
EPIGENETICKÉ ZMENY V PATOGENÉZE MALÍGNEHO MELANÓMU UVEY A MOŽNOSTI ICH TERAPEUTICKÉHO OVPLYVNENIA. PRIEHLAĎ
Autoři:
B. Smolková; L. Demková
Působiště autorů:
Ústav experimentálnej onkológie, Biomedicínske centrum Slovenskej akadémie vied, Bratislava
Vyšlo v časopise:
Čes. a slov. Oftal., 76, 2020, No. 2, p. 55-60
Kategorie:
Přehledový článek
doi:
https://doi.org/10.31348/2020/12
Souhrn
Uveálny melanóm (UM) je závažné onkologické ochorenie, ktoré vedie ku vzniku metastatickej choroby u viac ako 50 % pacientov. Napriek zlepšeniu lokálnej liečby, stále neexistuje efektívna terapia, ktorá by zabránila rozvoju metastáz. Preto si toto ochorenie vyžaduje intenzívny výskum, zameraný na identifikáciu nových liečebných stratégií.
V predklinických modeloch UM bolo dokázané, že epigenetické liečivá zvyšujú senzitivitu rezistentných nádorových buniek na liečbu. Úspešné použitie inhibítorov histón deacetyláz, ktoré u nádorových buniek indukovali zastavenie bunkového cyklu, reprogramovanie konzistentné s melanocytovou diferenciáciou a inhibíciu rastu nádorov v predklinických modeloch, dokazuje úlohu epigenetickej regulácie v metastázovaní UM. Hlbšie pochopenie úlohy epigenetických zmien v patogenéze UM by mohlo prispieť k objaveniu účinnejších liečiv, ktoré v kombinácii s tradičnými prístupmi môžu priniesť lepšie terapeutické výsledky pre pacientov s vysokým rizikom progresie ochorenia.
Klíčová slova:
malígny melanóm uvey – epigenetické zmeny – metylácia DNA – modifikácie histónov – nekódujúce RNA
ÚVOD
Malígny melanóm uvey (UM) je najčastejšie sa vyskytujúci vnútroočný nádor u dospelých. Jeho incidencia na Slovensku je 5,1 na 1 milión obyvateľov, čo znamená 32 až 54 novo-diagnostikovaných pacientov ročne [1]. Tumory najčastejšie vznikajú z melanocytov lokalizovaných v uveálnej vrstve oka, primárne v cievovke (choroidea; 85 %), ale môžu pochádzať aj z vráskovca (corpus ciliare; 5-8 %) alebo z dúhovky (iris; 3-5 %) [2].
Liečba primárneho UM sa v súčasnosti odkláňa od radikálnych postupov. Dominuje liečba ionizujúcim žiarením a kombinované prístupy, možná je liečba protónmi aj fotónmi. V našich podmienkach sa využíva brachyterapia episklerálnymi žiaričmi, napr. Ru106, liečba gama nožom alebo stereotaktická rádiochirurgia na lineárnom urýchľovači [3-5]. Prerastanie melanómu choroidey do očnice vedie k radikálnemu výkonu, nie iba enukleácii očnej gule, ale aj k exenterácii očnice [6]. Pri sledovaní prežívania pacientov po enukleácii a po liečbe ionizujúcim žiarením sa nezistili významné rozdiely [7,8]. Napriek nízkemu výskytu lokálnych rekurencií v súčasnosti nie je k dispozícii žiadna liečba, ktorá by zvyšovala celkové prežívanie metastatických pacientov. Rovnako neexistuje vysvetlenie, týkajúce sa primárnej rezistencie UM voči akejkoľvek systémovej liečbe. V štádiu klinického skúšania sú inhibítory tyrozínkinázy alebo Met (receptorový proteín rastového faktora hepatocytov), inhibítory históndeacetyláz (HDAC) alebo imunoterapia (NCT02068586, NCT02223819, NCT02068586, NCT01585194) [9].
