.
We further confirmed the importance of A2 in these clinical isolates by downmodulation of A2 expression in the VL-SL strain and this was associated with impaired survival in the visceral organs (Fig. S3). Overall, these results represent a validation of the importance of A2 in a natural setting and highlight the impact that alterations of A2 levels can have on the parasite's ability to survive in visceral organs. Nevertheless, virulence remained severely attenuated in CL-VL parasites in which A2 expression was partially restored (Fig. 4 C, D) relative to the VL-SL isolate (See also Fig. 1B, C), demonstrating that factors in addition to A2 are required for full virulence in the visceral organs.
Discussion
A major question concerning leishmaniasis in Sri Lanka is whether one or different L. donovani strains are responsible for cutaneous and visceral leishmaniasis. We provide compelling evidence here including fulfilment of Koch's postulates that different strains of L. donovani MON-37 are responsible for visceral and cutaneous disease in Sri Lanka. Most strikingly, the CL-SL clinical isolate was severely attenuated for survival in visceral organs in experimentally infected BALB/c mice, yet acquired the ability to cause cutaneous leishmaniasis in humans. Atypical cutaneous leishmaniasis in Sri Lanka caused by L. donovani is therefore most likely due to parasite-specific rather than host-specific determinants.
The L. donovani VL-SL and CL-SL clinical isolates examined here represent unique strains to investigate genetic determinants affecting disease tropism. There were no homozygous gene deletions and only one pseudogene (containing an additional 3 aa at the C-terminal) specific to the CL-SL strain that were wildtype in the VL-SL strain. SNPs and protein level variations are most likely responsible for disease tropism and pathology differences, and the Rag C and A2 genes identified within are examples of genes which may contribute to the different pathologies caused by the CL-SL and VL-SL strains. It will be of considerable interest to characterize additional CL-SL isolates and determine how widespread changes in these and other genes are on a larger population of parasites from cutaneous leishmaniasis patients in Sri Lanka.
Genome sequencing (Table 1, Fig. 4A) and transcription analysis (Table 4) provided evidence that A2 expression levels were higher in the VL-SL isolate relative to the CL-SL isolate and this was confirmed by Western blot (Fig. 4B). A2 is a multigene family present in L. donovani and L. infantum but found as pseudogenes in Old World cutaneous leishmaniasis species such as L. major and L. tropica [22]. Although A2 is present in New World cutaneous L. mexicana [3], [25], its size and sequence differ from Old World A2, and therefore may serve a different function. In L. donovani, A2 is required for survival in visceral organs and protects L. donovani against high temperature (fever) and oxidative stress [23], [24]. As demonstrated within, the expression of a single additional copy of the A2 gene in the CL isolate resulted in increased survival in the spleen. These observations argue that the L. donovani CL-SL strain has lost the threshold level of A2 expression necessary for survival in visceral organs and that this represents a major determinant of its attenuation.
It was of interest to identify a non-synonymous SNP in the CL-SL isolate in the Rag C gene which in higher eukaryotic cells is involved in the regulation of the mTOR pathway. There are 3 TOR genes in Leishmania which are all essential for Leishmania survival in macrophages or infected mice [20]. Although we did not identify SNPs in the TOR genes, this pathway could be compromised by polymorphism in the Rag C gene. Expression of the VL-SL Rag C gene in the CL-SL isolate significantly increased the ability of the CL-SL isolate to survive in visceral organs (Fig. 2) suggesting that this pathway plays an important role for visceral disease. It has also been demonstrated that targeting the TOR pathway with inhibitors represents a novel opportunity to treat trypanosomatid infections [26]. The observations made here further support this pathway as a drug target for visceral leishmaniasis.
Not only is the L. donovani CL-SL strain attenuated for survival in visceral organs, but it has also gained the ability to cause thousands of cutaneous leishmaniasis cases in Sri Lanka. It will be important to identify any gain of function enabling the CL-SL strain to cause cutaneous disease when other naturally-occurring strains of L. donovani are unable to do. This may require a better biological readout for the CL-SL L. donovani isolate and a better understanding of proteins currently classified as hypothetical.
