
Asociační studie vztahu čtrnácti polymorfismů kandidátních genů k dispozicím k závislosti na alkoholu
Authors:
O. Šerý; J. Lochman; K. Gláslová 1; R. Štaif; M. Novotný 1; V. Kmoch 1; P. Zvolský 1
Authors‘ workplace:
Laboratoř neurobiologie a molekulární psychiatrie, Laboratoře molekulární fyziologie, Ústav biochemie, Přírodovědecká fakulta MU, Brno
ředitel Ústavu biochemie doc. Ing. M. Mandl, CSc.
Psychiatrická klinika 1. LF UK a VFN, Praha
; přednosta prof. MUDr. J. Raboch, DrSc.
1
Published in:
Čes. a slov. Psychiat., 105, 2009, No. 4, pp. 153-162.
Category:
Original Article
Overview
Dispozice k závislosti na alkoholu jsou ovlivňovány mnoha environmentálními a genetickými faktory. Moderní molekulárně-genetické metody umožňují testování specifických genů, které ovlivňují patofyziologii komplexních chorob, jakou je také závislost na alkoholu. Mezi strategie pro odhalování predisponujících genů se řadí asociační studie. Provedli jsme asociační studii čtrnácti polymorfismů kandidátních genů ve vztahu k závislosti na alkoholu. Asociační studie byla provedena na 847 osobách. Objevili jsme vztah mezi závislostí na alkoholu a geny pro COMT, MAO-B, IL6, IL2, LILRA1 a PSG11. Všechny asociované geny ovlivňují neurovývoj centrální nervové soustavy. Předpokládáme vztah mezi geny, které se účastní neurovývoje CNS, a dispozicemi k závislosti na alkoholu.
Klíčová slova:
závislost na alkoholu, asociační studie, polymorfismus, gen, COMT, IL6, IL2, MAO, DAT1, DRD2.
ÚVOD
Od první práce, která nalezla vztah mezi genovým polymorfismem a závislostí na alkoholu, již uplynulo 18 let [2]. Tehdy autoři článku „Allelic association of human dopamine D2 receptor gene in alcoholism“ předběhli realitu. Postupem doby se ale podařilo nacházet reálné vztahy mezi jednotlivými geny a závislostí na alkoholu, a to při využití tzv. asociačních studií. Zatímco japonští výzkumníci se věnovali především polymorfismům a mutacím receptorů dopaminového systému, evropští a američtí vědci se pustili do výzkumu kandidátních genů např. z oblasti metabolismu neurotransmitérů, které jsou součástí odměňovacího systému. Bohužel, jak se ukázalo, ani jeden z přístupů vyhledávání kandidátních genů nebyl správný. Zatímco nebyla objevena žádná mutace dopaminového receptoru způsobující dispozice k závislosti na alkoholu, podařilo se prokázat vztah mezi závislostí na alkoholu a některými polymorfismy kandidátních genů z řad enzymů zúčastněných v metabolismu neurotransmitérů, jako jsou COMT či MAO. Nicméně žádný z vlivů dosud objevených polymorfismů nemá zásadní význam pro dispozice k závislosti na alkoholu. Je to dáno především skutečností, že dispozice k závislosti na alkoholu mají multifaktoriální charakter, a jak předpokládáme, na dispozicích k závislosti na alkoholu se podílí celá řada faktorů a celá řada genů.
V současnosti lze sledovat postupné zavádění nejnovějších technologií pro analýzu DNA také do psychiatrického výzkumu. Jedná se především o tzv. DNA čipy. Ty umožňují sledovat v rámci jedné analýzy trvající 2 dny až jeden milion polymorfismů u 96 osob a porovnávat pak vliv interakce velkého množství polymorfismů na fenotyp. DNA čipová technologie je tedy budoucností také pro psychiatrický výzkum, kdy se předpokládá jednak objev nových polymorfismů, které zásadním způsobem ovlivňují dispozice k závislosti na alkoholu, jednak se předpokládá nalezení zatím nepopsaných haplotypů a genových interakcí, které ovlivňují dispozice k závislosti na alkoholu.
Cílem naší práce bylo nalezení DNA polymorfismů, které se podílejí na dispozicích k závislosti na alkoholu. Polymorfismy byly vybírány také z těch genů, které se podílejí na vývoji, diferenciaci a modulaci dopaminového či opioidního systému v CNS. Do naší studie jsme zahrnuli dva polymorfismy s dosud neznámou funkcí, které byly objeveny pomocí celogenomové asociační studie závislosti na alkoholu [9].
METODIKA
Pacienti i kontrolní skupiny byly vybírány z věkové kategorie 25 - 50 let. Soubor pacientů závislých na alkoholu se skládal ze 444 osob. Pacienti závislí na alkoholu byli vybíráni na protialkoholních odděleních VFN v Praze, Apolinářská 4, a ženském PA oddělení v Lojovicích, dále na PA odděleních v Psychiatrické léčebně v Praze - Bohnicích, v Psychiatrické léčebně v Jihlavě a v Psychiatrickém protialkoholním oddělení v Červeném Dvoře. Pacienti před zahrnutím do studie podepsali informovaný souhlas.
Každá osoba ze souboru závislých na alkoholu prodělala pohovor s psychiatrem, v jehož rámci bylo provedeno zhodnocení osobní a rodinné anamnézy, a byla ověřena diagnóza podle MKN - 10 (F10.2), DSM IV (303.90) a MAST (Mitchigan Alcoholism Sreening Test).
Dále byla do studie zahrnuta skupina 403 kontrolních osob, které byly charakterizovány jako osoby bez závislosti či abúzu alkoholu a jiných drog (kromě kávy). Tento soubor osob byl získán na různých somatických odděleních VFN v Praze, na transfuzních stanicích v Praze a Brně, na vysokých školách a nejrůznějších pracovištích a ve spolupráci s vojenskými úřady v rámci odvodů vojáků základní služby v Praze.
Po podepsání informovaného souhlasu a vyplnění strukturovaného dotazníku určeného pro kontrolní skupinu (stručná osobní anamnéza s vyšetřením MAST), byly tyto informace ověřeny členem výzkumného týmu osobním pohovorem.
Osoby zařazené do kontrolní skupiny splňovaly další podmínky pro zařazení do studie: nekuřáctví (v současnosti i minulosti), nezneužívání psychoaktivních látek (v současnosti i minulosti) a vyloučení současného i dřívějšího zneužívání alkoholu (abúzu). Bylo také provedeno psychologické a psychiatrické vyšetření kontrolních osob (Lüscherova klinická diagnostika, M.I.N.I. – Mini mezinárodní neuropsychiatrické interview). A dále pomocí dotazníků CAGE a CSÉMY byl zjištěn vztah k alkoholu v posledních 12 měsících.
Po podepsání informovaného souhlasu, diagnóze a vyplnění dotazníků bylo pacientům i kontrolním osobám odebráno kvalifikovanou zdravotní sestrou nebo lékařem 5 ml žilní krve. Vzorek krve byl zmrazen na minus 20 stupňů Celsia a v termoboxu dopraven do laboratoře. Jednotlivé vzorky genomické DNA byly izolovány z krve s využitím komerčně dostupného izolačního kitu UltraClean BloodSpin Kit (MoBio, USA).
V rámci projektu byly studovány tyto polymorfismy kandidátních genů:
- TaqI A genu pro DRD2 (dopaminový receptor D2) [7]
- Val158Met genu pro COMT (katechol-O-metyltransferázu) [10]
- I/D genu pro ACE (angiotensinkonvertázu) [17]
- –174 genu pro IL-6 (interleukin 6) [24]
- delta 32. genu pro CCR5 (chemokinový receptor 5) [3]
- – 308 TNF alfa (tumor nekrotizující faktor alfa) – metodika viz. níže
- M235T genu pro AGT (angiotensinogen) [18]
- A/G ve 13. intronu genu pro MAO-B (monoaminoxidázu B) [6]
- –330 genu pro IL-2 (interleukin 2) – metodika viz. níže
- A118G (Asn40Asp) genu pro Mu opiodní receptor [21]
- 196 G/A (val66met) genu pro BDNF (Brain derived neurotrophic factor) [22]
- 40-bp VNTR genu pro DAT1 (dopaminový transportér) [19]
- polymorfismus tsc1189131 – metodika viz níže
- polymorfismus tsc0598556 – metodika viz níže
Detekce polymorfismu – 308 genu pro TNF alfa byla provedena podle metodiky navržené v naší laboratoři. Pro PCR reakci polymorfismu genu pro TNF alfa byly použity primery: 5´ - AGG CAA TAG GTT TTG AGG GGC AT – 3´ a 5 - CGG GGA AAG AAT CAT TCA ACC AG –3´. Amplifikační reakce byla prováděna v termálním cykléru Techne Touchgene Gradient. Po počáteční denaturaci, trvající 2,5 minuty při 94 °C, byla DNA amplifikována v tříkrokových cyklech: denaturace 30 s při 94°C, annealing 30 s při 66 °C, extenze 30 s při 72 °C. Po 35 cyklech byla použita finální extenze 5 minut při 72 °C. Následovala restrikční analýza: 20 μl PCR amplifikátu bylo smícháno s 2 U restrikčního enzymu NcoI (Fermentas, Litva). Poté následovala inkubace na vodní lázni při 37 °C po dobu 16 hodin. Gelová elektroforéza restrikčních fragmentů byla provedena na 2% agarózovém gelu (EliPhore) barveném ethidium bromidem při 80 V po dobu dvou hodin. Byly detekovány následující fragmenty: 264 bp, který značil genotyp „11“, 264 bp a 284 bp charakterizovaly genotyp „12“, 284 bp identifikoval genotyp „22“.
Detekce polymorfismu –330 genu pro IL-2 byla provedena podle metodiky navržené v naší laboratoři. Pro PCR reakci polymorfismu genu pro IL-2 byly použity primery: I2-2 5´ - caa gac tta gtg caa tgc aag – 3´a I2-5 5´ - tca cat gtt cag tgt agt ttc a – 3´. Po počáteční denaturaci, trvající 2,5 minuty při 94 °C, byla DNA amplifikována v tříkrokových cyklech: denaturace 30 s při 94 °C, annealing 30 s při 56,5 °C, extenze 30 s při 72 °C. Po 35 cyklech byla použita finální extenze 5 minut při 72 °C. Celková délka amplifikovaného fragmentu byla 450 bp. Následovala restrikční analýza: 20 μl PCR amplifikátu bylo smícháno s 5 U restrikčního enzymu Hsp92 II (Promega) a dále s 1,7 μl pufru K (Promega). Poté následovala inkubace na vodní lázni při 37 °C po dobu 7 hodin. Detekce restrikčních fragmentů byla provedena metodou iontoměničové HPLC (kolona TSK-gel DEAE NPR). K vymytí DNA z kolony byl použit gradient chloridu sodného. Byly detekovány následující fragmenty: 107 bp, který značil genotyp GG, 107 bp a 90 bp charakterizovaly genotyp TG, 90 bp identifikoval genotyp TT.
Detekce polymorfismu tsc1189131 (sekvence AC004784, gen v blízkosti pregnancy beta-1-glycoprotein genu) byla provedena podle metodiky navržené v naší laboratoři. K detekci byl využit automatizovaný systém AB7300 (Applied Biosystems). Pro RT-PCR reakci polymorfismu tsc1189131 byly použity primery: AFY19-1 : 5´ - act tga acc agt gac ctc taa - 3´ a AFY19-2 : 5´ - cta tcc cct tca ctc ttc ct - 3´. K detekci alel byly využity tyto sondy: TaqProbeC – agc aga gcg caa gga atg atc (Yakima Yellow) a TaqProbeT – agc aga gtg caa gga atg atc (FAM). Po počáteční denaturaci 10 minut při 95 °C byla DNA amplifikována v tříkrokových cyklech 40x: denaturace 20 s při 95 °C, annealing 20 s při 56 °C, extenze 32 s při 72 °C. Odlišné genotypy byly detekovány analýzou míry amplifikace, která byla mezi oběma alelami velmi odlišná.
Detekce polymorfismu tsc0598556 (sekvence AC009892) byla provedena podle metodiky navržené v naší laboratoři. K detekci byl využit automatizovaný systém AB7300 (Applied Biosystems). Pro RT-PCR reakci polymorfismu TSC0598556 byly použity primery: AFYNS-1 : 5´ - gca cag ggc tca gtg act tct - 3´ a AFYNS-2 : 5´ - agg gcg ttt tcc tgc act ct - 3´. K detekci alel byly využity tyto sondy: TaqProbeC – ttc cct tgc aga tcc tga gct (Yakima Yellow) a TaqProbeT – ttc ctt tgc aga tcc tga gct (FAM). Po počáteční denaturaci 10 minutách při 95 °C byla DNA amplifikována v tříkrokových cyklech 40x: denaturace 20 s při 95 °C, annealing 20 s při 59 °C, extenze 32 s při 72°C. Odlišné genotypy byly detekovány pomocí míry amplifikace pro jednotlivé sondy.
Statistické vyhodnocení výsledků bylo provedeno pomocí softwaru CSS Statistica (Verze 7). Byly vytvořeny kontingenční tabulky s počty osob pro jednotlivé genotypy studovaných genů. Z těchto tabulek se dále vycházelo při výpočtu genotypových a alelických frekvencí sledovaných polymorfismů. K přehlednému znázornění vztahů mezi sledovanými proměnnými byl použit Spearmanův korelační koeficient.
Výpočet rozdílu mezi genotypovými frekvencemi sledovaných polymorfismů byl proveden pomocí X2 testu, výpočet rozdílu mezi alelickými frekvencemi polymorfismů byl proveden pomocí Fisher-exact testu. Za signifikantní výsledek sledovaných genotypových a alelických frekvencí obou polymorfismů byla pokládána hladina pravděpodobnosti p <0,05. Z křížových tabulek pro varianty dvou polymorfismů se vycházelo při výpočtu vzájemné vazby polymorfismů studovaných genů. Tento výpočet byl proveden speciálním programem vyvinutým na LF MU v Brně doc. RNDr. Vladimírem Znojilem, CSc. Logistickou regresí byly spočítány na základě několika parametrů (věk, pohlaví a sledované polymorfismy genů) genetické zátěže konkrétních osob, které nemohou být uvedeny z etických důvodů.
VÝSLEDKY
Výsledky genotypových a alelických frekvencí společně s výsledky statistického srovnání jsou uvedeny v tabulkách 1–15. V tabulkách jsou uvedeny počty osob s daným genotypem, dále genotypové frekvence, alelické frekvence a výsledky vyhodnocení statistickými testy (Fisher exact test pro výpočet alelických frekvencí a X2 test pro výpočet genotypových frekvencí). Pro zkoumaný repetitivní polymorfismus 40-bp VNTR genu pro DAT1 jsou uvedeny dvě tabulky, v níž první uvádí počty nalezených opakování (repeticí) v souborech zkoumaných osob a druhá tabulka uvádí rozdělení nalezených repeticí do tří intervalů („dlouhé, středně dlouhé a krátké repetice“) za účelem dalšího statistického zpracování.

Při srovnání alelických frekvencí pomocí Fisher exact testu byl objeven statisticky signifikantní rozdíl ve frekvenci alely M polymorfismu genu pro COMT (p=0,012) ve skupině mužů, kde závislí muži měli frekvenci alely M 0,48 a muži z kontrolního souboru měli frekvenci alely M 0,57 (tab. 2).
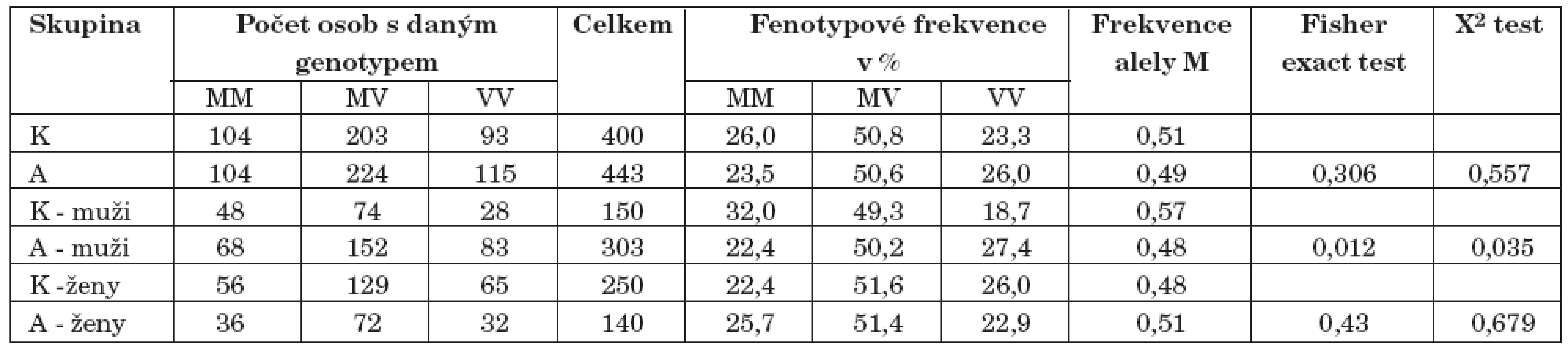
Byl nalezen rozdíl mezi genotypovými frekvencemi polymorfismu 174. genu pro IL-6 mezi celkovou skupinou alkoholiků a kontrolami (p=0,035). Naopak nebyl nalezen rozdíl v genotypové frekvenci při rozdělení souboru podle pohlaví. Nebyl také prokázán signifikantní rozdíl mezi alelickými frekvencemi celkového souboru (p=0,09), souboru mužů (p=0,082) a souboru žen (p=0,784) (tab. 3, tab. 4).

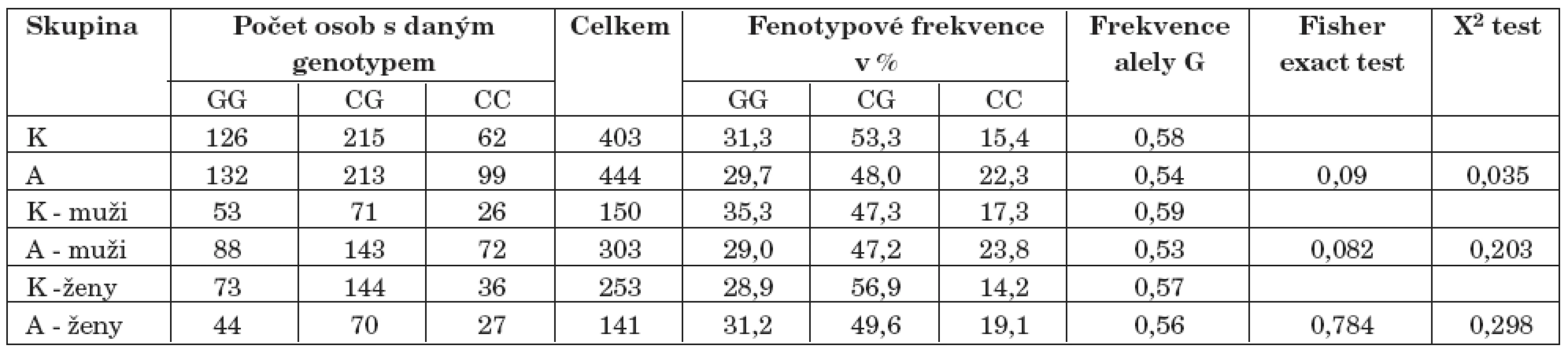
Ve studii byl nalezen signifikantní rozdíl v alelické frekvenci alely A polymorfismu genu pro MAO-B u mužů (p=0,023). A/G polymorfismus genu pro MAO-B byl genotypizován pouze v mužské části souboru, neboť tento polymorfismus se nalézá na pohlavně vázaném chromozomu (tab. 5 – tab. 8).




Statisticky signifikantní rozdíl byl objeven ve frekvenci alely T polymorfismu –330 genu pro IL-2, kde alela T byla nalezena u kontrolních osob ve frekvenci 0,74, u závislých osob ve frekvenci 0,65 (p=0,016). Při srovnání genotypových frekvencí pomocí X2testu byl rovněž objeven statisticky významný rozdíl v genotypových frekvencích polymorfismu –330 genu (p=0,024). Pohlavní rozdíl nebyl za pomocí Fisher exact testu ani X2 testu potvrzen (p >0,05) (tab. 9).

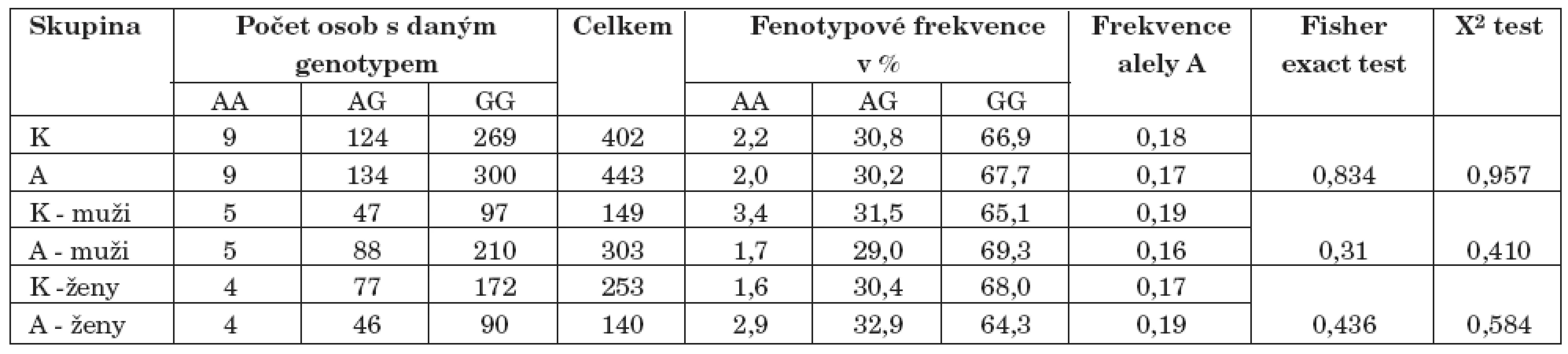
Dále byl ve studii potvrzen vztah mezi polymorfizmem tsc1189131 a závislostí na alkoholu (Fisher exact test p=0,026, X2 test p=0,051) v celkové populaci. Při rozdělení souborů osob podle pohlaví byl rozdíl v alelické frekvenci ještě výraznější u žen (p=0,018). Alela C se vyskytuje v souboru kontrolních osob více často než alela G, a to jak v celkové populaci, tak i v populaci rozdělené na muže a ženy (tab. 10 – tab. 14).




Při srovnání alelických frekvencí pomocí Fisher exact testu a srovnání genotypových frekvencí pomocí X2testu nebyl u ostatních polymorfismů zkoumaných kandidátních genů objeven vztah mezi soubory osob závislých na alkoholu a kontrol, a to ani při rozdělení studovaných souborů na muže a ženy (p>0,05).
Dále byly studovány vztahy dvojgenotypových frekvencí ve vztahu k závislosti na alkoholu, byl tedy studován vztah kombinace dvou genotypů. Při srovnání dvojgenotypových frekvencí tsc1189131 a genu pro IL6 byl nalezen statisticky signifikantní rozdíl mezi osobami závislými na alkoholu a kontrolami (p<0,01, p=0,006). Genotypy tsc1189131/IL6 TTCC a CTCC disponují ke vzniku závislosti na alkoholu a vyskytují se signifikantně častěji u osob závislých na alkoholu.

Na celkovém souboru osob byla objevena vazba mezi polymorfismy genů pro TNF alfa a angiotensinogen, COMT a DAT1, COMT a MAO B, COMT a BDNF a BDNF a Mu receptor (tab. 16). V souboru osob závislých na alkoholu byla objevena vazba mezi geny pro IL-2 a angiotensinogen, COMT a BDNF, COMT a MAO B, BDNF a MAO B (tab. 17). Na celkovém souboru kontrol byla objevena vazba mezi geny pro IL-6 a polymorfizmem tsc1189131, BDNF a Mu receptor, Mu receptor a polymorfizmem tsc0598556 (tab. 18).



DISKUSE
V rámci asociační studie dispozic k závislosti na alkoholu jsme zkoumali celkem 14 polymorfismů na souboru 847 osob. V naší studii byl potvrzen dříve objevený vztah mezi Val158Met polymorfismem genu pro COMT a dispozicemi k alkoholismu pouze v mužské části souboru (pro alelické frekvence p <0,02, pro genotypové frekvence p <0,04). Domníváme se, že tato souvislost je zprostředkována úlohou COMT ve vývoji mozku. Jak známo, COMT katalyzuje metylaci katecholových estrogenů na methoxy estrogeny – např. 2-methoxyestradiol, což je antiproliferativní metabolit. Úloha COMT v neuronálním vývoji by měla být dále studována. Podrobněji je tato souvislost popsána v naší předešlé publikaci [23].
Poprvé byl objeven vztah mezi –174 polymorfismem genu pro IL-6 a dispozicemi k závislosti na alkoholu na celkovém souboru osob (pro alelické frekvence p=0,09, pro genotypové frekvence p <0,04). Domníváme se, že vliv tohoto polymorfismu na dispozice k závislosti na alkoholu je vázán na účinek IL6 na přežívání neuronů a růst neuritů v průběhu vývoje CNS. Vztah mezi IL6 a dispozicemi k závislosti na alkoholu je blíže diskutován v naší předešlé publikaci [20].
Nově byl objeven vztah mezi závislostí na alkoholu a studovaným polymorfismem genu pro MAO B (pro alelické frekvence p <0,03), a to pouze u mužské části souboru. Domníváme se, že zásadním vlivem MAO B na dispozice k závislosti na alkoholu bude ovlivnění hladin fenylethylaminu (PEA). V čínské populaci vztah mezi polymorfismem MAO B a závislostí na alkoholu nebyl objeven [13].
Poprvé byl objeven vztah mezi genem pro IL-2 a závislostí na alkoholu na celkovém souboru osob (pro alelické frekvence p <0,02, pro genotypové frekvence p <0,03). Marcos a spol. [14] zjistili vztah mezi tímto polymorfismem a jaterní cirhózou u osob závislých na alkoholu. Ve studiích bylo zjištěno, že u osob s genotypem GG mají T lymfocyty vyšší produkci IL2 oproti heterozygotům TG a homozygotům TT [8]. IL2 podporuje přežívání kultivovaných neuronů a prodlužování jejich neuritů a ovlivňuje hypothalamo-hypofysární systém [16].
Chiu a spol. [9] použili DNA čipovou technologii na platformě Affymetrix a studovali vztah mezi 11 120 polymorfismy a závislostí na alkoholu na souboru 1614 osob. Objevili celkem 20 polymorfismů s významnějším vztahem k závislosti na alkoholu. Dva z těchto polymorfismů tsc1189131 (rs1963786) a tsc0598556 (rs272411) jsme vybrali také do naší studie a pokusili jsme se o zopakování asociace. V naší studii byl potvrzen vztah mezi polymorfismem tsc1189131 a závislostí na alkoholu na celkovém souboru (pro alelické frekvence p <0,03, pro genotypové frekvence p=0,068) a na souboru žen (pro alelické frekvence p <0,02 a pro genotypové frekvence p=0,051). Tento polymorfismus se nachází jako intronový polymorfismus v genu pro pregnancy specific beta-1-glycoprotein 11 (PSG11). Jak bylo zjištěno, PSG11 ovlivňuje mimo jiné sekreci IL6 a je produkován především placentárním syncytiotrofoblastem během těhotenství. Nelze tedy než se odkázat na námi objevený vztah mezi IL-6 a závislostí na alkoholu a znovu poukázat na možné ovlivnění dispozic k závislosti na alkoholu v době vývoje CNS. Navíc při studiu interakcí mezi polymorfismy byl zjištěn vztah mezi polymorfismem genu pro IL6 a polymorfismem tsc1189131, a to na souboru kontrolních osob. Znamená to, že se u souboru kontrolních osob častěji vyskytují některé dvojgenotypy než by bylo běžné v normální populaci. Jak jsme již při dřívějším výzkumu předznamenali, výběr nenávykových kontrolních osob se zdá být výraznějším kritériem než samotná závislost na alkoholu. Soubor kontrolních osob bez návyku na tabáku, drogách a alkoholu tvoří ve srovnání se souborem alkoholiků pomyslnou druhou část rozložení návykovosti v populaci. Právě interakce mezi IL6 a tsc1189131 v nenávykové části populace může být v dalším výzkumu vodítkem pro nalezení kombinace polymorfismů chránicích před závislostí na alkoholu.
Byl potvrzen také vztah mezi polymorfismem tsc0598556 a závislostí na alkoholu na ženské části souboru (pro genotypové frekvence p <0,03). Tento polymorfismus se nachází v intronu genu pro leukocyte immunoglobulin-like receptor (LILRA1). LILTRA1 se nachází jako imunoreceptor na monocytech a B buňkách a v menší míře na dendritických a NK buňkách. Lze předpokládat, že i zde bude polymorfismus ovlivňovat dispozice k závislosti na alkoholu ovlivněním vývoje CNS.
V naší studii jsme nezjistili vztah mezi závislostí na alkoholu a geny, které jsou součástí renin-angiotensinového systému (ACE a AGT). Samostatnou kapitolou je hypotetický vztah mezi Taql A polymorfismem genu pro DRD2 a závislostí na alkoholu, který se v naší studii neprokázal. Blíže je tento vztah komentován v naší předešlé práci [11].
Dopaminový přenašeč DAT hraje klíčovou roli v homeostatické regulaci dopaminergních neurotransmisích. Köhnke a spol. [12] ve své asociační studii porovnali soubor 216 osob závislých na alkoholu se 102 kontrolami. Ve studii objevili vyšší frekvenci alely 9 u osob závislých na alkoholu (0,48) než u kontrol (0,32). Ve studii, kterou provedli Wernicke a spol. [26], byl naopak zaznamenán vztah genotypu 10/10 k závislosti na alkoholu. Studie, kterou publikovali Foley a spol. [4], vztah VNTR polymorfismu genu pro DAT1 k závislosti na alkoholu nepotvrdila. Rovněž studie dalších autorů [5, 15] a naše studie nepotvrzují asociaci mezi VNTR polymorfismem genu pro DAT1 a alkoholovou závislostí.
Neurotrofiny jsou specifické růstové faktory, které se podílejí na lokální regulaci synaptické plasticity. Nejčastěji zkoumaným neurotrofinem je v posledních letech BDNF. BDNF byl zkoumán v souvislosti s patogenezí mnoha psychických onemocnění jako jsou mentální poruchy, deprese, úzkost, schizofrenie a drogová závislost. Val66Met polymorfismus genu pro BDNF byl rovněž zkoumán i v souvislosti se závislostí na alkoholu. Ve studii, kterou provedli Tsai a spol. [25], byl s negativním výsledkem zkoumán vztah tohoto polymorfismu k násilnému chování a agresivitě u osob závislých na alkoholu. V naší studii jsme neobjevili vztah Val66Met polymorfismu genu pro BDNF k alkoholové závislosti (p>0,05).
Předpokládá se vztah opiodních receptorů ke vzniku alkoholové závislosti. Příjem alkoholu může být výsledkem kompenzace dědičného deficitu endogenního opioidního systému. Alkohol interferuje s opioidním systémem a ovlivňuje mezolimbický systém. Alkohol má vliv na vazebné schopnosti opioidních receptorů a moduluje syntézu a sekreci opioidních peptidů. Alkohol stimuluje uvolnění dopaminu v mozku, které je regulováno OP1 a OP2 opioidními receptory. Jsou-li tyto receptory zablokovány, člověk necítí požitek z alkoholu.
Mezi nejčastěji zkoumané polymorfismy genu pro Mu opioidní receptor patří A118G (Asn40Asp) polymorfismus genu pro Mu opioidní receptor. V roce 2006 byla provedena meta-analýza vztahu tohoto polymorfismu k závislostem [1]. Meta-analýza zahrnovala celkem 8000 osob, výsledky 22 odborných článků a 28 typů sledovaných souborů osob. Vztah k závislostem nebyl pro A118G (Asn40Asp) polymorfismus genu pro Mu opioidní receptor v meta-analýze prokázán. Naše studie potvrzuje výsledek meta-analýzy.
ZÁVĚR
V naší devítileté studii jsme využili v dané době známé poznatky o molekulárních příčinách vzniku závislosti na alkoholu a do studie jsme kromě klasických kandidátních genů zahrnuli i ověření vlivu dvou nových polymorfismů objevených pomocí celogenomové asociační studie. Dále jsme přispěli k objevování dispozic k závislosti na alkoholu i propagací teorie, že dispozice k závislosti na alkoholu jsou do určité míry neurovývojová porucha a testovali jsme i vliv kandidátních genů z řad modulátorů vývoje centrálního nervového systému jakými jsou IL-6, IL-2 a BDNF. Jak se ukázalo, naše předpoklady byly správné a dokonce i vliv COMT na dispozice k závislosti na alkoholu lze vyvozovat spíše z jeho úlohy ve vývoji CNS než ve vztahu k metabolismu neurotransmitérů. Výsledky celé naší studie naznačují zajímavé interakce mezi studovanými polymorfismy a potvrzují hypotézu multigenního základu dispozic k alkoholové závislosti a složité funkční provázanosti kandidátních genů.
Pro praxi z našich výsledků vyplývá, že dispozice k závislosti na alkoholu se vytvářejí již v období vývoje CNS a nutně nesouvisejí s přímým účinkem alkoholu na CNS. Zatímco vliv genů na dispozice k závislosti nelze zatím přímo ovlivnit, lze úpravou prostředí zamezit propuknutí závislosti na alkoholu u osob s genetickou dispozicí. K takové úpravě prostředí u disponovaných osob patří především zajištění citového zázemí v období dospívání a celoživotní abstinence od alkoholu a jiných drog.
Poděkování
Tento projekt byl podpořen Interní grantovou agenturou Ministerstva zdravotnictví ČR–IGA MZ ČR č. NR9298-3/2007 a dále projektem Ministerstva školství, mládeže a tělovýchovy ČR č. MSM0021622413.
Doc. RNDr. Omar Šerý, Ph.D.
Laboratoř neurobiologie a molekulární psychiatrie
Laboratoře molekulární fyziologie
Ústav biochemie
Přírodovědecká fakulta MU
Kotlářská 2
611 37 Brno
e-mail: omarsery@sci.muni.cz
Sources
1. Arias, A., Feinn, R., Kranzler, H. R.: Association of an Asn40Asp (A118G) polymorphism in the mu-opioid receptor gene with substance dependence: a meta-analysis. Drug Alcohol Depend., 83, 2006, pp. 262-268.
2. Blum, K., Noble, E. P., Sheridan, P. J., Montgomery, A., Ritchie, T., Jagadeeswaran, P., Nogami, H., Briggs, A. H., Cohn, J. B.: Allelic association of human dopamine D2 receptor gene in alcoholism. JAMA, 263, 1990, pp. 2055-2060.
3. Eugen-Olsen, J., Iversen, A. K., Garred, P., Koppelhus, U., Pedersen, C., Benfield, T. L., Sorensen, A. M., Katzenstein, T., Dickmeiss, E., Gerstoft, J., Skinhøj, P., Svejgaard, A., Nielsen, J. O., Hofmann, B.: Heterozygosity for a deletion in the CKR-5 gene leads to prolonged AIDS-free survival and slower CD4 T-cell decline in a cohort of HIV-seropositive individuals. AIDS, 11, 1997, pp. 305-310.
4. Foley, P. F., Loh, E. W., Innes, D. J., Williams, S. M., Tannenberg, A. E., Harper, C. G., Dodd, P. R.: Association studies of neurotransmitter gene polymorphisms in alcoholic Caucasians. Ann. N Y Acad. Sci., 1025, 2004, pp. 39-46.
5. Franke, P., Schwab, S. G., Knapp, M., Gänsicke, M., Delmo, C., Zill, P., Trixler, M., Lichtermann, D., Hallmayer, J., Wildenauer, D. B., Maier, W.: DAT1 gene polymorphism in alcoholism: a family-based association study. Biol. Psychiatry, 45, 1999, pp. 652-654.
6. Garpenstrand, H., Ekblom, J., Forslund, K., Rylander, G., Oreland, L.: Platelet monoamine oxidase activity is related to MAOB intron 13 genotype. J. Neural. Transm., 107, 2000, pp. 523-30.
7. Grandy, D. K., Zhang, Y., Civelli, O.: PCR detection of the TaqA RFLP at the DRD2 locus. Hum. Mol. Genet., 2, 1993, p. 2197.
8. Hoffmann, S. C., Stanley, E. M., Darrin Cox, E., Craighead, N., DiMercurio, B. S., Koziol, D. E., Harlan, D. M., Kirk, A. D., Blair, P. J.: Association of cytokine polymorphic inheritance and in vitro cytokine production in anti-CD3/CD28-stimulated peripheral blood lymphocytes. Transplantation, 72, 2001, pp. 1444-1450.
9. Chiu, Y. F., Liu, S. Y., Tsai, Y. Y.: A comparison in association and linkage genome-wide scans for alcoholism susceptibility genes using single-nucleotide polymorphisms. BMC Genetics, 6, 2005, p. S89.
10. Jönsson, E. G., Goldman, D., Spurlock, G., Gustavsson, J. P., Nielsen, D. A., Linnoila, M., Owen, M. J., Sedvall, G. C.: Tryptophan hydroxylase and catechol-O-methyltransferase gene polymorphisms: relationships to monoamine metabolite concentrations in CSF of healthy volunteers. Eur. Arch. Psychiatry Clin. Neurosci., 247, 1997, pp. 297-302.
11. Kašparovská, J., Hříbková, H., Hloušková, R., Novotný, M., Gláslová, K., Šimek, V., Mikeš, V., Zvolský, P., Šerý, O.: TaqI A polymorfismus genu pro dopaminový receptor D2 a alkoholismus v České republice. Čes. a slov. Psychiat., 99, 2003, pp. 5-9.
12. Köhnke, M. D., Batra, A., Kolb, W., Köhnke, A. M., Lutz, U., Schick, S., Gaertner, I.: Association of the dopamine transporter gene with alcoholism. Alcohol Alcohol., 40, 2005, pp. 339-342.
13. Lu, R. B., Lee, J. F., Ko, H. C., Lin, W. W., Chen, K., Shih, J. C.: No association of the MAOA gene with alcoholism among Han Chinese males in Taiwan. Prog. Neuropsychopharmacol. Biol. Psychiatry, 26, 2002, pp. 457-461.
14. Marcos, M., Pastor, I., González-Sarmiento, R., Laso, F. J.: A new genetic variant involved in genetic susceptibility to alcoholic liver cirrhosis: -330T>G polymorphism of the interleukin-2 gene. Eur. J. Gastroenterol. Hepatol., 20, 2008, pp. 855-859.
15. Parsian, A., Zhang, Z. H.: Human dopamine transporter gene polymorphism (VNTR) and alcoholism. Am. J. Med. Genet., 74, 1997, pp. 480-482.
16. Raber, J., Sorg, O., Horn, T. F., Yu, N., Koob, G. F., Campbell, I. L., Bloom, F. E.: Inflammatory cytokines: putative regulators of neuronal and neuro-endocrine function. Brain Res. Brain Res. Rev., 26, 1998, pp. 320-326.
17. Rigat, B., Hubert, C., Corvol, P., Soubrier, F.: PCR detection of the insertion/deletion polymorphism of the human angiotensin converting enzyme gene (DCP1) (dipeptidyl carboxypeptidase 1). Nucleic Acids Res., 20, 1992, p. 1433.
18. Russ, A. P., Maerz, W., Ruzicka, V., Stein, U., Gross, W.: Rapid detection of the hypertension-associated Met235—>Thr allele of the human angiotensinogen gene. Hum. Mol. Genet., 2, 1993, pp. 609-610.
19. Shinohara, M., Mizushima, H., Hirano, M., Shioe, K., Nakazawa, M., Hiejima, Y., Ono, Y., Kanba, S.: Eating disorders with binge-eating behaviour are associated with the s allele of the 3’-UTR VNTR polymorphism of the dopamine transporter gene. J. Psychiatry Neurosci., 29, 2004, pp. 134-137.
20. Šerý, O., Hladilová, R., Novotný, M., Znojil, V., Zvolský, P.: Polymorfismus genu pro IL-6 má vztah k dispozicím k závislosti na alkoholu. Čes. a slov. Psychiat., 99, 2003, s. 185-188.
21. Šerý, O., Lochman, J., Častulík, L., Šťastný, F., Přikryl, R., Češková, E.: Polymorfismus genu pro OPMR1 má vztah ke schizofrenii: nový kandidátní gen. Čes. a slov. Psychiat., 105, 2009 (v tisku).
22. Šerý, O., Lochman, J., Zvolský, P., Hlinomazová, Z., Novotný, M.: Porucha barvocitu u alkoholiků a její souvislost s polymorfizmem genu pro BDNF. Čes. a slov. Psychiat., 104, 2008, s. 115-119.
23. Šerý, O., Mikes, V., Pitelová, R., Znojil, V., Zvolský, P.: The association between high-activity COMT allele and alcoholism. Neuro Endocrinol Lett., 27, 2006, pp. 231-235.
24. Šerý, O., Theirner, P., Hladilová, R., Štaif, R., Balaštiková, B., Drtílková, I.: Geny pro Il-6 a DRD2 souvisejí s hyperkinetickou poruchou. Čes. a slov. Psychiat., 99, 2003, s. 404-409.
25. Tsai, S. J., Liao, D. L., Yu, Y. W., Chen, T. J., Wu, H. C., Lin, C. H., Cheng, C. Y., Hong, C. J.: A study of the association of (Val66Met) polymorphism in the brain-derived neurotrophic factor gene with alcohol dependence and extreme violence in Chinese males. Neurosci. Lett., 381, 2005, pp. 340-343.
26. Wernicke, C., Smolka, M., Gallinat, J., Winterer, G., Schmidt, L. G., Rommelspacher, H.: Evidence for the importance of the human dopamine transporter gene for withdrawal symptomatology of alcoholics in a German population. Neurosci. Lett., 333, 2002, pp. 45-48.
Labels
Addictology Paediatric psychiatry PsychiatryArticle was published in
Czech and Slovak Psychiatry

2009 Issue 4
Most read in this issue
- Parazitární blud v psychiatrické ambulanci
- Metody diagnostiky disociativních symptomů
- Depresívne ochorenia a sclerosis multiplex
- Asociační studie vztahu čtrnácti polymorfismů kandidátních genů k dispozicím k závislosti na alkoholu