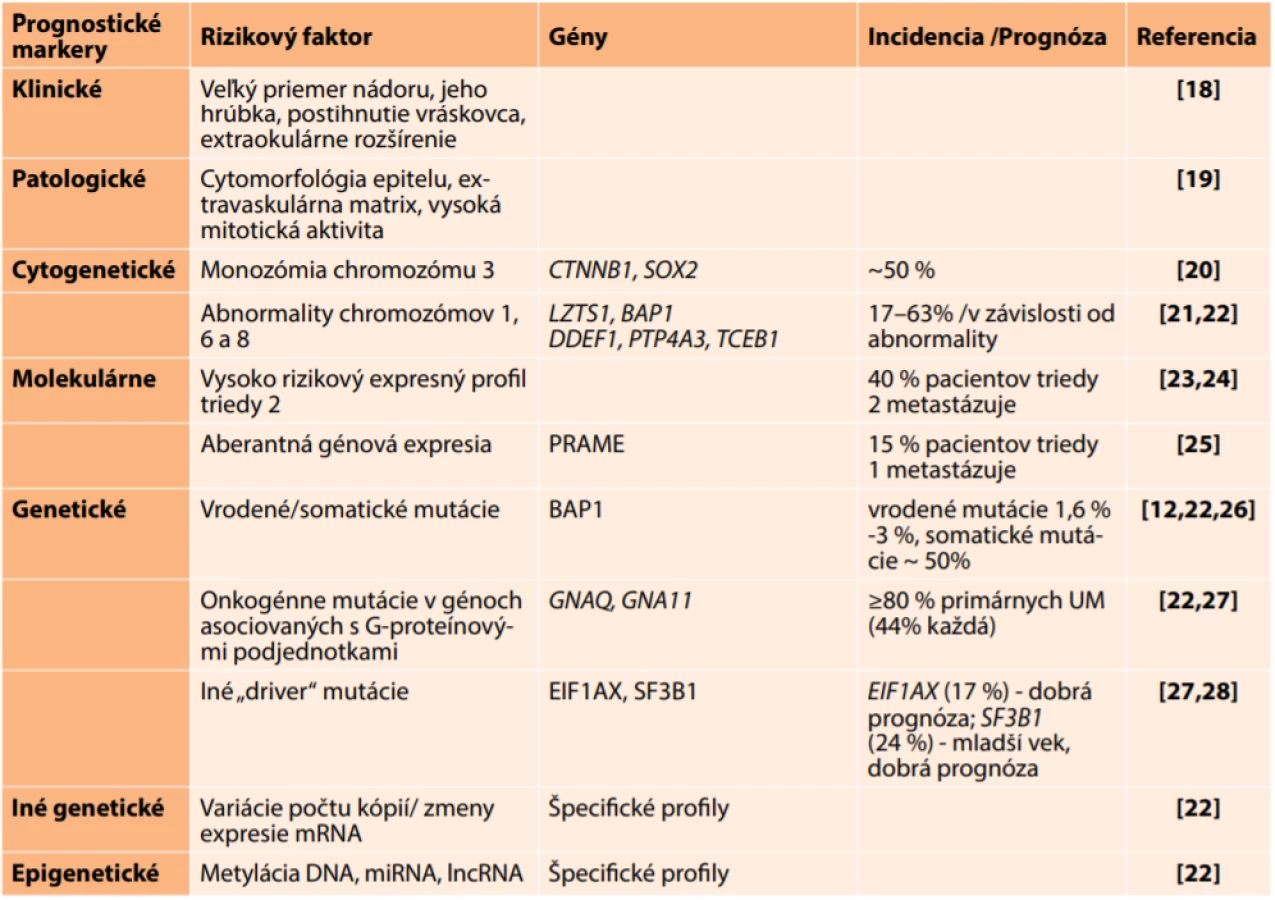
UM metastázuje až u 90 % prípadov do pečene. Mechanizmus, ktorý je základom pečeňového tropizmu, zatiaľ nie je známy. Metastatická choroba, ktorá vzniká u takmer 50 % pacientov, je asociovaná so zlou prognózou. Nedávno publikovaná metaanalýza, uskutočnená v súbore 912 pacientov s metastatickým UM, uvádza medián priemerného prežívania bez progresie 3,3 mesiaca a celkového prežívania 10,2 mesiaca [10]. Riziko vzniku metastáz je asociované s veľkosťou nádoru, extraokulárnym prerastaním, vysokou mitotickou aktivitou a epiteloidným typom nádorových buniek (Tabuľka 1). K najvýznamnejším molekulárnym markerom metastatického rizika patria monozómia chromozómu 3 a špecifický profil expresie mRNA v nádorovom tkanive. Expresný profil panelu 15-tich génov, komerčne dostupný pod názvom DecisionDx-UM, umožňuje rozdelenie pacientov na základe metastatického rizika do 3 skupín: Trieda 1A - veľmi nízke riziko s 2% pravdepodobnosťou metastatického šírenia; Trieda 1B - nízke riziko s 21% pravdepodobnosťou vzniku metastáz; Trieda 2 - vysoké riziko so 72% pravdepodobnosťou metastázovania počas nasledujúcich piatich rokov [11]. Kľúčovú úlohu v metastázovaní zohrávajú somatické mutácie génu BAP1 (BRCA1 - Associated Protein 1), vedúce k zníženiu alebo úplnej strate expresie Bap1 proteínu [12]. BAP1, lokalizovaný v oblasti 3p21, je tumor-supresorový gén, ktorý sa podieľa na epigenetickej regulácii expresie génov, významných počas vývoja a diferenciácie. Jeho inaktivácia vedie ku vzniku buniek, ktoré majú vlastnosti nádorových kmeňových buniek. Takýto fenotyp je asociovaný s agresívnym správaním nádorov a zlou prognózou [13]. S metastatickým rizikom sú spojené aj abnormality chromozómov 1, 6, 8 a 9, spolu so špecifickými mutáciami v ďalších génoch ako sú SF3B1 (Splicing Factor 3B Subunit 1A) alebo EIF1AX (eukaryotic translation initiation factor 1A, X-linked) [14]. Mutácie BRAF (B-Raf Proto-Oncogene, Serine/Threonine Kinase) a NRAS (NRAS Proto-Oncogene, GTPase), ktoré sú typické pre kožný melanóm (40 %–50 % a 15 %–20 %), sa u UM nevyskytujú [15,16].
Epigenetické mechanizmy
Epigenetické mechanizmy ako sú metylácia DNA, modifikácie histónov a nekódujúce RNA sú esenciálne pre normálny vývin a homeostázu organizmu. Navzájom interagujú a majú kľúčovú úlohu pri udržiavaní tkanivovo-špecifickej expresie génov a ochrane pred genetickou instabilitou. Ich narušenie môže viesť k zmenám vo funkcii génov, malígnej transformácii a môže mať dopad na jednotlivé signálne dráhy podieľajúce sa na metastázovaní. Úlohu epigenetickej deregulácie v patogenéze UM potvrdili výsledky niekoľkých štúdií. Jednou z najvýznamnejších je integrovaná multiplatformová analýza 80 UM, ktorá ukázala, že okrem známych genetických zmien sú so zlou prognózou pacientov asociované aj zmeny v metylácii DNA a expresii viacerých mikroRNA (miRNA) a dlhých nekódujúcich RNA (lncRNA) [22]. U nádorov s vysokým rizikom metastázovania je de-regulovaná aj expresia génov, kódujúcich samotné epigenetické regulačné enzýmy [29]. Epigenetická inaktivácia expresie môže mať zvlášť významnú úlohu predovšetkým v génoch lokalizovaných na chromozómoch s typickými abnormalitami v počte kópií ako sú chromozómy 1, 3, 6 alebo 8. Na chromozóme 3, monozómia ktorého je prítomná približne u 50 % pacientov s UM, je umiestnených niekoľko tumor-supresorových génov a génov, ktoré majú kľúčovú úlohu v hematogénnej diseminácii. Sú tu umiestnené napríklad gény RASSF1A (RAS association domain family 1), FHIT (Fragile Histidine Triad), BAP1 (BRCA1 - Associated Protein 1), CTNNB1 (Catenin Beta 1) alebo SOX2 (Sex-determining region Y (SRY)-Box2).
Metylácia DNA
Metylácia DNA patrí medzi najlepšie preštudované epigenetické mechanizmy. Ide o kovalentné pripojenie metylovej skupiny (-CH3) k bázam DNA, predovšetkým cytozínovému zvyšku v dinukleotidovej sekvencii CpG. Je katalyzovaná enzýmami DNA metyltransferázami (DNMTs) za prítomnosti S-adenozylmetionínu (SAM), ktorý sa uplatňuje ako donor metylovej skupiny. Metylácia / demetylácia je dôležitý mechanizmus na udržanie integrity genómu a zabezpečenie tkanivovo-špecifickej génovej expresie (Obrázok 1). V porovnaní s normálnymi bunkami, nádorové bunky majú narušený metylačný vzorec DNA buď znížením (hypometylácia), alebo zvýšením (hypermetylácia) počtu metylových skupín. V iniciácii onkologických ochorení sú významné predovšetkým hypermetylácia promótorov tumor-supresorových génov, hypometylácia protoonkogénov a globálna hypometylácia (ovplyvňujúca mimogénové a vnútrogénové oblasti), ktorá vedie k zvýšeniu chromozómovej instability. U pacientov s UM bola hypermetylácia DNA identifikovaná ako príčina inaktivácie viacerých génov, z ktorých väčšina sa podieľa na regulácii bunkového cyklu. Patria k nim gény APC, RASSF1A, RARB, LZTS1, CDH1, RB1, CDKN2A, PRAME a iné [17,30].

Modifikácie histónov
Na aktivitu génov majú veľký vplyv aj chemické modifikácie histónových proteínov, ktoré spolu s tesne ovinutým vláknom DNA tvoria komplex nazývaný nukleozóm. Je to základná jednotka chromatínu, vláknitej štruktúry, ktorá v závislosti od stupňa kondenzácie zohráva kľúčovú úlohu v kontrole expresie génov (Obrázok 2). Tvorba neaktívneho, kondenzovaného heterochromatínu, je viazaná s nízkym stupňom acetylácie a vysokým stupňom metylácie histónových molekúl. Menej kondenzovaný euchromatín je transkripčne aktívny [31]. Pre pochopenie úlohy modifikácií histónov v progresii UM, je prelomový objav úlohy Bap1 proteínu v regulácii diferenciácie zárodočných buniek. Strata expresie Bap1 bráni acetylácii histónu H3K27 v promótoroch kľúčových génov, regulujúcich diferenciáciu ektodermu, mezodermu a neurálnej lišty, čo vedie k zníženiu ich expresie [13]. Rovnováha medzi acetyláciou a deacetyláciou génu je určená relatívnymi aktivitami histón acetyltransferáz a HDAC. Inhibítory HDAC umožňujú re-expresiu epigeneticky inaktivovaných génov.
![Schematické znázornenie vplyvu modifikácií histónov a metylácie DNA
na kondenzáciu chromatínu. Voľná štruktúra chromatínu tzv. euchromatín, s acetyláciou a metyláciou histónových molekúl - bez metylácie DNA, je transkripčne
aktívny. Kondenzovaný heterochromatín s metylovanou DNA je transkripčne
inaktívny. Metylované CpG dinukleotidy na DNA sú označené čiernou guličkou,
nemetylované CpG dinukleotidy bielou guličkou, Ac – acetylácia, Met – metylácia
histónov. Upravené podľa [19].](https://pl-master.mdcdn.cz/media/cache/media_object_image_large/media/image_pdf/41d614cbf5bd8dcf04b2964796ff9ae7.jpeg)
Nekódujúce RNA
Medzi najlepšie preštudované nekódujúce RNA patria malé, 19-25 nukleotidové molekuly RNA, tzv. mikroRNA (miRNA). Primárnym mechanizmom ich fungovania je naviazanie sa na komplementárnu mRNA, čím inhibujú jej transláciu alebo mRNA degradujú (Obrázok 3) [32]. U bunkových línií, v nádorových tkanivách a periférnej krvi pacientov s UM boli popísané zmeny expresie viacerých miRNA, medzi nimi napríklad zvýšená expresia let-7b, miR-20a, miR-125b, miR-143, miR-146a, miR-155, miR-181, miR-193b, miR-199a, miR-223, miR-367, miR-454, miR-652, či znížená expresia miR-9, miR-34b/c, miR-124a, miR-137, miR-144, miR-145, miR-182, miR-204 a iných (viď prehľadový článok [17]). Aberantne exprimované miRNA hrajú dôležitú úlohu pri deregulácii onkogénnych dráh v UM, a môžu podporovať metastatické šírenie [33]. Okrem toho, že môžu byť zaujímavým diagnostickým a prognostickým biomarkerom, ponúkajú nám tiež sľubný terapeutický cieľ. Bolo dokázané, že funkciu špecifických miRNA je možné farmakologicky inhibovať komplementárnymi, chemicky modifikovanými oligonukleotidmi, ktoré sa nazývajú anti-miRs alebo antagomiRs. Preukázali sľubné výsledky v predklinickom vývoji [34] a mohli by kompenzovať zvýšenú expresiu génov onkogénnych dráh a tým pomáhať pri manažmente a liečbe UM. Expresia miRNA môže byť regulovaná prostredníctvom metylácie ich promótorov. Chen a kol. napr. uviedli, že hypometylačné činidlo 5-aza-2’-deoxycytidín (decitabín), umožnilo zvýšenie expresie miR-137 [35]. Podobne bola expresia miR-124a v bunkách UM obnovená po použití decitabínu a inhibítora histón deacetylázy, trichostatínu A [36]. Tieto zistenia sú dôkazom, že jednotlivé epigenetické mechanizmy, okrem plnenia svojej individuálnej úlohy, vzájomne na seba pôsobia na niekoľkých úrovniach a vzájomne interagujú.

Dlhé nekódujúce RNA sú definované ako transkripty RNA s dĺžkou väčšou ako 200 nukleotidov, bez schopnosti kódovať proteíny. Zasahujú do tumorigenézy nádorov, podieľajú sa na procesoch angiogenézy, bunkovej proliferácie, migrácie a apoptózy. U UM bola popísaná znížená expresia PAUPAR a zvýšená expresia viacerých lncRNA, napríklad ROR, HOXA11-AS, FTH1P3, PVT1, CYTOR, BANCR, PVT1/NEAT1 a MALAT1 (viď prehľadový článok [17]).
Potenciál pre epigenetickú terapiu
Strata BAP1 je spojená so stratou melanocytovej diferenciácie a zvýšeným metastatickým potenciálom. Inhibítory HDAC dokázali zvrátiť fenotypové účinky inaktivácie BAP1 indukciou morfologickej diferenciácie a zmenou vysokorizikového na nízkorizikový profil génovej expresie v bunkách UM [37]. Kyselina valproová, trichostatín A, tenovín-6, depsipeptid, panobinostat (LBH-589), vorinostat (kyselina suberanilohydroxámová) entinostat (MS-275), quisinostat, NaB, JSL-1, MC1568 a MC1575 preukázali sľubné protinádorové účinky aj v ďalších predklinických štúdiách u UM [38]. Môžu blokovať proliferáciu nádorových buniek, indukovať zastavenie ich rastu, terminálnu diferenciáciu, bunkovú smrť a inhibíciu angiogenézy. Ukázalo sa tým, že inhibítory HDAC a DNMT by mohli predstavovať alternatívnu adjuvantnú terapiu na predĺženie nádorovej dormancie. V súčasnosti prebiehajú klinické štúdie s kyselinou valproovou a vorinostatom (NCT02068586, NCT01587352), entinostatom (štúdia PEMDAC s pembrolizumabom, entinostatom, NCT02697630) a inhibítorom bromodomény BRD4 PLX2853 (NCT03297424) [39]. Inhibícia proteínov bromodomény a extraterminálnych (BET) proteínov ponúka nový terapeutický prístup pre UM. Ide o epigenetické regulátory tretej generácie, ktoré ovplyvňujú replikáciu DNA, remodeláciu a transkripciu chromatínu. Napriek regulačným schváleniam na liečbu niektorých hematologických malignít je problematika epigenetickej terapie u solídnych nádorov stále kontroverzná. Podľa niektorých autorov globálna hypometylácia DNA v dôsledku použitia inhibítora DNMT1 5-aza-2′-deoxycytidínu u myší viedla k chromozómovej instabilite a zvýšenému výskytu sekundárnych malignít [40,41]. Hoci boli tvrdenia o riziku spojenom s liečbou hypometylačnými látkami inými autormi kritizované [42], účinnosť prvej generácie epigenetických liečiv u pacientov so solídnymi nádormi bola sklamaním [43]. Vďaka vývoju nových zlúčenín a lepšiemu pochopeniu molekulárnej podstaty nádorových ochorení sa však zdá, že epigenetické liečivá by mohli mať dôležitú úlohu v synergii s klasickými terapeutickými prístupmi [43]. Využitie nanotechnológií by mohlo prispieť k zvýšeniu účinnosti a zníženiu tkanivovej toxicity takejto kombinovanej liečby.
ZÁVER
Epigenetické zmeny zohrávajú významnú úlohu v patogenéze onkologických ochorení. Majú reverzibilnú povahu a sú preto dobrým terapeutickým cieľom. V mnohých predklinických štúdiách bolo dokázané, že epigenetické liečivá umožňujú obnovu expresie aberantne inaktivovaných tumor-supresorových génov a zvyšujú senzitivitu rezistentných nádorových buniek na liečbu. Pre objavenie účinnejších liečiv pre adjuvantnú terapiu UM a liečbu metastatickej choroby je nevyhnutné akceptovanie významu epigenetických zmien a pochopenie ich úlohy v patogenéze a progresii tohoto ochorenia. V kombinácii s tradičnými terapeutickými postupmi ako sú imuno-, chemo - alebo rádioterapia by mohli epigenetické liečivá priniesť lepšie výsledky pre doteraz neliečiteľné, pokročilé štádiá UM.
Poďakovanie
Práca bola finančne podporená grantami APVV-17-0369 a VEGA 2/0052/18.
Prehlásenie
Autori článku prehlasujú, že vznik odborného článku, jeho publikovanie a zverejnenie nie je predmetom stretu záujmov a nie je podporené žiadnou farmaceutickou firmou.
Práca nebola zadaná inému časopisu ani inde vytlačená.
Do redakce doručeno dne: 19. 1. 2020
Do tisku přijato dne: 5. 3. 2020
Mgr. Božena Smolková, Ph.D.
Ústav experimentálnej onkológie, Biomedicínske centrum Slovenskej akadémie vied
Dúbravská cesta 9
845 05 Bratislava
Zdroje
1. Furdova, A., Olah, Z., Svetlosakova, Z., et al.: The current state of the evidence of malignant tumors of the eye and its adnexa (dg. C69) in the Slovak Republic and in the Czech Republic. Cesk Slov Oftalmol, 68(5);2012;195–201.
2. Vivet-Noguer, R., Tarin, M., Roman-Roman, S., et al.: Emerging Therapeutic Opportunities Based on Current Knowledge of Uveal Melanoma Biology. Cancers, 11(7);2019 : 1019 s.
3. Furdova, A., Sramka, M., Chorvath, M., et al.: Clinical experience of stereotactic radiosurgery at a linear accelerator for intraocular melanoma. Melanoma Res, 27(5);2017 : 463–8.
4. Furdova, A., Sramka, M., Chorvath, M., et al.: Stereotactic radiosurgery in intraocular malignant melanoma--retrospective study. Neuro Endocrinol Lett, 35(1);2014 : 28–36.
5. Furdova, A., Sramka, M., Waczulikova, I., et al.: Stereotactic Rediosurgery for Uveal Melanoma; Postradiation Complications. Cesk Slov Oftalmol, 71(5);2015 : 134–42.
6. Furdová, A., Ferková, A., Krásnik, V., et al.: Exenterácia očnice pre malígny melanóm choroidey v štádiu T4; možnosti epitetického riešenia. Čes a slov Oftal,71(3);2015.
7. Furdova, A., Babal, P., Kobzova, D., et al.: Uveal melanoma survival rates after single dose stereotactic radiosurgery. Neoplasma, 65(6);2018 : 965–71.
8. Furdova, A., Slezak, P., Chorvath, M., et al.: No differences in outcome between radical surgical treatment (enucleation) and stereotactic radiosurgery in patients with posterior uveal melanoma. Neoplasma, 57(4);2010 : 377–81.
9. Yang, J., Manson, D. K., Marr, B. P., et al.: Treatment of uveal melanoma: where are we now? Therapeutic advances in medical oncology, 10;2018 : 1–17.
10. Khoja, L., Atenafu, E. G., Suciu, S., et al.: Meta-Analysis in Metastatic Uveal Melanoma to Determine Progression-Free and Overall Survival Benchmarks: an International Rare Cancers Initiative (IRCI) Ocular Melanoma study. Ann Oncol, 30;2019 : 1370–1380.
11. Onken, M. D., Worley, L. A., Tuscan, M. D., et al.: An accurate, clinically feasible multi-gene expression assay for predicting metastasis in uveal melanoma. J Mol Diagn, 12(4);2010 : 461–8.
12. Harbour, J. W., Onken, M. D., Roberson, E. D., et al.: Frequent mutation of BAP1 in metastasizing uveal melanomas. Science, 330; 2010 : 1410–3.
13. Kuznetsov, J. N., Aguero, T. H., Owens, D. A., et al.: BAP1) regulates epigenetic switch from pluripotency to differentiation in developmental lineages giving rise to BAP1-mutant cancers. Science Advances, 5(9);2019: eaax1738.
14. Dogrusöz, M., Jager, M. J., Damato, B. Uveal melanoma treatment and prognostication. Asia Pac J Ophthalmol (Phila), 6(2); 2017 : 186–96.
15. Horkovicova, K., Markus, J., Krcova, I., et al.: Mutácia BRAF a možnosti identifikácie prognostických markerov metastázovania uveálneho melanómu. Cesk Slov Oftalmol, 72(4); 2016 : 149-56.
16. Cruz, F., 3rd, Rubin, B. P., Wilson, D., et al.: Absence of BRAF and NRAS mutations in uveal melanoma. Cancer Res, 63(18);2003 : 5761–6.
17. Smolkova, B., Horvathova Kajabova, V., Zmetakova, I., et al.: Role of epigenetic deregulation in hematogenous dissemination of malignant uveal melanoma. Neoplasma, 65(6);2018 : 840–54.
18. Shields, C. L., Kaliki, S., Furuta, M., et al.: American Joint Committee on Cancer classification of posterior uveal melanoma (tumor size category) predicts prognosis in 7731 patients. Ophthalmology, 120(10);2013 : 2066–71.
19. Kaliki, S., Shields, C. L., Shields, J. A. Uveal melanoma: Estimating prognosis. Indian Journal of Ophthalmology, 63(2);2015 : 93–102.
20. Damato, B., Coupland, S. E. Translating uveal melanoma cytogenetics into clinical care. Arch Ophthalmol, 127(4);2009 : 423–9.
21. Coupland, S. E., Lake, S. L., Zeschnigk, M., et al.: Molecular pathology of uveal melanoma. Eye (Lond), 27(2);2013 : 230–42.
22. Robertson, A. G., Shih, J., Yau, C., et al.: Integrative Analysis Identifies Four Molecular and Clinical Subsets in Uveal Melanoma. Cancer Cell, 32(2);2017 : 204–20 e15.
23. Tschentscher, F., Husing, J., Holter, T., et al.: Tumor classification based on gene expression profiling shows that uveal melanomas with and without monosomy 3 represent two distinct entities. Cancer Res, 63(10);2003 : 2578–84.
24. Onken, M. D., Worley, L. A., Ehlers, J. P., et al.: Gene expression profiling in uveal melanoma reveals two molecular classes and predicts metastatic death. Cancer Res, 64(20);2004 : 7205–9.
25. Field, M. G., Decatur, C. L., Kurtenbach, S., et al.: PRAME as an Independent Biomarker for Metastasis in Uveal Melanoma. Clinical Cancer Research, 22(5);2016 : 1234–42.
26. Gupta, M. P., Lane, A. M., DeAngelis, M. M., et al.: Clinical Characteristics of Uveal Melanoma in Patients With Germline BAP1 Mutations. JAMA Ophthalmol, 133(8);2015 : 881–7.
27. Decatur, C. L., Ong, E., Garg, N., et al.: Driver mutations in uveal melanoma: associations with gene expression profile and patient outcomes. JAMA ophthalmology, 134(7);2016 : 728–33.
28. Helgadottir, H., Höiom, V. The genetics of uveal melanoma: current insights. The Application of Clinical Genetics, 9;2016 : 147–55.
29. Herlihy, N., Dogrusoz, M., van Essen, T. H., et al.: Skewed expression of the genes encoding epigenetic modifiers in high-risk uveal melanoma. Invest Ophthalmol Vis Sci, 56(3);2015 : 1447–58.
30. Li, Y., Jia, R., Ge, S. Role of Epigenetics in Uveal Melanoma. International journal of biological sciences, 13(4);2017 : 426–33.
31. Svoreňová, M. L., Smolková, M. B. Úloha epigenetickej regulácie v procese epiteliálno-mezenchýmového prechodu, 12;2017 : 410–413.
32. Catalanotto, C., Cogoni, C., Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. International journal of molecular sciences, 17(10); 2016 : 1712.
33. Smit, K. N., Chang, J., Derks, K., et al.: Aberrant MicroRNA Expression and Its Implications for Uveal Melanoma Metastasis. Cancers, 11(6);2019 : 815.
34. Chakraborty, C., Sharma, A. R., Sharma, G., et al.: Therapeutic miRNA and siRNA: Moving from Bench to Clinic as Next Generation Medicine. Mol Ther Nucleic Acids, 8;2017 : 132–43.
35. Chen, X., Wang, J., Shen, H., et al.: Epigenetics, microRNAs, and carcinogenesis: functional role of microRNA-137 in uveal melanoma. Invest Ophthalmol Vis Sci, 52(3);2011 : 1193–9.
36. Chen, X., He, D., Dong, X. D., et al.: MicroRNA-124a is epigenetically regulated and acts as a tumor suppressor by controlling multiple targets in uveal melanoma. Invest Ophthalmol Vis Sci, 54(3); 2013 : 2248–56.
37. Landreville, S., Agapova, O. A., Matatall, K. A., et al.: Histone deacetylase inhibitors induce growth arrest and differentiation in uveal melanoma. Clin Cancer Res, 18(2);2012 : 408–16.
38. Moschos, M. M., Dettoraki, M., Androudi, S., et al.: The role of histone deacetylase inhibitors in uveal melanoma: current evidence. Anticancer research, 38(7);2018 : 3817–24.
39. Piperno-Neumann, S., Piulats, J. M., Goebeler, M., et al.: Uveal Melanoma: A European Network to Face the Many Challenges of a Rare Cancer. Cancers (Basel), 11(6);2019 : 817.
40. Gaudet, F., Hodgson, J. G., Eden, A., et al.: Induction of Tumors in Mice by Genomic Hypomethylation. Science, 300(5618);2003 : 489–92.
41. Eden, A., Gaudet, F., Waghmare, A., et al.: Chromosomal Instability and Tumors Promoted by DNA Hypomethylation. Science, 300(5618);2003 : 455.
42. Yang, A. S., Estecio, M. R., Garcia-Manero, G., et al.: Comment on „Chromosomal instability and tumors promoted by DNA hypomethylation“ and „Induction of tumors in nice by genomic hypomethylation“. Science, 302(5648);2003 : 1153.
43. Morel, D., Jeffery, D., Aspeslagh, S., et al.: Combining epigenetic drugs with other therapies for solid tumours - past lessons and future promise. Nat Rev Clin Oncol;2019.
Štítky
OftalmologieČlánek vyšel v časopise
Česká a slovenská oftalmologie

2020 Číslo 2
- Selektivní laserová trabekuloplastika nesnižuje nitroční tlak více než argonová laserová trabekuloplastika
- Progresi glaukomu je třeba hodnotit strukturálními i funkčními parametry
- Ztráta centrálního vidění po filtrujících operacích glaukomu
- Od PGF-2 alfa-isopropyl esteru k latanoprostu: přehled vývoje Xalatanu
- Compliance u pacientů s glaukomem
Nejčtenější v tomto čísle
- NEARTERITICKÁ FORMA PŘEDNÍ ISCHEMICKÉ NEUROPATIE OPTIKU - LÉČBA A RIZIKOVÉ FAKTORY
- Vzpomínka na doc. MUDr. Cigánka, CSc.
- BETAXOLOL, BRIMONIDIN A CARTEOLOL V LÉČBĚ NORMOTENZNÍCH GLAUKOMŮ.
- AFLIBERCEPT V LÉČBĚ VASKULÁRNÍ SERÓZNÍ ABLACE PIGMENTOVÉHO LISTU SÍTNICE: ROČNÍ VÝSLEDKY ANATOMICKÝCH A FUNKČNÍCH ZMĚN
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy