
Comparative Genomics Suggests an Independent Origin of Cytoplasmic Incompatibility in
Terrestrial arthropods are commonly infected with maternally inherited bacterial symbionts that cause cytoplasmic incompatibility (CI). In CI, the outcome of crosses between symbiont-infected males and uninfected females is reproductive failure, increasing the relative fitness of infected females and leading to spread of the symbiont in the host population. CI symbionts have profound impacts on host genetic structure and ecology and may lead to speciation and the rapid evolution of sex determination systems. Cardinium hertigii, a member of the Bacteroidetes and symbiont of the parasitic wasp Encarsia pergandiella, is the only known bacterium other than the Alphaproteobacteria Wolbachia to cause CI. Here we report the genome sequence of Cardinium hertigii cEper1. Comparison with the genomes of CI–inducing Wolbachia pipientis strains wMel, wRi, and wPip provides a unique opportunity to pinpoint shared proteins mediating host cell interaction, including some candidate proteins for CI that have not previously been investigated. The genome of Cardinium lacks all major biosynthetic pathways but harbors a complete biotin biosynthesis pathway, suggesting a potential role for Cardinium in host nutrition. Cardinium lacks known protein secretion systems but encodes a putative phage-derived secretion system distantly related to the antifeeding prophage of the entomopathogen Serratia entomophila. Lastly, while Cardinium and Wolbachia genomes show only a functional overlap of proteins, they show no evidence of laterally transferred elements that would suggest common ancestry of CI in both lineages. Instead, comparative genomics suggests an independent evolution of CI in Cardinium and Wolbachia and provides a novel context for understanding the mechanistic basis of CI.
Published in the journal:
. PLoS Genet 8(10): e32767. doi:10.1371/journal.pgen.1003012
Category:
Research Article
doi:
https://doi.org/10.1371/journal.pgen.1003012
Summary
Terrestrial arthropods are commonly infected with maternally inherited bacterial symbionts that cause cytoplasmic incompatibility (CI). In CI, the outcome of crosses between symbiont-infected males and uninfected females is reproductive failure, increasing the relative fitness of infected females and leading to spread of the symbiont in the host population. CI symbionts have profound impacts on host genetic structure and ecology and may lead to speciation and the rapid evolution of sex determination systems. Cardinium hertigii, a member of the Bacteroidetes and symbiont of the parasitic wasp Encarsia pergandiella, is the only known bacterium other than the Alphaproteobacteria Wolbachia to cause CI. Here we report the genome sequence of Cardinium hertigii cEper1. Comparison with the genomes of CI–inducing Wolbachia pipientis strains wMel, wRi, and wPip provides a unique opportunity to pinpoint shared proteins mediating host cell interaction, including some candidate proteins for CI that have not previously been investigated. The genome of Cardinium lacks all major biosynthetic pathways but harbors a complete biotin biosynthesis pathway, suggesting a potential role for Cardinium in host nutrition. Cardinium lacks known protein secretion systems but encodes a putative phage-derived secretion system distantly related to the antifeeding prophage of the entomopathogen Serratia entomophila. Lastly, while Cardinium and Wolbachia genomes show only a functional overlap of proteins, they show no evidence of laterally transferred elements that would suggest common ancestry of CI in both lineages. Instead, comparative genomics suggests an independent evolution of CI in Cardinium and Wolbachia and provides a novel context for understanding the mechanistic basis of CI.
Introduction
Bacterial symbionts of terrestrial arthropods are common, influential associates, known to affect fundamental aspects of the host life history, ecology, and evolution. These maternally inherited bacteria may, for example, provide essential nutrients supplementing their host's diet, confer protection against natural enemies, increase stress resistance, or influence host plant suitability [1]–[4]. Others have evolved sophisticated means of manipulating the arthropod host's reproduction in ways that cause the symbiont to spread within the host population [5]–[6]. Infection with reproductive manipulators may drive rapid evolution of host sex determination [7], affect genetic population structure, including reproductive isolation and speciation [8], as well as influence the evolution of sexual traits [9]. Reproductive manipulator symbionts may also be powerful tools in pest management for suppression or transformation of pest or vector populations [10]–[11].
The most common symbiont-induced reproductive manipulation, cytoplasmic incompatibility (CI), is also perhaps the most enigmatic. CI occurs, in the simplest case, when a symbiont-infected male host mates with an uninfected female. Affected host embryos die in early development. The symbiont spreads because of the decreased fitness of uninfected relative to infected female hosts [5]. The CI manipulation has been studied most extensively in Wolbachia pipientis, a member of the Alphaproteobacteria established in as many as 40% of terrestrial arthropod species [12] and in filarial nematodes [13]. The verbal model that best describes CI has been termed “modification/rescue” [14], where a factor that is important for the normal development of the insect embryo is modified in sperm cells and can be rescued only if a related strain is present in the eggs. In the fertilized oocyte of an incompatible mating of Drosophila or the parasitic wasp Nasonia vitripennis, CI Wolbachia leads to asynchrony of the timing of maternal and paternal chromosome condensation and segregation during the first embryonic mitotic division, disrupting embryonic development [15]–[16]. However, the molecular basis of CI in this uncultivable microbe remains largely unknown [5].
Genome analysis and expression studies of genes of diverse CI Wolbachia strains have revealed a number of genes with a potential role in CI [17]–[22], but our inability to cultivate these bacteria in a host-free environment, the lack of methods to genetically manipulate Wolbachia, and the absence of an independently evolved CI lineage with which to make comparisons has limited the progress in this area. Here we describe the genome of the only CI-inducing symbiont known that is distantly related to Wolbachia. Cardinium hertigii is a member of the Bacteroidetes, and the strain cEper1 infecting the parasitic wasp Encarsia pergandiella causes CI [23]. The tiny parasitic wasp host (∼18 µg, 1/1000 of the weight of Drosophila spp.) lays eggs in whiteflies, and larval wasps develop at the whiteflies' expense, emerging as adults from the whitefly remains. Related Cardinium strains have also been found in the arthropod groups Hymenoptera, Hemiptera, Diptera, Protura, Acari and Araneae, and an estimated 6–7% of all arthropods are infected with these bacteria [24]–[26]. The most recent analysis also places the nematode symbiont ‘Candidatus Paenicardinium endonii’ within the Cardinium clade, and Cardinium as sister group to the Acanthamoeba endosymbiont Amoebophilus asiaticus [26]–[28]. The Cardinium/Amoebophilus clade is only distantly related to other known insect symbiont lineages within the Bacteroidetes.
The genome sequence of Cardinium hertigii cEper1 reveals a highly reduced genome, both in terms of genome size and metabolic pathways, and a 58 kb cryptic plasmid. Cardinium encodes a set of proteins with the potential to interfere with eukaryotic cell cycle regulation. These proteins, some of which also occur in CI-inducing Wolbachia strains, are good candidates for effectors mediating CI. Despite its metabolically restricted genome, Cardinium encodes a complete biotin biosynthesis pathway, which suggests a potential role of Cardinium in host nutrition. Lastly, several lines of evidence suggest that protists have served as hosts for the progenitor of Cardinium before its adaptation to insects.
Results/Discussion
A highly reduced genome with features of both facultative symbionts and obligate nutritional symbionts of arthropods
The genome of Cardinium hertigii cEper1 consists of a single 887 kb chromosome and a 58 kb plasmid (pCher), with 841 protein coding genes (CDS) (Figure 1, Table 1). It is thus not only smaller than the genomes of free-living bacteria but also reduced compared to the genomes of the CI-inducing Wolbachia strains wMel, wRi, and wPip (1.27–1.48 Mb; [20]–[22]). The size of the Cardinium genome is actually closer in size to the described genomes of obligate (mutualist) symbionts of diverse insect hosts, which are typically highly reduced and range from 140 kb to 790 kb (Table S1) [29]–[30]. Other genomic features of Cardinium such as a low G+C content (36.6%) and a single (unlinked) set of rRNA genes are also common characteristics of intracellular bacterial symbionts. Cardinium differs from obligate symbionts in its abundance of transposable genetic elements (n = 104; 12.4% of all CDSs; Table S2), a feature more typical of facultative symbionts, which generally show a broader host range than obligate symbionts and are not required for host reproduction [29], [31]. In addition, while some obligate insect symbionts harbor small plasmids [32], Cardinium possesses a large cryptic plasmid. pCher contains 65 CDSs, most of which code for transposases and proteins with unknown function (Figure 1, Table 1). Plasmids of similar size have been reported from several rickettsial symbionts infecting arthropods [33]–[35].