One of the major observations from recent studies sequencing several different species of Leishmania is that although there are relatively few species-specific genes, there is considerably variation in individual gene and chromosome copy numbers [11], [12], [21]. We observed strain-specific differences in the somy of chromosomes 13 and 20 which were trisomic in the VL-SL strain and disomic in the CL-SL strain (Fig. 3A). This lead to an increased level of transcripts from chromosome 20 in the VL-SL isolate relative to the CL-SL isolate though the transcript levels from chromosome 13 are similar in the CL-SL and VL-SL strains (Fig. 3B). This suggests that certain genes encoded on chromosome 20 may play a role in VL-SL pathogenesis. Increased copy number for chromosome 20 would allow for simultaneous amplification and overexpression of these genes. Chromosome 20 encodes 125 hypothetical proteins. Non-hypothetical proteins of interest include glycosomal phosphoglycerate kinase C, one of the five genes with greater than four-fold differences in transcript levels compared to the CL-SL isolate, cytosolic phosphoglycerate kinase B, glycerol-3-phosphate dehydrogenase-like protein, eight calpain-like cysteine peptidases, two DNAj-like protein chaperones, a member of the glutaredoxin antioxidant family, five kinases and three phosphatase subunits. Calpains may play a role in resistance to miltefosine and calpain inhibitors arrest Leishmania growth [27].
Overall, this work provides valuable insight into the pathogenesis of visceral leishmaniasis. In particular, gene deletions or pseudogene formation do not appear to be required for L, donovani to lose the ability to survive in visceral organs and cause cutaneous disease. The ability of Leishmania parasites to cause visceral or cutaneous leishmaniasis may be determined by sequence polymorphisms or amplification of a few genes. This contributes to understanding L. donovani virulence and may help to identify intervention targets required for visceral organ infection by this important parasite.
Materials and Methods
Ethics statement
Isolation of L. donovani from the patients for this study was approved by the ethics review committee of the faculty of Medical Sciences, University of Sri Jayewardenepura, Sri Lanka (approval no 482/09). Written informed consent was obtained prior to parasite isolation from the two adult patients.
All BALB/c mouse infections (permit # 7395) were approved by the McGill University Animal Care Committee following guidelines from the Canadian Council on Animal Care (CCAC).
Sri Lanka L. donovani isolates and culture conditions
L. donovani parasites were isolated from two Sri Lankan patients from the Vavuniya district as previously shown geographically [8]. The VL-SL clinical isolate was from a bone marrow aspirate from a 53-year-old male with chronic fever, hepatosplenomegaly, low haemoglobin and seropositivity for rK39 anti-Leishmania antibodies as previously described [8]. The CL-SL clinical isolate was derived from a skin lesion on the nose from a 28 year old male. Biopsy samples were directly inoculated into Leishmania promastigote culture medium [10], and genomic DNA for genome sequencing was prepared within one month from these promastigote cultures. For axenic amastigote culture, Leishmania promastigotes were shifted to 37°C, pH 5.5 culture media to mimic the macrophage phagolysosome environment. Transfections were performed as previously described with the pKSneo-control plasmid, the pKSneo-A2 plasmid or the pKSneo-A2 (R) antisense plasmid [9]. Transfected parasites were maintained in media supplemented with 200 µg/mL G418 (Wisent).
Infection of BALB/c mice
Female BALB/c mice weighing 17–20 g were purchased from Charles River Breeding Laboratories and maintained in the animal care facility under pathogen-free conditions. BALB/c mice were infected by tail vein injection of stationary-phase promastigotes. Four weeks post infection, amastigotes were isolated from the liver and spleen, and the Leishmania parasite number determined by limiting dilution. The number of stationary promastigotes injected is indicated for each experiment in the Figure legends. Liver parasite burdens were also measured by counting the number of amastigotes in Giemsa-stained liver imprints, expressed as Leishman-Donovan Units (LDU): number of amastigotes per 1000 cell nuclei×liver weight [9]. For cutaneous infections, mice were infected subcutaneously with stationary-phase promastigotes in the hind footpads. Disease progression was monitored by weekly caliper measurement of footpad swelling.
Genomic DNA library preparation, sequencing and analysis
Genomic DNA was sheared into fragments of 100–1,200 bp (Nebulizer, Illumina) and was made into a paired-end DNA sequencing (DNA-seq) library using the Genomic DNA Sample Preparation Kit (Illumina, San Diego, USA). Libraries were sequenced using the Genome Analyzer IIx (Illumina) at the High Throughput Genomics Unit at the University of Washington. Reads were aligned against the reference genome (L. donovani BPK282/Ocl4, cloned line from Nepal). Somy and copy number information for each chromosome were calculated independently using custom written perl script entitled “find_copy_number.pl” (see supplementary methods, Text S1). Single nucleotide polymorphism and small indels were called by inputting alignment files (bam) from all the four libraries into GATK software [28]. A thorough manual inspection revealed around 30% of variant calls were false positives or incorrectly genotyped. Calls were then validated manually and reassigned genotypes if necessary. All the validated variants that were consistent within each group (VL-SL and CL-SL) but different between them were then analyzed in detail to study the effects on protein coding genes using SNP EFF(v3.2) tool [29]. Further details concerning genomic library preparation and sequencing are presented in Supplementary Methods (Text S1).
Total RNA preparation
Promastigote total RNA was extracted from late log phase promastigotes culture. Axenic amastigote total RNA was extracted from axenic amastigote cultures 22 hours after shifting the cells to amastigote culture conditions. B10R macrophages were infected with CL-SL or VL-SL amastigotes at an amastigote/macrophage ratio of 10/1 in suspension in DMEM medium for 12 hours. The percentage of infected macrophages was 72% for CL-SL and 95% for VL-SL; the average amastigotes per infected B10R cell was 3.6 for CL-SL and 3.3 for VL-SL. Infected macrophages were washed and centrifuged to remove extracellular amastigotes and directly suspended in Trizol reagent for total RNA extraction.
Construction and analysis of Spliced Leader libraries
SL RNA-seq libraries were prepared as previously described [30]. The SL RNA libraries were sequenced using the Genome Analyzer IIx (Illumina) at the High Throughput Genomics Unit at the University of Washington to generate 36-nt long single-end reads. Details of the mapping of high-throughput sequencing reads can be found in the supplementary methods (Text S1). The SL RNA-seq data have been submitted to the GEO database under accession no. GSE48475.
PCR, expression vector construction, transfection, RT-PCR, SDS-PAGE and western blotting
PCR cloning of VL-SL genes, expression vector constructions and transfection were performed as previously described [10]. To confirm that the corresponding VL-SL genes were expressed in transfected L. donovani CL-SL cells, the reverse transcription PCR (RT-PCR) reactions were performed with QIAGEN OneStep RT-PCR Kit. One primer was specific for the transfected VL-SL gene and other primer was specific to the expression vector sequence contained in the 3′ untranslated region (See supplementary Table S4 for primer sequences). 1 µg of total RNA extracted from these transfected CL-SL cells and pretreated with DNase I was used in each RT-PCR reaction. SDS-PAGE and Western blotting to detect A2 was performed as previously described [23]. A2 was detected with a 1∶8,000 dilution of mouse monoclonal anti-A2 antibody and tubulin with a 1∶2,000 dilution of mouse monoclonal anti-tubulin antibody (Oncogene).
Data access
All information was deposited in GenBank under bioproject ID PRJNA210295. DNA sequencing data can be accessed from the SRA database using accession no. SRS484822 and SRS484824. RNA-seq data has been deposited in the GEO database using accession no SRP026479. Open source software tools we used are referenced from methods section. The locally developed scripts are publicly available to download from github project ‘bifxscripts’ under GPLicence (https://github.com/bifxcore/bifxscripts/tree/master/Ldonovani_tropism)
Supporting Information
Zdroje
1. AlvarJ, VelezID, BernC, HerreroM, DesjeuxP, et al. (2012) Leishmaniasis Worldwide and Global Estimates of Its Incidence. Plos One 7: e35671.
2. MurrayHW, BermanJD, DaviesCR, SaraviaNG (2005) Advances in leishmaniasis. Lancet 366 : 1561–1577.
3. McCallLI, ZhangWW, MatlashewskiG (2013) Determinants for the development of visceral leishmaniasis disease. PLoS Pathog 9: e1003053.
4. KarunaweeraND, PratlongF, SiriwardaneHVYD, IhalamullaRL, DedetJP (2003) Lankan cutaneous leishmaniasis is caused by Leishmania donovani zymodeme MON-37. Transactions of the Royal Society of Tropical Medicine and Hygiene 97 : 380–381.
5. RanasingheS, WickremasingheR, MunasingheA, HulangamuwaS, SivanantharajahS, et al. (2013) Cross-Sectional Study to Assess Risk Factors for Leishmaniasis in an Endemic Region in Sri Lanka. Am J Trop Med Hyg 89 : 742–749.
6. SiriwardanaHVYD, NoyesHA, BeechingNJ, ChanceML, KarunaweeraND, et al. (2007) Leishmania donovani and cutaneous leishmaniasis, Sri Lanka. Emerging Infectious Diseases 13 : 476–478.
7. AbeygunasekaraPH, CostaYJ, SeneviratneN, RatnatungaN, Wijesundera MdeS (2007) Locally acquired visceral leishmaniasis in Sri Lanka. Ceylon Med J 52 : 30–31.
8. RanasingheS, ZhangWW, WickremasingheR, AbeygunasekeraP, ChandrasekharanV, et al. (2012) Leishmania donovani zymodeme MON-37 isolated from an autochthonous visceral leishmaniasis patient in Sri Lanka. Pathogens and global health 106 : 421–424.
9. ZhangWW, MatlashewskiG (1997) Loss of virulence in Leishmania donovani deficient in an amastigote-specific protein, A2. Proc Natl Acad Sci U S A 94 : 8807–8811.
10. ZhangWW, MatlashewskiG (2010) Screening Leishmania donovani-specific genes required for visceral infection. Mol Microbiol 77 : 505–517.
11. DowningT, ImamuraH, DecuypereS, ClarkTG, CoombsGH, et al. (2011) Whole genome sequencing of multiple Leishmania donovani clinical isolates provides insights into population structure and mechanisms of drug resistance. Genome Research 21 : 2143–2156.
12. RogersMB, HilleyJD, DickensNJ, WilkesJ, BatesPA, et al. (2011) Chromosome and gene copy number variation allow major structural change between species and strains of Leishmania. Genome Research 21 : 2129–2142.
13. ZhangWW, MatlashewskiG (2001) Characterization of the A2-A2rel gene cluster in Leishmania donovani: involvement of A2 in visceralization during infection. Mol Microbiol 39 : 935–948.
14. AzzouniF, MohlerJ (2012) Role of 5alpha-reductase inhibitors in prostate cancer prevention and treatment. Urology 79 : 1197–1205.
15. QiuXB, ShaoYM, MiaoS, WangL (2006) The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. Cellular and molecular life sciences: CMLS 63 : 2560–2570.
16. DhirV, GouldingD, FieldMC (2004) TbRAB1 and TbRAB2 mediate trafficking through the early secretory pathway of Trypanosoma brucei. Molecular and Biochemical Parasitology 137 : 253–265.
17. SauveAA, YounDY (2012) Sirtuins: NAD(+)-dependent deacetylase mechanism and regulation. Current Opinion in Chemical Biology 16 : 535–543.
18. JewellJL, GuanKL (2013) Nutrient signaling to mTOR and cell growth. Trends in Biochemical Sciences 38 : 233–242.
19. ClaytonCE (2002) Life without transcriptional control? From fly to man and back again (vol 21, pg 1881, 2002). Embo Journal 21 : 3917–3917.
20. da SilvaLM, BeverleySM (2010) Expansion of the target of rapamycin (TOR) kinase family and function in Leishmania shows that TOR3 is required for acidocalcisome biogenesis and animal infectivity. Proc Natl Acad Sci U S A 107 : 11965–11970.
21. SterkersY, LachaudL, BourgeoisN, CrobuL, Bastien, PagesM (2012) Novel insights into genome plasticity in eukaryotes: mosaic aneuploidy in Leishmania. Mol Micro 86 : 15–23.
22. ZhangWW, MendezS, GhoshA, MylerP, IvensA, et al. (2003) Comparison of the A2 gene locus in Leishmania donovani and Leishmania major and its control over cutaneous infection. J Biol Chem 278 : 35508–35515.
23. McCallLI, MatlashewskiG (2010) Localization and induction of the A2 virulence factor in Leishmania: evidence that A2 is a stress response protein. Mol Microbiol 77 : 518–530.
24. McCallLI, MatlashewskiG (2012) Involvement of the Leishmania donovani virulence factor A2 in protection against heat and oxidative stress. Exp Parasitol 132 : 109–115.
25. GhedinE, ZhangWW, CharestH, SundarS, KenneyRT, et al. (1997) Antibody response against a Leishmania donovani amastigote-stage-specific protein in patients with visceral leishmaniasis. Clin Diagn Lab Immunol 4 : 530–535.
26. Diaz-GonzalezR, KuhlmannFM, Galan-RodriguezC, Madeira da SilvaL, SaldiviaM, et al. (2011) The susceptibility of trypanosomatid pathogens to PI3/mTOR kinase inhibitors affords a new opportunity for drug repurposing. PLoS Negl Trop Dis 5: e1297.
27. BranquinhaMH, MarinhoFA, SangenitoLS, OliveiraSS, GoncalvesKC, et al. (2013) Calpains: potential targets for alternative chemotherapeutic intervention against human pathogenic trypanosomatids. Current medicinal chemistry 20 : 3174–3185.
28. DePristoMA, BanksE, PoplinR, GarimellaKV, MaguireJR, et al. (2011) A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat Genet 43 : 491.
29. CingolaniP, PlattsA, WangLL, CoonM, NguyenT, et al. (2012) A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w(1118); iso-2; iso-3. Fly 6 : 80–92.
30. ArmourCD, CastleJC, ChenR, BabakT, LoerchP, et al. (2009) Digital transcriptome profiling using selective hexamer priming for cDNA synthesis. Nature methods 6 : 647–649.
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2014 Číslo 7
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- Molecular and Cellular Mechanisms of KSHV Oncogenesis of Kaposi's Sarcoma Associated with HIV/AIDS
- Holobiont–Holobiont Interactions: Redefining Host–Parasite Interactions
- BCKDH: The Missing Link in Apicomplexan Mitochondrial Metabolism Is Required for Full Virulence of and
- Helminth Infections, Type-2 Immune Response, and Metabolic Syndrome
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy