
HLA Class I Binding of HBZ Determines Outcome in HTLV-1 Infection
CD8+ T cells can exert both protective and harmful effects on the virus-infected host. However, there is no systematic method to identify the attributes of a protective CD8+ T cell response. Here, we combine theory and experiment to identify and quantify the contribution of all HLA class I alleles to host protection against infection with a given pathogen. In 432 HTLV-1-infected individuals we show that individuals with HLA class I alleles that strongly bind the HTLV-1 protein HBZ had a lower proviral load and were more likely to be asymptomatic. We also show that in general, across all HTLV-1 proteins, CD8+ T cell effectiveness is strongly determined by protein specificity and produce a ranked list of the proteins targeted by the most effective CD8+ T cell response through to the least effective CD8+ T cell response. We conclude that CD8+ T cells play an important role in the control of HTLV-1 and that CD8+ cells specific to HBZ, not the immunodominant protein Tax, are the most effective. We suggest that HBZ plays a central role in HTLV-1 persistence. This approach is applicable to all pathogens, even where data are sparse, to identify simultaneously the HLA Class I alleles and the epitopes responsible for a protective CD8+ T cell response.
Published in the journal:
. PLoS Pathog 6(9): e32767. doi:10.1371/journal.ppat.1001117
Category:
Research Article
doi:
https://doi.org/10.1371/journal.ppat.1001117
Summary
CD8+ T cells can exert both protective and harmful effects on the virus-infected host. However, there is no systematic method to identify the attributes of a protective CD8+ T cell response. Here, we combine theory and experiment to identify and quantify the contribution of all HLA class I alleles to host protection against infection with a given pathogen. In 432 HTLV-1-infected individuals we show that individuals with HLA class I alleles that strongly bind the HTLV-1 protein HBZ had a lower proviral load and were more likely to be asymptomatic. We also show that in general, across all HTLV-1 proteins, CD8+ T cell effectiveness is strongly determined by protein specificity and produce a ranked list of the proteins targeted by the most effective CD8+ T cell response through to the least effective CD8+ T cell response. We conclude that CD8+ T cells play an important role in the control of HTLV-1 and that CD8+ cells specific to HBZ, not the immunodominant protein Tax, are the most effective. We suggest that HBZ plays a central role in HTLV-1 persistence. This approach is applicable to all pathogens, even where data are sparse, to identify simultaneously the HLA Class I alleles and the epitopes responsible for a protective CD8+ T cell response.
Introduction
Human T cell lymphotropic virus-type 1 (HTLV-1) is an oncogenic retrovirus that infects between 10 and 20 million people worldwide. Of these infected individuals, 1–6% develop adult T cell leukaemia/lymphoma (ATL/ATLL) and a further 2 to 3% develop a variety of chronic inflammatory syndromes including HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP); the rest remain lifelong asymptomatic carriers (ACs) of the virus.
Most HTLV-1-infected individuals mount a large, chronically activated CD8+ T cell response to HTLV-1 and it is unclear why this fails to eradicate the virus. Furthermore, there is evidence for both protective [1]–[3] and pathogenic effects [4]–[7] of HTLV-1-specific CD8+ T cells. The attributes of a protective anti-HTLV-1 response in vivo are unknown, although specificity for the viral protein Tax is a strong candidate. There are good reasons to believe that a Tax-specific CD8+ response [8] may be particularly protective. Firstly, Tax is the immunodominant HTLV-1 antigen [9], [10]. Secondly, HLA-A*02, which is associated with protection in southern Japan [11], binds several Tax epitopes [12], notably Tax 11–19, which is bound unusually strongly [13]. Thirdly, Tax is one of the first HTLV-1 proteins to be expressed and it has been shown, for HIV-1-infected cells in vitro, that CD8+ T cells specific to early viral proteins are particularly effective in viral control [14]. Finally, it has been shown that the selective pressure exerted on Tax is higher in asymptomatic carriers than in those that have developed HAM/TSP [15].
What constitutes an effective CD8+ T cell response is difficult to ascertain in any infection. Measurements of CD8+ T cell frequency, phenotype, function and specificity are informative but, because antigen load influences each of these factors, it can be difficult to ascertain if a particular immune profile is the cause or effect of good pathogen control [16]–[19]. An alternative approach is host genotype analysis. Polymorphisms in immune-related genes, particularly the HLA class I genes, have been associated with outcome in many human infections, notably Plasmodium falciparum, Mycobacterium tuberculosis, HIV-1, HTLV-1 and Hepatitis B Virus infection. The benefit of a genotypic analysis is that the direction of causality is unequivocal; the drawback is that, in common with all “omics” approaches to identify biomarkers, mechanistic insight is limited. Provided linkage disequilibrium can be ruled out, class I associations imply that the protective effect is mediated by CD8+ T or NK cells. However, why one particular allele should be protective remains unclear and so a class I association provides no information about how to manipulate the immune response to enhance protection.
The aim of this study was to develop a method to test the hypothesis that the effectiveness of an individual's HTLV-1-specific response and thus their proviral load and HAM/TSP risk was determined by the epitope binding properties of their HLA class I alleles. This method resulted in the identification of the viral protein HTLV-1 basic leucine zipper factor (HBZ) as a significant immunogenic target for both proviral load reduction and reduced disease risk. The HBZ gene was identified recently [20], it is encoded by the complementary strand of the HTLV-1 genome and its promoter lies in the 3′ LTR rather than the 5′ LTR. Our approach is generally applicable to all pathogens, including those in which few epitopes have been identified experimentally.
Results
Verification of epitope prediction software
Approximately 50 HLA class I-epitope pairs have been identified for HTLV-1 [12], [21]–[23] (mainly from the immunodominant protein Tax [24] in the context of A*02); this represents a small and non-random fraction of the ∼2200 nonamer epitopes that could be bound by the alleles of the Kagoshima cohort studied here (Methods). Therefore we used epitope prediction software to systematically predict HTLV-1 epitopes. The epitope prediction software that we used has been extensively validated for a number of other organisms including HIV-1 where it has provided useful insight [25]–[31], but because of the lack of experimental data, it has not previously been tested for HTLV-1. To validate the epitope prediction software, we measured experimentally the binding affinity of 200 HTLV-1 peptide-allele combinations (Table S1 in Supporting Information S1). We found a strong positive correlation between experimental measurement and the theoretical prediction for each of the two epitope prediction methods used namely Metaserver and Epipred (Metaserver: all P <0.00001, Spearman's rank correlation; Fig. 1. Epipred: all P <0.001, Spearman's rank correlation; Fig. S1 in Supporting Information S1). We conclude that these epitope prediction software packages accurately predict relative (i.e. rank order) HTLV-1 peptide binding affinities. Throughout this article figures in the main text are obtained using Metaserver, corresponding figures from Epipred are in Supporting Information S1. All conclusions were replicated by both methods and by an alternative metric (Supporting Information S1).

Protective class I alleles bind HBZ strongly
A number of associations between HLA class I alleles and proviral load or HAM/TSP risk in HTLV-1 infection have been identified in a population in southern Japan [3], [32]. We compared the predicted HTLV-1 peptide-binding affinities of the two protective alleles, A*0201 and Cw*0801, with those of the known detrimental allele, B*5401 (Methods). Peptides from the HTLV-1 protein HBZ bound to HLA-A*0201 and Cw*0801 significantly more strongly compared to B*5401 (P = 0.0002, Wilcoxon-Mann-Whitney; Fig. 2. Repeating the analysis with another protective allele from the A*02 family, namely A*0206 instead of A*0201 yielded identical conclusions P = 0.0007, Wilcoxon-Mann-Whitney, data not shown). These P values needs to be treated with caution because the rank of the binding affinity of one HBZ peptide for A*0201 may not be independent of the rank of the binding affinity of a second peptide to A*0201 and similarly for Cw*0801 and B*5401 (see Methods, independence of ranks). However, we also found that the difference in binding strength (i.e. the rank of the top A*0201 binding peptide minus the rank of the top B*5401 binding peptide) was significantly greater for HBZ than for other HTLV-1 proteins (P <0.001, binomial test). This statistic is based only on the top binding peptide so it does not assume different peptides have independent binding affinity ranks. Henceforth, we only considered the top binding peptide to avoid the potential problem of dependence (Methods).

Asymptomatic carriers bind HBZ more strongly than HAM/TSP patients
Having established that the known protective HLA class I alleles code for molecules that bind to peptides from HBZ more strongly than the known detrimental allele, we examined peptide binding by all alleles in the Kagoshima cohort. We compared the predicted epitopes for asymptomatic carriers (n = 202) and HAM/TSP patients (n = 230) from the Kagoshima cohort. We predicted the HTLV-1 peptides bound most strongly by each individual, given their HLA class I types and then tested for differences between the two subject groups (Methods). The results are shown in Table S2 in Supporting Information S1. One result remained highly statistically significant after correction for multiple comparisons and was consistent across both prediction methods: asymptomatic carriers have HLA class I alleles that bind more strongly to peptides from HBZ compared to HAM/TSP patients (Metaserver: P = 0.0002, Wilcoxon-Mann-Whitney; Fig. 3. Epipred: P <0.0001, Wilcoxon-Mann-Whitney; Fig. S2 in Supporting Information S1).

To test whether this association was caused solely by the known protective and detrimental HLA allele families, the analysis for HBZ was repeated excluding A*02 and B*54. The results showed that, amongst the HLA-A alleles, alleles from the A*02 family were responsible for the protective effect, whereas in HLA-B more than one allele family contributed significant effects. Overall, strong binding of HBZ peptides was associated with asymptomatic status, even when A*02, B*54 and Cw*08 were excluded from the analysis (Metaserver: P = 0.04, Wilcoxon-Mann-Whitney. Epipred: P = 0.006, Wilcoxon-Mann-Whitney; Table 1).

Individuals whose HLA class I genotype predisposed them to bind HBZ peptides strongly had a significantly lower proviral load
Next we investigated why strong binding of HBZ peptides was associated with remaining asymptomatic. One of the strongest correlates of HAM/TSP is a high HTLV-1 proviral load [33]. We therefore tested the hypothesis that strong binding of HBZ peptides was associated with a lower proviral load. The number of HLA class I alleles that each individual possessed that were predicted to strongly bind peptides from HBZ was plotted against their proviral load (Methods). We found that the number of HLA Class I alleles that an individual had that strongly bound HBZ peptides was significantly negatively correlated with their proviral load (Metaserver: P = 0.016, Spearman's rank correlation; Fig. 4. Epipred: P = 0.1, Spearman's rank correlation; Fig. S3 in Supporting Information S1). We tested this correlation independently in HAM/TSP patients and asymptomatic carriers and then combined the P values (rather than simply testing the whole cohort), so this result does not follow trivially from our previous observation than asymptomatic carriers bind HBZ significantly more strongly than HAM/TSP patients. An alternative metric, the binding strength of the top HBZ-binding peptide to each allele instead of the number of strongly binding alleles, yielded an identical conclusion i.e. there was a significant negative correlation between the proviral load and the strength of binding to HBZ peptides (Metaserver: P = 0.008, Spearman's rank correlation. Epipred: P = 0.003, Spearman's rank correlation).

HBZ peptide binding is a consistent predictor of proviral load
Next we compared our peptide-binding analysis of HLA class I genotype with a traditional frequency-based “presence or absence of an allele” analysis. Previously a “traditional” analysis yielded inconsistent results [3], [32], [34]. For example, A*02 was a significant predictor of load in ACs but not in patients with HAM/TSP. We therefore directly compared the ability of the novel peptide binding method and the traditional genotype method to predict proviral load in ACs and HAM/TSP patients (Table S3 in Supporting Information S1). This analysis showed that binding HBZ was a significant predictor of proviral load in both ACs and HAM/TSP patients (P = 0.001, P = 0.017), but confirmed the finding that in a traditional analysis HLA-A*02 (presence/absence) was a significant predictor in ACs only (P = 0.01) and HLA-B*54 for HAM/TSP patients only (P = 0.019). The proportion of variance in proviral load explained was also marginally higher for the peptide binding analysis than the traditional analysis. The observation that HBZ binding strength correlated with proviral load in both ACs and HAM/TSP patients suggests that peptide binding is the more fundamental predictor than HLA genotype.
HLA class I binding of peptides from different HTLV-1 proteins has a differential and correlated impact on both proviral load and HAM/TSP risk
Our findings demonstrate that the HTLV-1 protein that is associated with the most significant reduction in HAM/TSP risk when bound by HLA class I molecules (i.e. HBZ) is also, independently, associated with a significant reduction in proviral load when bound. We wished to investigate whether this relationship held across all proteins. We therefore produced two ranked lists of proteins. In the first list we ranked the HTLV-1 proteins according to whether they were bound more strongly by asymptomatic carriers or HAM/TSP patients (Fig. 5 x-axis; at the extremes ACs were significantly more likely to bind peptides from HBZ, HAM/TSP patients were significantly more likely to bind peptides from Env). This list could be viewed as the rank order of targets for a vaccine designed to reduce HAM/TSP risk. In the second list we ranked the proteins according to whether binding their peptides was associated with a lower proviral load (Fig. 5, y-axis; at the extremes, binding of HBZ was associated with a significantly lower proviral load, whereas binding of Env was associated with a significantly higher proviral load). This list could be viewed as the rank order of targets for a vaccine designed to reduce proviral load. We then compared these two sets of ranks and found them to be strongly positively correlated (Metaserver: RS = 0.86, P = 0.0005, Spearman's rank correlation; Fig. 5. Epipred: RS = 0.66, P = 0.02, Spearman's rank correlation; Fig. S4 in Supporting Information S1). That is, proteins whose peptides are bound strongly by asymptomatic carriers are, independently, those associated with a lower proviral load when bound. This observation has two important implications. Firstly, HLA class I binding of peptides from different proteins has a differential impact on both proviral load and HAM/TSP risk; i.e. CD8+ efficiency (ability to reduce proviral load and disease risk) is determined by protein specificity and we have established a list of protein targets of the most efficient response to the least efficient response. Secondly, the fact that across all alleles and across all proteins, peptide binding associated with immune control (reduced proviral load) is strongly correlated with prevention of HAM/TSP is the strongest evidence yet that the CD8+ T cell response can have a beneficial role in HTLV-1 infection.

The prevented fraction of disease, FP
We calculated the prevented fraction of disease attributable to the possession of one or more strong binding alleles to HBZ [3] (Methods). This showed that the possession of strong HBZ-binding HLA alleles prevented (Fp)≈48% (12.3% SD) of potential cases of HAM/TSP in the study population. However, although we found that a high proportion of potential HAM/TSP cases are prevented by strong HBZ binding, it should be noted that the strength of HBZ binding is not the only determinant of disease status: in a logistic regression model, the strength of HBZ binding alone could only correctly classify 55% of cases of HAM/TSP.
HBZ-specific CD8+ T cells can be detected ex vivo
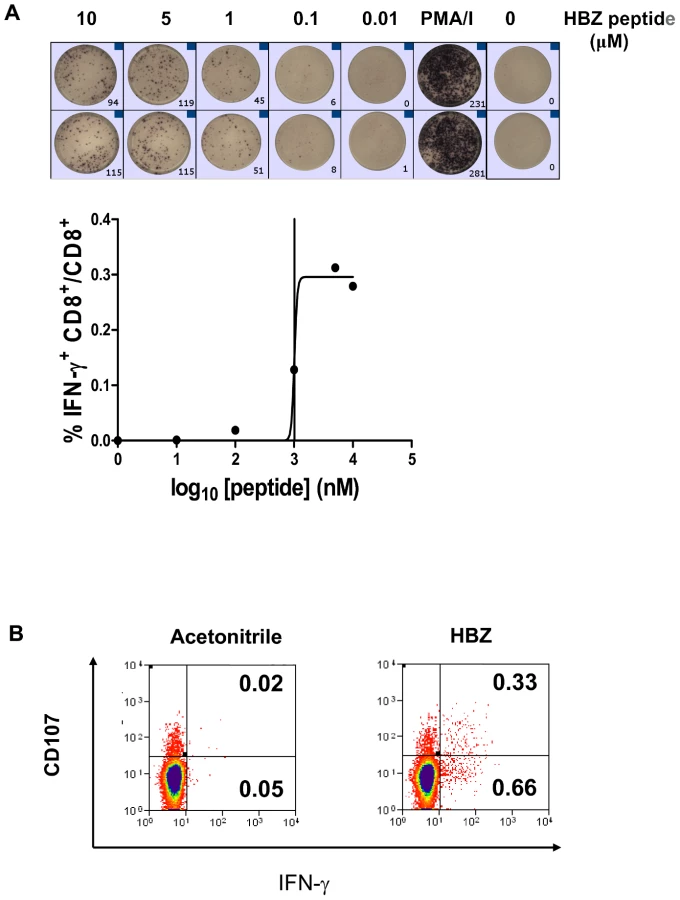
These results strongly imply that HBZ-specific CD8+ T cells play a protective role in HTLV-1 infection. HBZ immunogenicity has been studied in ATL patients [35], [36] but it is unknown whether a HBZ-specific CD8+ T cell response is generated or even whether HBZ protein is expressed in asymptomatic carriers and HAM/TSP patients. We therefore sought to identify HBZ-specific CD8+ T cells in fresh PBMCs from HTLV-1 infected individuals. We assayed IFN-γ production by ELISpot following stimulation in vitro with a pool of overlapping peptides that spanned the entire HBZ protein. Of 45 subjects tested, 31% had detectable HBZ-specific CD8+ T cells (Fig. 6). An independent CD8+ T cell assay, (CD107a mobilisation), confirmed that HBZ-specific CD8+ T cells are present in PBMC from infected individuals. We conclude that HBZ protein is expressed in vivo and is immunogenic.
Naturally infected cells can be lysed by an HBZ-specific CD8+ T cell clone
Recently, Sumeori et al established an HBZ-specific CD8+ T cell clone that recognised HBZ26–34 (GLLSLEEEL) in the context of HLA-A*0201 [37]. They showed that this clone was able to lyse an autologous B-lymphoblastoid cell line (B-LCL) that had been loaded with HBZ peptide but that cells from an ATL patient were resistant to killing. We investigated whether the same CD8+ T cell clone was able to kill naturally-infected cells from non-leukemic HTLV-I-infected individuals. First we confirmed the finding of Sumeori et al that autologous B-LCL loaded with HBZ26–34 peptide could be lysed by the CD8+ T cell clone (data not shown). Then we demonstrated, by a classical chromium release assay, that naturally-infected CD4+CD25+ cells from the PBMCs of 3 out of 4 HLA-A*0201+ non-leukemic patients were lysed by the CD8+ T cell clone but that cells from 3 out of 3 HLA-mismatched donors were not lysed (Fig. 7). We conclude that naturally-infected cells from AC and HAM/TSP patients are susceptible to lysis by an HBZ-specific clone.

The comparative immunogenicity of HBZ and Tax
How does the immunogenicity of HBZ compare to Tax? We compared the predicted top binding peptide from HBZ and Tax respectively to 43 HLA class I alleles (the maximum capacity of Metaserver). Peptides from Tax were predicted to bind significantly more strongly than peptides from HBZ (P = 0.00002, paired Wilcoxon-Mann-Whitney; Fig. 8A). Consistent with this prediction, the frequency of Tax-specific CD8+ T cells by IFN-γ ELISpot was also significantly greater compared to HBZ CD8+ T cells in 45 HTLV-1-infected individuals (P = 0.000006, paired Wilcoxon-Mann-Whitney; Fig. 8B).

Discussion
We show that strong predicted binding of peptides from the HTLV-1 protein HBZ is associated with a reduced risk of HAM/TSP and a reduced proviral load in a population with endemic HTLV-1 infection in southern Japan. We demonstrated that protection is not limited to a small subset of HLA class I alleles previously associated with disease status and proviral load (HLA-A*02 and HLA-Cw*08), but is generally associated with HLA class I alleles that bind strongly to HBZ. Given that a protein-specific HLA-restricted association is more likely to be mediated by CD8+ T cells than NK cells which show limited protein specificity we interpret this work in the context of the CD8+ T cell response.
Prior to this analysis, CD8+ T cells specific for the HTLV-1 protein Tax were often considered as the best candidate for ‘efficient’ or ‘protective’ CD8+ cells because of the immunodominance of Tax in the CD8+ T cell response [9], [24]. Our finding that binding of HBZ peptides rather than Tax peptides is protective raises the question: why is HBZ a critical target for the immune response?
HBZ functions by binding to cellular factors of the JUN and ATF/CREB families [37]. There are two major splice variants of the HBZ transcript, SP1 and SP2; the variant SP1 is more abundant and is the variant used in this study [38], [39]. The abundance of HBZ transcript has been previously correlated with disease severity [39]. Expression of HBZ suppresses Tax-mediated transactivation through the 5′ LTR [20], [40] and thereby inhibits expression of other HTLV-1 genes [20], [41]; HBZ can be expressed in the absence of transcription of other HTLV-1 genes. Additionally, HBZ RNA promotes the proliferation of infected T-lymphocytes [35]. This dual action – reduction of HTLV-1 expression and subsequent protection from immune surveillance, and enhancement of infected cell proliferation – probably confers a survival advantage on HBZ-expressing cells and is consistent with the observations that HBZ enhances persistence in HTLV-1 inoculated rabbits [41] and that ATL cells often have a hypermethylated or deleted 5′ LTR but an intact functional 3′ LTR [35]. We hypothesise that if HBZ-specific CD8+ T cells are weak or absent then infected cells that express HBZ but not other viral proteins will evade immune surveillance and proliferate rapidly, leading to an increase in proviral load. HBZ-specific CD8+ T cells would then play an important role in preventing this proliferation of provirus-positive cells and blocking this strategy of persistence. If this conclusion is correct that CD8+ T cell recognition of HBZ plays a central role in the control of HTLV-1 replication then one might expect that HBZ would have evolved to minimize class I binding. Consistent with this hypothesis, we find that the predicted binding affinity of HLA molecules to HBZ peptides is significantly weaker than that of Tax peptides and that the frequency of HBZ-specific CD8+ T cells is significantly lower than the frequency of Tax-specific CD8+ T cells. Although the low immunogenicity of HBZ is precisely what we predict given its central importance in maintaining HTLV-1 persistence, it is nevertheless striking that these low T cell frequency responses are so important. This result challenges the prevailing assumption that the immunodominant response to a pathogen is the most important.
We demonstrated using two different assays (IFNγ ELISpot and CD107 mobilisation) that HBZ-specific CTL are present in PBMC from HAM/TSP patients and ACs. We further show that naturally infected cells, isolated directly from HAM/TSP patients and ACs, are susceptible to lysis by an HBZ-specific CTL clone. Suemori et al have previously reported that the same HBZ-specific CTL clone was unable to lyse leukemic cells isolated from a patient with adult T cell leukemia [37]. The observation that aleukemic but not leukemic cells can be lysed may be because leukemic cells express lower levels of HLA: HBZ peptide on their surface or because leukemic cells can be inherently harder to lyse [42]–[44].
This approach to studying the association between HLA class I genotype and the outcome of infection has a number of strengths compared with a traditional frequency-based analysis. Firstly, it is more mechanistic: knowing that binding HBZ is associated with a reduced proviral load and disease risk compared with knowing that A*02 is associated with these outcomes is a simultaneously more fundamental and more applicable level of understanding. Secondly, identification of protective epitopes immediately suggests a practical approach to measure and enhance, via therapeutic vaccination, the efficiency of an individual's anti-viral response. Thirdly, because the same effect (e.g. HBZ binding) can be identified for many alleles it is less likely to be a spurious result of linkage disequilibrium or genetic stratification. Finally, effects due to multiple low-frequency alleles can be captured because analysis is made at the level of peptide binding rather than allelic frequency.
In summary, using a novel and generalizable approach, we have identified one of the constituents of an effective CD8+ T cell response in HTLV-1 infection.
Methods
Subjects
Kagoshima cohort
Two hundred and thirty individuals with HAM/TSP were compared with two hundred and two randomly selected HTLV-1 seropositive asymptomatic carriers (ACs) from the Kagoshima Red Cross Blood Transfusion Service. All cases and controls were of Japanese ethnic origin and resided in Kagoshima Prefecture, Japan. Full details of the cohort can be found in [3]. Each individual was HLA class I typed using PCR–sequence-specific primer reactions. Their proviral load in peripheral blood mononuclear cells was measured by quantitative PCR.
St Mary's cohort
IFNγ ELISpot, CD107a mobilisation and the chromium release assays were all carried out using cells from HTLV-1-infected individuals attending St Mary's Hospital. Ethics statement: All subjects attended the National Centre for Human Retrovirology at St. Mary's Hospital and donated blood after giving written informed consent as approved by the St. Mary's Hospital Research Ethics Committee.
Epitope prediction
We used two different algorithms to predict HLA class I epitopes: Metaserver and Epipred. Figures based on Metaserver predictions are in the main text, the corresponding figures for Epipred are in Supporting Information S1.
Metaserver
Metaserver is a combination of two web-based prediction methods that use artificial neural nets, NetCTL v1.2 [45] and NetMHC v3.0 [46], [47]. NetCTL is an integrated method that predicts TAP transport, proteasomal cleavage and HLA binding for 12 different class I alleles. NetMHC v3.0 predicts HLA-peptide binding for 43 HLA molecules. Metaserver combines the two methods and removes a normalising assumption (which held that all alleles bind the same number of peptides) to produce a technique that shows improved accuracy in epitope prediction [48] and predicts epitopes for 43 HLA molecules.
Epipred
In order to validate our results, we used a second, independent method of epitope prediction [49]. Epipred uses a logistic regression model that is trained on all available data across all HLA class I alleles and then specified for an individual allele.
Epitope prediction - allele coverage
Other than our initial comparison (protective against detrimental alleles), analysis was limited to A and B loci for two reasons: Metaserver does not have algorithms for the C loci and C loci predictors tend to be less accurate because of the lack of peptide-HLA-C experimental binding affinities to train the software. Metaserver provided coverage of 84% of the total count of A/B alleles in the Kagoshima cohort.
The missing alleles were: A*0207, A*0210, A*2603, A*3201, B*1301, B*1501, B*1508, B*1511, B*1518, B*2704, B*3701, B*3802, B*4005, B*4006, B*4601, B*4801, B*5201, B*5501, B*5504, B*5601, B*5603, B*5605, B*5705, B*5901, and B*6701.
We were able to obtain predictions for [A*0207, A*0210], A*2603 and [B*4005, B*4006] to a resolution of 2 digits by combining the predictions of other A02*, A26* and B40* predictors according to their frequency in Kagoshima.
Estimated number of epitope-allele combinations
We estimate that approximately 2,200 peptides could be bound by the alleles present in the Kagoshima cohort. This figure is 1% [45] of the 3,389 overlapping nonamers of the HTLV-1 proteome multiplied by the number of unique alleles (65) in the cohort.
Prediction quality
The accuracy of epitope prediction algorithms has increased to such an extent that the correlation between predicted binding affinities and measured binding affinity is as strong as the correlations of measurements between different laboratories [50]. The specificity of epitope predictors has been tested by predicting a set of CTL epitopes and subsequently verifying CD8+ T cell responses against these epitopes experimentally. Using this technique has yielded true-positive (correctly predicted) estimates of 62–80% [51]. Using the more direct approach of mass spectrometry to determine HLA-peptide binding yielded a true positive rate of greater than 98% [52]. Additionally, we verified the prediction software we used (Metaserver and Epipred) for HTLV-1 peptides.
The rank measure
Both prediction methods that we use produce a score for each peptide-HLA that represents the binding strength of that complex. In theory this score would allow us to compare predicted binding affinities between alleles. However, between allele comparisons can be problematic. Firstly, within-allele comparisons (i.e. predictions for different peptides to the same allele) are thought to be more comparable than predictions between alleles [45]. Secondly, whether or not a normalisation procedure should be applied for between-allele comparisons is still being debated in the community [48]. To avoid the potential problem of between-allele comparisons we used the rank measure technique introduced by Borghans et al. [53] in which she quantified the strength of peptide-HLA class I binding for peptides from a particular protein by ranking the binding score of peptides from the protein of interest to the allele amongst the binding score of peptides from the entire proteome to that allele; this approach has been successfully applied in the context of HIV infection [25], [28]. Specifically, we split each protein in the HTLV-1 reference sequence into overlapping nonamers offset by a single amino acid. Using the epitope prediction software, a predicted binding affinity score was calculated for each of these peptides to each HLA allele of interest. For each allele we ranked all nonamers from the proteome from the strongest to weakest predicted binding scores. This produced a list of rank values for each protein to that particular allele that quantified the binding relationship between that allele and the protein (an example is given in Table S4 in Supporting Information S1). To check for robustness we also repeated all calculations using an alternative to the rank measure: the raw predicted affinity score. We found that our conclusions were robust to the choice of method (Table S5 in Supporting Information S1).
Independence of ranks
We were concerned that the binding of the top 8 peptides from a protein to an allele may not be independent of one another. Since, the strength of the strongest binder provides information (i.e. an upper bound) about the strength of the second highest binder. For this reason, apart from Fig. 2, only the top rank for each protein-allele pair was used.
Experimental quantification of HLA class 1-peptide binding
The REVEAL HLA-peptide binding assay (ProImmune Ltd., Oxford, UK) was used to quantify peptide-HLA binding. For each allele-peptide combination that was tested, assembly of peptide-HLA complexes was quantified by ELISA with a conformation-dependent anti-HLA antibody. Samples of assembling peptide-HLA complexes were taken at a defined time point and snap-frozen in liquid nitrogen prior to analysis. The assembly for each peptide-HLA complex was then compared against a positive control peptide for that allele as the percentage of assembled peptide relative to that control. We selected four HLA class I alleles and 50 HTLV-1 peptides for each allele. The allele choice was based on allele frequency in the Kagoshima database and included 2 A alleles and 2 B alleles as well as alleles for which we knew that the epitope prediction tended to be poor. The 50 HTLV-1 nonamer peptides for each allele were selected to represent a range of predicted binding affinities, from weak to strong binding peptides. They originated from 4 HTLV-1 reference strain proteins: Tax, HBZ, Gag and Polymerase.
Protective versus detrimental alleles
Due to allele coverage (see above), it was necessary to use Metaserver for A*0201 and B*5401 and Epipred for Cw*0801. As the rank values were derived for each allele separately, it was acceptable to use different prediction methods for each allele in this case. Epipred predicts binding to allele families rather than individual alleles and so we calculated binding to Cw*08. The ranks of the strongest binding 8 peptides from each protein to the alleles A*0201 and Cw*08 (16 rank values) were compared against the ranks of the strongest binding 8 peptides to the allele B*5401 (8 rank values). A Wilcoxon-Mann-Whitney test was performed for each protein to test for differences between the two sets of rank values. The analysis was repeated using top 5 and top 10 as well as top 8 binding peptides, conclusions were robust to the choice of number of peptides (Results in Supporting Information S1). Finally, to avoid the potential problem of lack of independence of ranks (see “independence of ranks” above) we performed a binomial test on the difference in strength of binding of A*02 and B*54 to HBZ compared to all other HTLV-I proteins. The null hypothesis we tested was “the difference in binding of detrimental and protective alleles to HBZ is comparable to the other HTLV-1 proteins”. For each of the 12 HTLV-1 proteins we calculated the ranks of the single highest ranking peptide from that protein to A*02 and B*54. We then calculated the difference of these two ranks (detrimental – beneficial) for each of the 12 proteins and asked, using the Binomial test, whether the difference in binding for HBZ was larger than would be expected under the null hypothesis.
HAM/TSP versus asymptomatic carriers
The analysis was carried out on each HTLV-1 protein in turn. For each individual in the Kagoshima cohort, the rank of the top binding peptide from the HTLV-1 protein to each of the individual's A and B HLA class I alleles was found (see The Rank Measure). These ranks were then split into two groups – those from HAM/TSP patients and those from asymptomatic carriers (AC). The two sets of ranks (HAM/TSP vs. AC) were then compared for each protein using a Wilcoxon-Mann-Whitney test (null hypothesis: HAM/TSP patients and asymptomatic carriers bind the protein equally strongly).
Rank versus proviral load
We considered each HTLV-1 protein in turn. Firstly, we split the cohort by disease status (AC or HAM/TSP). Then, for each individual, we counted the number of alleles they possessed that were strong binders to the protein of interest and then tested for a correlation between the number of strong binders to the protein and proviral load using the Spearman rank correlation. A strong binding allele to a particular protein was defined as one that was in the top 40% of alleles. That is, the rank of the top binding peptide from the HTLV-1 protein to each of the individual's A and B HLA class I alleles was found (see The Rank Measure). This set of rank values (pooled HAM/TSP and AC) was then ordered from highest to lowest rank and the alleles that were represented in the top 40% of these ranks were defined as strong binding alleles to that protein. Importantly, for each protein, we looked at the relationship between strength of binding and proviral load separately in HAM/TSP patients and ACs and then combined the P values using Fisher's combined test (rather than simply looking at the relationship in the whole cohort). Therefore we could be confident that any relationship between protein binding and proviral load that we found did not follow trivially from a relationship between protein binding and disease status and the fact that asymptomatic carriers have a significantly lower load than HAM/TSP patients.
Our alternative metric for this method used the Rank Measure to quantify the strength of binding of peptides from each HTLV-1 protein to each individual's A and B alleles. We then tested for any correlation between these values and the individuals' proviral load for HAM/TSP patients and asymptomatic carriers.
Robustness of conclusions
All analysis was performed with two independent epitope prediction algorithms (Metaserver and Epipred) and with two different methods (rank method, raw score method); additionally an alternative approach to comparing protective v detrimental alleles (based on the binomial test) and to comparing proviral load with strength of binding were investigated. Conclusions were highly robust (Table S5 in Supporting Information S1).
Statistical analysis
All statistical analysis was carried out using the R Project for Statistical Computing [54]. The tests were non-parametric with the exception of multiple linear regression. All P values reported are 2-tailed. Fisher's combined probability test was used to combine P values.
Multivariate regression
General linear model analysis [55] was used to identify which factors were predictors of proviral load, either in ACs or patients with HAM/TSP.
Prevented fraction of disease, FP
To calculate the prevented fraction (Fp) of disease [3], [56], we used a 2×2 contingency table. The entries in the four cells were as follows: a (HAM/TSP, positive for protective genotype) = 183, b (HAM/TSP, negative for protective genotype) = 47, c (AC, positive for protective genotype) = 181, d (AC, negative for protective genotype) = 21. The fraction (Fp) of potential cases of HAM/TSP in the population that is prevented by the protective genotype is given by Fp = (1−R)×[1−(d×r1/b×r2)], where R = prevalence rate of HAM/TSP in the population (estimated as 1% of the HTLV-1-infected population), r1 = a+b and r2 = c+d. Fp is approximately normally distributed: the standard deviation is given by SD (Fp) = (1−R−Fp)×√[(c/d×r2)+(a/b×r1)].
Detection of HTLV-1-specific CD8+ T cells
Peripheral blood mononuclear cells (PBMC) were isolated from whole blood from HTLV-1 infected individuals by density gradient centrifugation.
IFNγ ELISpot
PBMC were depleted of CD4+ T cells using MACS beads (Miltenyi Biotec). The resulting cells were cultured in duplicate at a density of 100,000 cells per well in the presence of a range of concentrations of pooled overlapping 20mer peptides (offset by 6 amino acids) spanning HBZ, Tax, or with medium alone. After 6 hours, IFN-γ producing cells were detected by ELISpot (Mabtech). The threshold for a positive response to peptide was defined as greater than the mean plus two standard deviations of the number of spots in the medium only control.
CD107 mobilisation assay
1×106 PBMC were cultured in a 400 µl volume in the presence of 5 µl anti-CD107a-PE (eBioscience, CA) 1.4 µg/ml monensin (eBioscience), 20 µg/ml DNase (Sigma Aldrich, UK) with 2 µM HBZ peptide pool (Mimotopes, Australia), or the equivalent volume of peptide solvent, acetonitrile. After 5h, the cells were harvested and stained for 30 min with Live/Dead red (Invitrogen,CA), a fixable viability stain. Cells were fixed and permeablised using ebioscience FoxP3 staining buffer set according to the manufacturer's instructions, then stained with anti-CD3-APC-eFluor780, anti-CD4-eFluor450, anti-CD8-PECy5 (all eBioscience) anti-IFN-γ-FITC, anti-CD14-ECD, anti-CD19-ECD (all Beckman Coulter, France). Samples analysed by flow cytometry using a Cyan ADP (Beckman Coulter), and summit software (DAKO). Doublets, dead cells, monocytes and B cells were excluded from the analysis on the basis of forward and side scatter, pulse width, viability staining, and CD14 or CD19 expression. Antigen-specific CD8+ T cells were identified as CD3+CD8+ cells capable of producing IFN-γ and/or mobilising CD107 to the cell surface.
CD8+ lysis assay: chromium release
PBMC from HLA A*0201+ and HLA A*0201− HTLV-1 infected individuals were depleted of CD8+ cells, then enriched for CD25+ cells using MACS beads (Miltenyi Biotech, Germany), according to manufacturer's instructions. CD25+ cells were cultured for 16h to allow for viral antigen expression and presentation, then labelled with 51Cr by incubating for 1h in the presence of 50–100 µCi Na2CrO4 (MP Biomedicals, USA). Labelled cells were washed extensively and placed in culture in triplicate (40,000 cells/well) in the presence of defined ratios of HBZ-1, a CTL clone which recognises HBZ 26–34 in the context of HLA A*0201 [37], alone or in the presence of 5% Triton x-100 (Sigma Aldrich). As a control, 51Cr labelled B-LCL (autologous to the CTL clone) were cultured at the same ratios, with and without 1 µM HBZ 26–34 peptide. After 4h, culture supernatants were harvested, placed on a scintillation plate, and 51Cr release was assayed using a beta counter. Total specific lysis was calculated using the following formula: [chromium release (test well)−chromium release (no CTL control)]/ [chromium release (Triton−100% lysis)−chromium release (no CTL control)]*100, expressed as a percentage specific lysis of total cells. As not all CD4+CD25+T cells are infected, and thus do not represent targets for the CTL line, an estimate of specific lysis of infected cells was also calculated, making the conservative assumption that all the viral load is present in CD25+ cells, [57], and that the CTL line only kills infected cells. Percentage infected cells lysed was calculated using the following formula: [Percentage total cells lysed]/[fraction of CD4+CD25+ cells that are infected i.e. provirus positive].
HTLV-1 proteome
The reference strain is from [58], with the exception of HBZ, which was identified more recently and described in [35] (Supporting Information S1: HTLV-1 reference strain).
Supporting Information
Zdroje
1. VineAM
HeapsAG
KaftantziL
MosleyA
AsquithB
2004 The role of CTLs in persistent viral infection: cytolytic gene expression in CD8+ lymphocytes distinguishes between individuals with a high or low proviral load of human T cell lymphotropic virus type 1. J Immunol 173 5121 5129
2. AsquithB
MosleyAJ
BarfieldA
MarshallSEF
HeapsA
2005 A functional CD8+ cell assay reveals individual variation in CD8+ cell antiviral efficacy and explains differences in human T-lymphotropic virus type 1 proviral load. J Gen Virol 86 1515 1523
3. JefferyKJ
UsukuK
HallSE
MatsumotoW
TaylorGP
1999 HLA alleles determine human T-lymphotropic virus-I (HTLV-I) proviral load and the risk of HTLV-I-associated myelopathy. Proc Natl Acad Sci U S A 96 3848 3853
4. JacobsonS
2002 Immunopathogenesis of human T cell lymphotropic virus type I-associated neurologic disease. J Infect Dis 186 Suppl 2 S187 S192
5. KubotaR
KawanishiT
MatsubaraH
MannsA
JacobsonS
2000 HTLV-I specific IFN-gamma+ CD8+ lymphocytes correlate with the proviral load in peripheral blood of infected individuals. J Neuroimmunol 102 208 215
6. JacobsonS
ShidaH
McFarlinDE
FauciAS
KoenigS
1990 Circulating CD8+ cytotoxic T lymphocytes specific for HTLV-I pX in patients with HTLV-I associated neurological disease. Nature 348 245 248
7. GretenTF
SlanskyJE
KubotaR
SoldanSS
JaffeeEM
1998 Direct visualization of antigen-specific T cells: HTLV-1 Tax11-19 - specific CD8(+) T cells are activated in peripheral blood and accumulate in cerebrospinal fluid from HAM/TSP patients. Proc Natl Acad Sci U S A 95 7568 7573
8. SundaramR
SunY
WalkerCM
LemonnierFA
JacobsonS
2003 A novel multivalent human CTL peptide construct elicits robust cellular immune responses in HLA-A*0201 transgenic mice: implications for HTLV-1 vaccine design. Vaccine 21 2767 2781
9. KannagiM
HaradaS
MaruyamaI
InokoH
IgarashiH
1991 Predominant recognition of human T cell leukemia virus type I (HTLV-I) pX gene products by human CD8+ cytotoxic T cells directed against HTLV-I-infected cells. Int Immunol 3 761 767
10. GoonPK
BiancardiA
FastN
IgakuraT
HanonE
2004 Human T cell lymphotropic virus (HTLV) type-1-specific CD8+ T cells: frequency and immunodominance hierarchy. J Infect Dis 189 2294 2298
11. JefferyKJ
UsukuK
HallSE
MatsumotoW
TaylorGP
1999 HLA alleles determine human T-lymphotropic virus-I (HTLV-I) proviral load and the risk of HTLV-I-associated myelopathy. Proc Natl Acad Sci U S A 96 3848 3853
12. ParkerCE
NightingaleS
TaylorGP
WeberJ
BanghamCR
1994 Circulating anti-Tax cytotoxic T lymphocytes from human T-cell leukemia virus type I-infected people, with and without tropical spastic paraparesis, recognize multiple epitopes simultaneously. J Virol 68 2860 2868
13. HausmannS
BiddisonWE
SmithKJ
DingYH
GarbocziDN
1999 Peptide recognition by two HLA-A2/Tax11-19-specific T cell clones in relationship to their MHC/peptide/TCR crystal structures. J Immunol 162 5389 5397
14. BaalenCA
GuillonC
Baalen MvM
VerschurenEJ
BoersPH
2002 Impact of antigen expression kinetics on the effectiveness of HIV-specific cytotoxic T lymphocytes. Eur J Immunol 32 2644 2652
15. NiewieskS
DaenkeS
ParkerCE
TaylorG
WeberJ
1994 The transactivator gene of human T-cell leukemia virus type I is more variable within and between healthy carriers than patients with tropical spastic paraparesis. J Virol 68 6778 6781
16. BarberDL
WherryEJ
MasopustD
ZhuB
AllisonJP
2006 Restoring function in exhausted CD8 T cells during chronic viral infection. Nature 439 682 687
17. LichterfeldM
YuXG
MuiSK
WilliamsKL
TrochaA
2007 Selective depletion of high-avidity human immunodeficiency virus type 1 (HIV-1)-specific CD8+ T cells after early HIV-1 infection. J Virol 81 4199 4214
18. StreeckH
BrummeZL
AnastarioM
CohenKW
JolinJS
2008 Antigen load and viral sequence diversification determine the functional profile of HIV-1-specific CD8+ T cells. PLoS Med 5 e100
19. BanghamCR
2009 CTL quality and the control of human retroviral infections. Eur J Immunol 39 1700 1712
20. GaudrayG
GachonF
BasbousJ
Biard-PiechaczykM
DevauxC
2002 The complementary strand of the human T-cell leukemia virus type 1 RNA genome encodes a bZIP transcription factor that down-regulates viral transcription. J Virol 76 12813 12822
21. KubotaR
HanadaK
FurukawaY
ArimuraK
OsameM
2007 Genetic stability of human T lymphotropic virus type I despite antiviral pressures by CTLs. J Immunol 178 5966 5972
22. PiqueC
ConnanF
LevilainJP
ChoppinJ
DokhélarMC
1996 Among all human T-cell leukemia virus type 1 proteins, tax, polymerase, and envelope proteins are predicted as preferential targets for the HLA-A2-restricted cytotoxic T-cell response. J Virol 70 4919 4926
23. SundaramR
SunY
WalkerCM
LemonnierFA
JacobsonS
2003 A novel multivalent human CTL peptide construct elicits robust cellular immune responses in HLA-A*0201 transgenic mice: implications for HTLV-1 vaccine design. Vaccine 21 2767 2781
24. GoonPKC
BiancardiA
FastN
IgakuraT
HanonE
2004 Human T cell lymphotropic virus (HTLV) type-1-specific CD8+ T cells: frequency and immunodominance hierarchy. J Infect Dis 189 2294 2298
25. BorghansJAM
MølgaardA
de BoerRJ
KesmirC
2007 HLA Alleles Associated with Slow Progression to AIDS Truly Prefer to Present HIV-1 p24. PLoS ONE 2 e920
26. Vider-ShalitT
AlmaniM
SaridR
LouzounY
2009 The HIV hide and seek game: an immunogenomic analysis of the HIV epitope repertoire. Aids 23 1311 1318
27. Vider-ShalitT
SaridR
MamanK
TsabanL
LeviR
2009 Viruses selectively mutate their CD8+ T-cell epitopes–a large-scale immunomic analysis. Bioinformatics 25 i39 44
28. HoofI
KesmirC
LundO
NielsenM
2008 Humans with chimpanzee-like major histocompatibility complex-specificities control HIV-1 infection. Aids 22 1299 1303
29. SchellensIM
KesmirC
MiedemaF
van BaarleD
BorghansJA
2008 An unanticipated lack of consensus cytotoxic T lymphocyte epitopes in HIV-1 databases: the contribution of prediction programs. Aids 22 33 37
30. SchmidBV
KesmirC
de BoerRJ
2009 The distribution of CTL epitopes in HIV-1 appears to be random, and similar to that of other proteomes. BMC Evol Biol 9 184
31. KosmrljA
ReadEL
QiY
AllenTM
AltfeldM
Effects of thymic selection of the T-cell repertoire on HLA class I-associated control of HIV infection. Nature 465 350 354
32. JefferyKJ
SiddiquiAA
BunceM
LloydAL
VineAM
2000 The influence of HLA class I alleles and heterozygosity on the outcome of human T cell lymphotropic virus type I infection. J Immunol 165 7278 7284
33. NagaiM
UsukuK
MatsumotoW
KodamaD
TakenouchiN
1998 Analysis of HTLV-I proviral load in 202 HAM/TSP patients and 243 asymptomatic HTLV-I carriers: high proviral load strongly predisposes to HAM/TSP. J Neurovirol 4 586 593
34. VineAM
WitkoverAD
LloydAL
JefferyKJM
SiddiquiA
2002 Polygenic control of human T lymphotropic virus type I (HTLV-I) provirus load and the risk of HTLV-I-associated myelopathy/tropical spastic paraparesis. J Infect Dis 186 932 939
35. SatouY
YasunagaJ-i
YoshidaM
MatsuokaM
2006 HTLV-I basic leucine zipper factor gene mRNA supports proliferation of adult T cell leukemia cells. Proc Natl Acad Sci U S A 103 720 725
36. MatsuokaM
GreenP
2009 The HBZ gene, a key player in HTLV-1 pathogenesis. Retrovirology 6 71
37. SuemoriK
FujiwaraH
OchiT
OgawaT
MatsuokaM
2009 HBZ is an immunogenic protein, but not a target antigen for human T-cell leukemia virus type 1-specific cytotoxic T lymphocytes. J Gen Virol 90 1806 1811
38. UsuiT
YanagiharaK
TsukasakiK
MurataK
HasegawaH
2008 Characteristic expression of HTLV-1 basic zipper factor (HBZ) transcripts in HTLV-1 provirus-positive cells. Retrovirology 5 34
39. SaitoM
MatsuzakiT
SatouY
YasunagaJ-I
SaitoK
2009 In vivo expression of the HBZ gene of HTLV-1 correlates with proviral load, inflammatory markers and disease severity in HTLV-1 associated myelopathy/tropical spastic paraparesis (HAM/TSP). Retrovirology 6 19
40. BasbousJ
ArpinC
GaudrayG
PiechaczykM
DevauxC
2003 The HBZ factor of human T-cell leukemia virus type I dimerizes with transcription factors JunB and c-Jun and modulates their transcriptional activity. J Biol Chem 278 43620 43627
41. LiM
KesicM
YinH
YuL
GreenPL
2009 Kinetic Analysis of Human T-cell Leukemia Virus Type 1 Gene Expression in Cell Culture and Infected Animals. J Virol
42. AzumaT
OtsukiT
KuzushimaK
FroelichCJ
FujitaS
2004 Myeloma cells are highly sensitive to the granule exocytosis pathway mediated by WT1-specific cytotoxic T lymphocytes. Clin Cancer Res 10 7402 7412
43. JiangSB
OjciusDM
PersechiniPM
YoungJD
1990 Resistance of cytolytic lymphocytes to perforin-mediated killing. Inhibition of perforin binding activity by surface membrane proteins. J Immunol 144 998 1003
44. MullerC
TschoppJ
1994 Resistance of CTL to perforin-mediated lysis. Evidence for a lymphocyte membrane protein interacting with perforin. J Immunol 153 2470 2478
45. LarsenMV
LundegaardC
LamberthK
BuusS
BrunakS
2005 An integrative approach to CTL epitope prediction: a combined algorithm integrating MHC class I binding, TAP transport efficiency, and proteasomal cleavage predictions. Eur J Immunol 35 2295 2303
46. BuusS
LauemøllerSL
WorningP
KesmirC
FrimurerT
2003 Sensitive quantitative predictions of peptide-MHC binding by a ‘Query by Committee’ artificial neural network approach. Tissue Antigens 62 378 384
47. NielsenM
LundegaardC
WorningP
LauemøllerSL
LamberthK
2003 Reliable prediction of T-cell epitopes using neural networks with novel sequence representations. Protein Sci 12 1007 1017
48. MacnamaraA
KadolskyU
BanghamCRM
AsquithB
2009 T-Cell Epitope Prediction: Rescaling Can Mask Biological Variation between MHC Molecules. PLoS Comput Biol 5 e1000327
49. HeckermanD
KadieC
ListgartenJ
2007 Leveraging information across HLA alleles/supertypes improves epitope prediction. J Comput Biol 14 736 746
50. PetersB
BuiH-H
FrankildS
NielsonM
LundegaardC
2006 A community resource benchmarking predictions of peptide binding to MHC-I molecules. PLoS Comput Biol 2 e65
51. SchmidB
KesmirC
de BoerRJ
2008 The specificity and polymorphism of the MHC class I prevents the global adaptation of HIV-1 to the monomorphic proteasome and TAP. PLoS ONE 3 e3525
52. FortierM-H
CaronE
HardyM-P
VoisinG
LemieuxS
2008 The MHC class I peptide repertoire is molded by the transcriptome. J Exp Med 205 595 610
53. BorghansJAM
MølgaardA
de BoerRJ
KesmirC
2007 HLA Alleles Associated with Slow Progression to AIDS Truly Prefer to Present HIV-1 p24. PLoS ONE 2 e920
54. R Development Core Team 2008 R: A Language and Environment for Statistical Computing
55. CrawleyMJ
2007 The R Book: John Wiley and Sons, Ltd
56. KaslowRA
McNichollJ
HillAVS
2008 Genetic susceptibility to infectious diseases New York Oxford University Press xvi, 447
57. YamanoY
CohenCJ
TakenouchiN
YaoK
TomaruU
2004 Increased expression of human T lymphocyte virus type I (HTLV-I) Tax11-19 peptide-human histocompatibility leukocyte antigen A*201 complexes on CD4+ CD25+ T Cells detected by peptide-specific, major histocompatibility complex-restricted antibodies in patients with HTLV-I-associated neurologic disease. J Exp Med 199 1367 1377
58. SeikiM
HattoriS
HirayamaY
YoshidaM
1983 Human adult T-cell leukemia virus: complete nucleotide sequence of the provirus genome integrated in leukemia cell DNA. Proc Natl Acad Sci U S A 80 3618 3622
Štítky
Hygiena a epidemiologie Infekční lékařství LaboratořČlánek vyšel v časopise
PLOS Pathogens
2010 Číslo 9
- Parazitičtí červi v terapii Crohnovy choroby a dalších zánětlivých autoimunitních onemocnění
- Vakcíny proti klíšťové encefalitidě
- Kdy je nejlepší očkovat
- Možné vedlejší účinky očkování
- Imunogenita vakcín
Nejčtenější v tomto čísle
- Structure of the Extracellular Portion of CD46 Provides Insights into Its Interactions with Complement Proteins and Pathogens
- The Length of Vesicular Stomatitis Virus Particles Dictates a Need for Actin Assembly during Clathrin-Dependent Endocytosis
- Inhibition of TIR Domain Signaling by TcpC: MyD88-Dependent and Independent Effects on Virulence
- The Coevolution of Virulence: Tolerance in Perspective
Zvyšte si kvalifikaci online z pohodlí domova
Mazová zátka a její řešení
nový kurzVšechny kurzy