The representation of functional categories in the Cardinium genome based on the assignment of CDSs to NCBI clusters of orthologous genes (COGs, [36]) is similar to that of other endosymbionts with small genomes (Figure S1). For example, the gene set required for DNA repair and recombination is similarly reduced as in other facultative symbionts. While several proteins involved in recombination are not encoded (RecBCD, RecF, RecN, RecR), Cardinium has retained RecA, which is missing in most obligate symbionts [32]. The presence of this and other important components suggests that homologous recombination is still possible in Cardinium. The biosynthetic capabilities of Cardinium are very limited, similar to other intracellular insect symbionts and Cardinium's closest sequenced relative, Amoebophilus [37]. Cardinium is not able to synthesize most cofactors or any amino acids or nucleotides de novo. The tricarboxylic acid cycle is missing completely; an F-type ATPase is present but other components of a respiratory chain are lacking. Only the pay-off phase of glycolysis for the generation of ATP and NADH is present (Table S3, Figure 2). To compensate for its reduced metabolic capabilities Cardinium encodes 60 transport proteins (Table S4), facilitating the uptake of oligopeptides and amino acids via an oligopeptide transport system Opp A-F (CAHE_0240-0242, 0244 and 0245), ATP and other nucleotides via nucleotide transport proteins (CAHE_0018, 0158, 0160 and 0789), dicarboxylates via a C4-dicarboxylate transporter DcuAB (CAHE_0645 and 0647), and S-adenosylmethionine via an S-adenosylmethionine transporter (CAHE_0109), among others. Clearly, Cardinium is highly dependent on its intracellular environment and gains most key metabolites and energy in the form of ATP from its eukaryotic host cell.
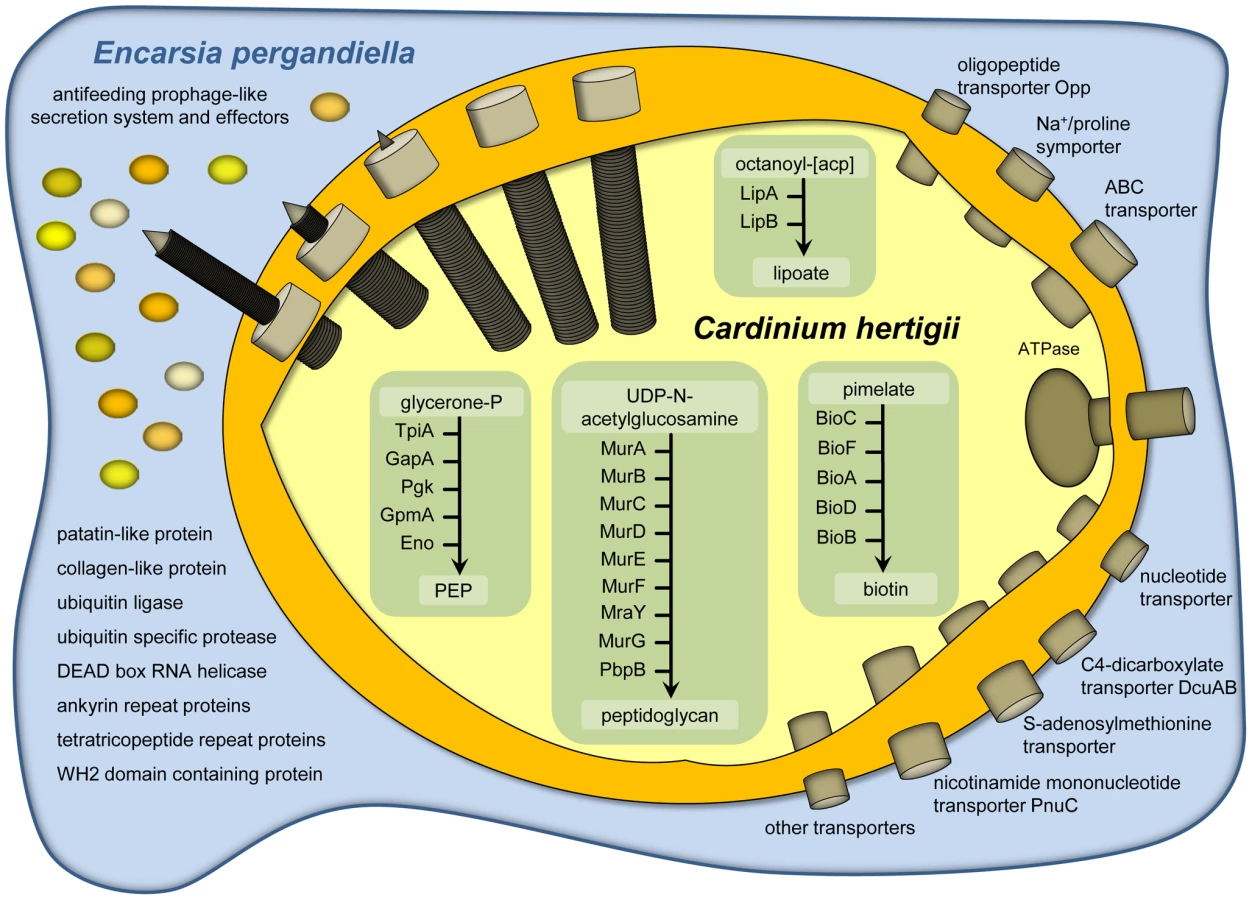
Potential role of retained biosynthetic pathways in host nutrition
Virtually the only complete biosynthetic pathways in the Cardinium genome are those for lipoate and biotin (Figure 2, Table S3). Lipoate is a highly conserved sulfur-containing cofactor involved in oxidative reactions, and also associated with pathogenesis and virulence of microbial pathogens [38]. Biotin is important for carboxylation reactions and cannot be synthesized by many multicelluar eukaryotes, including insects. This B-vitamin is thus an indispensable nutritional factor for insect growth and metamorphosis [39]. Vertebrate blood is deficient in B-vitamins and a complete biotin pathway is also present in the genome of a number of symbionts of blood-feeding hosts including the tsetse fly endosymbiont Wigglesworthia and the tick-associated Ehrlichia, Anaplasma, and Rickettsia species [35], [40]–[41]. It was also experimentally shown that the Wolbachia strain of the bedbug Cimex lectularius supplies various B-vitamins, including biotin, to compensate for the lack of these compounds in their insect host's blood diet [42]. The presence of the biotin pathway in Cardinium cEper1 despite of the lack or truncation of almost all other metabolic pathways is puzzling given the hosts' predaceous larval lifestyle, and that antibiotic curing of Cardinium does not lead to obvious fitness deficits in its host [23]. This does not rule out a possible benefit of supplemental B-vitamin provision that could partially compensate for what appear to be moderately severe fecundity costs (∼15%) to Cardinium infection [43]. It appears reasonably common for facultative, reproductive manipulator symbionts to simultaneously confer host fitness benefits [44]–[45]. On the other hand biotin is also essential for bacteria, and in the absence of alternative sources this pathway might be equally beneficial for Cardinium and its host.
A putative phage-derived protein secretion system
While many obligate symbionts of insects lack dedicated protein secretion systems, several facultative symbionts, including Wolbachia and Rickettsia species, Hamiltonella defensa and Sodalis glossinidius encode protein secretion systems well known from pathogenic microbes [46]–[48]. In Wolbachia, a type four (IV) secretion system is likely involved in mediating CI or other effects on their insect hosts [48]–[49]. No known protein secretion system is present in the genome of Cardinium, but we identified 16 genes arranged in five different genome regions that show highest similarity to antifeeding prophage (AFP)-like genes recently identified in Amoebophilus (amino acid sequence identity between 24% and 76%; E-value< = 1e−10; Figure 3C; Tables S5, S6) [50]. These AFP-like genes are somewhat similar to the putative defective prophage of the entomopathogen Serratia entomophila, which delivers toxins into the hemocytes of its insect host [51]. AFP-like genes are encoded also in other Bacteroidetes [52], with the phage tail sheath protein SCFP from the algicidal bacterium Saprospira sp. being one of the few characterized components. This protein forms characteristic cytoplasmic fibril structures in Saprospira [53]. Interestingly, transmission electron microscopy shows similar subcellular structures in Cardinium (Figure 3A, 3B) [54]–[56], suggesting the presence of an intact protein secretion system encoded by the AFP-like genes. The Cardinium AFP gene cluster lacks putative toxins that are the substrates of the Serratia and Photorhabdus AFPs. Instead, the AFP-like genes of Cardinium may encode a more general secretion system for proteins that are important for manipulation of the insect host cell, taking over the function of the type IV secretion system found in other reproductive manipulators such as Wolbachia. We were able to detect by PCR the three most highly conserved AFP-like genes (CAHE_0458, 0763, 0760) in four other Cardinium strains from three different Encarsia host species (Figure 3C, Table S7), suggesting that AFP-like genes are conserved among Cardinium strains displaying different phenotypes and likely serve an important function. Our hypothesis of a phage-derived protein secretion system in Cardinium parallels the finding that the type six (VI) secretion system shares a common origin with phage tail-associated protein complexes [57]–[58].

Candidate proteins for CI, host cell interaction, and host cell modulation
Typically, bacterial proteins for host cell interaction contain domains that are known to function in the context of a eukaryotic cell [59], including tetratricopeptide repeats (TPR), ankyrin repeats (ANK), leucine-rich repeats, and F - and U-box domains. Several Cardinium proteins contain characteristic TPR and ANK eukaryotic protein-protein interaction motifs (Table S8). In eukaryotic cells TPRs are often associated with multiprotein complexes and play important roles in the functioning of chaperones, transcription and protein transport complexes [60]. Proteins containing TPRs are also involved in the regulation of the eukaryotic cell cycle as components of the anaphase promoting complex (APC), a multi-subunit E3 ubiquitin ligase [61]. Proteins containing TPRs are also present in Amoebophilus and in CI-inducing Wolbachia strains, as well as in the mutualistic nematode-associated Wolbachia strain wBm [62].
ANK proteins play important roles in a variety of cellular processes in eukaryotes such as cell cycle regulation, cytoskeleton regulation, developmental and transcriptional regulation [63]. For example, the ANK protein PLUTONIUM has an important role in the regulation of DNA replication in early Drosophila development [64]. ANK proteins are also known from pathogenic intracellular bacteria such as Legionella pneumophila, Anaplasma phagocytophilum, and Coxiella burnetii, which use type IV secretion systems to translocate these bacterial effectors into their eukaryotic host cells [65]–[66]. Notably, among bacteria, Amoebophilus and CI-inducing Wolbachia strains encode the largest number of ANK proteins (54 ANK proteins in Amoebophilus, 60 in wPip, 35 in wRi, and 23 in wMel), and, while ANK proteins are virtually absent in other sequenced Bacteroidetes genomes and the mutualist Wolbachia strain wBm (five ANK proteins; [62]) Cardinium encodes 19 ANK proteins (14 encoded on the chromosome, five on the plasmid pCher). This overrepresentation of ANK proteins in CI-inducing but only distantly related Cardinium and Wolbachia strains suggests that this class of proteins comprises important mediators of host cell interaction possibly involved in CI. Indeed, it has been frequently suggested earlier that ANK proteins could play a role in Wolbachia CI [22], [67], although the evidence has been equivocal [19], [68].
Cardinium encodes a DEAD box RNA helicase (CAHE_0677). Eukaryotic homologs of this protein promote chromosome segregation in concert with the RNA interference pathway [69]. The DEAD box RNA helicase in Cardinium is conserved among five different Cardinium strains (Table S7), and shows the greatest similarity to Amoebophilus and to intracellular Alphaproteobacteria, including Wolbachia. In addition, the gene encoding this protein is located in a predicted operon with a gene (CAHE_0676) coding for a cold shock DNA-binding protein that is also conserved in CI-inducing Wolbachia strains.
Ubiquitination is a key regulatory process specific to eukaryotes and absent in bacteria. It is thus interesting that Cardinium encodes a protein with a putative RING domain ubiquitin ligase activity (CAHE_p0026; Figure S2) and an ubiquitin specific protease (USP, CAHE_0028; Figure S3). USPs are effector proteins that in bacteria are known in only a few pathogens and symbionts [70]–[71]. The Cardinium USP is conserved among five different strains (Table S7) and belongs to the CA clan of cysteine proteases; the three key domains, the catalytic cysteine box and two histidine boxes, are highly conserved among known and functionally characterized eukaryotic USPs [72]. This high degree of sequence conservation suggests that the Cardinium USP functions in the context of a eukaryotic cell and is able to manipulate the host's ubiquitin system. Ubiquitin proteases are involved in stabilizing/destabilizing proteins, signaling, DNA repair, histone structure, and cell-cycle progression [70], [73]. Among other proteins, eukaryotic USPs interact with cyclin-dependent kinases (CDKs) and with CDK inhibitor proteins (CKI). CDKs are associated with DNA replication initiation in the S-phase, nuclear envelope breakdown, chromosome condensation, assembly of mitotic spindle and changes in microtubule behavior in the M-phase [74]. In CI induced by Wolbachia, delayed nuclear envelope breakdown and histone H3 phosphorylation of mitotic male pronuclei relative to female pronuclei indicates a delayed activity of Cdk1 in the male pronuclei of insect embryos. As a consequence, male pronuclear chromosomes do not segregate properly during mitotic anaphase [5]. Interference of bacterial effectors with CDKs is thus one way in which reproductive incompatibility could be accomplished. If Cardinium used a similar mechanism for induction of CI as Wolbachia, this could be directly achieved via secretion of the Cardinium encoded USP and the counteracting ubiquitin ligase. In Wolbachia strains, which appear to lack USPs, this could be performed through other effectors targeting host USPs, for example ANK proteins [48], [67].
Although orthologs of some of these proteins were also detected in Cardinium strains that cause other phenotypes (Table S7), they are still likely to be good candidates for CI involvement. In addition, Cardinium encodes a number of other more general host interaction proteins. One such protein contains a WH2 motif and a proline-rich domain at the N-terminus (CAHE_0010). These two features are commonly found in actin binding proteins, such as the Sca2 protein in Rickettsia of the spotted fever group, used for bacterial motility within the eukaryotic host cell [75]. Similar proteins are also present in Wolbachia. Other known virulence factors present in Cardinium include a patatin-like phospolipase (CAHE_0286) that is most similar to patatin-like proteins encoded in WO prophages in Wolbachia [76], and a collagen-like protein containing collagen triple helix repeats (CAHE_0706). Collagen is mainly found in multicellular eukaryotes, but is also present in pathogenic bacteria and viruses [77] and has been associated with adhesion and invasion of eukaryotic cells [78].
Evolution from an ancestor in amoebae
Cardinium shares a number of genome characteristics with its closest sequenced relative, the amoeba symbiont Amoebophilus. Sixty-seven percent of all CDSs (n = 561) show similarity with Amoebophilus proteins (at least 25% sequence identity, at least 80% similarity in size). Further, their metabolic pathways are similarly truncated, encode similar transporters for the import of host-derived metabolites, and contain a notably large fraction of transposases or remnants of IS elements compared to other bacteria. The similarity of these genome features between Cardinium and Amoebophilus is striking considering the low degree of 16S rRNA sequence similarity (91%) between these symbionts, indicative of a large evolutionary distance. Consistent with its smaller size (47% relative to Amoebophilus) the Cardinium genome represents a subset of the Amoebophilus genome, with fewer CDSs (841 versus 1557), a greater degree of truncation of metabolic pathways (Figure S1), and the fewer functional transposase genes; 71% (74 out of 104) of the transposase genes are truncated or contain a frame shift compared to 43% in Amoebophilus [79]. Transposable elements are key mediators of genome plasticity; they are able to disrupt genes and to induce rearrangements such as inversions, duplications and deletions. They also play important roles in the shaping of symbiont genomes and in genome size reduction [29], [80]–[81]. The irregular genomic GC skew of Cardinium (Figure 1) is indicative of past activity of transposable elements. Distortion of the compositional strand bias is well known from other bacteria containing large numbers of transposases, including Wolbachia [20]–[22], [82]–[83]. The presence of a large proportion of transposase genes in the genomes of Cardinium and Amoebophilus is also consistent with the low degree of synteny in these relatives, indicating extensive reshuffling during the evolution of these bacteria from their last common ancestor (Figure S4).
The reduction in the capabilities of the Cardinium genome relative to Amoebophilus is also illustrated by cell wall biosynthesis. Both Cardinium and Amoebophilus are able to generate peptidoglycan, but they lack lipopolysaccharide (LPS) and show truncated phospholipid biosynthesis pathways. While Amoebophilus still encodes the complete MreBCD complex, RodA, and IspA considered necessary for a rod-shaped morphology [29], Cardinium lacks all of these genes with the exception of mreB (CAHE_0369) and indeed has a more coccoid appearance compared to Amoebophilus, a pattern also observed in other insect endosymbionts [29]. In general, the Cardinium genome represents a subset of the larger genome of the amoeba symbiont Amoebophilus. The large amount of inactivated transposase genes in the Cardinium genome suggests that it is undergoing further degradation and reduction.
In the Cardinium genome, we identified 68 genes (8% of all CDSs) that were possibly involved in past horizontal gene transfer (HGT) events (Table S9). A prominent example are the genes encoding the biotin synthesis pathway. Phylogenetic analysis suggests that Cardinium has originally lost all genes involved in biotin synthesis, and acquired the complete gene cluster by horizontal gene transfer, putatively from a donor related to rickettsiae (Figure S5). HGT among intracellular bacteria may occur among bacteria infecting the same hosts [84]–[86], and thus document ecological niches inhabited during the organism's evolutionary history. We used phylogenetic analysis to determine the putative HGT partners (donors or recipients) and infer additional possible hosts of the bacterial lineage leading to Cardinium. As expected, Cardinium contains a number of HGT-affected genes shared with partners generally found in arthropod hosts (38% of all HGT affected genes, Figures S6, S7; Table S9). In addition, there are many genes shared with a diverse assemblage of bacteria, and a few eukaryotic genes. Notably, 14% of the HGT-affected genes of Cardinium are shared with bacteria known to be associated with amoebae, e.g. Simkania negevensis and Legionella drancourtii, and 24% are shared with bacteria that have been reported to infect both amoebae and arthropods. The most likely explanation for the presence of genes from amoeba-associated bacteria is that prior to the adaptation to its arthropod host, Cardinium (or its ancestor) lived as a symbiont of amoebae or other protists, in which HGT with other amoeba-associated bacteria was facilitated. This notion is consistent with our observation that the Cardinium genome represents a subset of the genome of the sister lineage to Cardinium, the amoeba symbiont Amoebophilus. It is thus likely that the common ancestor of Cardinium and Amoebophilus lived as a symbiont of an amoeba or a protist. These unicellular eukaryotes are known to have contributed to the development of key features for survival in eukaryotic host cells by other intracellular bacteria [84], [87]–[88].
Independent origin of CI
At some point during its evolutionary history, Cardinium made the transition from amoebae to insect hosts and became a reproductive manipulator able to induce CI to facilitate its spread in host populations. Although Cardinium and Wolbachia share this phenotype, it is unknown whether the molecular mechanisms leading to CI are identical. If they were, and if the ability to cause CI originated in either one of the two groups and subsequently was acquired by the other through HGT during coinfection of the same host [89]–[91], one would expect to observe a set of genes in Cardinium and CI-inducing Wolbachia that likely mediate this phenotype and share a common evolutionary origin. Among the orthologous genes shared by Cardinium and Wolbachia there is not a single obvious case of a gene encoding a candidate effector involved in CI. Apart from the patatin-like phospolipase, which is considered a more general virulence factor, we identified only one orthologous gene (CAHE_0604) that was exclusive to Cardinium and some rickettsiae including the CI-inducing Wolbachia strains. This gene encodes a predicted integral membrane protein without any known functional domains and is thus unlikely to mediate CI. This suggests that there is no common evolutionary origin of CI in Cardinium and Wolbachia, and that the molecular mechanism of CI is either different in these two groups, or convergent.
It is striking, however, that comparison of the genomes of CI-inducing Wolbachia strains with the CI lineage of Cardinium revealed in both genomes a large number of proteins that contain eukaryotic domains and likely mediate host cell interaction and CI. These include a DEAD box RNA helicase, and many ANK and TPR proteins that are highly unusual in bacterial genomes and good candidates for CI effectors manipulating the eukaryotic cell cycle. Most of these proteins are highly divergent and show no sequence similarity beyond the presence of eukaryotic domains. This indicates an independent origin of genes involved in CI, most likely through independent HGT events and acquisition of host genes. This notion is further supported by the presence of ubiquitin modifying proteins in Cardinium, which might be involved in CI, and the absence of these in CI-inducing Wolbachia strains. Taken together, CI seems to be based on the exploitation of eukaryotic domains for host cell manipulation, and there is strong evidence for an independent emergence of the molecular mechanisms underlying CI in these two groups. In general, the Cardinium genome points to the utility of a comparative context for analysis of reproductive manipulation in symbiotic bacteria that are refractory to direct genetic manipulation, a fertile area for research in the coming years.
Materials and Methods
Nomenclature of Cardinium strains
No strain nomenclature has previously been adopted for Cardinium hertigii. In an effort to create a convenient and consistent system, strains have been named in this study following the strain nomenclature of Wolbachia pipientis [5]. Thus the genome reference strain is “cEper1”, where “c” refers to Cardinium, “Eper” refers to the host Encarsia pergandiella, and “1” simply denotes the first named strain from this host.
Rearing of Encarsia pergandiella wasps harboring Cardinium
Cardinium hertigii cEper1 is a symbiont of the minute parasitoid wasp Encarsia pergandiella (∼18 µg) that attacks whiteflies [23]. Wasps were originally collected from the whitefly Bemisia tabaci near Weslaco, Texas in October 2006, and kept in culture on B. tabaci on cowpea. Males of E. pergandiella develop as hyperparasites and were reared on another whitefly primary parasitoid, Eretmocerus eremicus. Prior to purification of Cardinium cells, wasps were reared on B. tabaci that were not infected with Rickettsia spp.
Purification of Cardinium cells and DNA isolation
For Cardinium purification, wasps were reared on dozens of whitefly-infested plants. Approximately 8,000 adult wasps were collected from emergence jars. The Cardinium purification protocol was modified from [92]. Wasps were surface-sterilized with 2.6% sodium hypochlorite and 0.5% SDS for 1 min, washed with sterile water, and homogenized by hand in buffer A (250 mM EDTA, 35 mM Tris-HCl, 250 mM sucrose, 25 mM KCl, 10 mM MgCl2) using a Dounce tissue grinder (Wheaton). The homogenate was transferred to a 1.5 ml centrifuge tube with an additional 1 ml of buffer A. Cellular debris was pelleted for 5 min at 80 g, 4°C. The supernatant was centrifuged for 5 min at 4000 g, 4°C. The resulting pellet was carefully resuspended in 1 ml of buffer A, then vortexed for 3 sec. Following a 5 min centrifugation at 300 g, the supernatant was loaded onto a 13 mm diameter filter cassette holder (Swinnnex filter holder, Millipore) containing a 0.8 to 8 µm pore size glass fiber prefilter (Millipore) and a strong protein binding 5 µm pore-size mixed cellulose ester membrane (Millipore). The supernatant was slowly pushed through the filter with a syringe. The filter cassette holder was washed with buffer A (without EDTA) until 1.5 ml of filtrate was obtained. The filtrate was centrifuged for 5 min at 5000 g, 4°C. Following resuspension of the pellet in buffer A (without EDTA), 10 units of DNase 1 (Roche) were added to the cell suspension and incubated for 30 min at 4°C to remove insect host DNA. The reaction was stopped with 100 µl 0.5 M EDTA. The tube was spun down for 5 min at 4100 g, 4°C, the pellet washed with 1 ml buffer A, then spun down again. The cell pellet was resuspended in 250 µl of TE buffer.
The purified Cardinium cells were mixed with 675 µl of DNA extraction buffer (100 mM Tris/HCl, 100 mM EDTA, 100 mM sodium phosphate, 1.5 M NaCl, 1% cetyltrimethylammonium bromide (CTAB) (w/v), 200 µg/ml proteinase K, pH 8.0; [93]), 10 µl of 20 mg/ml proteinase K (Roche) was added and the tube was incubated for 30 min at 37°C. Then 75 µl of 20% SDS was added, the tube was shaken and incubated at 65°C for 1 h, with gentle inversions every 15 to 20 min. Following the incubation, 1 ml of chloroform/isoamyalcohol (24∶1 v/v) was mixed in. The aqueous phase was recovered following centrifugation. Nucleic acids were precipitated by adding 0.6 volumes of isopropanol, holding at room temperature for 1 h, then centrifuging at 16,000 g for 20 min, 4°C. The pellet was washed with cold 70% ethanol and spun down for 5 min at max speed, 4°C. Ethanol was removed and the pellet allowed to air dry. The DNA pellet was resuspended in TE buffer with 7 units RNase/ml (RNaseA, Qiagen), and incubated for 20 min at 37°C.
Whole-genome amplification
The extracted Cardinium DNA was quantified using PicoGreen (Invitrogen), totaling approximately 2 ng, which was insufficient for library generation and sequencing, thus requiring amplification. To minimize bias, multiple displacement amplification (MDA) was performed on eight replicate reactions as follows. Approximately 0.1 ng of template DNA was denatured using alkaline solution and amplified using the Repli-g UltraFast Mini Kit (Qiagen) according to the manufacturer's instructions. MDA was performed overnight and the eight resulting MDA products were pooled prior to library generation and sequencing.
Sequencing, assembly, and gap closure
A combination of Illumina and 454 shotgun sequencing was performed on the pooled symbiont MDA DNA product. Two differing 454 standard libraries (one un-normalized, one normalized) were generated and sequenced totaling 300,490,911 bp. In addition, we generated and sequenced two 454 paired end libraries totaling 106,933,881 bp. An Illumina GAii shotgun library was constructed and sequenced (run mode 2×76 bp) generating 1,371,155,520 bp. All general aspects of library construction and sequencing can be found at http://www.jgi.doe.gov/. The Illumina GAii sequencing data was assembled with Velvet (http://genome.cshlp.org/content/18/5/821.short) with a hash length of 61 and with the following parameters -exp_cov 130 -cov_cutoff 1 -min_contig_lgth 100. The consensus sequences were shredded into 1.5 Kbp overlapped fake reads and assembled together with the 454 data. The velvet contig fake reads (17,983 reads, 9.2 Mbp) and the 454 pyrosequencing reads (400.3 Mbp) were assembled using the Newbler assembler version 2.4 (Roche) using the parameters –ace -g –mi 98 -ml 80 –rip. The Newbler assembly consisted of 20,306 contigs in 1,154 scaffolds. Illumina reads were additionally used to correct potential base errors and increase consensus quality using the software SeqMan NGen from DNASTAR. One scaffold consisting of 78 contigs was identified as the Cardinium chromosome based on BLAST searches against the ribosomal rRNA database Silva (release_102); another scaffold (6 contigs) was identified representing the Cardinium plasmid based on BLAST searches against the non-redundant sequence dataset (nr) at GenBank/EMBL/DBBJ. The gaps in both scaffolds were closed by manual refinement of the assembly and by PCR and Sanger sequencing in house and by LGC Genomics (Berlin, Germany).
Genome annotation and analysis
The genome was analyzed and automatically annotated using the Microbial Genome Analysis and Annotation Platform MaGe [94]. The automatic annotation was further refined by blastp against Swiss-Prot and UniProt using an E-value of 1e−5, a minimum amino acid identity of 30%, and minimum alignment overlaps of 40% as threshold values, and by manual annotation of selected genes. The circular view of the genome (Figure 1) was generated using the software GenVision (DNASTAR); the GC skew was calculated using the program CGView [95] with a sliding window size of 887 bp. Transposable genetic elements were identified using blastp. Data for NCBI clusters of orthologous genes (COGs, [36]) analysis were taken from the MaGe [94]. Biochemical pathway reconstruction was performed using KEGG [96] integrated in MaGe [94]. Classification of transport proteins into Transport Classification Database (TCDB) families was done using BLAST (http://www.tcdb.org/index.php) [97]. The antifeeding prophage (AFP)-like cluster was first identified by using blastp with proteins encoded on the AFP-like gene cluster of Amoebophilus and then by using either blastp or psi-blast with proteins of the AFP from Serratia. Putative host cell interaction proteins were further analyzed using blastp; protein domains were predicted using PFAM [98] and SMART [99]. Multiple amino acid sequence alignments were done using MAFFT [100]. Putative horizontal gene transfer candidate proteins were predicted by blastp of all Cardinium proteins against the non-redundant protein GenBank/EMBL/DDBJ sequence database (nr). Cardinium proteins with ten best blast hits to proteins from organisms outside the bacterial phylum Bacteroidetes were considered to potentially beinvolved in a past horizontal gene transfer (HGT) events. To further investigate this, the top 50 blast hits were used for amino acid sequence alignments with MUSCLE [101], and phylogenetic trees were reconstructed using the software MEGA5 [102]. Trees were calculated using the neighbor joining algorithm (2000 bootstrap resamplings) and the maximum likelihood algorithm (100 bootstrap resamplings). The nearest neighbor of putatively HGT affected genes of Cardinium was identified by the lowest number of internal nodes in the calculated trees. If there were several neighbors with the same number of nodes, the minimum sum of branch lengths was used as criterion. The sequences described in this paper have been deposited at GenBank/EMBL/DDBJ under accession numbers HE983995 (chromosome) and HE983996 (plasmid pCher). All contigs from the original Encarsia metagenome from which the Cardinium genome was reconstructed are also available at GenBank/EMBL/DDBJ.
PCR screening for putative host cell interaction genes in different Cardinium strains
Approximately 100 wasps from five Encarsia spp. cultures harboring different Cardinium hertigii strains, including the reference strain cEper1 were each collected in a 1.5 ml reaction tube, resuspended in 180 µl buffer ATL (QIAGEN DNeasy blood and tissue kit) and homogenized with a pellet pestle suitable for 1.5 ml microcentrifuge tubes. DNA from homogenized wasps was isolated with QIAGEN DNeasy blood and tissue kit as recommended in the manufacturer's protocol with the exception of the usage of 400 µg proteinase K (Roche) resuspended in 20 µl ddH2O instead of the proteinase K recommended by the manufacturer. A standard PCR cycling program with 35 cycles with primers specific for different Cardinium genes was used for the amplification (for primer sequences and annealing temperatures see Table S10). PCR included New England Biolabs Taq DNA Polymerase at a concentration of 0.8 units/20 µl reaction with ThermoPol Buffer. dNTPs were used at a final concentration of 1 mM. Primers were used at a concentration of 0.4 µM; BSA was added at 0.6 µg/µl.
Transmission electron microscopy of Cardinium cells
Transmission electron microscopy of Cardinium cells was performed as described elsewhere [55]. Ovaries of adult E. pergandiella wasps were fixed in 4% glutaraldehyde in 0.05 M cacodylate buffer overnight at 4°C. After postfixation in 2% OsO4 for 2 h, the samples were washed, en bloc-stained in 2% uranyl acetate, and dehydrated through an ethanol series (50, 70, 95, and 100%). The samples were then placed in propylene oxide and embedded in Epon. Serial sections were cut with an RMC MT7000 ultra microtome. The grids were stained with saturated uranyl acetate and lead citrate and viewed under a Philips Electronic Instruments CM12 transmission electron microscope.
Supporting Information
Zdroje
1. MoranNA, McCutcheonJP, NakabachiA (2008) Genomics and evolution of heritable bacterial symbionts. Annu Rev Genet 42 : 165–190.
2. HedgesLM, BrownlieJC, O'NeillSL, JohnsonKN (2008) Wolbachia and virus protection in insects. Science 322 : 702.
3. OliverKM, RussellJA, MoranNA, HunterMS (2003) Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc Natl Acad Sci U S A 100 : 1803–1807.
4. NakabachiA, YamashitaA, TohH, IshikawaH, DunbarHE, et al. (2006) The 160-kilobase genome of the bacterial endosymbiont Carsonella. Science 314 : 267.
5. WerrenJH, BaldoL, ClarkME (2008) Wolbachia: master manipulators of invertebrate biology. Nat Rev Microbiol 6 : 741–751.
6. SerbusLR, Casper-LindleyC, LandmannF, SullivanW (2008) The genetics and cell biology of Wolbachia-host interactions. Annu Rev Genet 42 : 683–707.
7. CordauxR, BouchonD, GreveP (2011) The impact of endosymbionts on the evolution of host sex-determination mechanisms. Trends Genet 27 : 332–341.
8. TelschowA, FlorM, KobayashiY, HammersteinP, WerrenJH (2007) Wolbachia-induced unidirectional cytoplasmic incompatibility and speciation: mainland-island model. PLoS ONE 2: e701 doi:10.1371/journal.pone.0000701..
9. JigginsFM, HurstGDD, MajerusMEN (2000) Sex-ratio-distorting Wolbachia causes sex-role reversal in its butterfly host. Proceedings of the Royal Society B-Biological Sciences 267 : 69–73.
10. ZabalouS, RieglerM, TheodorakopoulouM, StaufferC, SavakisC, et al. (2004) Wolbachia-induced cytoplasmic incompatibility as a means for insect pest population control. Proc Natl Acad Sci U S A 101 : 15042–15045.
11. WalkerT, JohnsonPH, MoreiraLA, Iturbe-OrmaetxeI, FrentiuFD, et al. (2011) The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 476 : 450–453.
12. ZugR, HammersteinP (2012) Still a host of hosts for wolbachia: analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS ONE 7: e38544 doi:10.1371/journal.pone.0038544..
13. TaylorMJ, HoeraufA (1999) Wolbachia bacteria of filarial nematodes. Parasitol Today 15 : 437–442.
14. WerrenJH (1997) Biology of Wolbachia. Annu Rev Entomol 42 : 587–609.
15. LassyCW, KarrTL (1996) Cytological analysis of fertilization and early embryonic development in incompatible crosses of Drosophila simulans. Mech Dev 57 : 47–58.
16. TramU, SullivanW (2002) Role of delayed nuclear envelope breakdown and mitosis in Wolbachia-induced cytoplasmic incompatibility. Science 296 : 1124–1126.
17. GavotteL, HenriH, StouthamerR, CharifD, CharlatS, et al. (2007) A Survey of the bacteriophage WO in the endosymbiotic bacteria Wolbachia. Mol Biol Evol 24 : 427–435.
18. SinkinsSP, WalkerT, LyndAR, StevenAR, MakepeaceBL, et al. (2005) Wolbachia variability and host effects on crossing type in Culex mosquitoes. Nature 436 : 257–260.
19. PapafotiouG, OehlerS, SavakisC, BourtzisK (2011) Regulation of Wolbachia ankyrin domain encoding genes in Drosophila gonads. Res Microbiol 162 : 764–772.
20. KlassonL, WalkerT, SebaihiaM, SandersMJ, QuailMA, et al. (2008) Genome evolution of Wolbachia strain wPip from the Culex pipiens group. Mol Biol Evol 25 : 1877–1887.
21. KlassonL, WestbergJ, SapountzisP, NaslundK, LutnaesY, et al. (2009) The mosaic genome structure of the Wolbachia wRi strain infecting Drosophila simulans. Proc Natl Acad Sci U S A 106 : 5725–5730.
22. WuM, SunLV, VamathevanJ, RieglerM, DeboyR, et al. (2004) Phylogenomics of the reproductive parasite Wolbachia pipientis wMel: a streamlined genome overrun by mobile genetic elements. PLoS Biol 2: e69 doi:10.1371/journal.pbio.0020069..
23. HunterMS, PerlmanSJ, KellySE (2003) A bacterial symbiont in the Bacteroidetes induces cytoplasmic incompatibility in the parasitoid wasp Encarsia pergandiella. Proc Biol Sci 270 : 2185–2190.
24. DuronO, HurstGD, HornettEA, JoslingJA, EngelstadterJ (2008) High incidence of the maternally inherited bacterium Cardinium in spiders. Mol Ecol 17 : 1427–1437.
25. DallaiR, MercatiD, GiustiF, GottardoM, CarapelliA (2011) A Cardinium-like symbiont in the proturan Acerella muscorum (Hexapoda). Tissue Cell 43 : 151–156.
26. NakamuraY, KawaiS, YukuhiroF, ItoS, GotohT, et al. (2009) Prevalence of Cardinium bacteria in planthoppers and spider mites and taxonomic revision of “Candidatus Cardinium hertigii” based on detection of a new Cardinium group from biting midges. Applied and Environmental Microbiology 75 : 6757–6763.
27. NoelGR, AtibalentjaN (2006) ‘Candidatus Paenicardinium endonii’, an endosymbiont of the plant-parasitic nematode Heterodera glycines (Nemata: Tylenchida), affiliated to the phylum Bacteroidetes. Int J Syst Evol Microbiol 56 : 1697–1702.
28. HornM, HarzenetterMD, LinnerT, SchmidEN, MullerKD, et al. (2001) Members of the Cytophaga-Flavobacterium-Bacteroides phylum as intracellular bacteria of acanthamoebae: proposal of ‘Candidatus Amoebophilus asiaticus’. Environ Microbiol 3 : 440–449.
29. McCutcheonJP, MoranNA (2011) Extreme genome reduction in symbiotic bacteria. Nat Rev Microbiol 10 : 13–26.
30. MoyaA, PeretoJ, GilR, LatorreA (2008) Learning how to live together: genomic insights into prokaryote-animal symbioses. Nat Rev Genet 9 : 218–229.
31. NewtonIL, BordensteinSR (2011) Correlations between bacterial ecology and mobile DNA. Curr Microbiol 62 : 198–208.
32. WuD, DaughertySC, Van AkenSE, PaiGH, WatkinsKL, et al. (2006) Metabolic complementarity and genomics of the dual bacterial symbiosis of sharpshooters. PLoS Biol 4: e188 doi:10.1371/journal.pbio.0040188..
33. BaldridgeGD, BurkhardtNY, FelsheimRF, KurttiTJ, MunderlohUG (2008) Plasmids of the pRM/pRF family occur in diverse Rickettsia species. Appl Environ Microbiol 74 : 645–652.
34. OgataH, RenestoP, AudicS, RobertC, BlancG, et al. (2005) The genome sequence of Rickettsia felis identifies the first putative conjugative plasmid in an obligate intracellular parasite. PLoS Biol 3: e248 doi:10.1371/journal.pbio.0030248..
35. GillespieJJ, JoardarV, WilliamsKP, DriscollT, HostetlerJB, et al. (2011) A Rickettsia genome overrun by mobile genetic elements provides insight into the acquisition of genes characteristic of obligate intracellular lifestyle. J Bacteriol 194 : 376–394.
36. TatusovRL, GalperinMY, NataleDA, KooninEV (2000) The COG database: a tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res 28 : 33–36.
37. Schmitz-EsserS, TischlerP, ArnoldR, MontanaroJ, WagnerM, et al. (2010) The genome of the amoeba symbiont “Candidatus Amoebophilus asiaticus” reveals common mechanisms for host cell interaction among amoeba-associated bacteria. J Bacteriol 192 : 1045–1057.
38. SpaldingMD, PriggeST (2010) Lipoic acid metabolism in microbial pathogens. Microbiology and Molecular Biology Reviews 74 : 200–228.
39. LipkeH, FraenkelG (1956) Insect nutrition. Annual Review of Entomology 1 : 17–44.
40. AkmanL, YamashitaA, WatanabeH, OshimaK, ShibaT, et al. (2002) Genome sequence of the endocellular obligate symbiont of tsetse flies, Wigglesworthia glossinidia. Nat Genet 32 : 402–407.
41. Dunning HotoppJC, LinM, MadupuR, CrabtreeJ, AngiuoliSV, et al. (2006) Comparative genomics of emerging human ehrlichiosis agents. PLoS Genet 2: e21 doi:10.1371/journal.pgen.0020021..
42. HosokawaT, KogaR, KikuchiY, MengXY, FukatsuT (2010) Wolbachia as a bacteriocyte-associated nutritional mutualist. Proc Natl Acad Sci U S A 107 : 769–774.
43. PerlmanSJ, KellySE, HunterMS (2008) Population biology of cytoplasmic incompatibility: maintenance and spread of Cardinium symbionts in a parasitic wasp. Genetics 178 : 1003–1011.
44. BrownlieJC, CassBN, RieglerM, WitsenburgJJ, Iturbe-OrmaetxeI, et al. (2009) Evidence for metabolic provisioning by a common invertebrate endosymbiont, Wolbachia pipientis, during periods of nutritional stress. PLoS Pathog 5: e1000368 doi:10.1371/journal.ppat.1000368..
45. HimlerAG, Adachi-HagimoriT, BergenJE, KozuchA, KellySE, et al. (2011) Rapid Spread of a Bacterial Symbiont in an Invasive Whitefly Is Driven by Fitness Benefits and Female Bias. Science 332 : 254–256.
46. PrestonGM (2007) Metropolitan Microbes: Type III Secretion in Multihost Symbionts. Cell Host Microbe 2 : 291–294.
47. CoombesBK (2009) Type III secretion systems in symbiotic adaptation of pathogenic and non-pathogenic bacteria. Trends in Microbiology 17 : 89–94.
48. DaleC, MoranNA (2006) Molecular interactions between bacterial symbionts and their hosts. Cell 126 : 453–465.
49. RancèsE, VoroninD, Tran-VanV, MavinguiP (2008) Genetic and functional characterization of the type IV secretion system in Wolbachia. J Bacteriol 190 : 5020–5030.
50. PenzT, HornM, Schmitz-EsserS (2011) The genome of the amoeba symbiont “Candidatus Amoebophilus asiaticus” encodes an afp-like prophage possibly used for protein secretion. Virulence 1 : 541–545.
51. HurstMR, BeardSS, JacksonTA, JonesSM (2007) Isolation and characterization of the Serratia entomophila antifeeding prophage. FEMS Microbiol Lett 270 : 42–48.
52. PerssonOP, PinhassiJ, RiemannL, MarklundBI, RhenM, et al. (2009) High abundance of virulence gene homologues in marine bacteria. Environ Microbiol 11 : 1348–1357.
53. FurusawaG, YoshikawaT, TakanoY, MiseK, FurusawaI, et al. (2005) Characterization of cytoplasmic fibril structures found in gliding cells of Saprospira sp. Can J Microbiol 51 : 875–880.
54. BigliardiE, SacchiL, GenchiM, AlmaA, PajoroM, et al. (2006) Ultrastructure of a novel Cardinium sp. symbiont in Scaphoideus titanus (Hemiptera: Cicadellidae). Tissue Cell 38 : 257–261.
55. Zchori-FeinE, GottliebY, KellySE, BrownJK, WilsonJM, et al. (2001) A newly discovered bacterium associated with parthenogenesis and a change in host selection behavior in parasitoid wasps. Proc Natl Acad Sci U S A 98 : 12555–12560.
56. Zchori-FeinE, PerlmanSJ, KellySE, KatzirN, HunterMS (2004) Characterization of a ‘Bacteroidetes’ symbiont in Encarsia wasps (Hymenoptera: Aphelinidae): proposal of ‘Candidatus Cardinium hertigii’. Int J Syst Evol Microbiol 54 : 961–968.
57. BonemannG, PietrosiukA, MogkA (2010) Tubules and donuts: a type VI secretion story. Mol Microbiol 76 : 815–821.
58. LeimanPG, BaslerM, RamagopalUA, BonannoJB, SauderJM, et al. (2009) Type VI secretion apparatus and phage tail-associated protein complexes share a common evolutionary origin. Proc Natl Acad Sci U S A 106 : 4154–4159.
59. JehlMA, ArnoldR, RatteiT (2011) Effective - a database of predicted secreted bacterial proteins. Nucleic Acids Res 39: D591–595.
60. BlatchGL, LassleM (1999) The tetratricopeptide repeat: a structural motif mediating protein-protein interactions. Bioessays 21 : 932–939.
61. SchreiberA, StengelF, ZhangZ, EnchevRI, KongEH, et al. (2011) Structural basis for the subunit assembly of the anaphase-promoting complex. Nature 470 : 227–232.
62. FosterJ, GanatraM, KamalI, WareJ, MakarovaK, et al. (2005) The Wolbachia genome of Brugia malayi: endosymbiont evolution within a human pathogenic nematode. PLoS Biol 3: e121 doi:10.1371/journal.pbio.0030121..
63. LiJ, MahajanA, TsaiMD (2006) Ankyrin repeat: a unique motif mediating protein-protein interactions. Biochemistry 45 : 15168–15178.
64. AxtonJM, ShamanskiFL, YoungLM, HendersonDS, BoydJB, et al. (1994) The inhibitor of DNA replication encoded by the Drosophila gene plutonium is a small, ankyrin repeat protein. EMBO J 13 : 462–470.
65. PanX, LuhrmannA, SatohA, Laskowski-ArceMA, RoyCR (2008) Ankyrin repeat proteins comprise a diverse family of bacterial type IV effectors. Science 320 : 1651–1654.
66. VothDE (2011) ThANKs for the repeat: Intracellular pathogens exploit a common eukaryotic domain. Cell Logist 1 : 128–132.
67. Iturbe-OrmaetxeI, BurkeGR, RieglerM, O'NeillSL (2005) Distribution, expression, and motif variability of ankyrin domain genes in Wolbachia pipientis. J Bacteriol 187 : 5136–5145.
68. DuronO, BoureuxA, EchaubardP, BerthomieuA, BerticatC, et al. (2007) Variability and expression of ankyrin domain genes in Wolbachia variants infecting the mosquito Culex pipiens. J Bacteriol 189 : 4442–4448.
69. PekJW, KaiT (2011) DEAD-box RNA helicase Belle/DDX3 and the RNA interference pathway promote mitotic chromosome segregation. Proc Natl Acad Sci U S A 108 : 12007–12012.
70. RytkonenA, HoldenDW (2007) Bacterial interference of ubiquitination and deubiquitination. Cell Host Microbe 1 : 13–22.
71. WilkesTE, DarbyAC, ChoiJH, ColbourneJK, WerrenJH, et al. (2010) The draft genome sequence of Arsenophonus nasoniae, son-killer bacterium of Nasonia vitripennis, reveals genes associated with virulence and symbiosis. Insect Mol Biol 19 Suppl 1 : 59–73.
72. HuM, LiP, LiM, LiW, YaoT, et al. (2002) Crystal structure of a UBP-family deubiquitinating enzyme in isolation and in complex with Ubiquitin Aldehyde. Cell 111 : 1041–1054.
73. NijmanSM, Luna-VargasMP, VeldsA, BrummelkampTR, DiracAM, et al. (2005) A genomic and functional inventory of deubiquitinating enzymes. Cell 123 : 773–786.
74. StifflerLA, JiJY, TrautmannS, TrustyC, SchubigerG (1999) Cyclin A and B functions in the early Drosophila embryo. Development 126 : 5505–5513.
75. HaglundCM, ChoeJE, SkauCT, KovarDR, WelchMD (2010) Rickettsia Sca2 is a bacterial formin-like mediator of actin-based motility. Nat Cell Biol 12 : 1057–1063.
76. KentBN, FunkhouserLJ, SetiaS, BordensteinSR (2011) Evolutionary genomics of a temperate bacteriophage in an obligate intracellular bacteria (Wolbachia). PLoS ONE 6: e24984 doi:10.1371/journal.pone.0024984..
77. RasmussenM, JacobssonM, BjorckL (2003) Genome-based identification and analysis of collagen-related structural motifs in bacterial and viral proteins. J Biol Chem 278 : 32313–32316.
78. PatersonGK, NieminenL, JefferiesJM, MitchellTJ (2008) PclA, a pneumococcal collagen-like protein with selected strain distribution, contributes to adherence and invasion of host cells. FEMS Microbiol Lett 285 : 170–176.
79. Schmitz-EsserS, PenzT, SpangA, HornM (2011) A bacterial genome in transition - an exceptional enrichment of IS elements but lack of evidence for recent transposition in the symbiont Amoebophilus asiaticus. BMC Evol Biol 11 : 270.
80. SiguierP, FileeJ, ChandlerM (2006) Insertion sequences in prokaryotic genomes. Curr Opin Microbiol 9 : 526–531.
81. TouchonM, RochaEP (2007) Causes of insertion sequences abundance in prokaryotic genomes. Mol Biol Evol 24 : 969–981.
82. NakayamaK, YamashitaA, KurokawaK, MorimotoT, OgawaM, et al. (2008) The Whole-genome sequencing of the obligate intracellular bacterium Orientia tsutsugamushi revealed massive gene amplification during reductive genome evolution. DNA Res 15 : 185–199.
83. ChoNH, KimHR, LeeJH, KimSY, KimJ, et al. (2007) The Orientia tsutsugamushi genome reveals massive proliferation of conjugative type IV secretion system and host-cell interaction genes. Proc Natl Acad Sci U S A 104 : 7981–7986.
84. BordensteinSR, ReznikoffWS (2005) Mobile DNA in obligate intracellular bacteria. Nat Rev Microbiol 3 : 688–699.
85. GeorgiadesK, MerhejV, El KarkouriK, RaoultD, PontarottiP (2011) Gene gain and loss events in Rickettsia and Orientia species. Biol Direct 6 : 6.
86. GimenezG, BertelliC, MolinerC, RobertC, RaoultD, et al. (2011) Insight into cross-talk between intra-amoebal pathogens. BMC Genomics 12.
87. MolmeretM, HornM, WagnerM, SanticM, Abu KwaikY (2005) Amoebae as training grounds for intracellular bacterial pathogens. Appl Environ Microbiol 71 : 20–28.
88. ToftC, AnderssonSG (2010) Evolutionary microbial genomics: insights into bacterial host adaptation. Nat Rev Genet 11 : 465–475.
89. WhiteJA, KellySE, CockburnSN, PerlmanSJ, HunterMS (2011) Endosymbiont costs and benefits in a parasitoid infected with both Wolbachia and Cardinium. Heredity 106 : 585–591.
90. SkaljacM, ZanicK, BanSG, KontsedalovS, GhanimM (2010) Co-infection and localization of secondary symbionts in two whitefly species. BMC Microbiol 10 : 142.
91. SirvioA, PamiloP (2010) Multiple endosymbionts in populations of the ant Formica cinerea. BMC Evol Biol 10 : 335.
92. BraigHR, ZhouWG, DobsonSL, O'NeillSL (1998) Cloning and characterization of a gene encoding the major surface protein of the bacterial endosymbiont Wolbachia pipientis. J Bacteriol 180 : 2373–2378.
93. ZhouJ, BrunsMA, TiedjeJM (1996) DNA recovery from soils of diverse composition. Appl Environ Microbiol 62 : 316–322.
94. VallenetD, LabarreL, RouyZ, BarbeV, BocsS, et al. (2006) MaGe: a microbial genome annotation system supported by synteny results. Nucleic Acids Res 34 : 53–65.
95. StothardP, WishartDS (2005) Circular genome visualization and exploration using CGView. Bioinformatics 21 : 537–539.
96. KanehisaM, ArakiM, GotoS, HattoriM, HirakawaM, et al. (2008) KEGG for linking genomes to life and the environment. Nucleic Acids Res 36: D480–484.
97. SaierMHJr, YenMR, NotoK, TamangDG, ElkanC (2009) The transporter classification database: recent advances. Nucleic Acids Res 37: D274–278.
98. FinnRD, TateJ, MistryJ, CoggillPC, SammutSJ, et al. (2008) The Pfam protein families database. Nucleic Acids Res 36: D281–288.
99. LetunicI, DoerksT, BorkP (2009) SMART 6: recent updates and new developments. Nucleic Acids Res 37: D229–232.
100. KatohK, TohH (2008) Recent developments in the MAFFT multiple sequence alignment program. Brief Bioinform 9 : 286–298.
101. EdgarRC (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 32 : 1792–1797.
102. KumarS, NeiM, DudleyJ, TamuraK (2008) MEGA: a biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief Bioinform 9 : 299–306.
103. BaslerM, PilhoferM, HendersonGP, JensenGJ, MekalanosJJ (2012) Type VI secretion requires a dynamic contractile phage tail-like structure. Nature 483 : 182–186.
Štítky
Genetika Reprodukční medicínaČlánek vyšel v časopise
PLOS Genetics
2012 Číslo 10
- Kazuistika – Perspektivy využití precizované medicíny v rámci personalizované specifické terapie onkologických pacientů
- Nobelova cena za chemii pro genetické nůžky: Objev, který změní naši budoucnost?
- Technologie na bázi RNA v klinické praxi: od přebarvených petúnií k terapii vzácných a dosud jen obtížně léčitelných chorob u lidí
- „Nepředstavovali jsme si, že náš výzkum povede přímo ke vzniku nových léků, dokonce ještě za našeho života“
- Bezplatné služby pro diagnostiku ATTRv amyloidózy pro kardiology
Nejčtenější v tomto čísle
- A Mutation in the Gene Causes Alternative Splicing Defects and Deafness in the Bronx Waltzer Mouse
- Mutations in (Hhat) Perturb Hedgehog Signaling, Resulting in Severe Acrania-Holoprosencephaly-Agnathia Craniofacial Defects
- Classical Genetics Meets Next-Generation Sequencing: Uncovering a Genome-Wide Recombination Map in
- Regulation of ATG4B Stability by RNF5 Limits Basal Levels of Autophagy and Influences Susceptibility to Bacterial Infection
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy